

Abstract
Theory predicts that mothers should adjust offspring sex ratios when the expected fitness gains or rearing costs differ between sons and daughters. Recent empirical work has linked biased offspring sex ratios to environmental quality via changes in relative maternal condition. It is unclear, however, whether females can manipulate offspring sex ratios in response to environmental quality alone (i.e. independent of maternal condition). We used a balanced within-female experimental design (i.e. females bred on both low- and high-quality diets) to show that female parrot finches (Erythrura trichroa) manipulate primary offspring sex ratios to the quality of the rearing environment, and not to their own body condition and health. Individual females produced an unbiased sex ratio on high-quality diets, but over-produced sons in poor dietary conditions, even though they maintained similar condition between diet treatments. Despite the lack of sexual size dimorphism, such sex ratio adjustment is in line with predictions from sex allocation theory because nutritionally stressed foster sons were healthier, grew faster and were more likely to survive than daughters. These findings suggest that mothers may adaptively adjust offspring sex ratios to optimally match their offspring to the expected quality of the rearing environment.
Keywords: sex allocation, nutrition quality, maternal diet, maternal condition, sex-specific costs
1. Introduction
Evolutionary theory predicts that when the fitness benefits of producing sons and daughters differ, females should adaptively maximize their fitness by adjusting offspring sex ratios to meet the prevailing social and environmental conditions [1,2]. This should be expected, for example, when constraints on current breeding conditions, such as partner quality and resource availability, impose differential fitness or rearing costs to producing sons and daughters. Yet, although relatively straightforward in theory, compelling empirical evidence for adaptive sex allocation in vertebrates remains elusive and often contradictory [3,4]. For example, despite a few striking examples of strategic sex-ratio adjustment [5–7], a large number of studies have found no evidence [8,9], and others suggest that biases in offspring sex ratios are not consistent within species or populations [10,11] or across years [12,13]. However, relatively few studies have attempted to investigate the sex-specific fitness costs and benefits to females (reviewed in [3,4,14]), and thus the net benefits to be gained from strategic sex ratio adjustment are often unclear or unknown.
To date, perhaps the best-supported studies have focused on maternal adjustment in relation to environmental factors, such as maternal condition and current nutritional quality. In this context, when resources are plentiful or high-quality, females are expected to produce more offspring of the sex that will gain the greatest benefit from increased investment or nutritional quality [1,15], and thus yield the highest return of fitness. Conversely, when conditions are harsh, females should produce offspring of the sex that suffers least from reduced environmental or parental condition [16,17]. For example, several studies on birds [16,18,19], reptiles [20] and mammals [21] (including humans [22,23]) have documented patterns of offspring sex-ratio variation consistent with these predictions. However, it is unclear whether mothers in different quality environments are adjusting offspring sex ratios in response to their own condition during egg production, or to their perception of the expected quality of the rearing environment. For example, because nutrition often directly affects individual body condition, most studies manipulating diet quality or quantity have, by association, also found a relationship between offspring sex ratios and maternal condition [18–20,24]. Furthermore, a number of studies manipulating maternal condition (without changing diet) have reported biased offspring sex ratios [16,25], suggesting that relative maternal condition and physiology at egg production (birds) or conception (mammals) may provide a mechanistic route for strategic sex allocation in vertebrates [21,26]. Consequently, it is unclear whether variation in nutritional quality alone, independent of maternal condition, can affect strategic sex allocation.
Here, we use a balanced within-female experimental design to test the relative effects of maternal condition and diet on the primary sex ratio of the blue-faced parrot finch (Erythrura trichroa), a small (ca 12–14 g) Australian estrildid finch where adult body condition appears largely insensitive to changes in nutritional quality. Although there is no discernable sexual dimorphism in adult body size in these socially monogamous finches [27], it is unknown whether there are sex-specific differences in life-history or fitness responses to resource quality. For example, previous studies have shown that sex-specific vulnerabilities during embryo and offspring development are not only related to sexual size dimorphism (i.e. larger sex has greater nutritional requirements [28,29]), but may also be influenced or exacerbated by sex-specific phenotypes (e.g. costs of testosterone in males [30,31]), clutch size [32] and genetic factors (e.g. heterogameity [1]), especially in poor conditions. Although males are traditionally thought to suffer greater mortality during development [28,33], studies of monomorphic species have also shown female-biased embryo and/or juvenile mortality under poor rearing conditions [34,35]. Therefore, to determine whether sons or daughters are the ‘weaker’ sex and more susceptible to nutritional stress, offspring were cross-fostered within and between different diet treatments (to decouple maternal investment from sex-specific responses to resource limitation), and their relative growth, health and survival monitored through juvenile development.
2. Material and methods
(a). Experimental design
Experiments were run from July 2010 to June 2011 using naive (virgin) wild-type birds from a large captive-bred colony (Martinsville, New South Wales, Australia). From hatching, all experimental birds had been reared and maintained on the same standard diet (containing approx. 16% protein), which consisted of a custom-mixed seed mix (red millet, white millet, Japanese millet, Siberian rye and canary seed), mineralized grit, and a daily supplement of freshly sprouted seeds (red millet, white millet, Japanese millet). Males and females were housed in single-sex groups prior to breeding.
In total, 56 females were randomly assigned to either a high-quality or low-quality diet, containing approximately 20 per cent or 8 per cent protein, respectively [36]. The low-quality diet consisted of a dry seed mix of three different seeds in equal proportions (red millet, Siberian rye and Japanese millet) and mineralized grit. In contrast, the high-quality diet included a dry seed mix of nine different seeds (red millet, white millet, Japanese millet, Siberian rye, canary seed, silk sorghum, niger and black lettuce), mineralized grit and a daily supplement (10 g) of freshly sprouted seeds (red millet, white millet and Japanese millet) and a specially designed softfood (egg, wheatgerm extract and multi-mineral powder). Females were kept on these diets in single-sex groups for a period of 12 weeks before being paired randomly with males. Pairs were housed in individual breeding cages, equipped with nest-boxes. For the 12 weeks prior to pairing, all males were caged in a separate room and fed the standard laboratory diet. Once paired, males were then fed on the same diet as their mate (i.e. in the same cage), and these diets were maintained during chick rearing. Females initiated egg-laying within 9.1 ± 7.1 days from pairing (8.9 ± 7.4 days on high-quality diets; 9.3 ± 6.7 days on low-quality diets).
Nests were checked daily for eggs, which were weighed (0.01 g). All broods were cross-fostered to isolate the effects of maternal investment at the time of laying from the post-hatching rearing environment [37]. Broods were either swapped between diet treatments (52 broods) or within the same diet treatment (44 broods) 1–2 days after hatching (1.5 ± 0.3 days old). No broods were abandoned. Offspring remained on their foster parent's experimental diet until independence (40 days), when offspring were removed from their carers and breeding pairs were separated and transferred to single-sex groups. Birds were given a six-week rest period (during which they all received the standard laboratory diet) before diet treatments were reversed. Thereafter, females that were previously on the high-quality diet received the low-quality diet, and vice versa. Males remained on the standard laboratory diet. After 12 weeks, all birds were re-paired with the same mate as in the first round, and the experiment was repeated.
(b). Sexing
Offspring sex was determined using a small blood sample (from chicks) or tissue (stored in alcohol) from unhatched embryos and dead chicks (less than two days) by PCR amplification of multiplexed sex-linked markers 3007F, 2987F and 3112R [38]. In 12 eggs (2.6% of eggs produced), there were no visible signs of embryonic development. Seven unsexed eggs were laid by females on the low-quality diet and five by females on the high-quality diet.
(c). Adult condition and health
To evaluate changes in condition, females were weighed, measured and blood sampled at pairing (i.e. after 12 weeks on their diet) and on the morning that the first egg was laid (13.2 ± 2.1 weeks on the diet). Males were also weighed at pairing and during egg-laying (at the same time as females). To standardize the procedures and minimize potential time effects, all measurements were taken at the same time of day (06.00–08.00 h). Approximately 150 μl of blood (two capillary tubes) were taken and transferred to heparinized microhaematocrit tubes, which were immediately centrifuged.
Haematocrit, or packed cell volume in relation to the whole volume of blood, reflects the erythrocyte content of blood, and is an increasingly used index of avian condition and physiological performance [39–42]. Haematocrit was calculated as the ratio of packed cell volume of whole blood (%) from the two centrifuged capillaries; within-individual measurements were highly repeatable (r = 0.98, F47,96 = 46.32, p < 0.001). As an index of nutritional condition [36,42,43], whole-blood haemoglobin concentration (g dl−1) was also measured on a 20 µl blood sample using a HemoCue photometer (HemoCue AB, Ängelholm, Sweden).
Cutaneous immune responsiveness was evaluated during egg production by measuring the extent of cutaneous swelling following injection of birds with a non-pathogenic antigen, phytohemagglutinin (PHA-P; Sigma). PHA is a lectin that stimulates innate and adaptive immune responses, including those giving rise to a measurable inflammatory response [44]. Each bird's right and left wing webs were injected with either PHA or sterile phosphate-buffered saline (PBS), respectively [45,46] and the consequent swellings measured with pressure-sensitive digital callipers (0.01 mm) 24 h later. The PHA response was calculated as the difference in swellings between the left wing web (control for the effect of injection on web wing thickness: PBS only) and the right wing web (0.2 mg PHA-P dissolved in 0.04 ml PBS).
(d). Offspring growth, condition and survival
Chicks were weighed every second day until 16 days old using an electronic balance (0.01 g) and their relative growth rates calculated (details in [46]). As an index of nutritional condition, whole-blood haemoglobin concentration (g dl−1) was measured on a 20 µl blood sample of 12-day-old nestlings using a HemoCue photometer [42]. To assess nestling immune function, PHA responses were measured in 12-day-old nestlings (as described for adults earlier). Offspring survival was measured at hatching (proportion of hatched young), and from hatching to independence (proportion surviving to 40 days).
(e). Statistical analysis
Within-individual changes in body weight, haemoglobin concentration and immune function, as well as clutch sizes, were analysed using a repeated-measures ANOVA with a structured blocking function (i.e. repeated measures for within-individual comparisons), with diet treatment and treatment order as fixed effects. Egg and chick measures were analysed using repeated-measures generalized linear models (GLM), with eggs or chicks within the nest as a repeated measure and breeding pair as a random factor. For the egg mass model, diet (of biological parents), treatment order and offspring sex were included as explanatory effects. For models of offspring health and development, pre-hatching environment (biological parents' diet), post-hatching rearing diet (foster parents diet), treatment order and offspring sex were included as potential explanatory effects.
Offspring sex ratio (number of males/total number of sexed eggs), as well as survival at hatching (number hatching/total number of eggs) and independence (number weaned/total number fostered after hatching) were analysed at the brood level by fitting generalized linear mixed models (GLMM). The error distribution was binomial with a logit link. Because there were no differences in clutch sizes between diet treatments or within females (see §3), GLMMs were not weighted by brood size. Pre-hatching environment and treatment order were included as fixed effects to test for diet-specific differences in offspring sex ratio. Pre-hatching parental diet, egg mass and offspring sex were included as explanatory terms in the hatchability model, while post-hatching rearing diet was also included as a fixed effect in tests of juvenile survival. Breeding pair and nest identity were included and retained as random repeated subjects, but did not constitute a significant random component in any model.
3. Results
(a). Male condition
There were no differences in pre-experimental body weight between males allocated to low- and high-quality diets (F1,55 = 0.25, p = 0.83). Between pairing and egg production (i.e. after an average of 9.1 ± 7.1 days on the experimental diets), there were also no within-male changes in body mass (F1,47 = 0.87, p = 0.52), or differences in male weights between the diet treatments (F1,47 = 0.13, p = 0.86).
(b). Female condition and immune function
There were no differences in pre-experimental body weight between females assigned to the two different diet treatments (F1,55 = 0.12, p = 0.90). Surprisingly, there were also no within-female differences in body weight between low- and high-quality diets at either pairing (F1,55 = 0.28, p = 0.77) or during egg-laying (F1,47 = 1.02, p = 0.32; figure 1a). Diet treatment also had no effect on maternal haemoglobin concentrations or haematocrit levels at initial pairing (haemoglobin: F1,47 = 0.05, p = 0.81; haematocrit: F1,47 = 0.19, p = 0.65) or during egg production (haemoglobin: F1,47 = 0.33, p = 0.74; haematocrit: F1,47 = 1.68, p = 0.19; figure 1b). Similarly, immune function was unaffected by diet treatments, with females showing similar PHA responses during egg-laying on both low- (2.36 ± 0.20) and high-quality diets (2.43 ± 0.22; F1,47 = 2.60, p = 0.11). There was no effect of treatment order on any condition or immune measures (F1,47 < 1.15, p > 0.21).
Figure 1.
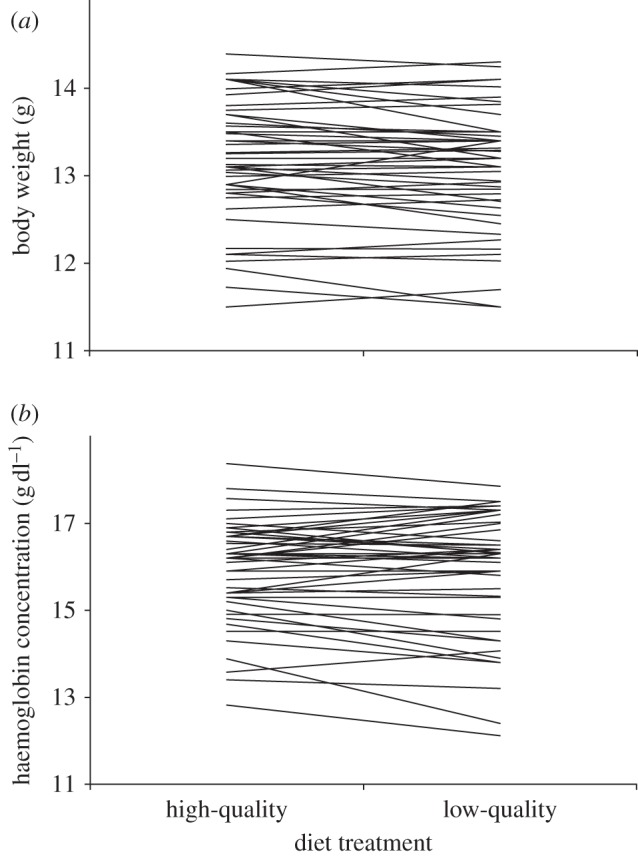
Within-female changes in (a) body weight and (b) haemoglobin concentrations at egg production for individual females breeding on high- and low-quality diets. Treatment order (i.e. whether first breeding on high- or low-quality diets) was randomized in the experiments.
(c). Clutch and egg size
Forty-eight females laid clutches on both the high- and low-quality diets. Two females did not lay a clutch on either diet, five laid only on the high-quality diet and one laid only on the low-quality diet. There were no within-female differences in clutch sizes between the two diet treatments (F1,47 = 0.73, p = 0.47) and no effect of treatment order on clutch size (F1,47 = 1.39, p = 0.24). In a paired, repeated measures GLM, there were no diet-related differences in egg size (F1,452 = 2.18, p = 0.14). There were also no sex-specific differences in the mass of male and female eggs in either diet treatment (F1,452 = 1.56, p = 0.21) and no effect of treatment order on egg mass (F1,95 = 2.18, p = 0.14).
(d). Sex ratios
When fed low-quality diets, females produced significantly male-biased primary sex ratios (72.9%; χ2 = 14.46, p < 0.01; n = 48 families, 229 offspring), whereas when fed the high-quality diet, females produced an unbiased sex ratio (48.6%; χ2 = 1.09, p = 0.28; n = 48 families, 224 offspring; figure 2). There were no treatment order effects in primary sex ratio adjustment (χ2 = 0.07, p = 0.79).
Figure 2.
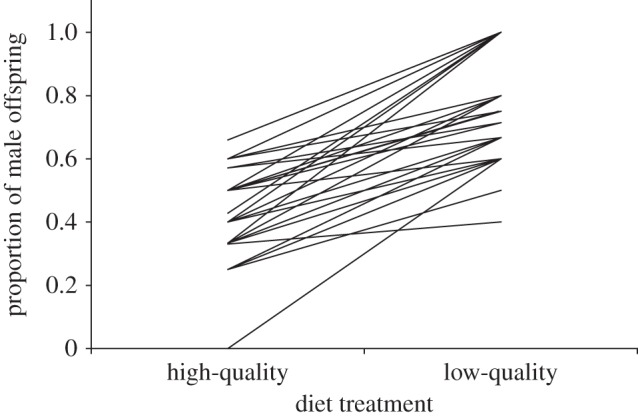
Primary sex ratio (proportion of sons) of clutches produced by individual females breeding on both high- and low-quality diets (lines connect the data points between the two clutches from individual females).
(e). Offspring development, health and survival
Sixteen of 453 (3.5%) embryos died before hatching. Embryo mortality was unaffected by diet (χ2 = 0.33, p = 0.55), offspring sex (χ2 = 0.92, p = 0.33) or egg mass (χ2 = 0.58, p = 0.44). There were also no diet-related differences in offspring weight at hatching (F1,95 = 2.06, p = 0.15), or after experimentally cross-fostering offspring within and between high- and low-quality diets (F1,95 = 0.73, p = 0.39). However, foster offspring reared on low-quality diets grew significantly more slowly than offspring reared on high-quality diets (F1,95 = 13.97, p < 0.001; figure 3a) and this effect was independent of their pre-hatching environment (i.e. biological parents' diet: F1,95 = 0.05, p = 0.81). In particular, although there was no sexual dimorphism in hatchling weight (F1,95 = 1.89, p = 0.17), sons reared on low-quality diets grew faster than daughters (sex × diet: F1,95 = 37.27, p < 0.001; sex: F1,95 = 17.05, p < 0.001). As a consequence, foster daughters reared on low-quality diets fledged later (26.40 ± 3.35 days) than sons (20.85 ± 2.45 days) (sex × diet: F1,95 = 47.61, p < 0.001; sex: F1,95 = 35.77, p < 0.001; diet: F1,95 = 38.19, p < 0.001).
Figure 3.
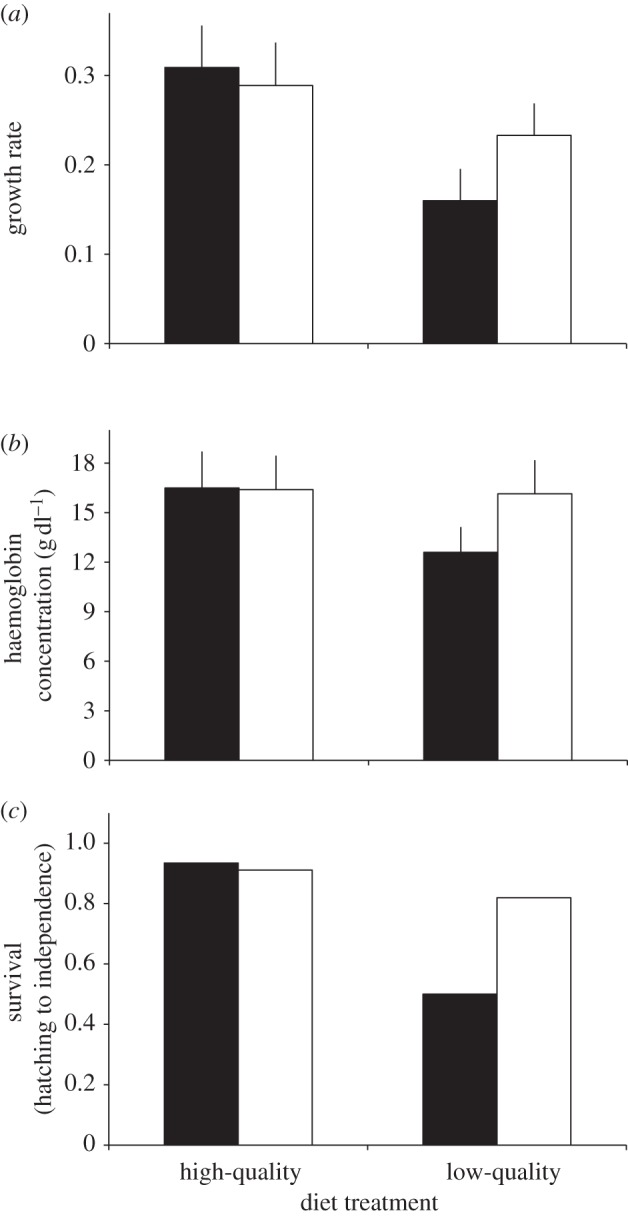
Foster daughters reared on low-quality diets had (a) lower growth rates, (b) lower haemoglobin concentrations and (c) higher mortality than sons, and offspring (both daughters (black bars) and sons (white bars)) reared on high-quality diets. Values are presented as predicted means (+s.d.) from GLMM.
By using haemoglobin concentration as an index of nutritional condition, foster offspring reared on high-quality diets were in better condition than foster offspring reared on low-quality diets (F1,95 = 10.63, p < 0.001). Specifically, foster sons had higher haemoglobin concentrations than daughters on low-quality diets (sex × diet: F1,95 = 9.35, p < 0.001; sex: F1,95 = 8.81, p < 0.001; figure 3b). Similarly, foster sons reared on low-quality diets had higher PHA responses than daughters (sex × diet: F1,95 = 16.27, p < 0.001; sex: F1,95 = 16.41, p < 0.001; diet: F1,95 = 16.81, p < 0.001), but there were no sex-specific immune functions differences for sons and daughters reared on high-quality diets.
Seventy-eight of 440 (17.7%) offspring died between hatching and independence. More fostered offspring reared on low-quality diets (24.9%) died before reaching independence than on high-quality diets (9.5%) (χ2 = 9.91, p = 0.002). Furthermore, mortality was more severe for daughters (than sons) reared on low-quality diets (sex × diet: χ2 = 23.47, p < 0.001; sex: χ2 = 13.41, p < 0.001; figure 3c). There was no effect of egg mass (χ2 = 1.44, p = 0.19) or pre-hatching environment (χ2 = 0.08, p = 0.81) on post-hatching mortality.
4. Discussion
Manipulation of diet quality prior to breeding had no detectable effect on the body condition, health and immune function of females. However, despite this, the brood sex ratio at egg-laying differed between the two diets. Females laid significantly more male eggs in poor dietary conditions than they did in favourable dietary conditions. Such differential maternal investment is likely to be adaptive because sons appear to be less vulnerable to nutritional stress than daughters. Although there was no sexual dimorphism in egg and hatching size, foster sons reared on poor-quality diets grew faster and were healthier (i.e. higher haemoglobin levels and cutaneous immune responses) than daughters. Both haemoglobin [42,43] and PHA [39,47,48] responses in nestlings have been found to predict health and survival probability in a number of bird species, and are thought to reflect nutritional deficiencies and increased disease susceptibility of poorer-quality offspring. Consistent with this, we found that malnourished sons fledged earlier than daughters and were more likely to survive until parental independence. Indeed, daughters appear highly vulnerable to nutritional stress during development, with more than 51.5 per cent of daughters reared on low-quality diets dying (compared with 7.3% of sons). Since we experimentally controlled for post-hatching maternal effects (i.e. cross-fostering within and between treatments), these fitness effects are likely to be indicative of sex-specific responses to nutritional stress in their early rearing environment. Because of the lack of sexual size dimorphism in parrot finches [27], these sex-specific fitness effects could not have been predicted a priori. Therefore, these results highlight the importance of experimentally testing the effect of environmental conditions on the relative development of sons and daughters.
Previous studies on birds and mammals have reported sex-specific offspring mortality during early development, with sons (rather than daughters) typically found to suffer higher mortality in poor rearing conditions [28,33]. In general, the larger body sizes of males (e.g. higher growth and metabolic rates [28]) and their greater physiological costs (e.g. androgens that trade-off with immune function; [30,31,49]) have been invoked to explain their increased susceptibility to poor environmental or nutritional conditions. Furthermore, work on mammals also suggests that the greater mortality of sons might be due to their heterogameity [1]. The heterogametic sex (XY males in mammals; ZW females in birds) is thought to always be at a disadvantage, because they are susceptible to the expression of sex-linked deleterious recessive alleles. This hypothesis might explain why daughters in this study, and in the related and monomorphic zebra finch [34,35,50], are more vulnerable than sons to malnutrition during early development. Theoretically, heterogameity would predispose daughters to greater mortality and reduced performance during development [1,2]. However, it remains untested whether heterogameity is the main or sole driver of daughter-biased mortality in these studies. Furthermore, it seems unlikely that heterogameity provides a general explanation across birds [32]. Male-biased mortality has been reported in a number of avian species [33,51] and other factors, such as clutch size and sibling rivalry [32], have also been invoked to explain sex-specific vulnerability. Further work is needed to identify the role of both genetic and phenotypic factors in determining sex-specific embryo and juvenile development.
In contrast to their offspring, adult parrot finches appear to be largely insensitive to dietary changes in their environment. Despite a nutritionally compromised diet (which had substantially reduced protein content), female parrot finches on low-quality diets maintained comparable body weight, haemoglobin concentrations, haematocrit and PHA responses compared with when they were fed high-quality diets. Because relative ‘condition’ is a complex measure, the indices used here may not have accurately or completely measured changes in condition to nutritional stress. However, these morphological and physiological measures have typically been associated with condition, health and survival in a number of bird species [39,41,44], including the closely related Gouldian finch (Erythrura gouldiae) [36,42], and even in their own offspring (this study). Interestingly, in contrast to other Erythrura finches (i.e. parrot and Gouldian finches) that have highly specialized diets [27], blue-faced parrot finches are generalist foragers, often switching food resources to use the most prevalent seeds available in their natural environments [52]. Perhaps as a consequence, they have become increasingly popular as pet birds in captivity, and are renowned as ‘good doers’ that thrive on diets considered substandard for most estrildid finches [27]. In particular, although blue-faced parrot finches will readily feed on small insects, they do not require such high protein sources for maintenance or breeding [27], which may help explain why adults in this study maintained comparable body condition and immune function on the low-quality protein-deprived diets.
Since males were also fed the same experimental diet as females (once they were paired to females), one possibility is that paternal condition may influence sex allocation [53]. For example, fathers in inferior condition may reduce provisioning and care to their offspring, potentially providing direct benefits to females for adaptive sex allocation. However, female parrot finches initiated egg-laying very rapidly (average of nine days from pairing), and during this short time male body weights did not change on either diet treatment, suggesting that male condition is unlikely to directly influence female allocation decisions. Another possibility is that nutritional quality could affect differential fertilization, perhaps through differences in sperm quality and/or viability. In lizards, for example, male-biased sex ratios may arise because sperm with male sex-determining loci have increased viability in the female reproductive tract [54]. However, although the quality and viability of avian sperm can change rapidly in response to environmental conditions [55], and can affect offspring viability [56,57], it is unclear how males could directly influence sex determination in birds; females are the heterogametic sex (producing Z- and W-bearing ova) and therefore they control offspring sex prior to ovulation [26]. Nevertheless, there is growing evidence for the role of paternal effects in adaptive sex allocation (e.g. lizards [53,54]), and further studies in birds are required to rigorously test these possibilities.
Variation in diet quality has been linked to variation in offspring sex ratios in a number of species [15,18,20]. In these studies, however, females in poorer environments were also in poorer body condition. In general, females in relatively poorer condition are expected to produce proportionately more offspring of the sex that either benefits most or suffers least from their compromised condition, whereas the reverse is expected for high-condition females [1]. In this study, however, there were no diet-related changes in maternal body condition, but females nevertheless overproduced sons on low-quality diets. This suggests that females are not adjusting offspring sex ratios in relation to their own relative condition per se, but rather that females are adjusting sex ratios in response to the expected rearing environment for their offspring, presumably because sons are less costly than daughters to rear in poor conditions. Altering offspring sex ratios in response to the quality of the local environment is likely to be advantageous, as it should allow mothers to best match the phenotype of their offspring to the prevailing conditions, and thus maximize their fitness. Future studies are needed to determine the relative long-term costs and benefits of such strategic sex allocation to both mothers and offspring.
Acknowledgements
Thanks to Erik Wapstra and an anonymous reviewer for very helpful comments on the manuscript. This work was supported by an Australian Research Council Discovery Grant (Australian Post-doctoral Fellowship; to S.R.P.) and a Macquarie University Vice-Chancellor's Innovation Research Fellowship (S.R.P.). The Animal Ethics Committee at Macquarie University approved this research.
References
- 1.Trivers R. L., Willard D. E. 1973. Natural selection of parental ability to vary the sex ratio of offspring. Science 179, 90–91 10.1126/science.179.4068.90 (doi:10.1126/science.179.4068.90) [DOI] [PubMed] [Google Scholar]
- 2.Charnov E. L. 1982. The theory of sex allocation. Princeton, NJ: Princeton University Press [Google Scholar]
- 3.West S. A. 2009. Sex allocation. Princeton, NJ: Princeton University Press [Google Scholar]
- 4.Cockburn A., Legge S., Double M. C. 2002. Sex ratios in birds and mammals: can the hypotheses be disentangled? In Sex ratios: concepts and research methods (ed. Hardy I. C. W.), pp. 266–286 Cambridge, UK: Cambridge University Press [Google Scholar]
- 5.Pryke S. R., Griffith S. C. 2009. Genetic incompatibility drives sex allocation and maternal investment in a polymorphic finch. Science 323, 1605–1607 10.1126/science.1168928 (doi:10.1126/science.1168928) [DOI] [PubMed] [Google Scholar]
- 6.Heinsohn R., Legge S., Barry S. 1997. Extreme bias in sex allocation in Eclectus parrots. Proc. R. Soc. Lond. B 264, 1325–1329 10.1098/rspb.1997.0183 (doi:10.1098/rspb.1997.0183) [DOI] [Google Scholar]
- 7.Komdeur J., Daan S., Tinbergen J., Mateman C. 1997. Extreme adaptive modification in sex ratio of the Seychelles warbler's eggs. Nature 385, 522–525 10.1038/385522a0 (doi:10.1038/385522a0) [DOI] [Google Scholar]
- 8.Parker T. H. 2005. No evidence for adaptive differential sex allocation in red junglefowl. Auk 122, 1161–1168 10.1642/0004-8038(2005)122[1161:NEFADS]2.0.CO;2 (doi:10.1642/0004-8038(2005)122[1161:NEFADS]2.0.CO;2) [DOI] [Google Scholar]
- 9.Radford A. N., Blakey J. K. 2000. Is variation in brood sex ratios adaptive in the great tit (Parus major)? Behav. Ecol. 11, 294–298 10.1093/beheco/11.3.294 (doi:10.1093/beheco/11.3.294) [DOI] [Google Scholar]
- 10.Rosivall B., Török J., Haselquist D., Bensch S. 2004. Brood sex ratio adjustment in collared flycatchers (Ficedula albicollis): results differ between populations. Behav. Ecol. Sociobiol. 56, 346–351 10.1007/s00265-004-0796-3 (doi:10.1007/s00265-004-0796-3) [DOI] [Google Scholar]
- 11.Dreiss A., Moyen R. M., White F., Moller A. P., Danchin E. 2006. Sex ratio and male sexual characters in a population of blue tits, Parus caeruleus. Behav. Ecol. 17, 13–19 10.1093/beheco/ari089 (doi:10.1093/beheco/ari089) [DOI] [Google Scholar]
- 12.Korsten P., Lessells C. M., Mateman A. C., van der Velde M., Komdeur J. 2006. Primary sex ratio adjustment to experimentally reduced male UV attractiveness in blue tits. Behav. Ecol. 17, 539–546 10.1093/beheco/arj061 (doi:10.1093/beheco/arj061) [DOI] [Google Scholar]
- 13.Radford A., Blakey J. 2000. Intensity of nest defence is related to offspring sex ratio in the great tit Parus major. Proc. R. Soc. Lond. B 267, 535–538 10.1098/rspb.2000.1033 (doi:10.1098/rspb.2000.1033) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.West S. A., Sheldon B. C. 2002. Constraints in the evolution of sex ratio adjustment. Science 295, 1685–1688 10.1126/science.1069043 (doi:10.1126/science.1069043) [DOI] [PubMed] [Google Scholar]
- 15.Appleby B. M., Petty S. J., Blakey J. K., Rainey P., Macdonald D. W. 1997. Does variation of sex ratio enhance reproductive success of offspring in tawny owls (Strix aluco). Proc. R. Soc. Lond. B 264, 1111–1116 10.1098/rspb.1997.0153 (doi:10.1098/rspb.1997.0153) [DOI] [Google Scholar]
- 16.Nager R. G., Monaghan P., Griffiths R., Houston D. C., Dawson R. 1999. Experimental demonstration that offspring sex ratio varies with maternal condition. Proc. Natl Acad. Sci. USA 96, 570–573 10.1073/pnas.96.2.570 (doi:10.1073/pnas.96.2.570) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Cameron E. Z., Linklater W. L. 2002. Sex bias in studies of sex bias: the value of daughters to mothers in poor condition. Anim. Behav. 63, F5–F8 10.1006/anbe.2001.1902 (doi:10.1006/anbe.2001.1902) [DOI] [Google Scholar]
- 18.Rutstein A. N., Slater P. J. B., Graves J. A. 2004. Diet quality and resource allocation in the zebra finch. Biol. Lett. 271, S286–S289 10.1098/rsbl.2003.0154 (doi:10.1098/rsbl.2003.0154) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Rutkowska J., Cichon M. 2002. Maternal investment during egg laying and offspring sex: an experimental study of zebra finches. Anim. Behav. 64, 817–822 10.1006/anbe.2002.1973 (doi:10.1006/anbe.2002.1973) [DOI] [Google Scholar]
- 20.Warner D. A., Lovern M. B., Shine R. 2007. Maternal nutrition affects reproductive output and sex allocation in a lizard with environmental sex determination. Proc. R. Soc. B 274, 883–890 10.1098/rspb.2006.0105 (doi:10.1098/rspb.2006.0105) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Cameron E. Z., Linklater W. L. 2007. Extreme sex ratio variation in relation to change in condition around conception. Biol. Lett. 3, 395–397 10.1098/rsbl.2007.0089 (doi:10.1098/rsbl.2007.0089) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Cameron E. Z., Dalerum F. 2009. A Trivers–Willard effect in contemporary humans: male-biased sex ratios among billionaires. PLoS ONE 4, e4195. 10.1371/journal.pone.0004195 (doi:10.1371/journal.pone.0004195) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Pollet T. V., Fawcett T. W., Buunck A. P., Nettle D. 2009. Sex-ratio biasing towards daughters among lower-ranking co-wives in Rwanda. Biol. Lett. 5, 765–768 10.1098/rsbl.2009.0394 (doi:10.1098/rsbl.2009.0394) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Pike T. W. 2005. Sex ratio manipulation in response to maternal condition in pigeons: evidence for pre-ovulatory follicle selection. Behav. Ecol. Sociobiol. 58, 407–408 10.1007/s00265-005-0931-9 (doi:10.1007/s00265-005-0931-9) [DOI] [Google Scholar]
- 25.Kalmbach E., Nager R. G., Griffiths R., Furness R. W. 2001. Increased reproductive effort results in male-biased offspring sex ratio: an experimental study in a species with reversed sexual size dimorphism. Proc. R. Soc. Lond. B 268, 2175–2179 10.1098/rspb.2001.1793 (doi:10.1098/rspb.2001.1793) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Pike T. W., Petrie M. 2003. Potential mechanisms of avian sex manipulation. Biol. Rev. 78, 553–574 10.1017/S1464793103006146 (doi:10.1017/S1464793103006146) [DOI] [PubMed] [Google Scholar]
- 27.Evans S., Fidler M. 1990. Parrot finches. London, UK: Artillery House [Google Scholar]
- 28.Clutton-Brock T. H., Albon S. D., Guinness F. E. 1985. Parental investment and sex-differences in juvenile mortality in birds and mammals. Nature 313, 131–133 10.1038/313131a0 (doi:10.1038/313131a0) [DOI] [Google Scholar]
- 29.Kalmbach E., Benito M. M. 2007. Sexual size dimorphism and offspring vulnerability in birds. In Sex, size and gender roles: evolutionary studies of sexual size dimorphism (eds Fairburn D. J., Blanckenhorn W. U., Szekely T.), pp. 133–142 Oxford, UK: Oxford University Press [Google Scholar]
- 30.Zuk M. 1990. Reproductive strategies and disease susceptibility: an evolutionary viewpoint. Parasitol. Today 6, 231–233 10.1016/0169-4758(90)90202-F (doi:10.1016/0169-4758(90)90202-F) [DOI] [PubMed] [Google Scholar]
- 31.Müller W., Groothuis T. G. G., Eising C. M., Dijkstra C. 2005. An experimental study on the causes of sex-biased mortality in the black-headed gull—the possible role of testosterone. J. Anim. Ecol. 74, 735–741 10.1111/j.1365-2656.2005.00964.x (doi:10.1111/j.1365-2656.2005.00964.x) [DOI] [Google Scholar]
- 32.Jones K. S., Nakagawa S., Sheldon B. C. 2009. Environmental sensitivity in relation to size and sex in birds: meta-regression analysis. Am. Nat. 174, 122–133 10.1086/599299 (doi:10.1086/599299) [DOI] [PubMed] [Google Scholar]
- 33.Griffiths R. 1992. Sex-biased mortality in the lesser black-backed gull Larus fuscus during the nestling stage. Ibis 134, 237–244 10.1111/j.1474-919X.1992.tb03805.x (doi:10.1111/j.1474-919X.1992.tb03805.x) [DOI] [Google Scholar]
- 34.Bradbury R. B., Blakey J. K. 1998. Diet, maternal condition, and offspring sex ratio in the zebra finch, Poephila guttata. Proc. R. Soc. Lond. B 265, 895–899 10.1098/rspb.1998.0375 (doi:10.1098/rspb.1998.0375) [DOI] [Google Scholar]
- 35.Kilner R. 1998. Primary and secondary sex ratio manipulation by zebra finches. Anim. Behav. 56, 155–164 10.1006/anbe.1998.0775 (doi:10.1006/anbe.1998.0775) [DOI] [PubMed] [Google Scholar]
- 36.Pryke S. R., Rollins L. A., Griffith S. C. 2011. Context-dependent sex allocation: constraints on the expression and evolution of maternal effects. Evolution 65, 2792–2799 10.1111/j.1558-5646.2011.01391.x (doi:10.1111/j.1558-5646.2011.01391.x) [DOI] [PubMed] [Google Scholar]
- 37.Pryke S. R., Griffith S. C. 2009. Postzygotic genetic incompatibility between sympatric color morphs. Evolution 63, 793–798 10.1111/j.1558-5646.2008.00584.x (doi:10.1111/j.1558-5646.2008.00584.x) [DOI] [PubMed] [Google Scholar]
- 38.Fridolfsson A. K., Ellegren H. 1999. A simple and universal method for molecular sexing of non-ratite birds. J. Avian Biol. 30, 116–121 10.2307/3677252 (doi:10.2307/3677252) [DOI] [Google Scholar]
- 39.Saino N., Bolzern A. M., Møller A. P. 1997. Immunocompetence, ornamentation, and viability of male barn swallows (Hirundo rustica). Proc. Natl Acad. Sci. USA 94, 549–552 10.1073/pnas.94.2.549 (doi:10.1073/pnas.94.2.549) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Potti J., Moreno J., Merino S., Frias O., Rodriguez R. 1999. Environmental and genetic variation in the haematocrit of fledgling pied flycatchers Ficedula hypoleuca. Oecologia 120, 1–8 10.1007/s004420050826 (doi:10.1007/s004420050826) [DOI] [PubMed] [Google Scholar]
- 41.Fair J., Whitaker S., Pearson B. 2007. Sources of variation in haematocrit in birds. Ibis 149, 535–552 10.1111/j.1474-919X.2007.00680.x (doi:10.1111/j.1474-919X.2007.00680.x) [DOI] [Google Scholar]
- 42.Pryke S. R., Astheimer L. B., Griffith S. C., Buttemer W. A. 2012. Hormonal basis of life-history trade-offs: effects of diet on condition, behavior and reproduction in genetic finch morphs. Am. Nat. 179, 375–390 10.1086/664078 (doi:10.1086/664078) [DOI] [PubMed] [Google Scholar]
- 43.Bańbura J., Bańbura M., Kaliński A., Skwarska J., Stomczyński R., Wawrzyniak J., Zieliński P. 2007. Habitat and year-to-year variation in haemoglobin concentration in nestling blue tits Cyanistes caeruleus. Comp. Biochem. Physiol. Mol. Integr. Physiol. 148, 572–577 10.1016/j.cbpa.2007.07.008 (doi:10.1016/j.cbpa.2007.07.008) [DOI] [PubMed] [Google Scholar]
- 44.Martin L. B., Han P., Lewittes J., Kuhlman J. R., Klasing K. C., Wikelski M. 2006. Phytohemagglutinin-induced skin swelling in birds: histological support for a classic immunoecological technique. Funct. Ecol. 20, 290–299 10.1111/j.1365-2435.2006.01094.x (doi:10.1111/j.1365-2435.2006.01094.x) [DOI] [Google Scholar]
- 45.Pryke S. R., Astheimer L. B., Buttemer W. A., Griffith S. 2007. Frequency-dependent tradeoffs between competing colour morphs. Biol. Lett. 3, 494–497 10.1098/rsbl.2007.0213 (doi:10.1098/rsbl.2007.0213) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Pryke S. R., Griffith S. C. 2009. Socially mediated trade-offs between aggression and parental effort in competing color morphs. Am. Nat. 174, 455–464 10.1086/605376 (doi:10.1086/605376) [DOI] [PubMed] [Google Scholar]
- 47.Tella J. L., Bortolotti G. R., Forero M. G., Dawson R. D. 2000. Environmental and genetic variation in T-cell-mediated immune response of fledgling American kestrels. Oecologia 123, 453–459 10.1007/s004420000331 (doi:10.1007/s004420000331) [DOI] [PubMed] [Google Scholar]
- 48.Christe P., de Lope F., Gonzalez G., Saino N., Møller A. P. 2001. The influence of environmental conditions on immune responses, morphology and recapture probability of nestling house martins (Delichon urbica). Oecologia 126, 333–338 10.1007/s004420000527 (doi:10.1007/s004420000527) [DOI] [PubMed] [Google Scholar]
- 49.Uller T., Olsson M. 2003. Prenatal exposure to testosterone increases ectoparasite susceptibility in the common lizard (Lacerta vivipara). Proc. R. Soc. Lond. B 270, 1867–1870 10.1098/rspb.2003.2451 (doi:10.1098/rspb.2003.2451) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Martins T. L. F. 2004. Sex-specific growth in zebra finch nestlings: a possible mechanism for sex ratio adjustment. Behav. Ecol. 15, 174–180 10.1093/beheco/arg094 (doi:10.1093/beheco/arg094) [DOI] [Google Scholar]
- 51.Whittingham L. A., Dunn P. O. 2001. Survival of extra-pair and within-pair young in tree swallows. Behav. Ecol. 12, 496–500 10.1093/beheco/12.4.496 (doi:10.1093/beheco/12.4.496) [DOI] [Google Scholar]
- 52.Craig M. D. 2003. An ecological study of the blue-faced parrot-finch (Erythrura trichoa macgillivrayi) near Yungaburra, Australia. Emu 103, 363–368 10.1071/MU02047 (doi:10.1071/MU02047) [DOI] [Google Scholar]
- 53.Cox R. M., Duryea M. C., Najarro M., Calsbeek R. 2011. Paternal condition drives sex-ratio bias in a lizard that lacks parental care. Evolution 65, 220–230 10.1111/j.1558-5646.2010.01111.x (doi:10.1111/j.1558-5646.2010.01111.x) [DOI] [PubMed] [Google Scholar]
- 54.Olsson M., Schwartz T., Uller T., Healey M. 2007. Sons are made from old stores: sperm storage effects on sex ratio in a lizard. Biol. Lett. 3, 491–493 10.1098/rsbl.2007.0196 (doi:10.1098/rsbl.2007.0196) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Immler S., Pryke S. R., Birkhead T. R., Griffith S. C. 2010. Pronounced within-individual plasticity in sperm morphometry across social environments. Evolution 64, 1634–1643 10.1111/j.1558-5646.2009.00924.x (doi:10.1111/j.1558-5646.2009.00924.x) [DOI] [PubMed] [Google Scholar]
- 56.Birkhead T. R., Møller A. P. 1998. Sperm competition in birds: evolutionary causes and consequences. London, UK: Academic Press [Google Scholar]
- 57.Pryke S. R., Rollins L. A., Griffith S. C. 2010. Females use multiple mating and genetically loaded sperm competition to target compatible genes. Science 329, 964–967 10.1126/science.1192407 (doi:10.1126/science.1192407) [DOI] [PubMed] [Google Scholar]