

Abstract
The zebrafish (Danio rerio) has recently become a common model in the fields of genetics, environmental science, toxicology, and especially drug screening. Zebrafish has emerged as a biomedically relevant model for in vivo high content drug screening and the simultaneous determination of multiple efficacy parameters, including behaviour, selectivity, and toxicity in the content of the whole organism. A zebrafish behavioural assay has been demonstrated as a novel, rapid, and high-throughput approach to the discovery of neuroactive, psychoactive, and memory-modulating compounds. Recent studies found a functional similarity of drug metabolism systems in zebrafish and mammals, providing a clue with why some compounds are active in zebrafish in vivo but not in vitro, as well as providing grounds for the rationales supporting the use of a zebrafish screen to identify prodrugs. Here, we discuss the advantages of the zebrafish model for evaluating drug metabolism and the mode of pharmacological action with the emerging omics approaches. Why this model is suitable for identifying lead compounds from natural products for therapy of disorders with multifactorial etiopathogenesis and imbalance of angiogenesis, such as Parkinson's disease, epilepsy, cardiotoxicity, cerebral hemorrhage, dyslipidemia, and hyperlipidemia, is addressed.
1. Introduction
The zebrafish (Danio rerio) is a tropical freshwater fish that has become one of the most popular vertebrate model organisms in biological research. The zebrafish has traditionally been used as a model for studying developmental biology and embryology. Recently, zebrafish has become famous in the fields of genetics, environmental science, toxicological studies, and especially drug screening [1, 2].
Zebrafish are small, even adults are only 3-4 cm long, and are suitable, therefore, for animal studies in laboratories with limited space. Their high fecundity enables each adult female to produce hundreds of eggs per mating at intervals of only a few days. The embryos grow and develop rapidly. By 120 h after fertilization (hpf), the heart, liver, brain, pancreas, kidney, and other organs are completely developed. Zebrafish cardiovascular, nervous systems and metabolic pathways are highly similar to those of mammals at the anatomical, physiological, and molecular levels. The zebrafish genome is highly similar to the human genome, with approximately 87% similarity [2]. Their pharmacological response is comparable with that of human, suggesting applications in identifying test compounds with therapeutic potential. The larvae are only 1–4 mm long and can survive in a single well of a standard 384-well plate for several days by using the nutrients stored in the yolk sac. Assay studies require only a small quantity (10–100 ng) of test compounds, such as small molecules, which are easily absorbed through the skin and gills, or directly by swallowing after 72 hpf. Early zebrafish embryos and larvae are optically transparent, which allows real-time imaging in vivo. These advantageous features combine and make zebrafish an ideal model for studying the biological activity profiling of natural products containing complex chemical components.
2. Relevance and Predictability of Drug Response between Zebrafish and Human
Using zebrafish as a model for drug screening will always raise the question of whether the beneficial effect of a drug lead compound observed in zebrafish would have clinical relevance. Although it has been shown that the zebrafish and human genomes are highly similar, a study should be done to compare the physiological response of human and zebrafish after exposure to a series of drugs. Mittelstadt has tested the effect of nine drugs with QT prolongation in zebrafish and found eight of these compounds (except procainamide) induced dissociation between the atrium and ventricular rates (Table 1) [3]. A similar study was done by Berghmans, who measured the atrial and ventricular rates of zebrafish in response to seven known QT-induced drugs and 2 negative controls and found that 7 of the 9 compounds, including the negative controls, showed the expected effects (Table 1) [1]. Two studies focused on the optomotor response were followed independently by Berghmans and Richards They both found zebrafish showed a high percentage of predictability (~78% and ~70%) of drug response (Table 1) [1, 4]. The zebrafish is also a good model for screening drugs with potential seizure liability. Winter reported the animal model offered 72% overall predictability as 13 out of 17 positive controls and 5 out of 8 negative controls showed their predicted effects (Table 1) [5]. Orally active anti-VEGF agents including sunitinib malate and ZM323881 effectively blocked hypoxia-induced retinal neovascularization in zebrafish. [6]. Two known antiangiogenic compounds, SU5416 and TNP470, which has shown antiangiogenic activity in mammalian system, have also demonstrated reduced vessel formation in zebrafish [7]. A range of known sedative compounds such as clozapine, fluoxetine, melatonin, diazepam, and pentobarbital have comparable response in zebrafish and all of these compounds resulted in reduced locomotor activity. [8–10]. Also, zebrafish also showed comparable responses to toxins for inducing pathologic consequences mimicking Parkinson's disease and epilepsy which will be addressed in later chapters. These evidences suggested that zebrafish demonstrate a good correlation with clinical relevance and support its potential as a model for pharmacological assessment [1, 3–5].
Table 1.
Discrepancies and similarities of the effect of drugs in human and zebrafish.
| Area of evaluation in zebrafish | Test compounds | Proportion of drugs with expected effects (%) | Reference |
|---|---|---|---|
| Inhibition of hERG or QTc prolongation | Study 1: Amiodarone, bepridil, cisapride, haloperidol, pimozide, procainamide, D,L-sotalol, terfenadine, thioridazine | All compounds, except for procainamide | [3] |
| Study 2: Negative controls: amoxicillin, aspirin Positive controls: chlorpromazine, cisapride, cromakalim, isoprenaline, moxifloxacin, nicotine, verapamil |
7 out of 9 compounds, including negative controls | [1] | |
|
| |||
| Visual safety or optomotor response | Study 1: 27 compounds, including 19 with positive and 8 with negative effects on inhibition of optomotor response | About 70% in overall showed the predicted drug effects. | [4] |
| Study 2: Negative control: aspirin Positive controls: chloroquine, chlorpromazine, diazepam, nicotine, ouabain, phenytoin, atropine, lithium |
7 out of 9 compounds including negative control | [1] | |
|
| |||
| Seizure liability | 25 drugs including 17 positive and 8 negative controls | 72% in overall | [5] |
|
| |||
| Gut contraction | Negative controls: aspirin and moxifloxacin Positive controls: amoxicillin, chlorpromazine, cisapride, cromakalim, isoprenaline, nicotine, nitrendipine, and verapamil |
5 out of 10 compounds including negative controls | [1] |
3. Similarity of Drug Metabolism between Zebrafish and Mammals: Omics Approach Provides a Clue
It is a common phenomenon that many compounds that occur naturally in metabolic tissues, such as liver and gastrointestinal tract, are inactive in vitro but are bioactivated in vivo into an active metabolite. One pioneer study with a zebrafish screen searched for cell-cycle modulators and identified 14 active candidates from a library of 2000 compounds (0.7% positive hits). The cell cycle-modulating activities of the active compounds identified from the zebrafish screen were validated in cell lines derived from both zebrafish and mammals [11]. Interestingly, only half of the active compounds were shown to be active in both embryos and either one of the cell lines, showing that some compounds are active in in vivo but inactive in vitro. The use of zebrafish for drug screening as well as for various pharmacological studies has received increasing attention in the fields of drug absorption, metabolism, distribution, and excretion [12]. However, there are few reports in the literature of detailed systematic studies investigating the fate of drugs after absorption as well as identification of the enzymes involved in drug metabolism in zebrafish larvae.
Our recent study addressed drug absorption and metabolism in zebrafish embryos and larvae. We used LC-MS/MS to identify and profile the metabolites of icaritin and its glycoside derivatives in zebrafish larvae [13]. Icaritin is a prenylated flavonoid compound that is regarded as an active ingredient of Herba Epimedii, which has been widely used in China as a medicinal herb for the treatment of infertility, osteoporosis, and weakness of the limbs. The result showed clearly that the metabolic pathway involving icaritin and its glycoside derivatives in zebrafish larvae is similar to that reported in mammals (Figure 1). The first step in the pathway is the enzymatic removal of the sugar moiety of these compounds after consumption in the cells of the gastrointestinal mucosa or by enzymes secreted by the colon flora [14]. Hydrolysis of the flavonoid derivative produces the free aglycone, which is conjugated by sulfation, glucuronidation, or methylation or in different combinations with steps that are controlled by phase II metabolism enzymes.
Figure 1.
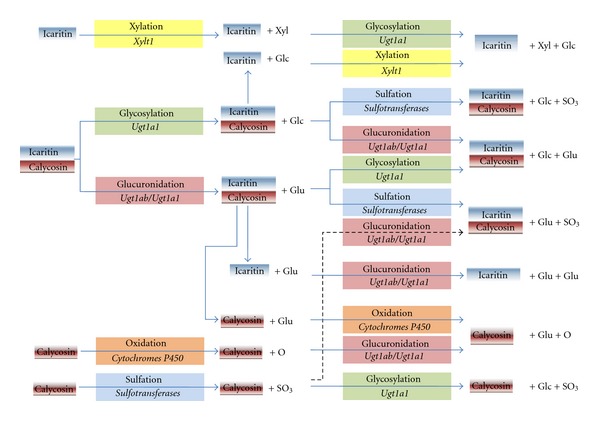
The metabolism of icaritin and calycosin in zebrafish embryos and larvae. The proposed routes of how icaritin and calycosin are metabolized in zebrafish embryos and larvae are summarized and some drug metabolism enzymes are identified by omics approach. The process and the corresponding gene are shown in each colored box. Glc: glycosylated group; Glu: glucuronidated group; Xyl: xylated group; SO3: sulfonated group; Xylt1: Xylosyltransferase 1; Ugt1a1, Ugt1ab: UDP-glucuronosyltransferase.
In order to investigate whether zebrafish larvae express the essential drug-metabolizing enzymes that are involved in the proposed metabolic pathways for the production of icaritin and its glycoside derivatives, combined transcriptomic and proteomic approaches were used to identify these enzymes [13]. In fact, transcriptomic profiling procedures identified 51 unique mRNA transcripts (out of a total of 13,310 nonredundant mRNA transcripts) that belong to three categories of key enzymes involved in phase I drug metabolism [15], including the cytochrome P450 family, flavin-containing monooxygenases, and epoxide hydrolases in zebrafish larvae. Moreover, mRNA transcripts of several key phase II drug metabolism enzymes [16], including UDP-glucuronosyltransferase, sulfotransferases, catechol-O-methyltransferase, and glutathione-S-transferases, were identified. However, the proteomic approach identified only three proteins (out of 2998 distinct proteins) that belong to the glutathione-S-transferases, a major type of phase II detoxification enzymes. The result illustrates that the metabolism of icaritin and its derivatives in both zebrafish larvae and mammalian models are highly conserved.
In addition, calycosin, an active constituent in Radix Astragali, was found to promote angiogenesis in zebrafish and human endothelial cells involving activation of the estrogen receptor and mitogen-activated protein kinase (MAPK) signaling pathway [17]. Our recently accepted paper characterizes drug absorption and metabolism using calycosin as a probe in zebrafish larvae [18]. Ten metabolites of calycosin produced by glucuronidation, glycosylation, sulfation, oxidation, or combinations of any two of these metabolisms in zebrafish larvae were identified by LC-MS/MS (Figure 1). The results showed the kinetic changes of calycosin and its metabolites in zebrafish larvae. This study identified drug metabolites previously identified in mammals, reconfirming the conservation of drug metabolism systems in zebrafish and identified novel metabolites, providing insight into the possibility of the discovery of novel drug metabolite diversity in zebrafish. In addition, the abundance of calycosin and its metabolites were increased steadily during 24 h after treatment [18], which reflects the difference of common drug administration between zebrafish and mammals. Unlike the common drug administration routes, such as gastric irrigation and oral administration, used in rodents, the drug treatment for zebrafish was usually performed by keeping the whole fish in a drug-containing incubation medium. This method keeps the zebrafish in an environment of constant drug concentration and drug compounds are continuously taken into the body through both the GI tract and the respiratory systems. Future in-depth systematic investigation of absorption, distribution, metabolism, and excretion (ADME) in zebrafish is warranted.
The high similarity of phase I and phase II metabolisms in zebrafish may be attributed to the highly conserved genetic expression profiles in liver as well as gut microbiota with human and mice counterparts, respectively [19, 20]. Drug screening in other small invertebrate model organisms, such as the fruit fly Drosophila melanogaster, has identified some very promising lead compounds, particularly for antiaging [21]. Nonetheless, the proof of concept of the highly conserved drug metabolism between zebrafish and mammals strongly supports the usefulness of zebrafish as a vertebrate model rather than other invertebrate model organisms for drug discovery as well as drug metabolism studies.
4. Presence of Blood-Brain Barrier (BBB) in Zebrafish
The BBB is crucial for the maintenance of a stable environment with the regulation of ionic balance and nutrient transport and the blockage of potentially toxic molecules. The intrinsic complexity of the cell-matrix-cell interactions of the neural-vascular unit has made analysis of gene function difficult in cell culture, tissue explants, and even animal models. The zebrafish has emerged as a premier vertebrate model for analyzing the complex cellular interactions in vivo and the genetic mechanisms of embryonic development [22]. Brain endothelial cells show immunoreactivity to Claudin-5 and Zonula Occludens-1 (ZO-1), implying the presence of tight junctions in these cells. The expression of Claudin-5 and ZO-1 was detected in cerebral microvessels starting from 3 dpf, concomitant with maturation of the BBB [23]. Zhang et al. observed that zebrafish embryos develop BBB functions by 3 dpf, with earlier expression of Claudin-5 in the central arteries at 2 dpf [24].
Our recent study of the neuroprotective effect of quercetin shed light on the presence of functional BBB in zebrafish larvae at 3 dpf and the role of BBB permeability in determining the beneficial effect of a neuroprotective drug in Parkinson's disease (PD) in in vivo. Quercetin is one of the commonest naturally occurring flavonoids. Although it and structurally related flavonoids have been shown to have a neuroprotective capacity in various in vitro and in vivo experimental models [25–27], the neuroprotective effect of quercetin remains controversial. Nevertheless, quercetin did not protect substantia nigra neurons from an oxidative insult in vivo, probably due to inefficiency in passing through the BBB in in vivo conditions [28]. There is an urgent need for appropriate in vivo studies in order to confirm the neuroprotective effect of quercetin and to identify the reason for the discrepancy between findings in vitro and in vivo. In order to address this controversy, we administered quercetin at different maturation stages of the BBB in zebrafish and we found it can prevent but not rescue the DA neuronal injury induced by 6-OHDA [28]. When quercetin was administered to zebrafish larvae before 3 dpf when BBB is not well established, it could spread rapidly throughout the brain and exert a protective effect against 6-OHDA toxicity. However, when quercetin was administered to zebrafish after 3 dpf, the matured BBB posed an obstacle to quercetin entering the brain, preventing it from rescuing 6-OHDA insult in dopaminergic (DA) neurons. This result supports earlier findings of the presence of BBB in zebrafish by 72 hpf [23, 24].
5. Behaviour Screen in Zebrafish
Zebrafish displays learning, sleeping, drug addiction, and neurobehavioral phenotypes that are quantifiable and comparable with those in human [10, 29]. A zebrafish behavioural assay has been demonstrated as a novel, rapid, and high-throughput approach to the discovery of neuroactive, psychoactive, and memory-modulating compounds [30–32]. In the past, a major obstacle to the discovery of psychoactive drugs was the inability to predict how small molecules will alter complex behaviours. Recently, Rihel et al. reported that the multidimensional nature of zebrafish phenotypes enabled the hierarchical clustering of molecules with comparable effects. This behavioural profiling revealed conserved functions of psychotropic molecules and predicted the mechanism of action of poorly characterized compounds [30]. In addition, Kokel and his colleagues used automated screening assays to evaluate thousands of chemical compounds and found that diverse classes of neuroactive molecules led to distinct patterns of locomotor behaviour. They concluded that a zebrafish behaviour assay can rapidly identify novel psychotropic chemicals and predict their molecular targets [31].
6. Zebrafish Bioassay Screening for Selectivity
Toxicity is now the first obstacle to drug development. From 2003–2010, the overall success rate for drugs passing from Phase I to FDA approval was only 9% [33]. A high percentage of drug developments failed at different stages, including animal testing or clinical trial, owing to nontolerated side effects and toxicity. As in vitro studies, which are usually cell based or molecular based, such as enzymatic or ligand-binding assays, drug screening with these assays predict the potential therapeutic action toward a specific molecular target and/or cell type; however, hidden toxicity and side effects due to interactions of the drug or its metabolites with other molecular targets, are not fully known.
Recently, a number of drugs were withdrawn from the market due to their human ether-a-gogo-related (hERG) cardiac toxicity [34]. The hERG potassium ion channel has a major role during the repolarization of the cardiac action potential, and the blockade of this ion channel can lead to prolongation of the QT interval, which is closely associated with torsade de pointes, a potentially lethal heart arrhythmia [35]. As a result, hERG (IKr) preclinical safety data are an essential part of any investigation of new drug submissions recommended in the FDA ICH guideline [36]. Zebrafish may present a good alternative model for large-scale screening of drug toxicity on QT prolongation through the ERG channel. hERG and its zebrafish homolog (zERG) have a high degree of similarity as zERG shows 99% conserved amino acid sequence in drug-binding and pore domains with the human ortholog [37]. Inhibition or knockdown of the zERG gene resulted in characteristic arrhythmia with 2 : 1 atrioventricular blockage (2 atrial beats coupled to 1 ventricular beat) [37]. The pharmaceutical industry has changed strategy by prescreening compound libraries for hERG cardiac toxicity before screening for therapeutic targets. According to the ICH S7A guidelines, CNS studies including behavior, learning and memory, neurochemistry, optomotor, and/or electrophysiology examinations are recommended before product approval [38]. Zebrafish may be a good model for the CNS assessment, since the animal possesses matched defined area in brain including hypothalamus and olfactory bulb [39]. The hippocampus was proposed to be located in the lateral zone of the pallium in zebrafish [39, 40]. In addition, important neurotransmitter systems such as the cholinergic, 5-hydroxytryptaminergic, dopaminergic, and noradrenergic pathways are also present in zebrafish brain [41, 42]. Zebrafish also has comparable neurological pharmacological response including locomotor activity [10], circadian pacemaking [43], and drug addiction [44] to human counterpart. These evidences support that zebrafish may be physiologically relevant model for screening out neurotoxic compounds.
Assessment of gastrointestinal complications may also be important during drug development, since the adverse reactions may result in death caused by gastrointestinal bleeding [45]. The zebrafish displayed similar physiology in gastrointestinal system with human. For example, the small intestine is lined with most of the cell types except Paneth cells [46, 47]; the peristalsis is controlled by a pair of smooth muscles and regulated by enteric nervous system [48]. However, it did not have a stomach [49] and a submucosa layer containing connective tissue to separate the epithelium from smooth muscle layer [46]. Moreover, in the study of the effect of 10 known compounds on gastrointestinal contraction in zebrafish, 5 out of 10 compounds showed expected effect [1] (Table 1). The relatively low predictability was due to the low reproducibility of cromakalim, nicotine, and nitrendipine in duplicated experiments [1] (Table 1). Nevertheless, zebrafish still has the potential for predicting adverse effects in gastrointestinal system [1]. There is increasing research on predicting the toxicity of a compound and excluding those compounds predicted to be toxic early in the drug discovery process [50].
Efficacy and toxicity are two important criteria for a drug to be marketed and the zebrafish model allows simultaneous measurement of these two parameters. The survival rate and/or mortality are/is a common and direct parameter used to indicate the toxicity of a compound. The beating heart of the embryo is the golden parameter used to indicate the living status of drug-treated zebrafish embryos. Thus, the lethal toxicity of a compound to zebrafish embryos reflected by the heartbeat rate could be monitored simultaneously with observation of the activity, such as antiangiogenesis, associated with the compound of interest. Moreover, other signs of toxicity, such as delayed development of zebrafish embryos, can be observed from the lower level of pigmentation in body and eyes, larger yolk sac, and shorter trunk in response to drug treatment. For instance, in our on-going screening of antiangiogenesis activities of a series of methoxyflavone derivatives, we identified structural modification in a single chemical group of the same scaffold, which exhibited higher potency of antiangiogenic activity and lower toxicity to zebrafish embryos [51]. This pilot study serves as proof of concept, suggesting the advantage of zebrafish over HUVEC cells as an angiogenic assay is that the zebrafish allows content screening of both activity and in vivo toxicity.
Along with studying an antiangiogenic compound in zebrafish, we could evaluate the selectivity of molecular action, such as cell-cycle arrest, to blood vessel cells in a live organism. Zebrafish embryos were trypsinized into a live cell suspension which was stained with the DNA-staining dye DRAQ5 (Biostatus Ltd., UK) for subsequent cell-cycle analysis (Figure 2). The differential effect of the compound on the cell cycle of endothelial GFP-expressing cells and the non-GFP-expressing cells could be determined by flow cytometry. Using this technique, the resveratrol derivative trans-3,5,4′-trimethoxystilbene (TMS) was found to induce cell-cycle arrest more significantly in endothelial cells (in about 20–30% of GFP-positive cells and in only 5–10% of GFP-negative cells) in zebrafish embryos (Figure 3), confirming that TMS exerted a more specific cytotoxic effect on endothelial cells than on other cell types in vitro and, more importantly, in vivo [52]. However, there was still an overall increase in G2/M phase cells in the whole cell population, indicating that TMS caused cell-cycle arrest in some other cell types (Figure 3). This finding provides a solution to the controversial issue regarding whether resveratrol and related compounds cause cell-cycle arrest through the G1 or G2/M phase in cell culture in vitro [53]. This study in zebrafish embryos, which showed the induction of G2/M cell-cycle arrest in GFP-positive endothelial cells by TMS in a whole live organism, provides insight into the physiological relevance of the compound. The concept of determining selectivity of antiangiogenic action on zebrafish endothelial cells was supported by the results of a similar study, in which nobiletin (5,6,7,8,3′,4′-hexamethoxyflavone) exhibited an effect on cell-cycle arrest differently via inducing G0/G1 phase accumulation in GFP-positive endothelial cells [54]. Besides analysis of the cell cycle, a similar approach could probably be adopted to probe cells for different cellular physiological parameters, such as oxidative stress and mitochondrial function, by different stains. This approach allows examination of how a candidate selective drug may act on specific cell types in a live organism.
Figure 2.
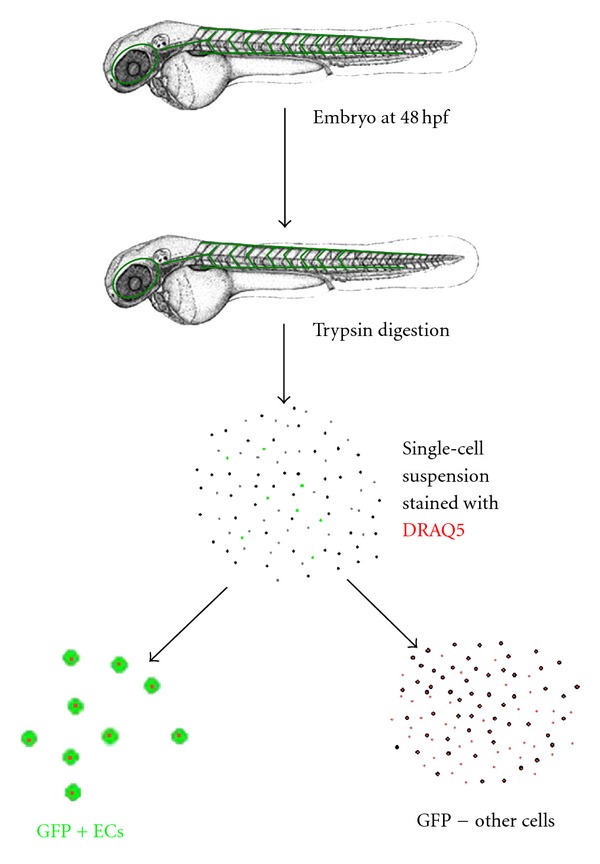
A diagram showing the processing of zebrafish embryos for isolating endothelial cells followed by staining with DRAQ5. Tg(fli-1:EGFP) zebrafish embryos are firstly trypsinized into a cell suspension, stained with DRAQ5, and separated into GFP expressing endothelial cells and others.
Figure 3.
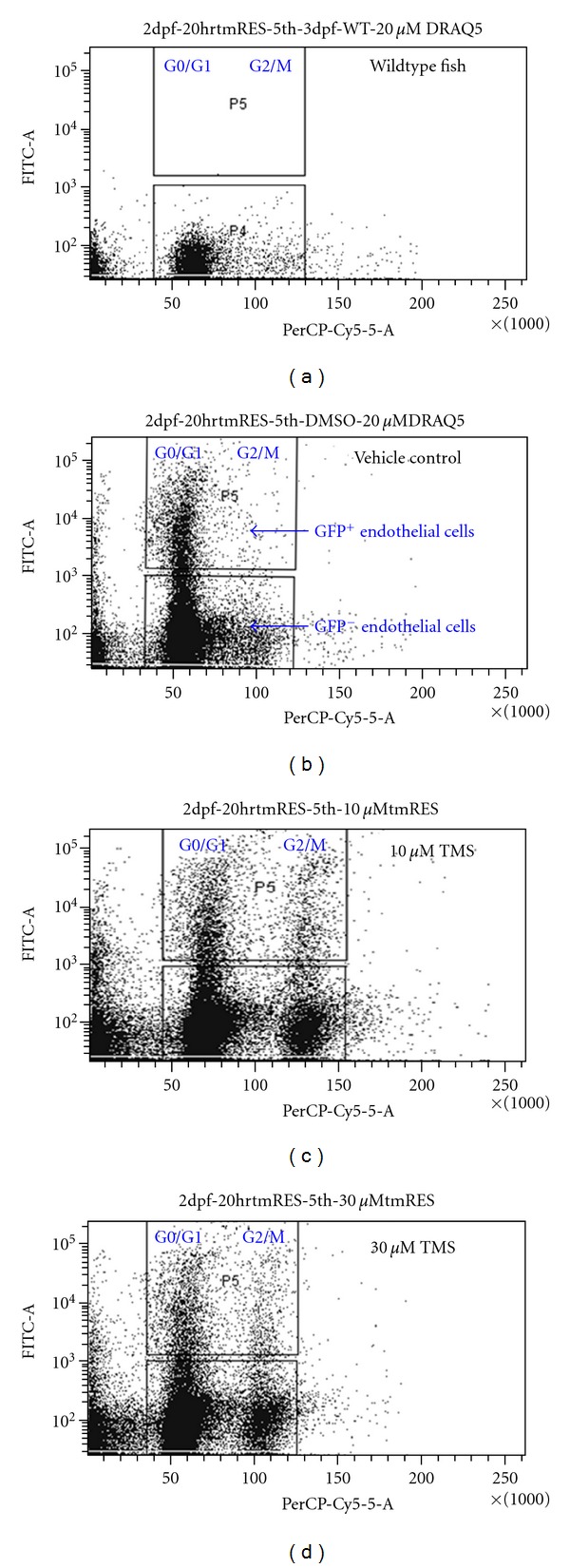
Cell-cycle analysis of zebrafish embryos after treatment with different concentration of an antiangiogenesis compound named trans-3,5,4′-trimethoxystilbene (TMS). Tg(fli-1:EGFP) zebrafish embryos treated with TMS for 20 h were then trypsinized and DRAQ5-stained for cell-cycle studies by flow cytometry. (a) Wildtype embryos did not show GFP-expressing-cells. (b) Tg(fli-1:EGFP) showed GFP expressing and non-GFP expressing cells. (c) 10 μM TMS and (d) 30 μM TMS exerted G2/M cell-cycle arrest preferentially in endothelial cells.
7. Phenomics and Biological Activity Profiling
Phenomics was originally an area of biology that involved studies of phenotype as a whole organism. Image-based bioassays reflecting changes of locomotor behaviour in the phenotype of different cell types, organs, and physiological systems in wildtype or transgenic zebrafish offer the opportunity to assess multiple pharmacological activities of a chemical compound (Figure 4). Pharmacological action of a compound could be decoded by a system biology approach through data mining of the multidimensional phenotypic data of an organism [55] together with measurement of the relative levels of mRNA transcripts (transcriptome), proteins (proteome), and metabolite components (metabolome). Recently, this omics approach has been incorporated increasingly into drug discovery and toxicology. Omics data provide much more information than typical phenotypic assay, including observable changes of morphology in the embryo as well as behaviour and mortality. By coupling omics data with an existing phenotypic end-point assay, more details of the mechanism and the toxicity of a chemical could be used to explain the cause-and-effect relationship. Even though the phenotypic changes are the same, such as vitellogenin, which indicate the estrogenic exposure in fish was upregulated by 17b-estradiol, bisphenol A, and genistein [56], the modes of action may differ. This possibility can be shown by differential gene expression induced by these chemicals based on transcriptome analysis [56].
Figure 4.
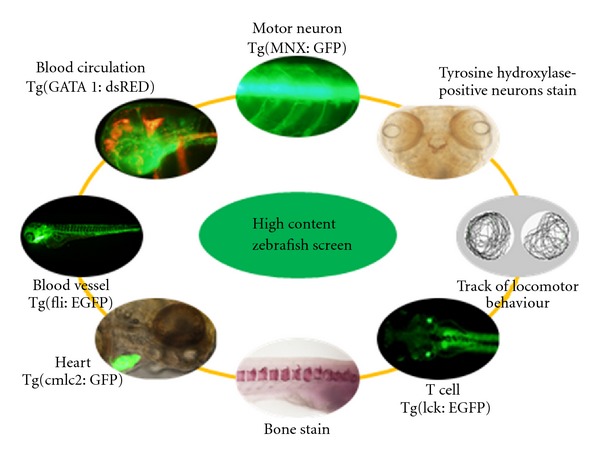
The examples of zebrafish model for high content drug screening. Image-based bioassays reflecting the physiological changes in either wildtype or transgenic zebrafish enable the assessment of multiple pharmacological activities of a chemical compound.
8. Integrative Transcriptomic and Proteomic Analysis of Zebrafish
Because the therapeutic action of a drug on normalizing pathological change can originate from different cellular pathways in the complex regulatory network, an in vivo study could provide considerably more information than in vitro assays using purified targets. mRNA transcripts and proteins are the primary molecules responsible for biological functions in cells and the ability to examine the transcriptome and the proteome of an organism provides a robust overview of the physiological changes taking place and could greatly augment target-oriented biological data. Therefore, we recently used an RNA-seq technology for transcriptome profiling and a fully automatable RP-RP 2DLC system for shotgun proteomics to address the drug metabolism system of zebrafish and the downstream transcriptional effect of a drug. The recent advancement of deep sequencing and 2D RP-RP LC-MS/MS technology identified a total of 12,560 mRNA transcripts (obtained from about 5 million reads per RNA sample) with matched annotated genes [13, 57] and 1752 unique proteins from the zebrafish lysate, respectively, in a single analysis [58].
The current transcriptome profiling tools used in zebrafish are microarray and RNA-seq. RNA-seq or deep sequencing of RNA samples using the next generation of sequencing technology is becoming a popular transcriptome profiling tool because it is an open platform that does not require predefined probes. In principle, RNA-seq profiles known and novel transcripts and it yields data with higher resolution, wider dynamic range, and lower background noise, while it only requires smaller amounts of RNA sample than microarrays [59].
By contrast, proteomic studies have used mainly integrated technologies, including separation of proteins by 2D polyacrylamide gel electrophoresis (2D-PAGE) and identification of proteins by matrix-assisted, laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS). Another major advantage of LC-MS/MS over conventional 2D PAGE is that the high sensitivity of LC allows for faster detection and direct identification of wider range of proteins, including high molecular weight proteins and very acidic or basic proteins, all of which are problematic when using methods such as 2D PAGE [58].
After transcriptomics, proteomics is often considered the next step in the study of biological systems. The integrated study of the two approaches is often considered as a study of causality. The focus is especially on identifying some biological response that initiates at the transcriptional level and exhibits functional information at the protein level. Transcriptomics has advantages over proteomics by allowing larger scale and higher throughput of analysis and about 10 times more coverage of detected gene targets in a single run of a zebrafish sample, while proteomics has advantages over transcriptomics in terms of potentially observing functional change in protein expression and posttranslational levels.
The methodology of conventional immunology depends on the availability of antibodies and a priori knowledge of the targets and, in general, is not amenable to global monitoring. Moreover, it is now known that the transcript level is often not correlated with the protein expression level, and the proteomic approach defines responses at the protein level that are probably not regulated at the transcription level, thus providing additional information [60]. Therefore, utilization of integrated transcriptomics and proteomics could provide more confirmatory evidence for the identification of molecular targets involved in the biological response of zebrafish to drug treatment. One of the advantages of integrating transcriptomics and proteomics lies in its neutrality to a priori knowledge and targets. The observation-driven result could lead to insights into previously unsuspected targets. The recent advancement of deep sequencing technology and the 2D LC-MS/MS system provides an unprecedented opportunity to formulate a system biology approach to unravel the resulting effect of holistic action of a drug or natural product containing multiple components through either interacting with a specific or multiple targets in a whole organism.
9. Zebrafish Disease Models
A highly relevant disease model should be developed when exploring the pathophysiological mechanism and the biological activity of any drug compound. Zebrafish models have been developed in several therapeutic areas for investigating human diseases. Disease models are created by mutation or inactivation of genes, treatment with chemicals, or even modification of a diet.
10. Neurodegenerative Disease Models
10.1. Parkinson's Disease
PD is the second most common neurodegenerative disease characterized by progressive loss of DA neurons in the substantia nigra pars compacta. The etiology of PD is not completely understood but increasing evidence suggests that oxidative damage induced by reactive oxygen species (ROS) and reactive nitric species (RNS) neuroinflammation, excitotoxicity, and apoptosis are involved in the progression of DA neurodegeneration [61].
Recently, the zebrafish has been demonstrated to be an appropriate model for PD [62]. The DA system in the posterior tuberculum of the ventral diencephalon is comparable with the nigrostriatal system in human [63]. PD-related neurotoxins cause the loss of DA neurons, reduced expression of tyrosine hydroxylase (TH) (Table 2) and the impairment of motor behaviour in zebrafish that are comparable with the pathophysiological features observed in other animal models [64]. In addition, clinical and experimental neuroprotective agents (nomifensine, a DAT inhibitor; L-deprenyl, an MAO-B inhibitor) (Table 2) have been demonstrated to be active in protecting zebrafish from neuronal insult [65]. Either knockdown or mutation of important genes, including PARKIN and LRRK2, contributes to a significant decrease in the number DA neurons (Table 2). Taken together, the results of earlier studies suggest that the zebrafish is a good alternative species for a PD model and offers great opportunity for screening and discovery of novel PD therapeutic agents.
Table 2.
Potential marker genes for PD.
| Gene | Function | Assessment method | Reference |
|---|---|---|---|
| Tyrosine hydroxylase (TH) | Catalytic conversion of the amino acid L-tyrosine to dihydroxyphenylalanine | Immunostaining, locomotion behaviour test | [28, 64, 66] |
| Dopamine transporter (DAT) | Membrane-spanning protein for pumping neurotransmitter DA back into cytosol from the synaptic region | Whole mount in situ hybridization (WISH), swimming behaviour | [65] |
| Vesicular monoamine transporter 2 (VMAT2) | Integral membrane protein for transporting neurotransmitter carrying monoamine structure, for example, dopamine and norepinephrine from cellular cytosol into synaptic vesicles | Visualization in VMAT2: GFP transgenic fish | [67] |
| MAO-B | Catalytic oxidation of monoamines | Monoamine oxidase enzyme histochemistry | [66] |
| PARKIN (PARK2) | Gene knockdown leads to complex I deficiency and dopaminergic neuronal cell loss | WISH, whole-mount antibody immunofluorescence, behaviour analysis | [68] |
| LRRK2 | Genetic mutant caused loss of DA neuron and locomotive defect | WISH, swimming behaviour | [69] |
The brain structure and function of the zebrafish are very similar to those of other vertebrates [70]. The anatomy of the zebrafish brain DA system was studied recently, and a region anatomically similar to the striatum was identified in the forebrain [71]. Neurotoxins, such as MPTP, 6-OHDA, and rotenone, are known to induce DA neuron loss in animal models. Among those neurotoxins, MPTP/MPP+ is the best characterized toxin to generate model of PD and has proved useful for studying the striatal circuitry involved in PD pathophysiology [72]. Exposure of zebrafish to MPTP caused profound loss of tyrosine hydroxylase-positive (TH+) neurons and downregulated TH mRNA expression in contrast to vehicle-treated healthy zebrafish (Figure 5) leading to a deficit in locomotor behaviour (Figure 6). Earlier studies revealed that 6-OHDA is taken up selectively by the plasma membrane dopamine transporter and subsequently accumulates in the mitochondria, resulting in the formation of ROS and RNS [73]. In addition, neuroinflammation plays a key role in 6-OHDA-induced DA neuron damage in vivo [74]. We measured the gene expression of proinflammatory mediators in 6-OHDA-treated zebrafish by quantitative real-time PCR and showed that 6-OHDA caused overexpression of IL-1β, TNF-α, and COX-2, several-fold higher than that of untreated control fish [75]. These proinflammatory genes play important roles in the etiology of PD [76]. It has been shown that the level of the COX-2 protein is upregulated in substantia nigra DA neurons in PD patients and in animal models [77]. The inhibition of COX-2 and TNF-α has provided neuroprotection in rats [76]. Our current iTRAQ-based shotgun proteomics study in a zebrafish model for PD suggested the potential involvement of both TNF-α/NF-κB and oxidative phosphorylation pathways in 6-OHDA-induced neurodegeneration in zebrafish (unpublished data). However, given that, all reported promising studies on this chemical induced PD experimental zebrafish model, more researches need to be done to differentiate systemic toxicity and selective neuronal toxicity of the neurotoxins. In addition, generation of transgenic zebrafish expressing fluorescent protein specifically in DA neuron, that allows tracking the kinetic change of living DA neurons in vivo, is a viable strategy to replace the postimmunochemical staining of TH-positive neurons.
Figure 5.
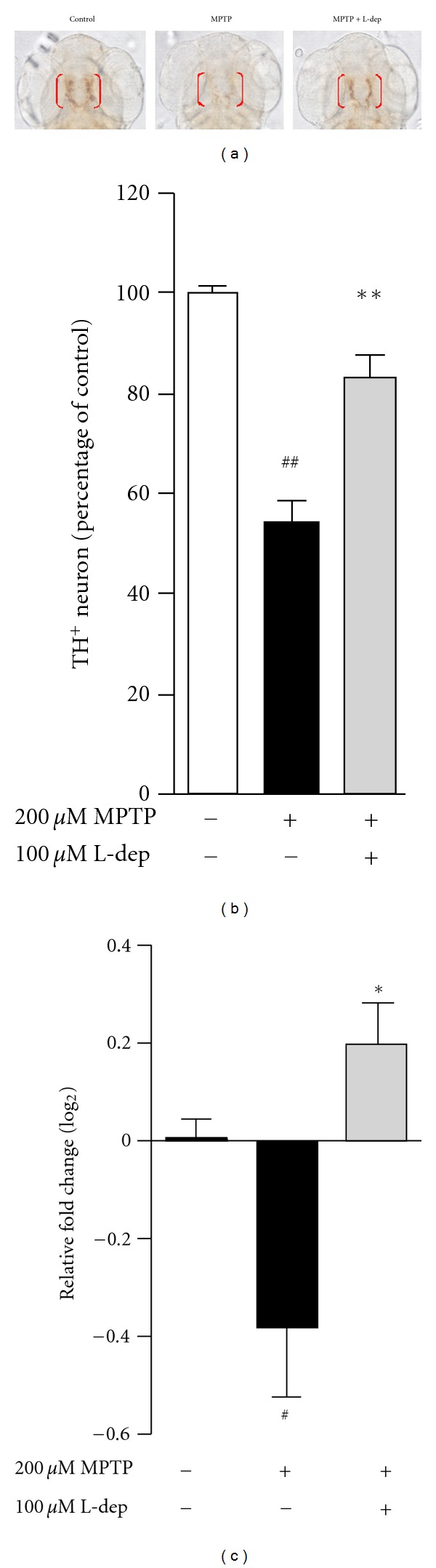
MPTP induces DA neuron loss in zebrafish. (a) Representative picture of anti-TH whole mount immunostaining. TH+ neurons in diencephalic region were indicated by bracket, dorsal view. L-dep, L-deprenyl (selegiline), a selective MAO-B inhibitor, was used as positive control. (b) Counting of TH+ neuron. (c) Relative fold change of th gene expression as compared to control, MPTP downregulated th gene expression. # P < 0.05 and ## P < 0.01 compared with untreated control. *P < 0.05 and **P < 0.01 compared with MPTP treated alone.
Figure 6.
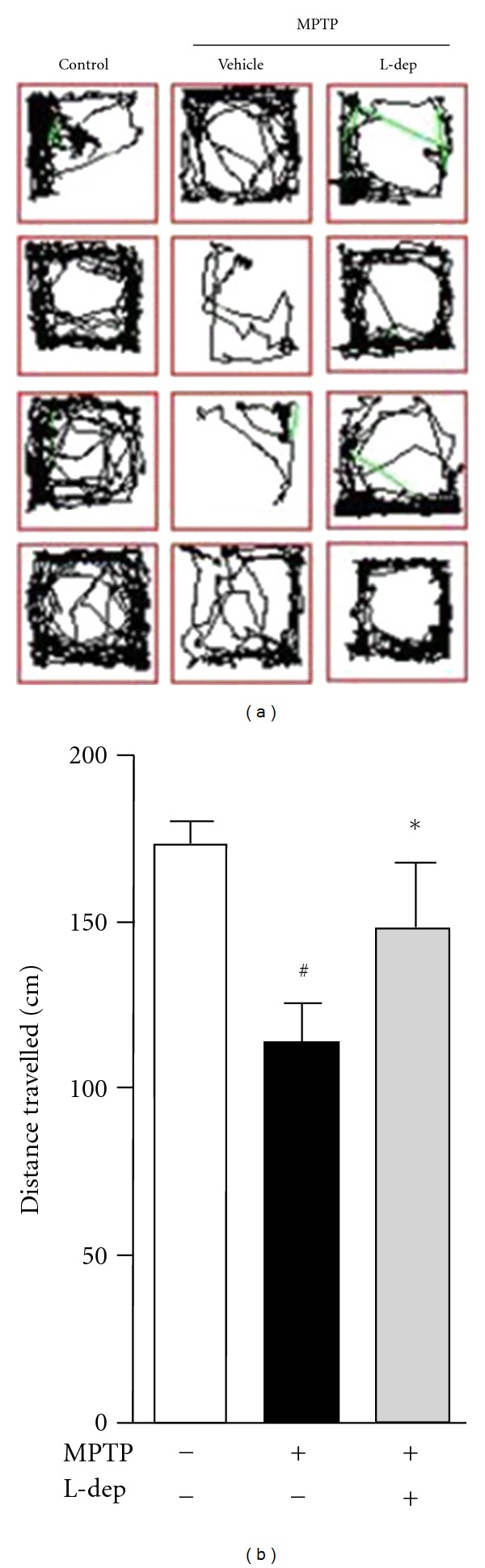
MPTP induces deficit of swimming behavior in zebrafish. (a) Typical swimming patterns of control and MPTP-treated zebrafish. Lines show the track of zebrafish movement. Zebrafish treated with MPTP was less active as compared to the control. (b) Quantitative analysis of total distance travelled. # P < 0.05 compared with untreated control. *P < 0.05 compared with MPTP treated alone.
10.2. Epilepsy
Epilepsy is a common neurological disorder characterized by the recurrent appearance of spontaneous seizures due to neuronal hyperactivity, and the disease afflicts nearly 50 million people worldwide [78]. Recent studies showed that the pathogenesis of epilepsy involves altered distribution of GABA receptors (Table 3), enhanced activity of excitatory circuits, neuronal loss, and synaptic reorganization [79–82]. A number of genes encoding transcription factors, synaptic receptors, ion channels, and glucose transporters have shown altered mRNA expression in rodent models of epilepsy (Table 3). These findings suggest potential gene markers other than c-fos. Although some antiepileptic drugs (AEDs) are marketed, there is no drug capable of reversing the cause of pathological changes in the brain [83] and some disease subtypes, such as temporal lobe epilepsy, are even resistant to current pharmacotherapies [84]. This problem calls for large-scale screening of new candidates of AED, but this is difficult to achieve in a rodent model.
Table 3.
Potential marker genes for epilepsy.
| Function | Gene | Assessment method | References |
|---|---|---|---|
| Transcription factor | c-Fos | Immunohistochemistry, In situ hybridization, real-time PCR | [85] |
| c-Jun | Electrophoretic mobility-shift assay | [86] | |
| CREB | Real-time PCR, northern blot | [87] | |
| Zac 1 | Immunohistochemistry, In situ hybridization | [88] | |
| Receptor | NMDAR1 | Immunohistochemistry, Western blot | [89] |
| GABA(A)-receptor delta | Immunohistochemistry | [81] | |
| Ion channel | Kv1.2 and Kv4.2 | In situ hybridization | [90] |
| Transporter | GLUT1 and GLUT3 | In situ hybridization, Western blot | [91] |
A PTZ-induced epilepsy model of zebrafish was established by Baraban in 2005 [85], who reported the upregulation of c-fos in the CNS region of zebrafish exposed to PTZ. After exposure to PTZ, the larval zebrafish shows three stages of seizure: a dramatic increase in total distance travelled at Stage I, rapid whirlpool-like circling swimming behaviour at Stage II, and culmination in clonus-like convulsions leading to loss of posture at Stage III. Current AEDs can stop the seizure at Stages I and II and, therefore, epileptic zebrafish at both stages are suitable for drug screening [92].
10.3. Heart Disease and Cardiotoxicity
Mutations found in cardiac troponin T type 2 (TNNT2) [93] and T-box-5 (Tbx5) are implicated in cardiomyopathy. Severe heart defect was observed in zebrafish carrying the mutated TNNT2. Mutation in Tbx5 leads to the maldevelopment of heart and upper limbs known as Holt-Oram syndrome [94]. Zebrafish carrying the same mutation have comparable deformed heart and pectoral fins [95]. In fact, troponin T was considered as a biomarker in congenital heart failure from dilated cardiomyopathy [96, 97]. Other biomarkers, such as myosin light chain-I [96, 97], cardiotrophin [98], and endothelin-1 [99], are proposed to have diagnostic value in congestive heart failure and hypertension (Table 4).
Table 4.
Potential biomarkers for human heart disease.
| Gene | Function | Assessment method | Associated cardiovascular disease |
|---|---|---|---|
| Troponin T | Myocardial contraction | ELISA | Congestive heart failure [96, 97] |
| Heart fatty acid binding protein | Carrier proteins for fatty acids and other lipophilic substances, such as eicosanoids and retinoids | ELISA | Congestive heart failure [96, 97] |
| Myosin light chain-I | Myocardial contraction | ELISA | Congestive heart failure [96, 100] |
| Creatine kinase MB | Energy metabolism | ELISA | Congestive heart failure [96] |
| Cardiotrophin-1 | Response to stress and humoral factors such as angiotensin II | ELISA | Hypertension [98] |
| Endothelin-1 | Potent endothelium-derived vasoconstrictor peptide | Radioimmunoassay | Heart failure [99] |
The zebrafish is a good model for studying cardiotoxicity. The cardiac function can be studied in zebrafish embryos through assessment of heart rate, heart morphology, cardiac myocytes number, and heart size [101]. Recently, we explored the cardiotoxicity of chemotherapeutic agents such as sunitinib malate (Sutent; SU11248; Pfizer). Sutent is a multitargeted tyrosine kinase inhibitor with antiangiogenic activity. It has been approved for first-line and adjuvant treatment of renal cell carcinoma. However, long-term angiogenesis inhibition would involve unwanted side effects, including cardiac and renal toxicity in patients with cancer [102]. Our study showed that Sutent deteriorates heart function through induction of pericardial edema and decrease in heart rate in zebrafish embryos (Figure 7).
Figure 7.
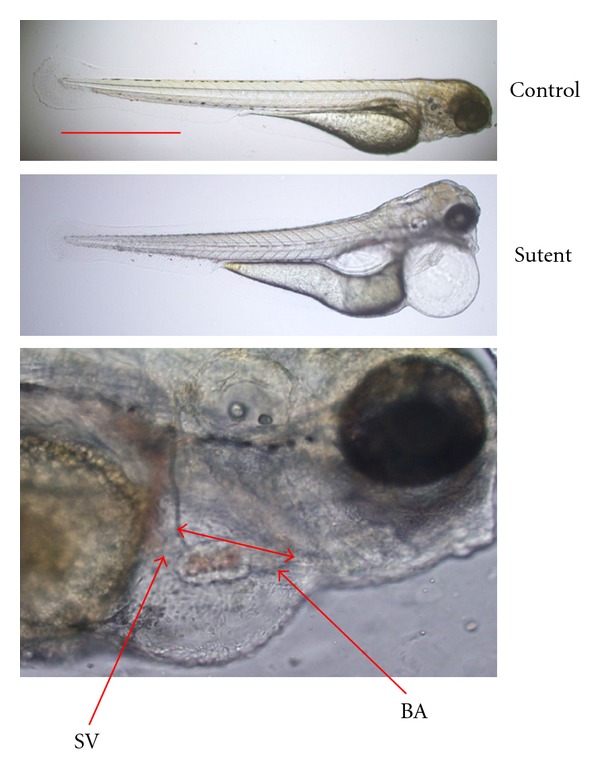
Sutent-induced cardiotoxicity in zebrafish embryos. Embryos at 5 dpf were treated with Sutent for 72 h followed by assessment of cardiac function. (a) Pericardial edema was observed after administration of Sutent compared to the control. The severity of pericardial edema was quantified by measuring the distance between sinus venosus (SV) and bulbus arteriosus (BA). (b) Embryo heart rate was decreased by treatment with Sutent in dose-dependent manner.
11. Cerebral Hemorrhage Model
Cerebral hemorrhage, also known as hemorrhagic stroke, occurs when a blood vessel in the brain becomes weak and bursts, allowing blood to leak into the brain. Atorvastatin, a 3-hydroxy-3-methylglutaryl coenzyme-A (HMG-CoA) reductase inhibitor, reduces cholesterol, ameliorates, and vascular atherosclerosis and improves cardiovascular morbidity and mortality [103]. Pretreatment with atorvastatin significantly reduced infarct volume induced by permanent middle cerebral artery occlusion in animal studies [104]. Clinical studies showed patients with postischemic-stroke treatment with atorvastatin showed improving neurological recovery [105]. However, this beneficial effect is partly counteracted by an increased risk of hemorrhagic stroke [106]. Moreover, atorvastatin induced intracranial hemorrhages in wildtype fish [107] and induced cerebral hemorrhage in a zebrafish model (Figure 8), which offers an opportunity to screen cerebrovascular-protective compounds.
Figure 8.
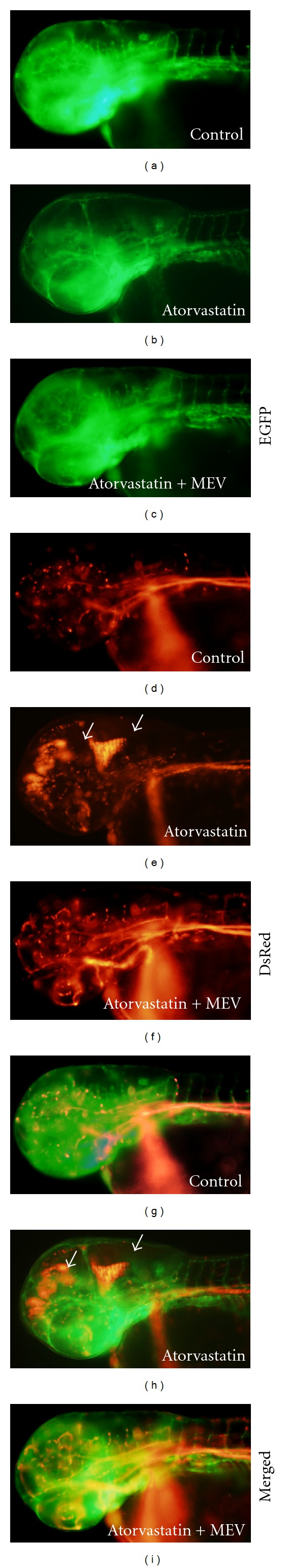
Atorvastatin-induced cerebral hemorrhage in zebrafish embryos. Tg(fli1:EGFP); Tg(gata1:dsred) homozygous double transgenic zebrafish embryos at 24 hpf were treated with atorvastatin alone or with atorvastatin and mevalonate (MEV) in combination for 24 h. Images showing blood vessels (EGFP, green) were superimposed on images showing blood flow (DsRed, red). Hemorrhage observed in the atorvastatin treatment group was prevented by cotreatment with MEV. Fluorescent microscopic images are at magnification 100x.
12. Dyslipidemia and Hyperlipidemia
The zebrafish model can be used in the study of lipid metabolism. The quenched fluorescent phospholipid substrate N-((6-(2,4-dinitrophenyl)amino)hexanoyl)-1-palmitoyl-2-BODIPY-FL-pentanoyl-sn-glycero-3-phosphoethanolamine (PED6) taken up by zebrafish larvae can fluoresce after cleavage by phospholipases in the intestine. It has been reported that this assay can be used to detect the fat-free (ffr) mutation, which likely results in disturbed lipid processing through impaired intestinal phospholipase activity [108] and reduced protease activity [109]. A research team led by Stoletov has developed a hypercholesterolemic (HCD) model in zebrafish utilizing a fluorescent cholesteryl ester to observe vascular lipid accumulation and fluorescent dextran in the endothelial cell layer disorganization after an HCD diet [110]. The reliability of the model was further supported by accumulation of macrophages, increased phospholipase A2 activity, and elevated levels of oxidized phosphatidylcholines in zebrafish fed an HCD diet compared to those fed a normal diet [110, 111]. Another research group led by Jin has demonstrated the antiatherosclerotic effect of turmeric and laurel aqueous extracts using this HCD model [112]. These disease models have proved to be highly relevant to human diseases and showed a number of conserved phenotype between zebrafish and human. Moreover, the difficulty of studying the atherogenic events in a temporal manner has been overcome due to its optical transparency.
13. Searching for Active Compounds from Natural Products
Many natural products exhibit a range of biological activity that is probably due to interaction of their complex chemical constituents with multiple targets in the body, which opens new avenues for therapy of disorders, with multifactorial etiopathogenesis such as neurodegeneration. The physiological complexity of zebrafish is similar to that of mammals, providing a suitable model for the study of human diseases as well as throughput drug screens. Using a whole organism as a model allows a more comprehensive and simultaneous analysis of the range of biological activity and toxicity of a chemical or multiple chemicals compared to an in vitro assay. Zebrafish embryos and early larvae are optically transparent, allowing screens with a measurable phenotypic readout using imaging microscopy for assessing pathological changes in Parkinson's disease, epilepsy, heart disease or cardiotoxicity, cerebral hemorrhage, and hyperlipidemia. This approach allows live and continuous observation on individuals which are often inapplicable in other in vivo models. More importantly, invasive approaches are often applied in these animal models, so reassessment of individuals may not be possible. For example, cerebral hemorrhage in rodent models was commonly done by intraparenchymal infusion of either autologous blood or bacterial collagenase. The hematoma size and location were evaluated with histologic analysis [113, 114]; hyperlipidemia in rodent models was achieved by feeding ApoE deficient mice with high fat diet for eight weeks and the atherosclerotic lesion was also observed by histology [115, 116]. Also, visual observation with imaging microscopy may require less technical skills and also far more convenient. For example, the evaluation of cardiotoxicity in zebrafish was determined by heart rate, heart morphology, cardiac myocyte number, and heart size. However, in rodent models, the heart function was often assessed by electrocardiogram or echocardiogram which requires intensive technical and labor input. Using rodent disease models for early stage drug screening may sound inapplicable since high-throughput studies are usually required. In addition, zebrafish model enables the observation of any pharmacological effect(s) on multiple targets underlying the pathway of a disease or a normal physiological process can be observed. The zebrafish model is, therefore, very suitable for identifying the off-target effects or multiple targets due to the holistic action of natural products.
14. Identifying Angiogenic Compounds from Natural Products
Angiogenesis is the establishment of the mature blood vessel network through expansion and remodeling of the vascular primordium. Blood vessel formation through angiogenesis involves the induction of new sprouts, coordinated and directed endothelial cell migration, proliferation, sprout fusion (anastomosis), and lumen formation [117]. Under normal conditions, tiny vessels do not increase in size or number, except in wound healing, embryonic development, and development of the corpus luteum. In fact, many diseases are associated with an imbalance in the regulation of angiogenesis, in which either excessive or insufficient blood vessel formation occurs.
To evaluate the angiogenic response in zebrafish, transgenic fish expressing green fluorescent protein (GFP) specifically in endothelial cells, for example, Tg(fli-1:EGFP) and Tg(fli-1:nEGFP), are recently used for rapid analysis of changed vasculature in live embryos in response to drugs [118]. In fact, zebrafish is an excellent animal model for the study of angiogenesis, with many antiangiogenic drugs eliciting responses similar to those in mammalian systems [119]. During the vasculature development, subintestinal vein vessels (SIVs) originate from the duct of Cuvier at 48 hpf and form a vascular basket in the yolk sac during the next 24 h. The angiogenic response was evaluated visually with respect to the following criteria: (1) the appearance of spikes or sprouts projecting from the subintestinal vessel basket or the lengthening of such spikes; (2) the extension of the basket into the yolk region with more than seven vertical branches within the basket [120]; (3) statistical increases in diameter compared to the medium control; (4) the ectopic growth of newly formed blood vessels from SIVs and increased numbers of SIVs in the endothelial cells [121]. Sprout formation was seen as the main characteristic in proangiogenesis [17, 120, 121]. Recently, we demonstrated the feasibility of drug screening in a zebrafish model and found the antiangiogenesis effect of a resveratrol derivative [52], indirubin [122], nobiletin [54], and sinensetin [51] as well as proangiogenesis effects of Angelica sinensis extract [123, 124], Panax notoginseng extract [125], and Radix Astragali extract [17].
Angiogenesis plays an important role in the development of human chronic inflammatory diseases, including cancer, psoriasis, rheumatoid arthritis, macular degeneration, and diabetes retinopathy [126, 127]. There is growing evidence that chronic inflammation and angiogenesis are codependent, involving increased cellular infiltration and proliferation as well as overlapping roles of regulatory growth factors and cytokines [126]. Persistent inflammation is linked with the progression of cancer, as proinflammatory cytokines are detected frequently in tumor tissue[128]. In rheumatoid arthritis, the formation of pannus [129], which is an inflammatory connective tissue mass rich in blood vessels, is apparently because angiogenic factors, such as VEGF, stimulate encephalitogenic T cells and induce more severe and prolonged encephalomyelitis [130]. Besides angiogenic factors, transcription factors such as NF-κB plays a central role in the signaling of apoptosis and inflammation [131]. NF-κB expression is associated with VEGF in the development and progression of tumorigenesis [132]. Signaling by the cyclooxygenase-2 (COX-2) downstream of NF-κB may play a key role in the tumorigenesis of a variety of human malignancies by stimulating cell proliferation and angiogenesis [133]. Moreover, a recent study showed that chronic inflammation in benign prostatic hyperplasia causes an overexpression of COX-2, which induces the increased expression of Bcl-2 and VEGF [134].
Pharmacology of many anti-inflammatory drugs revealed at least some part of their efficacy is due to their antiangiogenic effect [126]. Tocotrienol, a member of the vitamin E family, possesses anticancer properties acting through regulating multiple signaling pathways, including anti-inflammation and antiangiogenesis [135]. The extract of Physalis angulate shows antimetastatic and antiangiogenic activity in human oral squamous carcinoma and human umbilical vein endothelial cells, probably due to its anti-inflammatory properties [136]. Indirubin inhibits inflammatory reactions by suppressing the production of interferon-γ and interleukin-6, which is a well-known inflammatory cytokine [137]. Interestingly, it also displays antiangiogenic activity by inducing HUVEC apoptosis and cell-cycle arrest at the G0/G1 phase [122]. Resveratrol and its derivative exert antiangiogenic and vascular-disrupting effects in zebrafish through downregulation of VEGFR2 and cell-cycle modulation [52]. The anti-inflammatory property of resveratrol is reported to prevent an increase in the levels of serum amyloid A, tumor necrosis factor-α, interleukin (IL-6), IL-1β, and nuclear transcription factor-κB in colitis-associated disease [138].
Angiogenesis deficiencies are associated with numerous human cardiovascular and cerebrovascular diseases (e.g., ischemic cardiac and cerebral problems). Our previous discovery of a pro-angiogenic herb called Angelica sinensis by zebrafish assay leading to development of a wound healing formulation for diabetic foot ulcer patients [124, 139]. Our recent work presented, for the first time, that a chemical-induced blood vessel loss in zebrafish in vivo could mimic angiogenesis deficiencies associated with human disease conditions and be used to identify pro-angiogenic agents. VEGFR tyrosine kinase inhibitor II (VRI), a pyridinyl-anthranilamide compound that displays antiangiogenic properties, strongly inhibits the kinase activities of both VEGF receptor 1 and 2. Treatment of the zebrafish with VRI induces significant blood vessel loss in ISV (intersegmental vessels) and DLAV (dorsal longitudinal anastomotic vessels). For instance, we identified a polysaccharide fraction (50000 D < MW and DM < 0.1 μm) isolated from Astragali Radix partially restores the chemical-induced blood vessel loss in the zebrafish model [140]. This is also the first study to prove the concept of screening the bioactivity of polysaccharides in live zebrafish, whose drug metabolism systems were shown recently to have a high degree of functional similarity to that of mammals. Since polysaccharides isolated from natural products usually undergo the enzymatic breakdown of the sugar moiety in the cells of the gastrointestinal mucosa, or by enzymes secreted by the colon flora, to become active metabolites after oral consumption by humans, the study of the bioactivity of the polysaccharides required the development of an in vivo assay equipped with mammalian-equivalent drug metabolism systems. Our findings provide insight into a new angiogenesis deficiency zebrafish model for screening vascular regenerative agents as well as the important roles of various substances from Chinese medicines for the treatment of various pathological conditions associated with deficient angiogenesis, such as ageing, stroke, ulcers, and cardiovascular diseases [140, 141].
15. Identifying Anti-Parkinson's Disease Compounds from Natural Products
PD patients usually suffer primarily from the death of dopaminergic (DA) neurons in the substantia nigra. Recent research in the pharmacotherapy of PD has identified numerous agents for the symptomatic control of motor impairments, but none is able to prevent, slow, or halt the progression of the disease [142]. The main obstacle to developing neuroprotective therapies is our limited understanding of the key molecular events that provoke neurodegeneration. Earlier studies highlighted the pathological involvement of oxidative stress, neuroinflammation, excitotoxicity, and apoptosis in neurodegenerative diseases [143]. Because PD, as well as other neurodegenerative disorders, usually has multifactorial etiopathogenesis, multiple drug therapy is required to address the varied pathological aspects [144]. Multiple drug strategy has been the essence of the rationales used for formulating traditional Chinese medicines (TCMs) for thousands of years. TCMs contain a mixture of chemical components from a single herb or a combination of several herbs and thus versatile functions and possess great potential in the multitarget approach for improved treatment of complicated diseases, such as PD.
By combining whole mount immunostaining and a behavioural screen, we have identified the neuroprotective activity of a few TCMs, including Fructus Alpinia oxyphylla extract (AOE) and Eriocaulon buergerianum extract (EBE) [75, 145]. Recently, increasing evidence suggests the beneficial effects of Fructus AOE on various neurodegenerative diseases. Treatment with aqueous AOE attenuated the death of cortical astrocytes induced by amyloid-β (Aβ) in vitro, prevented ischemia-induced learning disability, and rescued hippocampal CA1 neurons from lethal ischemic damage in mice [146]. Treatment with the ethanolic AOE in the presence of glutamate significantly enhanced viability and reduced apoptosis in a cortical neuron culture [147]. We found that ethanolic AOE prevented and restored 6-OHDA-induced DA neurodegeneration and attenuated the deficit of locomotor activity in zebrafish [75]. In addition, the aquatic plant EB (Gujingcao) is a TCM with anti-inflammatory and antimicrobial properties [148]. In the Chinese Pharmacopoeia (2005), the capitulum of EB is one of the most frequently used Chinese medicinal herbs, with flavonoids, volatile oils, anthraquinone, naphthopyranones, protocatechuic acid, and c-tocopheryl acetate being the bioactive constituents [149]. Flavonoids such as patuletin hispidulin, quercetin, quercetagetin, and quercetagetin derivatives and volatile oil such as palmitic acid, (Z,Z)-9,12-octacosane-dienoic acid are the two major classes of chemicals in EB [148]. EB demonstrates significant therapeutic effects on headache, toothache, nasosinusitis, night blindness, glaucoma, retinochoroiditis, conjunctivitis, and other eye diseases [150]. The results of our study suggested that EBE has profound neuroprotective activity in zebrafish, including the dose-dependent recovery of DA neuron loss caused by 6-OHDA in vivo and inhibition of the 6-OHDA-induced decrease of total movement distance in zebrafish [145]. We found that quercetin was one of the active neuroprotective constituents in EBE [28]. All these groundwork warrants further study of how the interaction of multiple components in these natural products elicits neuroprotection.
16. Conclusions
Zebrafish offers interesting possibilities for the simultaneous assessment of efficacy and toxicity of target compounds, which is not easily addressed with current rodent models. With its physiological similarities to human, many disease models could be established for identifying the off-target and the targeted effects of target compounds. More importantly, it allows integrative studies of transcriptomics and proteomics for identifying drug metabolic pathways and known or novel molecular targets involved in the biological response of zebrafish to drug treatment.
Acknowledgments
This study was supported by grants from the Science and Technology Development Fund of Macau SAR (Ref. nos. 045/2007, 058/2009, 014/2011 and 078/2011/A3) and the Research Committee, University of Macau (Ref. no. UL017).
References
- 1.Berghmans S, Butler P, Goldsmith P, et al. Zebrafish based assays for the assessment of cardiac, visual and gut function—potential safety screens for early drug discovery. Journal of Pharmacological and Toxicological Methods. 2008;58(1):59–68. doi: 10.1016/j.vascn.2008.05.130. [DOI] [PubMed] [Google Scholar]
- 2.Lieschke GJ, Currie PD. Animal models of human disease: zebrafish swim into view. Nature Reviews Genetics. 2007;8(5):353–367. doi: 10.1038/nrg2091. [DOI] [PubMed] [Google Scholar]
- 3.Mittelstadt SW, Hemenway CL, Craig MP, Hove JR. Evaluation of zebrafish embryos as a model for assessing inhibition of hERG. Journal of Pharmacological and Toxicological Methods. 2008;57(2):100–105. doi: 10.1016/j.vascn.2007.10.004. [DOI] [PubMed] [Google Scholar]
- 4.Richards FM, Alderton WK, Kimber GM, et al. Validation of the use of zebrafish larvae in visual safety assessment. Journal of Pharmacological and Toxicological Methods. 2008;58(1):50–58. doi: 10.1016/j.vascn.2008.04.002. [DOI] [PubMed] [Google Scholar]
- 5.Winter MJ, Redfern WS, Hayfield AJ, Owen SF, Valentin JP, Hutchinson TH. Validation of a larval zebrafish locomotor assay for assessing the seizure liability of early-stage development drugs. Journal of Pharmacological and Toxicological Methods. 2008;57(3):176–187. doi: 10.1016/j.vascn.2008.01.004. [DOI] [PubMed] [Google Scholar]
- 6.Cao R, Jensen LDE, Söll I, Hauptmann G, Cao Y. Hypoxia-induced retinal angiogenesis in zebrafish as a model to study retinopathy. PLoS ONE. 2008;3(7) doi: 10.1371/journal.pone.0002748.e2748 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Serbedzija GN, Flynn E, Willett CE. Zebrafish angiogenesis: a new model for drug screening. Angiogenesis. 1999;3(4):353–359. doi: 10.1023/a:1026598300052. [DOI] [PubMed] [Google Scholar]
- 8.Airhart MJ, Lee DH, Wilson TD, Miller BE, Miller MN, Skalko RG. Movement disorders and neurochemical changes in zebrafish larvae after bath exposure to fluoxetine (PROZAC) Neurotoxicology and Teratology. 2007;29(6):652–664. doi: 10.1016/j.ntt.2007.07.005. [DOI] [PubMed] [Google Scholar]
- 9.Boehmler W, Carr T, Thisse C, Thisse B, Canfield VA, Levenson R. D4 Dopamine receptor genes of zebrafish and effects of the antipsychotic clozapine on larval swimming behaviour. Genes, Brain and Behavior. 2007;6(2):155–166. doi: 10.1111/j.1601-183X.2006.00243.x. [DOI] [PubMed] [Google Scholar]
- 10.Zhdanova IV, Wang SY, Leclair OU, Danilova NP. Melatonin promotes sleep-like state in zebrafish. Brain Research. 2001;903(1-2):263–268. doi: 10.1016/s0006-8993(01)02444-1. [DOI] [PubMed] [Google Scholar]
- 11.Murphey RD, Stern HM, Straub CT, Zon LI. A chemical genetic screen for cell cycle inhibitors in zebrafish embryos. Chemical Biology and Drug Design. 2006;68(4):213–219. doi: 10.1111/j.1747-0285.2006.00439.x. [DOI] [PubMed] [Google Scholar]
- 12.Rawls JF, Samuel BS, Gordon JI. Gnotobiotic zebrafish reveal evolutionarily conserved responses to the gut microbiota. Proceedings of the National Academy of Sciences of the United States of America. 2004;101(13):4596–4601. doi: 10.1073/pnas.0400706101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Li ZH, Alex D, Siu SO, et al. Combined in vivo imaging and omics approaches reveal metabolism of icaritin and its glycosides in zebrafish larvae. Molecular BioSystems. 2011;7(7):2128–2138. doi: 10.1039/c1mb00001b. [DOI] [PubMed] [Google Scholar]
- 14.Walle T. Absorption and metabolism of flavonoids. Free Radical Biology and Medicine. 2004;36(7):829–837. doi: 10.1016/j.freeradbiomed.2004.01.002. [DOI] [PubMed] [Google Scholar]
- 15.Brunton LL, Goodman LS, Blumenthal D, et al. Goodman and Gilman’s Manual of Pharmacology and Therapeutics. New York, NY, USA: McGraw-Hill Professional; 2007. [Google Scholar]
- 16.Jancova P, Anzenbacher P, Anzenbacherova E. Phase II drug metabolizing enzymes. Biomedical papers of the Medical Faculty of the University Palacký. 2010;154(2):103–116. doi: 10.5507/bp.2010.017. [DOI] [PubMed] [Google Scholar]
- 17.Tang JY, Li S, Li ZH, et al. Calycosin promotes angiogenesis involving estrogen receptor and mitogen-activated protein kinase (MAPK) signaling pathway in zebrafish and HUVEC. PLoS ONE. 2010;5(7) doi: 10.1371/journal.pone.0011822.e11822 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Hu G, Siu SO, Li S, et al. Metabolism of calycosin, an isoflavone from astragali radix, in zebrafish larvae. Xenobiotica. 2012;42(3):294–303. doi: 10.3109/00498254.2011.617015. [DOI] [PubMed] [Google Scholar]
- 19.Siew HL, Yi LW, Vega VB, et al. Conservation of gene expression signatures between zebrafish and human liver tumors and tumor progression. Nature Biotechnology. 2006;24(1):73–75. doi: 10.1038/nbt1169. [DOI] [PubMed] [Google Scholar]
- 20.Rawls JF, Mahowald MA, Ley RE, Gordon JI. Reciprocal gut microbiota transplants from zebrafish and mice to germ-free recipients reveal host habitat selection. Cell. 2006;127(2):423–433. doi: 10.1016/j.cell.2006.08.043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Bauer JH, Goupil S, Garber GB, Helfand SL. An accelerated assay for the identification of lifespan-extending interventions in Drosophila melanogaster. Proceedings of the National Academy of Sciences of the United States of America. 2004;101(35):12980–12985. doi: 10.1073/pnas.0403493101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Watanabe K, Nishimura Y, Oka T, et al. In vivo imaging of zebrafish retinal cells using fluorescent coumarin derivatives. BMC Neuroscience. 2010;11, article 116 doi: 10.1186/1471-2202-11-116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Jeong JY, Kwon HB, Ahn JC, et al. Functional and developmental analysis of the blood-brain barrier in zebrafish. Brain Research Bulletin. 2008;75(5):619–628. doi: 10.1016/j.brainresbull.2007.10.043. [DOI] [PubMed] [Google Scholar]
- 24.Zhang J, Piontek J, Wolburg H, et al. Establishment of a neuroepithelial barrier by Claudin5a is essential for zebrafish brain ventricular lumen expansion. Proceedings of the National Academy of Sciences of the United States of America. 2010;107(4):1425–1430. doi: 10.1073/pnas.0911996107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Woodgate A, MacGibbon G, Walton M, Dragunow M. The toxicity of 6-hydroxydopamine on PC12 and P19 cells. Molecular Brain Research. 1999;69(1):84–92. doi: 10.1016/s0169-328x(99)00103-5. [DOI] [PubMed] [Google Scholar]
- 26.Singh S, Das T, Ravindran A, et al. Involvement of nitric oxide in neurodegeneration: a study on the experimental models of Parkinson’s disease. Redox Report. 2005;10(2):103–109. doi: 10.1179/135100005X38842. [DOI] [PubMed] [Google Scholar]
- 27.Lin YC, Uang HW, Lin RJ, Chen IJ, Lo YC. Neuroprotective effects of glyceryl nonivamide against microglia-like cells and 6-hydroxydopamine-induced neurotoxicity in SH-SY5Y human dopaminergic neuroblastoma cells. Journal of Pharmacology and Experimental Therapeutics. 2007;323(3):877–887. doi: 10.1124/jpet.107.125955. [DOI] [PubMed] [Google Scholar]
- 28.Zhang ZJ, Cheang LCV, Wang MW, Lee SMY. Quercetin exerts a neuroprotective effect through inhibition of the iNOS/NO system and pro-inflammation gene expression in PC12 cells and in zebrafish. International Journal of Molecular Medicine. 2011;27(2):195–203. doi: 10.3892/ijmm.2010.571. [DOI] [PubMed] [Google Scholar]
- 29.Panula P, Sallinen V, Sundvik M, et al. Modulatory neurotransmitter systems and behavior: towards zebrafish models of neurodegenerative diseases. Zebrafish. 2006;3(2):235–247. doi: 10.1089/zeb.2006.3.235. [DOI] [PubMed] [Google Scholar]
- 30.Rihel J, Prober DA, Arvanites A, et al. Zebrafish behavioral profiling links drugs to biological targets and rest/wake regulation. Science. 2010;327(5963):348–351. doi: 10.1126/science.1183090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Kokel D, Bryan J, Laggner C, et al. Rapid behavior-based identification of neuroactive small molecules in the zebrafish. Nature Chemical Biology. 2010;6(3):231–237. doi: 10.1038/nchembio.307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Wolman MA, Jain RA, Liss L, et al. Chemical modulation of memory formation in larval zebrafish. Proceedings of the National Academy of Sciences the United States of America. 2011;108(37):15468–15473. doi: 10.1073/pnas.1107156108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Chatrchyan S, Khachatryan V, Sirunyan AM, et al. Search for neutral minimal supersymmetric standard model Higgs bosons decaying to tau pairs in pp collisions at radicals = 7 TeV. Physical Review Letters. 2011;106(23):15 pages. doi: 10.1103/PhysRevLett.106.231801.231801 [DOI] [PubMed] [Google Scholar]
- 34.Shah RR. Pharmacogenetic aspects of drug-induced torsade de pointes: potential tool for improving clinical drug development and prescribing. Drug Safety. 2004;27(3):145–172. doi: 10.2165/00002018-200427030-00001. [DOI] [PubMed] [Google Scholar]
- 35.Roden DM. Drug-induced prolongation of the QT interval. The New England Journal of Medicine. 2004;350(10):1013–1022. doi: 10.1056/NEJMra032426. [DOI] [PubMed] [Google Scholar]
- 36.FDA. ICH Guidance for Industry: S7B Nonclinical Evaluation of the Potential for Delayed Ventricular Repolarization (QT Interval Prolongation) by Human Pharmaceuticals. U.S. Department of Health and Human Services, 2005. [PubMed]
- 37.Langheinrich U, Vacun G, Wagner T. Zebrafish embryos express an orthologue of HERG and are sensitive toward a range of QT-prolonging drugs inducing severe arrhythmia. Toxicology and Applied Pharmacology. 2003;193(3):370–382. doi: 10.1016/j.taap.2003.07.012. [DOI] [PubMed] [Google Scholar]
- 38.FDA. ICH Guidance for Industry: S7A Safety Pharmacology Studies for Human Pharmaceutical. U.S. Department of Health and Human Services, 2001.
- 39.Tropepe V, Sive HL. Can zebrafish be used as a model to study the neurodevelopmental causes of autism? Genes, Brain and Behavior. 2003;2(5):268–281. doi: 10.1034/j.1601-183x.2003.00038.x. [DOI] [PubMed] [Google Scholar]
- 40.Portavella M, Torres B, Salas C. Avoidance response in goldfish: emotional and temporal involvement of medial and lateral telencephalic pallium. Journal of Neuroscience. 2004;24(9):2335–2342. doi: 10.1523/JNEUROSCI.4930-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Rink E, Wullimann MF. Connections of the ventral telencephalon (subpallium) in the zebrafish (Danio rerio) Brain Research. 2004;1011(2):206–220. doi: 10.1016/j.brainres.2004.03.027. [DOI] [PubMed] [Google Scholar]
- 42.Wullimann MF, Mueller T. Teleostean and mammalian forebrains contrasted: evidence from genes to behavior. Journal of Comparative Neurology. 2004;475(2):143–162. doi: 10.1002/cne.20183. [DOI] [PubMed] [Google Scholar]
- 43.Cahill GM. Clock mechanisms in zebrafish. Cell and Tissue Research. 2002;309(1):27–34. doi: 10.1007/s00441-002-0570-7. [DOI] [PubMed] [Google Scholar]
- 44.Guo S. Linking genes to brain, behavior and neurological diseases: what can we learn from zebrafish? Genes, Brain and Behavior. 2004;3(2):63–74. doi: 10.1046/j.1601-183x.2003.00053.x. [DOI] [PubMed] [Google Scholar]
- 45.Pirmohamed M, James S, Meakin S, et al. Adverse drug reactions as cause of admission to hospital: prospective analysis of 18 820 patients. British Medical Journal. 2004;329(7456):15–19. doi: 10.1136/bmj.329.7456.15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Wallace KN, Akhter S, Smith EM, Lorent K, Pack M. Intestinal growth and differentiation in zebrafish. Mechanisms of Development. 2005;122(2):157–173. doi: 10.1016/j.mod.2004.10.009. [DOI] [PubMed] [Google Scholar]
- 47.Rich A, Leddon SA, Hess SL, et al. Kit-like immunoreactivity in the zebrafish gastrointestinal tract reveals putative ICC. Developmental Dynamics. 2007;236(3):903–911. doi: 10.1002/dvdy.21086. [DOI] [PubMed] [Google Scholar]
- 48.Holmberg A, Schwerte T, Pelster B, Holmgren S. Ontogeny of the gut motility control system in zebrafish Danio rerio embryos and larvae. Journal of Experimental Biology. 2004;207(23):4085–4094. doi: 10.1242/jeb.01260. [DOI] [PubMed] [Google Scholar]
- 49.Wallace KN, Pack M. Unique and conserved aspects of gut development in zebrafish. Developmental Biology. 2003;255(1):12–29. doi: 10.1016/s0012-1606(02)00034-9. [DOI] [PubMed] [Google Scholar]
- 50.Guo L, Guthrie H. Automated electrophysiology in the preclinical evaluation of drugs for potential QT prolongation. Journal of Pharmacological and Toxicological Methods. 2005;52(1):123–135. doi: 10.1016/j.vascn.2005.04.002. [DOI] [PubMed] [Google Scholar]
- 51.Lam IK, Alex D, Wang YH, et al. In vitro and in vivo structure and activity relationship analysis of polymethoxylated flavonoids: identifying sinensetin as a novel antiangiogenesis agent. Molecular Nutrition and Food Research. 2012;56(6):945–956. doi: 10.1002/mnfr.201100680. [DOI] [PubMed] [Google Scholar]
- 52.Alex D, Leong EC, Zhang ZJ, et al. Resveratrol derivative, trans-3,5,4′-trimethoxystilbene, exerts antiangiogenic and vascular-disrupting effects in zebrafish through the downregulation of VEGFR2 and cell-cycle modulation. Journal of Cellular Biochemistry. 2010;109(2):339–346. doi: 10.1002/jcb.22405. [DOI] [PubMed] [Google Scholar]
- 53.Notas G, Nifli AP, Kampa M, Vercauteren J, Kouroumalis E, Castanas E. Resveratrol exerts its antiproliferative effect on HepG2 hepatocellular carcinoma cells, by inducing cell cycle arrest, and NOS activation. Biochimica et Biophysica Acta. 2006;1760(11):1657–1666. doi: 10.1016/j.bbagen.2006.09.010. [DOI] [PubMed] [Google Scholar]
- 54.Lam KH, Alex D, Lam IK, et al. Nobiletin, a polymethoxylated flavonoid from citrus, shows anti-angiogenic activity in a zebrafish in vivo model and HUVEC in vitro model. Journal of Cellular Biochemistry. 2011;112(11):3313–3321. doi: 10.1002/jcb.23257. [DOI] [PubMed] [Google Scholar]
- 55.Houle D, Govindaraju DR, Omholt S. Phenomics: the next challenge. Nature Reviews Genetics. 2010;11(12):855–866. doi: 10.1038/nrg2897. [DOI] [PubMed] [Google Scholar]
- 56.Kausch U, Alberti M, Haindl S, Budczies J, Hock B. Biomarkers for exposure to estrogenic compounds: gene expression analysis in zebrafish (Danio rerio) Environmental Toxicology. 2008;23(1):15–24. doi: 10.1002/tox.20306. [DOI] [PubMed] [Google Scholar]
- 57.Li S, Lou S, Lei BU, et al. Transcriptional profiling of angiogenesis activities of calycosin in zebrafish. Molecular BioSystems. 2011;7(11):3112–3121. doi: 10.1039/c1mb05206c. [DOI] [PubMed] [Google Scholar]
- 58.Siu SO, Lam MPY, Lau E, Kong RPW, Lee SMY, Chu IK. Fully automatable two-dimensional reversed-phase capillary liquid chromatography with online tandem mass spectrometry for shotgun proteomics. Proteomics. 2011;11(11):2308–2319. doi: 10.1002/pmic.201100110. [DOI] [PubMed] [Google Scholar]
- 59.Wang Z, Gerstein M, Snyder M. RNA-Seq: A revolutionary tool for transcriptomics. Nature Reviews Genetics. 2009;10(1):57–63. doi: 10.1038/nrg2484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Wit MD, Keil D, van der Ven K, Vandamme S, Witters E, Coen WD. An integrated transcriptomic and proteomic approach characterizing estrogenic and metabolic effects of 17 α-ethinylestradiol in zebrafish (Danio rerio) General and Comparative Endocrinology. 2010;167(2):190–201. doi: 10.1016/j.ygcen.2010.03.003. [DOI] [PubMed] [Google Scholar]
- 61.Barnham KJ, Haeffner F, Ciccotosto GD, et al. Tyrosine gated electron transfer is key to the toxic mechanism of Alzheimer’s disease β-amyloid. The FASEB Journal. 2004;18(12):1427–1429. doi: 10.1096/fj.04-1890fje. [DOI] [PubMed] [Google Scholar]
- 62.Flinn L, Bretaud S, Lo C, Ingham PW, Bandmann O. Zebrafish as a new animal model for movement disorders. Journal of Neurochemistry. 2008;106(5):1991–1997. doi: 10.1111/j.1471-4159.2008.05463.x. [DOI] [PubMed] [Google Scholar]
- 63.Holzschuh J, Ryu S, Aberger F, Driever W. Dopamine transporter expression distinguishes dopaminergic neurons from other catecholaminergic neurons in the developing zebrafish embryo. Mechanisms of Development. 2001;101(1-2):237–243. doi: 10.1016/s0925-4773(01)00287-8. [DOI] [PubMed] [Google Scholar]
- 64.Anichtchik OV, Kaslin J, Peitsaro N, Scheinin M, Panula P. Neurochemical and behavioural changes in zebrafish Danio rerio after systemic administration of 6-hydroxydopamine and 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine. Journal of Neurochemistry. 2004;88(2):443–453. doi: 10.1111/j.1471-4159.2004.02190.x. [DOI] [PubMed] [Google Scholar]
- 65.McKinley ET, Baranowski TC, Blavo DO, Cato C, Doan TN, Rubinstein AL. Neuroprotection of MPTP-induced toxicity in zebrafish dopaminergic neurons. Molecular Brain Research. 2005;141(2):128–137. doi: 10.1016/j.molbrainres.2005.08.014. [DOI] [PubMed] [Google Scholar]
- 66.Sallinen V, Torkko V, Sundvik M, et al. MPTP and MPP+ target specific aminergic cell populations in larval zebrafish. Journal of Neurochemistry. 2009;108(3):719–731. doi: 10.1111/j.1471-4159.2008.05793.x. [DOI] [PubMed] [Google Scholar]
- 67.Wen L, Wei W, Gu W, et al. Visualization of monoaminergic neurons and neurotoxicity of MPTP in live transgenic zebrafish. Developmental Biology. 2008;314(1):84–92. doi: 10.1016/j.ydbio.2007.11.012. [DOI] [PubMed] [Google Scholar]
- 68.Flinn L, Mortiboys H, Volkmann K, Kster RW, Ingham PW, Bandmann O. Complex I deficiency and dopaminergic neuronal cell loss in parkin-deficient zebrafish (Danio rerio) Brain. 2009;132(6):1613–1623. doi: 10.1093/brain/awp108. [DOI] [PubMed] [Google Scholar]
- 69.Sheng D, Qu D, Kwok KHH, et al. Deletion of the WD40 domain of LRRK2 in zebrafish causes parkinsonism-like loss of neurons and locomotive defect. PLoS Genetics. 2010;6(4) doi: 10.1371/journal.pgen.1000914. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Parng C, Ton C, Lin YX, Roy NM, McGrath P. A zebrafish assay for identifying neuroprotectants in vivo . Neurotoxicology and Teratology. 2006;28(4):509–516. doi: 10.1016/j.ntt.2006.04.003. [DOI] [PubMed] [Google Scholar]
- 71.Rink E, Wullimann MF. The teleostean (zebrafish) dopaminergic system ascending to the subpallium (striatum) is located in the basal diencephalon (posterior tuberculum) Brain Research. 2001;889(1-2):316–330. doi: 10.1016/s0006-8993(00)03174-7. [DOI] [PubMed] [Google Scholar]
- 72.Blum D, Torch S, Lambeng N, et al. Molecular pathways involved in the neurotoxicity of 6-OHDA, dopamine and MPTP: contribution to the apoptotic theory in Parkinson’s disease. Progress in Neurobiology. 2001;65(2):135–172. doi: 10.1016/s0301-0082(01)00003-x. [DOI] [PubMed] [Google Scholar]
- 73.Sharma R, McMillan CR, Tenn CC, Niles LP. Physiological neuroprotection by melatonin in a 6-hydroxydopamine model of Parkinson’s disease. Brain Research. 2006;1068(1):230–236. doi: 10.1016/j.brainres.2005.10.084. [DOI] [PubMed] [Google Scholar]
- 74.Cicchetti F, Brownell AL, Williams K, Chen YI, Livni E, Isacson O. Neuroinflammation of the nigrostriatal pathway during progressive 6-OHDA dopamine degeneration in rats monitored by immunohistochemistry and PET imaging. European Journal of Neuroscience. 2002;15(6):991–998. doi: 10.1046/j.1460-9568.2002.01938.x. [DOI] [PubMed] [Google Scholar]
- 75.Zhang ZJ, Cheang LCV, Wang MW, et al. Ethanolic extract of fructus Alpinia oxyphylla protects against 6-hydroxydopamine-induced damage of PC12 cells in vitro and dopaminergic neurons in zebrafish. Cellular and Molecular Neurobiology. 2011:1–14. doi: 10.1007/s10571-011-9731-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Hunter RL, Dragicevic N, Seifert K, et al. Inflammation induces mitochondrial dysfunction and dopaminergic neurodegeneration in the nigrostriatal system. Journal of Neurochemistry. 2007;100(5):1375–1386. doi: 10.1111/j.1471-4159.2006.04327.x. [DOI] [PubMed] [Google Scholar]
- 77.Esposito E, Di Matteo V, Benigno A, Pierucci M, Crescimanno G, Di Giovanni G. Non-steroidal anti-inflammatory drugs in Parkinson’s disease. Experimental Neurology. 2007;205(2):295–312. doi: 10.1016/j.expneurol.2007.02.008. [DOI] [PubMed] [Google Scholar]
- 78.Baraban SC. Emerging epilepsy models: insights from mice, flies, worms and fish. Current Opinion in Neurology. 2007;20(2):164–168. doi: 10.1097/WCO.0b013e328042bae0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Dugladze T, Vida I, Tort AB, et al. Impaired hippocampal rhythmogenesis in a mouse model of mesial temporal lobe epilepsy. Proceedings of the National Academy of Sciences of the United States of America. 2007;104(44):17530–17535. doi: 10.1073/pnas.0708301104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Houser CR, Zhang NH, Peng ZC. Alterations in the distribution of GABAA receptorsin epilepsy. Epilepsia. 2010;51(5, article 47) [Google Scholar]
- 81.Penschuck S, Lüscher B, Fritschy JM, Crestani F. Activation of the GABA(A)-receptor δ-subunit gene promoter following pentylenetetrazole-induced seizures in transgenic mice. Molecular Brain Research. 1997;51(1-2):212–219. doi: 10.1016/s0169-328x(97)00243-x. [DOI] [PubMed] [Google Scholar]
- 82.Prinz AA. Understanding epilepsy through network modeling. Proceedings of the National Academy of Sciences of the United States of America. 2008;105(16):5953–5954. doi: 10.1073/pnas.0802299105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Löscher W. Animal models of epilepsy for the development of antiepileptogenic and disease-modifying drugs. A comparison of the pharmacology of kindling and post-status epilepticus models of temporal lobe epilepsy. Epilepsy Research. 2002;50(1-2):105–123. doi: 10.1016/s0920-1211(02)00073-6. [DOI] [PubMed] [Google Scholar]
- 84.Morimoto K, Fahnestock M, Racine RJ. Kindling and status epilepticus models of epilepsy: rewiring the brain. Progress in Neurobiology. 2004;73(1):1–60. doi: 10.1016/j.pneurobio.2004.03.009. [DOI] [PubMed] [Google Scholar]
- 85.Baraban SC, Taylor MR, Castro PA, Baier H. Pentylenetetrazole induced changes in zebrafish behavior, neural activity and c-fos expression. Neuroscience. 2005;131(3):759–768. doi: 10.1016/j.neuroscience.2004.11.031. [DOI] [PubMed] [Google Scholar]
- 86.Lukasiuk K, Kaczmarek L. AP-1 and CRE DNA binding activities in rat brain following pentylenetetrazole induced seizures. Brain Research. 1994;643(1-2):227–233. doi: 10.1016/0006-8993(94)90029-9. [DOI] [PubMed] [Google Scholar]
- 87.Wang P, Wang WP, Zhang S, Wang HX, Lou Y, Fan YH. Impaired spatial learning related with decreased expression of calcium/calmodulin-dependent protein kinase IIα and cAMP-response element binding protein in the pentylenetetrazol-kindled rats. Brain Research. 2008;1238:108–117. doi: 10.1016/j.brainres.2008.07.103. [DOI] [PubMed] [Google Scholar]
- 88.Valente T, Domínguez MI, Bellmann A, Journot L, Ferrer I, Auladell C. Zac1 is up-regulated in neural cells of the limbic system of mouse brain following seizures that provoke strong cell activation. Neuroscience. 2004;128(2):323–336. doi: 10.1016/j.neuroscience.2004.06.056. [DOI] [PubMed] [Google Scholar]
- 89.Chen WF, Chang H, Huang LT, et al. Alterations in long-term seizure susceptibility and the complex of PSD-95 with NMDA receptor from animals previously exposed to perinatal hypoxia. Epilepsia. 2006;47(2):288–296. doi: 10.1111/j.1528-1167.2006.00420.x. [DOI] [PubMed] [Google Scholar]
- 90.Tsaur ML, Sheng M, Lowenstein DH, Jan YN, Jan LY. Differential expression of K+ channel mRNAs in the rat brain and down-regulation in the hippocampus following seizures. Neuron. 1992;8(6):1055–1067. doi: 10.1016/0896-6273(92)90127-y. [DOI] [PubMed] [Google Scholar]
- 91.Nehlig A, Rudolf G, Leroy C, Rigoulot MA, Simpson IA, Vannucci SJ. Pentylenetetrazol-induced status epilepticus up-regulates the expression of glucose transporter mRNAs but not proteins in the immature rat brain. Brain Research. 2006;1082(1):32–42. doi: 10.1016/j.brainres.2006.01.078. [DOI] [PubMed] [Google Scholar]
- 92.Siebel AM, Rico EP, Capiotti KM, et al. In vitro effects of antiepileptic drugs on acetylcholinesterase and ectonucleotidase activities in zebrafish (Danio rerio) brain. Toxicology In Vitro. 2010;24(4):1279–1284. doi: 10.1016/j.tiv.2010.03.018. [DOI] [PubMed] [Google Scholar]
- 93.Sehnert AJ, Huq A, Weinstein BM, Walker C, Fishman M, Stainier DYR. Cardiac troponin T is essential in sarcomere assembly and cardiac contractility. Nature Genetics. 2002;31(1):106–110. doi: 10.1038/ng875. [DOI] [PubMed] [Google Scholar]
- 94.Basson CT, Bachinsky DR, Lin RC, et al. Mutations in human TBX5 cause limb and cardiac malformation in Holt-Oram syndrome. Nature Genetics. 1997;15(1):30–35. doi: 10.1038/ng0197-30. [DOI] [PubMed] [Google Scholar]
- 95.Garrity DM, Childs S, Fishman MC. The heartstrings mutation in zebrafish causes heart/fin Tbx5 deficiency syndrome. Development. 2002;129(19):4635–4645. doi: 10.1242/dev.129.19.4635. [DOI] [PubMed] [Google Scholar]
- 96.Goto T, Takase H, Toriyama T, et al. Circulating concentrations of cardiac proteins indicate the severity of congestive heart failure. Heart. 2003;89(11):1303–1307. doi: 10.1136/heart.89.11.1303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Setsuta K, Seino Y, Ogawa T, Arao M, Miyatake Y, Takano T. Use of cytosolic and myofibril markers in the detection of ongoing myocardial damage in patients with chronic heart failure. American Journal of Medicine. 2002;113(9):717–722. doi: 10.1016/s0002-9343(02)01394-3. [DOI] [PubMed] [Google Scholar]
- 98.Pemberton CJ, Raudsepp SD, Yandle TG, Cameron VA, Richards AM. Plasma cardiotrophin-1 is elevated in human hypertension and stimulated by ventricular stretch. Cardiovascular Research. 2005;68(1):109–117. doi: 10.1016/j.cardiores.2005.05.014. [DOI] [PubMed] [Google Scholar]
- 99.Stewart DJ, Cernacek P, Costello KB, Rouleau JL. Elevated endothelin-1 in heart failure and loss of normal response to postural change. Circulation. 1992;85(2):510–517. doi: 10.1161/01.cir.85.2.510. [DOI] [PubMed] [Google Scholar]
- 100.Hansen MS, Stanton EB, Gawad Y, et al. Relation of circulating cardiac myosin light chain 1 isoform in stable severe congestive heart failure to survival and treatment with flosequinan. American Journal of Cardiology. 2002;90(9):969–973. doi: 10.1016/s0002-9149(02)02663-2. [DOI] [PubMed] [Google Scholar]
- 101.Burns CG, Milan DJ, Grande EJ, Rottbauer W, MacRae CA, Fishman MC. High-throughput assay for small molecules that modulate zebrafish embryonic heart rate. Nature Chemical Biology. 2005;1(5):263–264. doi: 10.1038/nchembio732. [DOI] [PubMed] [Google Scholar]
- 102.Kappers MHW, van Esch JHM, Sleijfer S, Danser AJ, van den Meiracker AH. Cardiovascular and renal toxicity during angiogenesis inhibition: clinical and mechanistic aspects. Journal of Hypertension. 2009;27(12):2297–2309. doi: 10.1097/HJH.0b013e3283309b59. [DOI] [PubMed] [Google Scholar]
- 103.Nicholls SJ, Brandrup-Wognsen G, Palmer M, Barter PJ. Meta-analysis of comparative efficacy of increasing dose of Atorvastatin versus Rosuvastatin versus Simvastatin on lowering levels of atherogenic lipids (from VOYAGER) American Journal of Cardiology. 2010;105(1):69–76. doi: 10.1016/j.amjcard.2009.08.651. [DOI] [PubMed] [Google Scholar]
- 104.Yrjänheikki J, Koistinaho J, Kettunen M, et al. Long-term protective effect of atorvastatin in permanent focal cerebral ischemia. Brain Research. 2005;1052(2):174–179. doi: 10.1016/j.brainres.2005.06.004. [DOI] [PubMed] [Google Scholar]
- 105.Lampl Y, Lorberboym M, Gilad R, et al. Early outcome of acute ischemic stroke in hyperlipidemic patients under atorvastatin versus simvastatin. Clinical Neuropharmacology. 2010;33(3):129–134. doi: 10.1097/WNF.0b013e3181d47863. [DOI] [PubMed] [Google Scholar]
- 106.Huisa BN, Stemer AB, Zivin JA. Atorvastatin in stroke: a review of SPARCL and subgroup analysis. Vascular Health and Risk Management. 2010;6(1):229–236. doi: 10.2147/vhrm.s6795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Gjini E, Hekking LH, Küchler A, et al. Zebrafish Tie-2 shares a redundant role with Tie-1 in heart development and regulates vessel integrity. Disease Models and Mechanisms. 2011;4(1):57–66. doi: 10.1242/dmm.005033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Farber SA, Pack M, Ho SY, et al. Genetic analysis of digestive physiology using fluorescent phospholipid reporters. Science. 2001;292(5520):1385–1388. doi: 10.1126/science.1060418. [DOI] [PubMed] [Google Scholar]
- 109.Hama K, Provost E, Baranowski TC, et al. In vivo imaging of zebrafish digestive organ function using multiple quenched fluorescent reporters. American Journal of Physiology. 2009;296(2):G445–G453. doi: 10.1152/ajpgi.90513.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Stoletov K, Fang L, Choi SH, et al. Vascular lipid accumulation, lipoprotein oxidation, and macrophage lipid uptake in hypercholesterolemic zebrafish. Circulation Research. 2009;104(8):952–960. doi: 10.1161/CIRCRESAHA.108.189803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Fang L, Harkewicz R, Hartvigsen K, et al. Oxidized cholesteryl esters and phospholipids in zebrafish larvae fed a high cholesterol diet: macrophage binding and activation. The Journal of Biological Chemistry. 2010;285(42):32343–32351. doi: 10.1074/jbc.M110.137257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Jin S, Hong JH, Jung SH, Cho KH. Turmeric and laurel aqueous extracts exhibit in vitro anti-atherosclerotic activity and in vivo hypolipidemic effects in a zebrafish model. Journal of Medicinal Food. 2011;14(3):247–256. doi: 10.1089/jmf.2009.1389. [DOI] [PubMed] [Google Scholar]
- 113.Rosenberg GA, Mun-Bryce S, Wesley M, Kornfeld M. Collagenase-induced intracerebral hemorrhage in rats. Stroke. 1990;21(5):801–807. doi: 10.1161/01.str.21.5.801. [DOI] [PubMed] [Google Scholar]
- 114.Bullock R, Mendelow AD, Teasdale GM, Graham DI. Intracranial haemorrhage induced at arterial pressure in the rat. Part 1: description of technique, ICP changes and neuropathological findings. Neurological Research. 1984;6(4):184–188. doi: 10.1080/01616412.1984.11739687. [DOI] [PubMed] [Google Scholar]
- 115.Wilson KM, McCaw RB, Leo L, et al. Prothrombotic effects of hyperhomocysteinemia and hypercholesterolemia in ApoE-deficient mice. Arteriosclerosis, Thrombosis, and Vascular Biology. 2007;27(1):233–240. doi: 10.1161/01.ATV.0000251607.96118.af. [DOI] [PubMed] [Google Scholar]
- 116.Plump AS, Smith JD, Hayek T, et al. Severe hypercholesterolemia and atherosclerosis in apolipoprotein E-deficient mice created by homologous recombination in ES cells. Cell. 1992;71(2):343–353. doi: 10.1016/0092-8674(92)90362-g. [DOI] [PubMed] [Google Scholar]
- 117.Franco CA, Liebner S, Gerhardt H. Vascular morphogenesis: a Wnt for every vessel? Current Opinion in Genetics and Development. 2009;19(5):476–483. doi: 10.1016/j.gde.2009.09.004. [DOI] [PubMed] [Google Scholar]
- 118.Lawson ND, Weinstein BM. In vivo imaging of embryonic vascular development using transgenic zebrafish. Developmental Biology. 2002;248(2):307–318. doi: 10.1006/dbio.2002.0711. [DOI] [PubMed] [Google Scholar]
- 119.Langheinrich U. Zebrafish: a new model on the pharmaceutical catwalk. BioEssays. 2003;25(9):904–912. doi: 10.1002/bies.10326. [DOI] [PubMed] [Google Scholar]
- 120.Raghunath M, Sy Wong Y, Farooq M, Ge R. Pharmacologically induced angiogenesis in transgenic zebrafish. Biochemical and Biophysical Research Communications. 2009;378(4):766–771. doi: 10.1016/j.bbrc.2008.11.127. [DOI] [PubMed] [Google Scholar]
- 121.Nicoli S, De Sena G, Presta M. Fibroblast growth factor 2-induced angiogenesis in zebrafish: the zebrafish yolk membrane (ZFYM) angiogenesis assay. Journal of Cellular and Molecular Medicine. 2009;13(8):2061–2068. doi: 10.1111/j.1582-4934.2008.00432.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Alex D, Lam IK, Lin Z, Lee SMY. Indirubin shows anti-angiogenic activity in an in vivo zebrafish model and an in vitro HUVEC model. Journal of Ethnopharmacology. 2010;131(2):242–247. doi: 10.1016/j.jep.2010.05.016. [DOI] [PubMed] [Google Scholar]
- 123.Lu XL, Luo D, Yao XL, et al. Dl-3n-butylphthalide promotes angiogenesis via the extracellular signal-regulated kinase 1/2 and phosphatidylinositol 3-kinase/Akt-endothelial nitric oxide synthase signaling pathways. Journal of Cardiovascular Pharmacology. 2012;59(4):352–362. doi: 10.1097/FJC.0b013e3182443e74. [DOI] [PubMed] [Google Scholar]
- 124.Lam HW, Lin HC, Lao SC, et al. The angiogenic effects of Angelica sinensis extract on HUVEC in vitro and zebrafish in vivo . Journal of Cellular Biochemistry. 2008;103(1):195–211. doi: 10.1002/jcb.21403. [DOI] [PubMed] [Google Scholar]
- 125.Hong SJ, Wan JB, Zhang Y, et al. Angiogenic effect of saponin extract from Panax notoginseng on HUVECs in vitro and zebrafish in vivo . Phytotherapy Research. 2009;23(5):677–686. doi: 10.1002/ptr.2705. [DOI] [PubMed] [Google Scholar]
- 126.Jackson JR, Seed MP, Kircher CH, Willoughby DA, Winkler JD. The codependence of angiogenesis and chronic inflammation. The FASEB Journal. 1997;11(6):457–465. [PubMed] [Google Scholar]
- 127.Carmeliet P, Jain RK. Angiogenesis in cancer and other diseases. Nature. 2000;407(6801):249–257. doi: 10.1038/35025220. [DOI] [PubMed] [Google Scholar]
- 128.Balkwill F, Mantovani A. Inflammation and cancer: back to Virchow? The Lancet. 2001;357(9255):539–545. doi: 10.1016/S0140-6736(00)04046-0. [DOI] [PubMed] [Google Scholar]
- 129.Hau M, Kneitz C, Tony HP, Keberle M, Jahns R, Jenett M. High resolution ultrasound detects a decrease in pannus vascularisation of small finger joints in patients with rheumatoid arthritis receiving treatment with soluble tumour necrosis factor α receptor (etanercept) Annals of the Rheumatic Diseases. 2002;61(1):55–58. doi: 10.1136/ard.61.1.55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Mor F, Quintana FJ, Cohen IR. Angiogenesis-inflammation cross-talk: vascular endothelial growth factor is secreted by activated T cells and induces Th1 polarization. Journal of Immunology. 2004;172(7):4618–4623. doi: 10.4049/jimmunol.172.7.4618. [DOI] [PubMed] [Google Scholar]
- 131.Bonizzi G, Karin M. The two NF-κB activation pathways and their role in innate and adaptive immunity. Trends in Immunology. 2004;25(6):280–288. doi: 10.1016/j.it.2004.03.008. [DOI] [PubMed] [Google Scholar]
- 132.Meteoglu I, Erdogdu IH, Meydan N, Erkus M, Barutca S. NF-κB expression correlates with apoptosis and angiogenesis in clear cell renal cell carcinoma tissues. Journal of Experimental and Clinical Cancer Research. 2008;27(1, article 53) doi: 10.1186/1756-9966-27-53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Leung WK, To KF, Go MY, et al. Cyclooxygenase-2 upregulates vascular endothelial growth factor expression and angiogenesis in human gastric carcinoma. International Journal of Oncology. 2003;23(5):1317–1322. [PubMed] [Google Scholar]
- 134.Kim BH, Kim CI, Chang HS, et al. Cyclooxygenase-2 overexpression in chronic inflammation associated with benign prostatic hyperplasia: Is it related to apoptosis and angiogenesis of prostate cancer? Korean Journal of Urology. 2011;52(4):253–259. doi: 10.4111/kju.2011.52.4.253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Kannappan R, Gupta SC, Kim JH, Aggarwal BB. Tocotrienols fight cancer by targeting multiple cell signaling pathways. Genes and Nutrition. 2012;7(1):43–52. doi: 10.1007/s12263-011-0220-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Hseu YC, Wu CR, Chang HW, et al. Inhibitory effects of Physalis angulata on tumor metastasis and angiogenesis. Journal of Ethnopharmacology. 2011;135(3):762–771. doi: 10.1016/j.jep.2011.04.016. [DOI] [PubMed] [Google Scholar]
- 137.Kunikata T, Tatefuji T, Aga H, Iwaki K, Ikeda M, Kurimoto M. Indirubin inhibits inflammatory reactions in delayed-type hypersensitivity. European Journal of Pharmacology. 2000;410(1):93–100. doi: 10.1016/s0014-2999(00)00879-7. [DOI] [PubMed] [Google Scholar]
- 138.Singh UP, Singh NP, Singh B, et al. Resveratrol (Trans-3,5,4′-trihydroxystilbene) induces silent mating type information regulation-1 and down-regulates nuclear transcription factor-κB activation to abrogate dextran sulfate sodium-induced colitis. Journal of Pharmacology and Experimental Therapeutics. 2010;332(3):829–839. doi: 10.1124/jpet.109.160838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Zhao H, Deneau J, Che GO, et al. Angelica sinensis isolate SBD.4: composition, gene expression profiling, mechanism of action and effect on wounds, in rats and humans. European Journal of Dermatology. 2012;22(1):58–67. doi: 10.1684/ejd.2011.1599. [DOI] [PubMed] [Google Scholar]
- 140.Hu G, Mahady GB, Li S, et al. Polysaccharides from astragali radix restore chemical-induced blood vessel loss in zebrafish. Vascular Cell. 2012;4(1, article 2) doi: 10.1186/2045-824X-4-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Tse HY, Hui MN, Li L, et al. Angiogenic efficacy of simplified 2-herb formula (NF3) in zebrafish embryos in vivo and rat aortic ring in vitro . Journal of Ethnopharmacology. 2012;139(2):447–453. doi: 10.1016/j.jep.2011.11.031. [DOI] [PubMed] [Google Scholar]
- 142.Dunnett SB, Björklund A. Prospects for new restorative and neuroprotective treatments in Parkinson’s disease. Nature. 1999;399(6738, supplement):A32–A39. doi: 10.1038/399a032. [DOI] [PubMed] [Google Scholar]
- 143.Barnham KJ, Masters CL, Bush AI. Neurodegenerative diseases and oxidatives stress. Nature Reviews Drug Discovery. 2004;3(3):205–214. doi: 10.1038/nrd1330. [DOI] [PubMed] [Google Scholar]
- 144.Weinreb O, Amit T, Bar-Am O, Youdim MBH. Rasagiline: a novel anti-Parkinsonian monoamine oxidase-B inhibitor with neuroprotective activity. Progress in Neurobiology. 2010;92(3):330–344. doi: 10.1016/j.pneurobio.2010.06.008. [DOI] [PubMed] [Google Scholar]
- 145.Wang M, Zhang Z, Cheang LC, Lin Z, Lee SM. Eriocaulon buergerianum extract protects PC12 cells and neurons in zebrafish against 6-hydroxydopamine-induced damage. Chinese Medicine. 2011;6, article 16 doi: 10.1186/1749-8546-6-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Koo BS, Lee WC, Chang YC, Kim CH. Protective effects of alpinae oxyphyllae fructus (Alpinia oxyphylla MIQ) water-extracts on neurons from ischemic damage and neuronal cell toxicity. Phytotherapy Research. 2004;18(2):142–148. doi: 10.1002/ptr.1382. [DOI] [PubMed] [Google Scholar]
- 147.Yu X, An L, Wang Y, Zhao H, Gao C. Neuroprotective effect of Alpinia oxyphylla Miq. fruits against glutamate-induced apoptosis in cortical neurons. Toxicology Letters. 2003;144(2):205–212. doi: 10.1016/s0378-4274(03)00219-4. [DOI] [PubMed] [Google Scholar]
- 148.Ho JC, Chen CM. Flavonoids from the aquatic plant Eriocaulon buergerianum . Phytochemistry. 2002;61(4):405–408. doi: 10.1016/s0031-9422(02)00218-2. [DOI] [PubMed] [Google Scholar]
- 149.Yang LX, Huang KX, Li HB, et al. Design, synthesis, and examination of neuron protective properties of alkenylated and amidated dehydro-silybin derivatives. Journal of Medicinal Chemistry. 2009;52(23):7732–7752. doi: 10.1021/jm900735p. [DOI] [PubMed] [Google Scholar]
- 150.Lo YC, Shih YT, Chen IJ, Wu YC. San-Huang-Xie-Xin-Tang protects against activated microglia- and 6-OHDA-induced toxicity in neuronal SH-SY5Y cells. Evidence-Based Complementary and Alternative Medicine. 2011;2011:11 pages. doi: 10.1093/ecam/nep025.429384 [DOI] [PMC free article] [PubMed] [Google Scholar]