

Abstract
All nonmammalian vertebrates studied can regenerate inner ear mechanosensory receptors (i.e., hair cells) (Corwin and Cotanche, 1988; Lombarte et al., 1993; Baird et al., 1996), but mammals possess only a very limited capacity for regeneration after birth (Roberson and Rubel, 1994). As a result, mammals experience permanent deficiencies in hearing and balance once their inner ear hair cells are lost. The mechanisms of hair cell regeneration are poorly understood. Because the inner ear sensory epithelium is highly conserved in all vertebrates (Fritzsch et al., 2007), we chose to study hair cell regeneration mechanism in adult zebrafish, hoping the results would be transferrable to inducing hair cell regeneration in mammals. We defined the comprehensive network of genes involved in hair cell regeneration in the inner ear of adult zebrafish with the powerful transcriptional profiling technique digital gene expression, which leverages the power of next-generation sequencing (‘t Hoen et al., 2008). We also identified a key pathway, stat3/socs3, and demonstrated its role in promoting hair cell regeneration through stem cell activation, cell division, and differentiation. In addition, transient pharmacological inhibition of stat3 signaling accelerated hair cell regeneration without overproducing cells. Taking other published datasets into account (Sano et al., 1999; Schebesta et al., 2006; Dierssen et al., 2008; Riehle et al., 2008; Zhu et al., 2008; Qin et al., 2009), we propose that the stat3/socs3 pathway is a key response in all tissue regeneration and thus an important therapeutic target for a broad application in tissue repair and injury healing.
Introduction
The sensory epithelium of the inner ear is mainly composed of two types of cells: hair cells and supporting cells (Fritzsch et al., 2006). Inner ear hair cells are the basic mechanosensory receptors for hearing and balance (Vollrath et al., 2007), while supporting cells provide a variety of functions including being the stem cells for replacing hair cells in most vertebrates (Balak et al., 1990; Baird et al., 1996; Jones and Corwin, 1996). All nonmammalian vertebrates studied show the ability to regenerate their inner ear hair cells (Cruz et al., 1987; Corwin and Cotanche, 1988; Lombarte et al., 1993; Baird et al., 1996). However, in mammals, loss of inner ear hair cells caused by acoustic overexposure (McGill and Schuknecht, 1976), otoxic drugs (Lim, 1976) or aging (Soucek et al., 1986) is the major cause of permanent auditory and vestibular deficiencies because mammals lose regenerative ability after birth (Roberson and Rubel, 1994). Because the inner ear sensory epithelium is highly conserved in all vertebrates (Fritzsch et al., 2007), numerous studies have been done in nonmammalian vertebrates to understand the mechanism of hair cell regeneration (Brignull et al., 2009). However, our understanding of the mechanisms involved is still very limited because it is a complex, multistaged process (Stone and Cotanche, 2007).
In this study, we used the powerful profiling technique digital gene expression (DGE) (‘t Hoen et al., 2008; Morrissy et al., 2009) to study the hair cell regeneration in zebrafish at high resolution to get a more comprehensive view of the process. In zebrafish, spontaneous and damage-induced hair cell production has been demonstrated in both the inner ear (Bang et al., 2001; Higgs et al., 2002; Schuck and Smith, 2009) and the neuromasts (Harris et al., 2003), a mechanosensory structure highly similar to the sensory epithelia of the inner ear (Nicolson, 2005) and thus an excellent model for studying hair cell regeneration (Harris et al., 2003; Hernández et al., 2007; Ma et al., 2008; Behra et al., 2009). In addition, zebrafish are commonly used as a genetic/genomic model organism, making zebrafish a valuable system for studying the molecular mechanisms of hair cell regeneration in adult vertebrates in a systematic fashion.
By analyzing the expression profiles from inner ear tissues during regeneration, we identified a key pathway, stat3/socs3, and demonstrated its role in promoting regeneration. Signal transducer and activator of transcription 3 (stat3) is a transcription factor. Its downstream targets include stat3 itself and suppressor of cytokine signaling 3 (socs3) (Leonard and O'Shea, 1998). Socs3 protein antagonizes the activation of Stat3 in the cytosol as a negative feedback (Leonard and O'Shea, 1998). The self-restrictive pathway is known to be involved in the following various biological processes: cell proliferation, cell migration, immune responses, cell survival (Yoshimura, 2009; Yu et al., 2009), as well as regeneration in skin (Sano et al., 1999; Zhu et al., 2008), liver (Dierssen et al., 2008; Riehle et al., 2008), fins (Schebesta et al., 2006), and retinas (Qin et al., 2009). Comparing our data with those in other publications, we propose that the stat3/socs3 pathway is a key response in all tissue regeneration and thus a potential therapeutic target for tissue repair.
Materials and Methods
Animal husbandry.
Zebrafish were maintained under approved animal protocols as previously described (Westerfield, 2000) and in compliance with guidelines for animal care from NIH and the University of Maryland.
Noise exposure of adult zebrafish.
Adult wild-type TAB-5 (Amsterdam et al., 1999) mixed-sex zebrafish (∼1 year old) were exposed to white noise (100–10,000 Hz, 150–170 dB re 1 μPa) for 48 h at 28–29°C according to a protocol modified from Smith et al. (2006). After exposure, the fish were maintained under regular husbandry conditions until killed. The control fish were not exposed to noise.
Tag profiling and tag mapping.
The tag profiling data of inner ear tissues were generated by Illumina. Tags detected only once were discarded. The tag sequences were mapped against transcriptome and genomic sequence databases: Refseq RNAs, UniGene, Ensembl RNAs, Ensembl RNA ab initio, and the zebrafish genome (Zv.8). UniGene IDs were used as the primary index (except for those Refseq RNAs without corresponding UniGene IDs). The expression level of a certain gene (identified by a unique UniGene ID) was the summation of counts of the tags that were “unambiguously mapped” to the gene normalized by the total counts of all tags obtained in the profile in the expression unit called transcripts per million.
Analyses of expression profiling data.
The sample clustering analysis was done using Genesifter software (Geospiza). Candidate genes were identified for further analysis if they showed significant differences in expression level during regeneration compared with the control sample (i.e., ≥1.5-fold increase or ≥2-fold decrease in expression level with a p value <0.01, as determined by χ2 test or Fisher's exact test. Pathway analysis was done with MetaCore software (GeneGo).
Quantitative RT-PCR.
Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and bactin1 were used as reference genes. The relative change in expression level of a gene X in an experimental sample compared with the control sample was calculated as 2 × (ΔCtEXPx − ΔCtCONx), where ΔCtEXPx = (CtEXPref − CtEXPx) and ΔCtCONx = (CtCONref − CtCONx). Ct is the cycle number at which amplification rises above the background threshold. All primers are listed in Table 1.
Table 1.
Sequences of primers and morpholinos
| Experiment | Target gene | Sequence |
|---|---|---|
| qRT-PCR | atoh1a | F: 5′-GCG AAG AAT GCA CGG ATT GAA CCA-3′ |
| R: 5′-TGC AGG GTT TCG TAC TTG GAG AGT-3′ | ||
| bactin1 | F: 5′-GAC CCA GAC ATC AGG GAG TGA TGG-3′ | |
| R: 5′-AGG TGT GAT GCC AGA TCT TCT CCA TG-3′ | ||
| bcl6 | F: 5′-TGT TCT GCT CAA CCT GAA CCG ACT-3′ | |
| R: 5′-TAG AAG AGC CCA CTG CAT GCC ATA-3′ | ||
| dld | F: 5′-TCC AAC CCT TGC TCG AAT GAT GCT-3′ | |
| R: 5′-TCG ATG TTG TCT TCG CAG TGC GTT-3′ | ||
| gapdh | F: 5′-ACT CCA CTC ATG GCC GTT AC-3′ | |
| R: 5′- TGA GCT GAG GCC TTC TCA AT-3′ | ||
| jak1 | F: 5′-ACG AGT GCT TGG GAA TGG CTG TTT-3′ | |
| R: 5′-AGT TGC GTT GCT TAA TGG TGC GGT-3′ | ||
| mmp2 | F: 5′-GCT GGT GTG CAA CCA CTG AAG ATT-3′ | |
| R: 5′-AAG ACA CAG GGT GCT CCA TCT GAA-3′ | ||
| mmp9 | F: 5′-AAA TCG AGA AGC TCG GCC TAC CAA-3′ | |
| R: 5′- TCC TCT GTC AAT CAG CTG AGC CTT-3′ | ||
| socs3a | F: 5′-TAA AGC AGG GAA GAC AAG AGC CGA-3′ | |
| R: 5′-TGG AGA AAC AGT GAG AGA GCT GGT-3′ | ||
| socs3b | F: 5′-CGG ATA ACG CTT TGA AGC TGC CTT-3′ | |
| R: 5′-TAC TAT GCG TTA CCA TGG CGC TCT-3′ | ||
| stat3 | F: 5′-AGT GAA AGC AGC AAA GAG GGA GGA-3′ | |
| R: 5′-TGA GCT GCT GCT TAG TGT ACG GTT-3′ | ||
| In situ hybridization probe synthesis | stat3 | F: 5′-ATT AAC CCT CAC TAA AGG GAC GCA AGT TCA ACA TCC TTG GCA CT-3′ |
| R: 5′-TAA TAC GAC TCA CTA TAG GGA GAT TTG GCT CGG AGA GAG AAA GCA GT-3′ | ||
| socs3a | F: 5′-ATT AAC CCT CAC TAA AGG GAA AGA CTG TGA ACG GAC ACA CGG AT-3′ | |
| R: 5′-TAA TAC GAC TCA CTA TAG GGA GAA GTG TCT GGC ATG AGA AGG CTG AA-3′ | ||
| Morpholino | socs3a | MO1: CCCTGAGCTGCCGGGAAGCAGATCT |
| MO2: CGTGTAATATACAGAGTGTCGAGTC | ||
| stat3 | MO1: GCCATGTTGACCCCTTAATGTGTCG | |
| tp53 | MO1: GCGCCATTGCTTTGCAAGAATTG |
A list of primers for qRT-PCR and in situ hybridization probe synthesis are listed. The bold sequences are T3 [forward (F)] and T7 [reverse (R)] promoters included in the primers for probe synthesis. Both morpholinos targeting socs3a gave rise to similar phenotypes. Results in the article are from embryos injected with MO1. stat3 MO was synthesized based on sequences from Yamashita et al. (2002), which was also used by Sepich et al. (2005) and Kassen et al. (2009) for knocking down stat3 expression in zebrafish embryos. tp53 MO was synthesized based on sequences from Kratz et al. (2006).
Whole-mount in situ hybridization.
Regular whole-mount in situ hybridization (ISH) of zebrafish embryos was done as previously described (Oxtoby and Jowett, 1993). Fluorescent whole-mount in situ hybridization was performed according to the regular in situ hybridization protocol, except that the TSA Cyanine 3 system (PerkinElmer) was used for developing fluorescent signals. All primers for probe synthesis are listed in Table 1.
Morpholino and mature mRNA injection.
The injections were done in fertilized eggs of TAB-5 (Amsterdam et al., 1999) or Tg(cldnb:GFP) (Haas and Gilmour, 2006) lines. Morpholinos (MOs) against stat3, socs3a, and tp53 (Table 1) were injected at a concentration of 500 μm. The mature mRNA of socs3a was injected at a concentration of 37.8 ng/μl.
Induction of lateral line hair cell death in larval zebrafish.
The 5 d postfertilization (dpf)-old larval fish were treated with 10 μm CuSO4 for 2 h at 28.5°C as previously described (Hernández et al., 2007) to induce complete lateral line hair cell loss.
Quantification of hair cell numbers and mitotic events during lateral line hair cell regeneration.
After CuSO4 treatment to induce lateral line hair cell death, 5-dpf-old Tg(pou4f3:GFP) larvae (Xiao et al., 2005) were then raised at 28.5°C in the Holtfreter's buffer containing S3I-201 (400 μm, EMD Biosciences) or DMSO (40 μl/10 ml buffer, Sigma). Lateral line hair cells (GFP-positive cells) in living larvae were quantified at 24, 48, and 72 h post-treatment (hpt). The mitotic events during lateral line hair cell regeneration were quantified by a bromodeoxyuridine (BrdU) incorporation assay (Ma et al., 2008).
Immunohistochemistry.
The primary and secondary antibodies used include the rabbit myosin VI and myosin VIIa antibodies (1:200 dilution, Proteus Biosciences), rabbit anti-stat3pS727 (1:50 dilution, Abcam), mouse anti-BrdU with Alexa Fluor 546 conjugate (1:200 dilution, Invitrogen), goat GFP antibody (FITC conjugated, 1:200 dilution, Abcam), and Alexa Fluor 568 (or 488) goat anti-rabbit IgG (1:1000 dilution, Invitrogen). The inner ear sensory epithelia were dissected as previously described (Liang and Burgess, 2009).
Microscopy and image analysis.
To capture images of the entire saccular epithelia, several overlapping pictures were taken for each epithelium with Zeiss Vision software and tiled with Photoshop 7.0 software (Adobe). The imaging in the living larvae was done using an Axiovert200M and an Apotome Grid Confocal (Zeiss). The antibody staining and fluorescent in situ hybridization results of embryos/larvae were visualized under AxiovertNLO confocal microscope (Zeiss) with Zeiss AIM software. The regular in situ hybridization results were captured using a Leica MZ16F microscope with Leica FireCam software.
Statistical analysis.
χ2 tests (genes with tag count >5) and Fisher's exact tests (genes with tag count ≤5) were used to compare the expression levels of the genes quantified by tag profiling. The calculation was done using R software. Expression data from inner ear samples collected at different time points during regeneration were compared with those from the control sample. One-way ANOVA, post hoc test, and Student's t test were used to compare hair cell numbers. The calculations were done using Excel software (Microsoft).
Results
Inner ear hair cell regeneration in adult zebrafish
We induced inner ear hair cell loss in adult zebrafish by modifying a previously published noise exposure protocol (Smith et al., 2006) (Fig. 1A). Using this setup, we could consistently induce hair cell loss in the anterior-medial region of the saccular macula (Fig. 1B), but not in the utricular or lagenar macula (data not shown), which is consistent with the proposed function of saccule as the major auditory end organ in otophysians (Fay, 1978). We confirmed complete elimination of the hair cells (as opposed to hair cell bundle damage) with phalloidin (staining hair cell bundles) and myosin VI/VIIa antibodies (staining hair cell bodies) (Fig. 1C). When we quantified the hair cells in a 20 × 20 μm area at 40% of the total length of the anterior–posterior axis of the epithelium at 24 h intervals from 0 to 120 h postexposure (hpe), we found that the hair cells regrew rapidly and returned to control levels in 96 h (Fig. 1C,D), consistent with a previous publication (Schuck and Smith, 2009).
Figure 1.
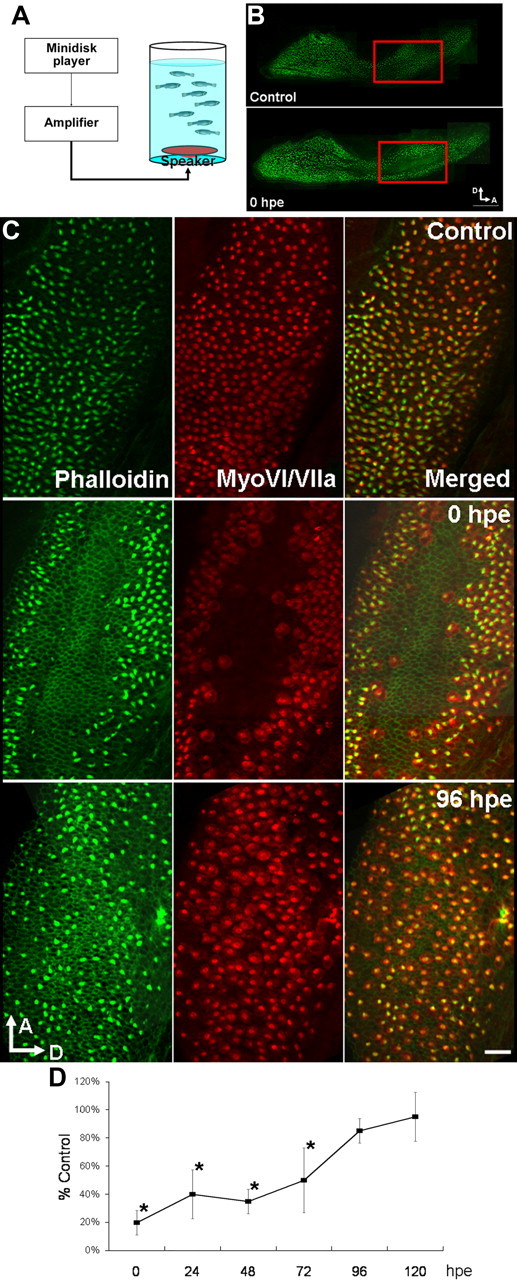
Saccular hair cells were eliminated by noise exposure and subsequently regenerated. A, Hair cell loss in the saccule was consistently induced by the noise exposure apparatus. B, Phalloidin staining of the saccular hair cells; hair cell loss occurred in the anterior-medial area of the saccular sensory epithelium (red boxes; scale bar, 100 μm). C, Closer examination of the damaged area with phalloidin (green channel) and anti-myosin VI/VIIa (red channel) staining confirmed the complete elimination of hair cell bodies (middle; scale bar, 20 μm) as well as the repopulation of hair cells to control level by 96 hpe in the damaged area (bottom). D, Quantification of hair cell number after sound exposure (one-way ANOVA and post hoc test, n = 3, *p < 0.05; all error bars in this publication demonstrate SD). D, Dorsal; A, anterior.
Candidate gene/pathway identification by DGE
Based on the time line of hair cell regeneration in adult fish (Fig. 1C,D), we collected inner ear tissue samples at 0, 24, 48, and 96 hpe and generated the expression profiles from the samples using the DGE tag-profiling technique (Morrissy et al., 2009). The raw data from profiling included sequences of 944,347 unique tag sequences (20 nt) and a “count” associated with each tag sequence (Table 2). The count is a direct quantification of the copy number of the transcript that the tag represents. The total count number of all the tag sequences was 15,744,948. After filtering the raw data (see Materials and Methods), we mapped 303,342 unique tags from a total of 15,103,943 sequences to transcriptome and genome databases (Table 2). Approximately 60% of the tags were mapped to at least one known/predicted transcript or at least one location on the genome (Fig. 2A). Although ∼40% of the unique tag sequences failed to be mapped, the total proportion of unmapped tags only accounted for ∼7.2% of the total tag counts. When one mismatch was allowed during the mapping, only ∼7% of the tag sequences failed to be mapped, suggesting that the failure in mapping most likely resulted from sequencing errors and/or single-nucleotide polymorphisms. In this study, we adopted conservative criteria, only analyzing tags with no mismatch in mapping and matching to a single UniGene entry.
Table 2.
Summary of tag profiling results
| Total |
After filtration |
|||
|---|---|---|---|---|
| Count | Unique tag | Count | Unique tag | |
| Control | 3,339,753 | 327,240 | 3,199,598 | 187,085 |
| 0 hpe | 2,758,836 | 279,532 | 2,646,064 | 166,760 |
| 24 hpe | 3,615,956 | 317,505 | 3,486,384 | 187,933 |
| 48 hpe | 3,476,027 | 331,687 | 3,337,409 | 193,069 |
| 96 hpe | 2,554,376 | 289,177 | 2,434,488 | 169,289 |
| Total | 15,744,948 | 944,347 | 15,103,943 | 303,342 |
The numbers of tag counts and unique tag sequences from each of the five expression profiles in raw data (under the “Total” column) and after the removal of the tag sequences that were detected only once in five profiles (under the “After filtration” column).
Figure 2.
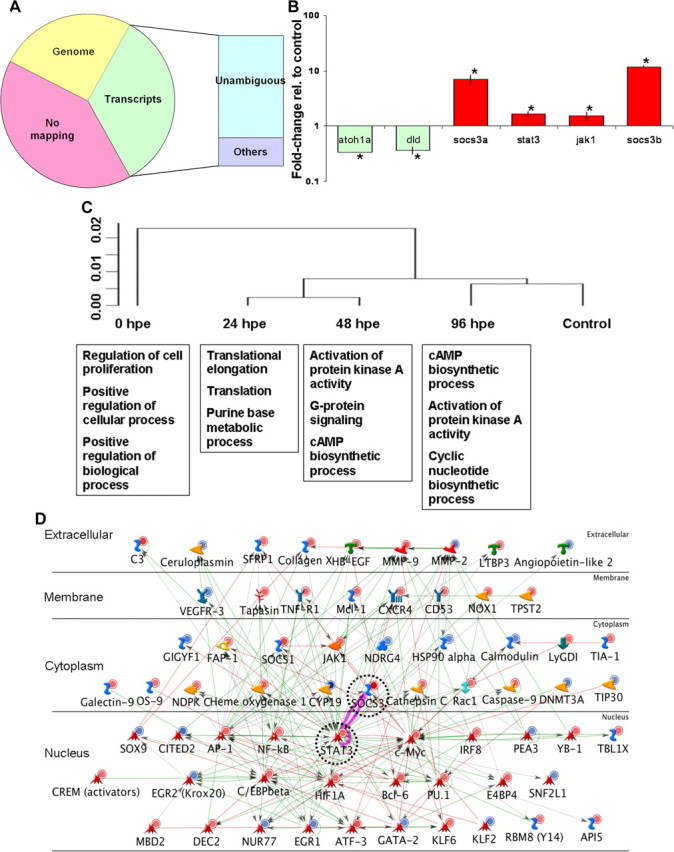
Analyses and confirmation of tag-profiling results from inner ear tissues collected during hair cell regeneration. A, A total number of 303,342 unique-sequence tag sequences from inner ear tissues collected at five time points were mapped against transcriptome and genome databases. Approximately 34% of the tag sequences were mapped to one or more known/predicted transcripts and ∼17% to unique loci in genome (without a known transcript), leaving ∼49% of the sequences without transcriptome or genome mapping. Among those tags mapped to known/predicted transcripts, ∼78% were unambiguously mapped. B, Some of the candidate genes (0 hpe) identified by tag profiling were confirmed by qRT-PCR using GAPDH as a reference gene (n = 3, one-tailed t test, *p = 1.81e-4, 3.78e-4, 9.41e-3, 8.76e-3, 0.0272, 1.578e-4). C, Clustering analysis of the five expression profiles with Genesifter showed the relationship between different profiles (top diagram). The clustering results had the predicted relationship where the 0 hpe expression was the most different from control and then the samples progressively returned to “normal” over time. Pathway analysis showed cell signaling pathways represented by the candidate genes that are critical for specific phases of regeneration [e.g., cell proliferation-related at 0 hpe and cell differentiation-related at later time points (bottom boxes)]. D, Pathway analysis of the candidate genes (identified at 0 hpe) involved in inner ear hair cell regeneration highlighted the interactions between stat3 and socs3 (dashed circles) with other identified candidate genes. The known interactions (red: positive; green: negative; gray: unspecified) between the human orthologs of the candidate genes were extracted in batches to predict the candidate pathways involved in the hair cell regeneration. Different colors of the circles indicate those genes being upregulated (red) or downregulated (blue) during hair cell regeneration.
A total of 80,604 unique tags were unambiguously mapped to a known/predicted transcript (Fig. 2A) (i.e., the tag could be assigned to one and only one of the known/predicted transcripts). Those 80,640 tags were used for gene identification and expression level calculations. Clustering analysis of the five expression profiles with Genesifter (Geospiza) software based on the calculation of the overall differences between profiles showed that the 96 hpe profile shared the highest similarity with the control profile, followed by the 24 hpe and the 48 hpe profiles, which were highly similar to each other, and then by a 0 hpe profile that differed the most from the control profile (Fig. 2C, top). Essentially the transcriptional profiles were most different immediately after noise exposure and slowly returned to normal over the course of 4 d.
We used χ2 tests and Fisher's exact tests to compare the expression level of each gene during regeneration to the control level. A total of 2269 genes with significant changes in their expression levels at one or more time points during regeneration were identified as candidate genes (data available upon request). Measureable changes in expression levels detected by DGE were confirmed in several candidate genes (selected because of already known functions in hair cell development or because of direct interest in the genes) with quantitative RT-PCR (qRT-PCR) (Fig. 2B). We performed a pathway analysis for the 2269 candidate genes using the Metacore (GeneGo) software package. The functions of the most enriched interaction network calculated from the candidate genes in each experimental time point are listed in Figure 2C (bottom). While the enriched pathways identified by Metacore were generally broad, the fact that the different collected time points had very different enrichment profiles, indicated distinct phases in the regeneration process. The results of profile distance calculation and enrichment analysis corresponded nicely with the distinct morphological changes in the saccular epithelium during regeneration, as well as with the known cellular mechanisms during hair regeneration in other nonmammalian vertebrates (Stone and Cotanche, 2007).
Because we were interested in identifying early signaling events that trigger regeneration, we prioritized our candidates for functional analysis from the highly enriched networks that were apparent at the earliest measured time point (0 hpe, Fig. 2D). Metacore and Pathway Studio both identified a large interconnected pathway with the hub of that pathway centering on stat3 (Fig. 2D). Confirming the potential importance, the gene socs3a (a zebrafish homolog of human SOCS3), a downstream target of stat3 (and a negative-feedback inhibitor for stat3 activity) was one of the largest transcriptional changes at 0 hpe (≈10-fold increase). By 24 hpe, stat3 and socs3a signaling return to normal levels, suggesting that this response is an important early regenerative response but is not necessary in later stages of regeneration. With the help of the analysis, we identified the stat3/socs3 pathway as the dominant signaling pathway to become activated at the earliest time point of recovery with >50 relevant genes in the pathway responding to hair cell damage (Fig. 2D). No other known signaling pathway had as many connected nodes as the stat3/socs3a node in the 0 hpe responses. Related genes in the stat3/socs3 pathway, such as socs3b (a paralog of socs3a in zebrafish), Janus kinase 1 (jak1), and matrix metalloproteinase 9 (mmp9), also showed significant changes in their expression levels (Fig. 2B; data available on request).
Self-restrictive feedback between stat3 and socs3a in zebrafish
In mammals, there is a self-restrictive feedback mechanism between stat3 and socs3: stat3 activates the transcription of socs3, which in turn, antagonizes stat3 activation (Leonard and O'Shea, 1998). To test whether there is a similar feedback mechanism between stat3 and socs3a in zebrafish, we temporarily knocked down stat3 or socs3a in zebrafish embryos using MO injection. We detected a decrease in the mRNA levels of both stat3 and socs3a in stat3 morphants but an increase in the mRNA levels of both genes in socs3a morphants (Table 3). This is the expected result for a negative-feedback loop. To further confirm the knock-down studies, we injected mature mRNA of socs3a to zebrafish embryos and found a reduction in stat3 expression level, mimicking the effects of stat3-MO injection (Table 3). These data demonstrate that the self-restrictive feedback relation between stat3 and socs3a in mammals is maintained in zebrafish.
Table 3.
Summary of qRT-PCR results in morpholino- and mRNA-injected embryos
| Target gene |
|||
|---|---|---|---|
| Atoh1a | Stat3 | Socs3a | |
| socs3a MO (36 hpf) | 1.35 (9.91e-3) | 1.76 (0.0284) | 8.37 (0.0324) |
| socs3a mRNA (12 hpf) | 0.615 (2.70e-3) | 0.720 (0.0235)a | |
| stat3 MO (32 hpf) | 0.547 (0.0105) | 0.294 (1.77e-4) | 0.600 (0.041) |
In developing embryos, stat3 and socs3a were knocked down respectively with MO injection, and socs3a was also temporarily overexpressed with mRNA injection. Changes in mRNA levels of atoh1a, stat3, and socs3a in MO/mRNA-injected embryos compared to control embryos were determined with qRT-PCR. The numbers in each cell stand for fold change and p value (in braces).
aData collected from a 6 hpf embryo.
The stat3/socs3a pathway is involved in hair cell production in zebrafish development
In addition to the hair cells in their inner ears, zebrafish, like other fishes and amphibians, also possess the mechanosensory lateral line organ for detecting water movement over the body (Montgomery et al., 2000; McHenry et al., 2009). The neuromasts in the lateral line are composed of hair cells and supporting cells that are highly similar to those in the inner ear sensory epithelium (Nicolson, 2005). In addition, the hair cell regeneration in the lateral line neuromasts shares a similar molecular mechanism to that in the inner ear (Harris et al., 2003; Ma et al., 2008; Behra et al., 2009).
Both stat3 (Oates et al., 1999; Thisse and Thisse, 2004) and socs3a (Fig. 3A) are expressed in the neuromasts at 5 dpf, implying a normal function in those tissues for either development or homeostasis. In addition, stat3 is known to be expressed in the anterior region of the otic vesicle at earlier developmental stages (Oates et al., 1999; Thisse and Thisse, 2004). As an initial test of the roles for stat3 and socs3 in hair cell regeneration, we tested the effects of morpholino inhibition of each gene on normal inner ear development. Testing two different morpholinos for socs3a and a previously published and verified one for stat3 (Yamashita et al., 2002), knocking down stat3 or socs3a had a clear inhibitory effect on hair cell production (Fig. 3B–E). In stat3 morphants, the anterior otolith in the otic vesicles was often missing while the posterior otolith appeared approximately normal (Fig. 3B). Correspondingly, a closer examination of the maculae in the otic vesicle revealed a significantly decreased number of hair cells in the anterior macula in those morphants while the hair cell number in the posterior macula remained normal (Fig. 3C,D). Coinjection with tp53 MO resulted in an identical phenotype (data not shown), suggesting that the hair cell-related phenotypes are gene specific. The phenotypes of the morphants corresponded nicely with the expression pattern of both genes detected by in situ hybridization (Oates et al., 1999; Thisse and Thisse, 2004) (Fig. 3A). The phenotypes also serve as an indirect confirmation of the involvement stat3/socs3 pathway in hair cell regeneration, as hair cell production during development and regeneration will, to some extent, share similar genetic programs (Stone and Rubel, 1999; Cafaro et al., 2007; Ma et al., 2008; Daudet et al., 2009).
Figure 3.
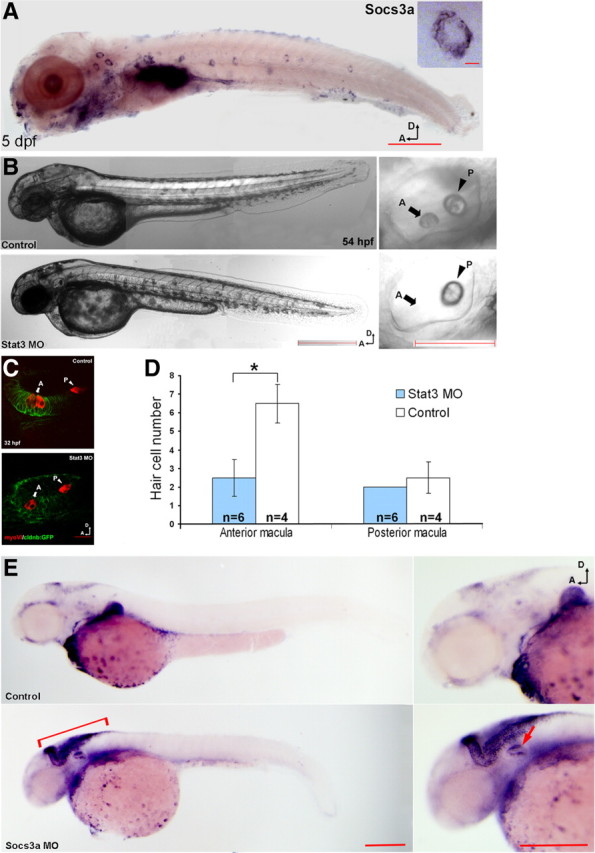
stat3 and socs3a are involved in hair cell production during zebrafish larval development. A, socs3a expression was detected in anterior and posterior lateral line neuromasts in 5 dpf larvae by in situ hybridization; there is a close-up of one neuromast in the top right corner (scale bars: top, 10 μm; bottom, 1 mm). Expression of socs3a is similar to stat3 expression (Oates et al., 1999; Thisse and Thisse, 2004). B, The anterior otolith (arrows labeled with “A”) in stat3 morphants was typically missing, while the posterior otolith (arrowheads labeled with “P”) appeared approximately normal (scale bars: left, 500 μm; right, 100 μm). C, MyosinVI antibody staining (red channel) of the hair cells in the anterior sensory macula (arrows labeled with A) and posterior macula (arrowheads labeled with P) in control larvae and stat3 morphants (scale bar, 20 μm) at 32 hpf. cldnb:GFP (green channel) was used to outline the otic vesicle. D, The stat3 morphants showed a significant reduction in the number of hair cells in the anterior macula, but not in the posterior macula at 32 hpf (n = 4 (control)/6 (MOs), one-tailed t test, *p = 2.79e-4). E, Both stat3 and socs3a morphants possessed fewer posterior lateral line neuromasts (n = 10 in all groups, one-tailed t test, *p = 2.52e-08, 2.41e-04) with a smaller number of hair cells per neuromast (n = 10 in all groups, one-tailed t test, *p = 1.10e-06, 1.56e-10) compared with control larvae at 2.5 dpf. In socs3a morphants (bottom), an expansion of atoh1a expression was detected by in situ hybridization in the brain area (bracket) as well as the otic vesicle (arrow) compared with control (top) (scale bars: left, 1 mm; right, 500 μm). D, Dorsal; A, anterior; MO, morpholino.
To further understand the function of the stat3/socs3 pathway in hair cell production, we measured the mRNA level of atonal homolog 1a (atoh1a) in stat3 and socs3a morphants. atoh1a is one of the zebrafish homologs of atoh1, an essential gene required for hair cell fate commitment (Bermingham et al., 1999; Woods et al., 2004; Millimaki et al., 2007). Both the knocking down of stat3 and the overexpression of socs3a resulted in a reduction in atoh1a mRNA level (Table 3) but did not change the overall distribution or timing of expression based on in situ hybridization (data not shown). In contrast, a strong increase in atoh1a mRNA level was observed in socs3a morphants where stat3 was hyperactivated (Table 3), demonstrating that stat3/socs3a are positive/negative regulators of atoh1a expression. In addition, the cross talk between stat3/socs3 and atoh1 was further confirmed by in situ hybridization of the socs3a morphants (Fig. 3E). At 24 and 36 hpf, there is little detectable difference in atoh1a expression between socs3a morphants and wild-type fish by in situ hybridization (data not shown). But at 48 and 72 hpf, there is a clear expansion of atoh1a expression in the otic vesicle (Fig. 3E, arrowhead), the hindbrain, and the spinal chord (Fig. 3E, bracket), suggesting the stat3–atoh1a regulatory relationship is not unique to the sensory epithelia of the inner ear and lateral line, but is a more general pathway for regulation. These results suggest that the stat3/socs3 pathway is involved in the hair cell differentiation process possibly by directly or indirectly expanding atoh1a expression.
Stat3 activation during hair cell regeneration and development in lateral line neuromasts
Because the lateral line neuromasts are located on the surface of the body, it makes the lateral line a convenient model for testing hair cell regeneration. Expression of both stat3 (Thisse and Thisse, 2004) and socs3a (Fig. 3A) was detected in the neuromasts at 5 dpf, so we continued with additional functional studies of the stat3/socs3a pathway during regeneration in larval neuromasts. At 5 dpf, the number of hair cells in the lateral line neuromasts reaches a relatively stable level (Ma et al., 2008; Behra et al., 2009), which enables us to differentiate regeneration of hair cells from normal growth and development.
We induced hair cell death in the lateral line neuromasts with CuSO4, a protocol ensuring complete lateral line hair cell elimination within 2 h in 5 dpf larvae (Hernández et al., 2007), which is followed subsequently by regeneration of hair cells to control levels within 72 h (Behra et al., 2009). We performed in situ hybridization in larval fish after 2 h of CuSO4 treatment and found an increase in the level of stat3 expression in the lateral line neuromasts when hair cell death was induced (Fig. 4A). In addition, we also found changes in the location of stat3 expression induced by CuSO4-triggered hair cell death (Fig. 4A), which was further confirmed by fluorescent whole-mount in situ hybridization in ET20 larvae that marks the mantle cells of the neuromasts with GFP expression the outermost layer of nonsensory cells in the neuromast (Parinov et al., 2004) (Fig. 4B). RNA probes against stat3 were mostly detected in the GFP-positive cells of the neuromasts in untreated ET20 larvae (Fig. 4B, top). When hair cell death was induced, stat3 expression was no longer limited to the mantle cells, but was expressed strongly throughout the supporting cells of the neuromasts. The in situ hybridization data confirmed the activation of stat3 induced by hair cell death in the lateral line neuromasts was in the supporting cell population, matching expression increases observed by DGE during inner ear hair cell regeneration.
Figure 4.
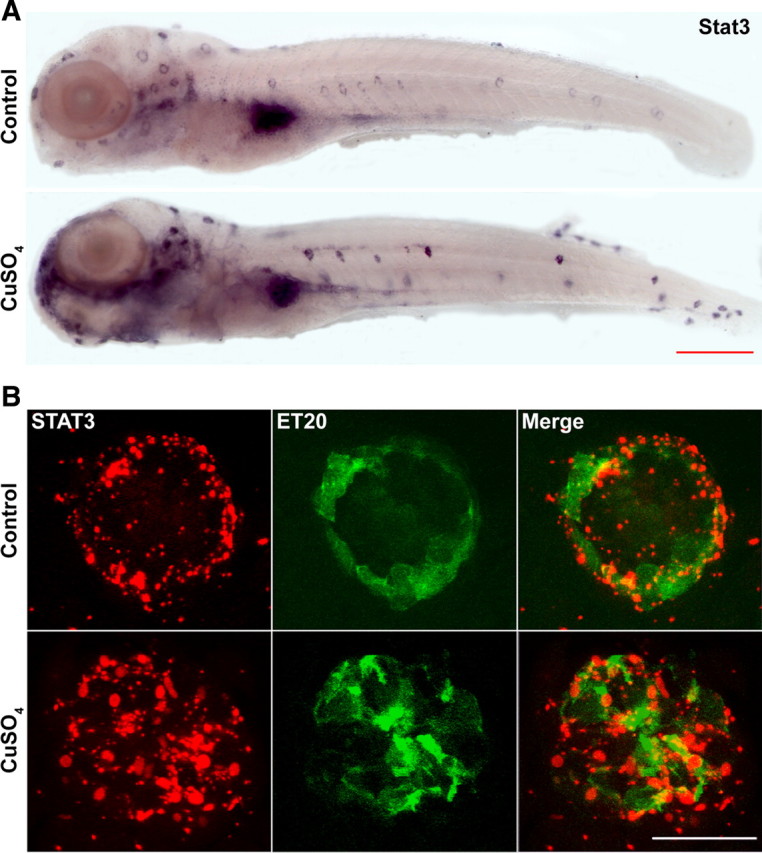
Hair cell death in the lateral line system resulted in an increase in stat3 expression level in the nonsensory cells in the neuromasts. A, The results of regular whole-mount in situ hybridization targeting stat3 mRNA in control (top) CuSO4-treated (bottom) larvae showed an increase in stat3 expression when hair cell death took place. Scale bar, 1 mm. B, Costaining of neuromasts with fluorescent detection of probes targeting stat3 mRNA (red channel) and FITC-conjugated anti-GFP antibody (green channel) in ET20 transgenic larvae showed an increase in nonsensory cells in the neuromasts after hair cell death. Scale bar, 20 μm.
The activation of mammalian stat3 protein is mostly characterized by the phosphorylation of either Y705 (Yoshimura, 2009; Yu et al., 2009) or S727, which functions concurrently with Y705 (Wen et al., 1995) or independently (Liu et al., 2003; Qin et al., 2008). The corresponding amino acid (S751) and the flanking amino acid sequences are highly conserved in zebrafish stat3 when compared with the human protein, so we first tracked the activation of stat3 in the neuromasts after CuSO4 treatment with an antibody that recognizes STAT3 S727 phosphorylation (STAT3pS727). We found a significant increase in STAT3pS727-positive nuclei in the neuromasts at 12 hpt with CuSO4 in the CuSO4-treated larvae relative to control (Fig. 4A,B). Double labeling with GFP in Tg(scm1:GFP) (Behra et al., 2009) demonstrates that the cells with nuclear staining of STAT3pS727 are the supporting cells (Fig. 4A). The activation and the nuclear import of stat3 protein in the supporting cells subsequent to hair cell death indicate the activation of stat3 signaling during hair cell regeneration.
Hair cell production during development and regeneration will, to some extent, share similar genetic programs (Stone and Rubel, 1999; Cafaro et al., 2007; Ma et al., 2008; Daudet et al., 2009), so we checked whether stat3 activation in the lateral line neuromasts is similar during development and regeneration. In 3 dpf larvae, most cells in the neuromasts showed homogeneous STAT3pS727 labeling only in their nuclei (Fig. 5A′, top). Double-labeling with scm1:GFP demonstrates that those cells are supporting cells (Fig. 5A′, top). In addition, in differentiating hair cells, the nuclear labeling of STAT3pS727 antibody decreased and became spotty, with cytoplasmic labeling becoming predominant, suggesting that the stat3 was becoming inactive in these cells (Fig. 5A′, top, close-ups). The nuclear import of stat3 in supporting cells during hair cell regeneration in 5 dpf and older larvae seems to be recapitulating the stat3 activation during immature neuromast development in younger larvae.
Figure 5.
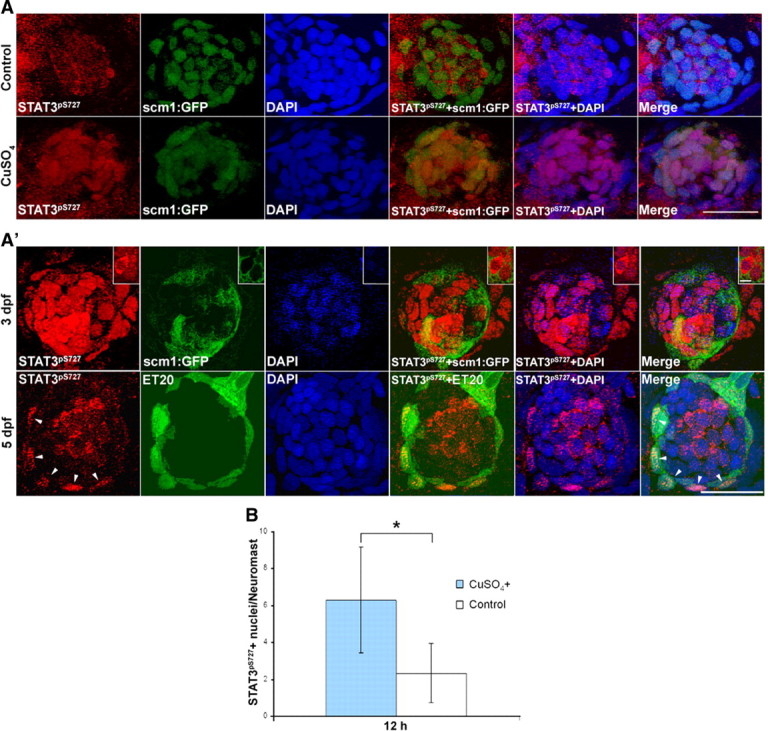
Phosphorylation and nuclear import of stat3 proteins were detected in the regenerating and developing neuromasts. A, Costaining of anti-STAT3pS727 with DAPI in Tg(scm1:GFP) larvae confirmed anti-STAT3pS727 labeling mainly in the nuclei of supporting cells (GFP positive) at 12 hpt with copper compared with untreated controls. Scale bar, 20 μm. A′, In 3 dpf larvae, the majority of the supporting cell nuclei (GFP-positive cells) were labeled with STAT3pS727 antibody (top). Differentiating hair cells showed decreased STAT3pS727 nuclear staining, but an increase of STAT3pS727 labeling in the cytoplasm (top, close-ups; scale bar: close-ups, 5 μm). Nuclear anti-STAT3pS727 labeling decreased dramatically in more mature neuromasts in 5 dpf larvae (bottom), except for a small group of cells labeled with GFP (arrowheads) in ET20 transgenic larvae (scale bars, 20 μm). B, Quantification of activated phospho-STAT3. The increase in nuclear localized phospho-stat3 was statistically significant (n = 10/9, one-tailed t test, p = 9.83e-4). S727 in human STAT3 corresponds to S751 in zebrafish STAT3 by sequence alignment.
It is noteworthy that in older larvae, a significant level of labeling was still detectable in a subgroup of mantle cells that are marked by GFP in the ET20 transgenic line (Fig. 5A′, bottom). The significance of this cell population having activated stat3 is not yet clear, but it seems to represent a distinct cellular compartment in the lateral line neuromasts.
Inhibition of stat3-promoted hair cell regeneration
To further clarify the function of stat3, we compared the hair cell regeneration processes with and without the presence of a stat3 inhibitor. The majority of the chemical inhibitors were ototoxic and could not be used for regeneration studies (data not shown). One published inhibitor, S3I-201, was not toxic on its own and was used to test the role of stat3 in hair cell regeneration. S3I-201 is a cell-permeable chemical that binds to the SH2 domain of mammalian stat3 protein and reportedly blocks the dimerization of phosphorylated (activated) stat3 molecules (Siddiquee et al., 2007). After exposing Tg(pou4f3:GFP) larvae (in which the hair cells are labeled with GFP) (Xiao et al., 2005) to CuSO4, we quantified the regenerating hair cells and performed a BrdU incorporation assay (Ma et al., 2008). The S3I-201-treated larvae showed a significant increase in BrdU incorporation in the neuromasts compared with DMSO-treated larvae (control) at 24 hpt, but not at 48 or 72 hpt (Fig. 6A). Accordingly, the S3I-201-treated larvae had more hair cells per neuromast at 48 hpt, but not at 24 or 72 hpt (Fig. 6A), which is consistent with an expected lag between supporting cell division and subsequent hair cell differentiation. In essence, the hair cells regenerated faster in the S3I-201-treated embryos than in controls. Similar results were observed when hair cell quantification was done by myosin VI antibody staining (data not shown). S3I-201 did not have a significant impact on hair cell numbers in larvae that had not been treated with CuSO4 (data not shown).
Figure 6.
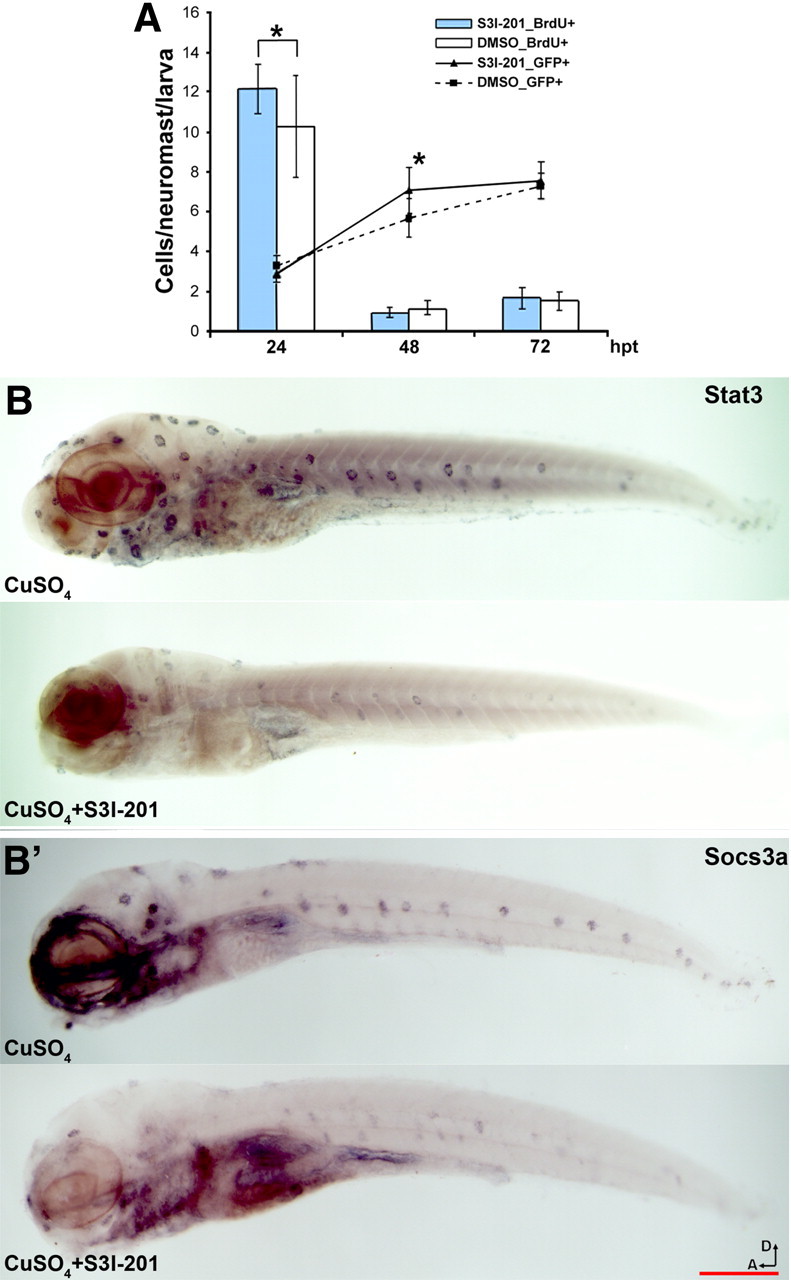
S3I-201 promoted lateral line hair cell regeneration by downregulating stat3/socs3 signaling. A, Quantification of hair cells after lateral line hair cell loss induced by CuSO4 treatment; larvae incubated with S3I-201 had more hair cells [GFP-positive cells in Tg(pou4f3:GFP)] per neuromast at 48 hpt with CuSO4 compared with the control larvae incubated with DMSO (n = 12/9, one-tailed t test, *p = 5.34e-3), but not at 24 or 72 hpt (line graph). BrdU incorporation assays showed a significantly higher number of BrdU-positive cells per neuromast in larvae incubated with S3I-201 at 24 hpt (n = 16/10; one-tailed t test, *p = 9.24e-3), but not at later time points (bar graph). B, B′, 5 dpf larvae fish were treated with CuSO4 with or without the presence of S3I-201 for 1 h and examined with in situ hybridization. stat3 (B) and socs3a (B′) expression levels were both reduced in larvae treated with both CuSO4 and S3I-201 (B and B′, top) compared with those treated with only CuSO4 (B and B′, bottom). Scale bar, 1 mm. D, Dorsal; A, anterior.
To confirm the inhibitory function of S3I-201 in hair cell regeneration triggered by CuSO4 ototoxicity, we performed ISH to examine the mRNA levels of stat3 and socs3a in the lateral line neuromasts after 1 h of CuSO4 treatment with or without the presence of S3I-201. The ISH data demonstrated a reduction in both stat3 (Fig. 6B) and socs3a (Fig. 6B′) expression in the neuromasts cotreated with both CuSO4 and S3I-201 compared with those treated only with CuSO4. Our qRT-PCR results were in accordance with the ISH results in that stat3 expression in larvae treated with both CuSO4 and S3I-201 was only 81% of the level in larvae treated only with CuSO4 (p = 0.0014, n = 3 for both groups). Since stat3 activates its own expression, the data suggest that S3I-201 stimulated hair cell regeneration by moderately inhibiting stat3 activity.
Discussion
Recent studies have demonstrated the power of using the DGE platform for measuring transcriptional changes (‘t Hoen et al., 2008; Morrissy et al., 2009). Using DGE, we generated a comprehensive list of candidate genes involved in the inner ear hair cell regeneration in adult zebrafish (data available upon request). One major pathway that is initiated in the earliest responses centers on stat3 and socs3a. This pathway is broadly implicated in the initial signaling responses of hair cell regeneration in the zebrafish inner ear. We went on to show that stat3/socs3 signaling played an important role in cell division and hair cell differentiation during both normal development and tissue regeneration.
Stat3 signaling: activated during development and regeneration
To some extent, hair cell production during both development and regeneration share similar genetic programs (Stone and Rubel, 1999; Cafaro et al., 2007; Ma et al., 2008; Daudet et al., 2009). In this study, we identified stat3 and socs3a as important regulators in zebrafish hair cell regeneration in the inner ear (data available upon request) and lateral line system (Fig. 5A). Correspondingly, we also found them required for normal hair cell production during the larval development (Fig. 3B–E). The examination of stat3 activity in lateral line neuromasts during development and regeneration further confirms the similarity shared between the two processes. Activated, nuclear-localized stat3 protein was detected in a large number of supporting cells and differentiating hair cells in 3 dpf larvae (Fig. 5A′) and was downregulated in those same cells in the neuromasts of more mature 5 dpf larvae (Fig. 5A). However, when hair cell death was triggered in 5 dpf larvae, the stat3 protein was again activated and imported into the nuclei of the supporting cells (Fig. 5A), recapitulating the active stat3 observed during neuromast development (Fig. 5A′).
Another interesting observation is the specific overlap of STAT3pS727-positive cells and the GFP-positive mantle cells of ET20 larvae (Fig. 5A′), which is further supported by the enrichment of stat3 mRNA in those GFP-positive cells (Fig. 4A). This suggests that the mantle cells have a specific function in the neuromast that requires activated stat3. Previous publications suggest two types of stem cell-like cells in the neuromasts, one type involved in long-term cell replacement (i.e., the true stem cells), and the other in an amplification/differentiation role (Williams and Holder, 2000; Ma et al., 2008). The role of the ET20:GFP-labeled (and stat3-activated) mantle cells may regulate the cell division of adjacent stem cells through a nonautonomous mechanism similar to the role of stat92E in escort cells for regulating the stem cell division in the Drosophila ovary (Decotto and Spradling, 2005). These ET20:GFP and STAT3pS721-positive cells would represent part of the niche environment necessary to promote normal cell turnover as well as tissue regeneration. Alternatively, the mantle cells might directly divide asymmetrically to repopulate the supporting cells of the neuromasts after damage or cell turnover. The triggering of cell differentiation seemed to be achieved, at least in part, through activation of atoh1 (Fig. 3E), a key regulator of hair cell differentiation (Fig. 7). This pathway seems to be used in more than one tissue type as socs3a inhibition results in an expansion of atoh1a expression in both the otic vesicle and in the brain. It is noteworthy that while atoh1a expression is expanded in the otic vesicle with a hyperactive stat3, the number of hair cells was not (data not shown), suggesting that more factors are necessary for hair cell differentiation.
Figure 7.

The working model of the stat3/socs3 pathway and the interaction of stat3/socs3 with atoh1 during hair cell regeneration in zebrafish. stat3 positively regulates the function of atoh1, a key player in promoting hair cell fate commitment, in a direct or indirect manner. In addition, the balancing of the stat3/socs3 pathway is crucial to hair cell differentiation: either hyperactivation or hypoactivation of stat3 inhibits hair cell differentiation. The self-restrictive loop helps to balance the expression of stat3 spatially and temporally to ensure the appropriate replenishment of the lost hair cells.
A previous microarray study did show basal-level expression of stat3 in newborn rat cochlea that could be elevated by stress in explant cultures (Gross et al., 2008). It was also reported that stat3 showed a temporal expression pattern similar to gata2 and C/EBP during the in vitro differentiation of conditional cell lines derived from mouse otocyst (Holley et al., 2007). Little is known about the functions of stat3 and socs3 in the inner ear in mammals other than that activated stat3 protein is detected in the outer hair cells in mouse cochlea late in embryonic development (Hertzano et al., 2004). The persistence of stat3 expression in a subset of supporting cells in zebrafish that is not detected in the mammalian cochlea might represent a key difference that enables nonmammalian vertebrates to regenerate hair cells. A more detailed comparison study focusing on the stat3/socs3 signaling pathway in the inner ears in mice and zebrafish would help us further understand key regulators of hair cell regeneration. In addition, it is also intriguing to explore whether cell cycle progression can be triggered in supporting cells by genetic and/or pharmacological enhancement of stat3 activity in damaged mammalian sensory epithelia.
The stat3/socs3 pathway may serve as a common initiator in a variety of regenerative processes
The stat3/socs3 pathway is best known as the mediator of a wide variety of biological processes, including cell proliferation, cell migration, immune response, and cell survival (Yoshimura, 2009; Yu et al., 2009). Studies in mouse skin (Sano et al., 1999; Zhu et al., 2008) and liver (Dierssen et al., 2008; Riehle et al., 2008) further confirm the importance of the stat3/socs3 pathway for proper wound healing. For example, in microarray datasets from other zebrafish regeneration experiments, a strong induction of socs3 expression was consistently observed during the regeneration of fins (Schebesta et al., 2006) and retinas (Qin et al., 2009). stat3 expression was also found to be upregulated in dividing cells during zebrafish retinal photoreceptor regeneration (Kassen et al., 2009). The early activation of stat3 in the supporting cells subsequent to hair cell death is consistent with these previous studies and suggests that this is a common pathway for healing. stat–jak signaling has been found to play a critical role in the development and regeneration of the epithelial cells in the Drosophila midgut by initiating cell division and inducing differentiation (Jiang et al., 2009), a dual process that is highly similar to what we demonstrated during regeneration of the inner ear sensory epithelia. In zebrafish inner ear and lateral line regeneration, the activation of stat3 signaling in supporting cells precedes the cell division, suggesting that the gene is playing a role in cell cycle progression of those cells. A recent study supports this idea as it reported leukemia inhibitory factor promoted cell division in a cochlea-derived cell line, HEI-OC1, through the stat3/jak2 signaling pathway (Chen et al., 2010). This suggests that stat signaling is a fundamental initiating mechanism of tissue regeneration that operates through cell division and differentiation and has been conserved for over the 300 million years of evolution from invertebrates to vertebrates. Together, all these data strengthen our hypothesis that stat3/socs3 is a central activating mechanism of stem cell populations in most, if not all, forms of regeneration.
Although stat3 activation can be considered as a promoting factor for a regenerative process, we showed that a mild inhibition of stat3 via a chemical antagonist (Fig. 6B,B′) could accelerate hair cell regeneration by promoting supporting cell division (Fig. 6A). Because stat3 is also known to be required for the maintenance of pluripotency of various types of stem cells (Raz et al., 1999), and we did identify a small group of putative stem cells in the neuromasts with persisting stat3 activity under quiescent/undamaged conditions (Fig. 5A′), a possible explanation for the observation that inhibiting stat3 modestly enhanced regeneration at early time points is that S3I-201 forced the premature division and differentiation of these stem cells, resulting in an increase in BrdU incorporation at 24 hpt and in hair cell number at 48 hpt (Fig. 6A). However, we do not believe such a mechanism contributed much, if any, to the difference between S3I-201-treated larvae and control larvae during regeneration. First, we observed an accelerated regeneration without overproduction or underproduction of hair cells (Fig. 6A), suggesting that we were simply accelerating the normal process. Meanwhile, undamaged larvae raised with S3I-201 from 5 to 8 dpf did not show any significant difference in the number of the lateral hair cells compared with control, showing it was not the release of a cell cycle block in the mantle (ET20-positive) cells that explains the accelerated appearance of the hair cells. In addition, the damage-induced stat3 activation was observed in all of the supporting cells (Fig. 4B), the main source of new hair cells during regeneration (Ma et al., 2008). While we cannot completely rule out the contribution of any other mechanism, the data here do support our hypothesis that S3I-201 promoted the division of the supporting cells through mild downregulation of the damage-induced stat3 activity in those cells. In essence, S3I-201 increased the rate of stat3 inactivation that naturally occurs through socs3a activation, allowing the cells to proceed to mitosis and cell differentiation sooner.
We believe that our results emphasize the balance of stat3/socs3 signaling as being crucial for a successful healing process. Two examples in the literature demonstrated that excessive stat3 activity can block cell cycle progression and inhibit regeneration similar to what we observed; prolonged overexpression of IL-6 and the resulting hyperactivation of stat3 inhibited regeneration after hepatectomy in mice (Wüstefeld et al., 2000; Jin et al., 2006). Moreover, excessive stat3 activity in fatty livers was associated with the cell cycle arrest after hepatectomy (Torbenson et al., 2002). Accordingly, keeping the cytokine signaling tempered by the negative feedback of socs3 is known to be crucial for proper regeneration (Taub, 2004). Tissue-specific knockout of socs3 in mice led to an upregulation in stat3 activity but delayed skin wound healing due to the increased neutrophil and macrophage infiltration and elevated chemokine expression (Zhu et al., 2008). In addition, we are not ruling out the possibility that stat3 may play more than one role in more than one type of cell in tissue regeneration. It is known that stat3 is activated in hepatocytes as well as myeloid cells after hepatectomy (Wang et al., 2010). However, while knocking out stat3 in hepatocytes inhibits cell cycle progression and regeneration, knocking out stat3 in myeloid cells leads to the opposite results (Wang et al., 2010). Previous data also suggest a complicated cross talk of stat3 signaling pathway in different cell types during regeneration (Wang et al., 2010). Given the known active involvement of myeloid cells in lateral line hair cell death in zebrafish larvae (d'Alençon et al., 2010), it is very likely that S3I-201 treatment disturbed the stat3 signaling in not only supporting cells but also myeloid cells and even the cross talk between these two types of cells. Our data together with data from many others demonstrate crucial yet complicated roles of the stat3/socs3 pathway during tissue regeneration. We have developed a model for stat3 that requires initial activation of stat3 with a critical negative-feedback role for socs3a. Further studies are necessary to work out the exact roles of each gene in this complex and dynamic process. While stat3 activation could potentially be therapeutic to injuries in a wide variety of tissues, caution needs to be taken for such application, given the multifaceted functions of stat3 in the injury and healing process.
Footnotes
This research was supported by the Intramural Research Program of the National Human Genome Research Institute, National Institutes of Health (S.M.B.). We thank A. Popper for use of the sound exposure equipment and helpful discussions, and D. Bodine and W. Pavan for critical reading of the manuscript.
The authors declare no competing financial interests.
References
- Amsterdam A, Burgess S, Golling G, Chen W, Sun Z, Townsend K, Farrington S, Haldi M, Hopkins N. A large-scale insertional mutagenesis screen in zebrafish. Gene Dev. 1999;13:2713–2724. doi: 10.1101/gad.13.20.2713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baird RA, Steyger PS, Schuff NR. Mitotic and nonmitotic hair cell regeneration in the bullfrog vestibular otolith organs. Ann NY Acad Sci. 1996;781:59–70. doi: 10.1111/j.1749-6632.1996.tb15693.x. [DOI] [PubMed] [Google Scholar]
- Balak KJ, Corwin JT, Jones JE. Regenerated hair cells can originate from supporting cell progeny: evidence from phototoxicity and laser ablation experiments in the lateral line system. J Neurosci. 1990;10:2502–2512. doi: 10.1523/JNEUROSCI.10-08-02502.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bang PI, Sewell WF, Malicki JJ. Morphology and cell type heterogeneities of the inner ear epithelia in adult and juvenile zebrafish (Danio rerio) J Comp Neurol. 2001;438:173–190. doi: 10.1002/cne.1308. [DOI] [PubMed] [Google Scholar]
- Behra M, Bradsher J, Sougrat R, Gallardo V, Allende ML, Burgess SM. Phoenix is required for mechanosensory hair cell regeneration in the zebrafish lateral line. PLoS Genet. 2009;5:e1000455. doi: 10.1371/journal.pgen.1000455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bermingham NA, Hassan BA, Price SD, Vollrath MA, Ben-Arie N, Eatock RA, Bellen HJ, Lysakowski A, Zoghbi HY. Math1: an essential gene for the generation of inner ear hair cells. Science. 1999;284:1837–1841. doi: 10.1126/science.284.5421.1837. [DOI] [PubMed] [Google Scholar]
- Brignull HR, Raible DW, Stone JS. Feathers and fins: non-mammalian models for hair cell regeneration. Brain Res. 2009;1277:12–23. doi: 10.1016/j.brainres.2009.02.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cafaro J, Lee GS, Stone JS. Atoh1 expression defines activated progenitors and differentiating hair cells during avian hair cell regeneration. Dev Dyn. 2007;236:156–170. doi: 10.1002/dvdy.21023. [DOI] [PubMed] [Google Scholar]
- Chen HC, Ma HI, Sytwu HK, Wang HW, Chen CC, Liu SC, Chen CH, Chen HK, Wang CH. Neural stem cells secrete factors that promote auditory cell proliferation via a leukemia inhibitory factor signaling pathway. J Neurosci Res. 2010;88:3308–3318. doi: 10.1002/jnr.22492. [DOI] [PubMed] [Google Scholar]
- Corwin JT, Cotanche DA. Regeneration of sensory hair cells after acoustic trauma. Science. 1988;240:1772–1774. doi: 10.1126/science.3381100. [DOI] [PubMed] [Google Scholar]
- Cruz RM, Lambert PR, Rubel EW. Light microscopic evidence of hair cell regeneration after gentamicin toxicity in chick cochlea. Arch Otolaryngol Head Neck Surg. 1987;113:1058–1062. doi: 10.1001/archotol.1987.01860100036017. [DOI] [PubMed] [Google Scholar]
- d'Alençon CA, Pena OA, Wittmann C, Gallardo VE, Jones RA, Loosli F, Liebel U, Grabher C, Allende ML. A high-throughput chemically induced inflammation assay in zebrafish. BMC Biol. 2010;8:151. doi: 10.1186/1741-7007-8-151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Daudet N, Gibson R, Shang J, Bernard A, Lewis J, Stone J. Notch regulation of progenitor cell behavior in quiescent and regenerating auditory epithelium of mature birds. Dev Biol. 2009;326:86–100. doi: 10.1016/j.ydbio.2008.10.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Decotto E, Spradling AC. The Drosophila ovarian and testis stem cell niches: Similar somatic stem cells and signals. Dev Cell. 2005;9:501–510. doi: 10.1016/j.devcel.2005.08.012. [DOI] [PubMed] [Google Scholar]
- Dierssen U, Beraza N, Lutz HH, Liedtke C, Ernst M, Wasmuth HE, Trautwein C. Molecular dissection of gp130-dependent pathways in hepatocytes during liver regeneration. J Biol Chem. 2008;283:9886–9895. doi: 10.1074/jbc.M705483200. [DOI] [PubMed] [Google Scholar]
- Fay RR. Phase-locking in goldfish saccular nerve fibres accounts for frequency discrimination capacities. Nature. 1978;275:320–322. doi: 10.1038/275320a0. [DOI] [PubMed] [Google Scholar]
- Fritzsch B, Pauley S, Beisel KW. Cells, molecules and morphogenesis: the making of the vertebrate ear. Brain Res. 2006;1091:151–171. doi: 10.1016/j.brainres.2006.02.078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fritzsch B, Beisel KW, Pauley S, Soukup G. Molecular evolution of the vertebrate mechanosensory cell and ear. Int J Dev Biol. 2007;51:663–678. doi: 10.1387/ijdb.072367bf. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gross J, Machulik A, Moller R, Fuchs J, Amarjargal N, Ungethüm U, Kuban RJ, Szczepek AJ, Haupt H, Mazurek B. mRNA expression of members of the IGF system in the organ of Corti, the modiolus and the stria vascularis of newborn rats. Growth Factors. 2008;26:180–191. doi: 10.1080/08977190802194317. [DOI] [PubMed] [Google Scholar]
- Haas P, Gilmour D. Chemokine signaling mediates self-organizing tissue migration in the zebrafish lateral line. Dev Cell. 2006;10:673–680. doi: 10.1016/j.devcel.2006.02.019. [DOI] [PubMed] [Google Scholar]
- Harris JA, Cheng AG, Cunningham LL, MacDonald G, Raible DW, Rubel EW. Neomycin-Induced hair cell death and rapid regeneration in the lateral line of zebrafish (Danio rerio) J Assoc Res Otolaryngol. 2003;4:219–234. doi: 10.1007/s10162-002-3022-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hernández PP, Olivari FA, Sarrazin AF, Sandoval PC, Allende ML. Regeneration in zebrafish lateral line neuromasts: Expression of the neural progenitor cell marker sox2 and proliferation-dependent and-independent mechanisms of hair cell renewal. Dev Neurobiol. 2007;67:637–654. doi: 10.1002/dneu.20386. [DOI] [PubMed] [Google Scholar]
- Hertzano R, Montcouquiol M, Rashi-Elkeles S, Elkon R, Yücel R, Frankel WN, Rechavi G, Möröy T, Friedman TB, Kelley MW, Avraham KB. Transcription profiling of inner ears from Pou4f3ddl/ddl identifies Gfi1 as a target of the Pou4f3 deafness gene. Hum Mol Genet. 2004;13:2143–2153. doi: 10.1093/hmg/ddh218. [DOI] [PubMed] [Google Scholar]
- Higgs DM, Souza MJ, Wilkins HR, Presson JC, Popper AN. Age- and size-related changes in the inner ear and hearing ability of the adult zebrafish (Danio rerio) J Assoc Res Otolaryngol. 2002;3:174–184. doi: 10.1007/s101620020035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- ‘t Hoen PAC, Ariyurek Y, Thygesen HH, Vreugdenhil E, Vossen RHAM, de Menezes RX, Boer JM, van Ommen GB, den Dunnen JT. Deep sequencing-based expression analysis shows major advances in robustness, resolution and inter-lab portability over five microarray platforms. Nucleic Acids Res. 2008;36:e141. doi: 10.1093/nar/gkn705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holley MC, Kneebone A, Milo M. Information for gene networks in inner ear development: a study centered on the transcription factor gata2. Hear Res. 2007;227:32–40. doi: 10.1016/j.heares.2006.04.006. [DOI] [PubMed] [Google Scholar]
- Jiang H, Patel PH, Kohlmaier A, Grenley MO, McEwen DG, Edgar BA. Cytokine/Jak/Stat signaling mediates regeneration and homeostasis in the Drosophila midgut. Cell. 2009;137:1343–1355. doi: 10.1016/j.cell.2009.05.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jin X, Zimmers TA, Perez EA, Pierce RH, Zhang Z, Koniaris LG. Paradoxical effects of short- and long-term interleukin-6 exposure on liver injury and repair. Hepatology. 2006;43:474–484. doi: 10.1002/hep.21087. [DOI] [PubMed] [Google Scholar]
- Jones JE, Corwin JT. Regeneration of sensory cells after laser ablation in the lateral line system: hair cell lineage and macrophage behavior revealed by time-lapse video microscopy. J Neurosci. 1996;16:649–662. doi: 10.1523/JNEUROSCI.16-02-00649.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kassen SC, Ramanan V, Montgomery JE, T Burket C, Liu CG, Vihtelic TS, Hyde DR. Time course analysis of gene expression during light-induced photoreceptor cell death and regeneration in albino zebrafish. Dev Neurobiol. 2007;67:1009–1031. doi: 10.1002/dneu.20362. [DOI] [PubMed] [Google Scholar]
- Kassen SC, Thummel R, Campochiaro LA, Harding MJ, Bennett NA, Hyde DR. CNTF induces photoreceptor neuroprotection and Müller glial cell proliferation through two different signaling pathways in the adult zebrafish retina. Exp Eye Res. 2009;88:1051–1064. doi: 10.1016/j.exer.2009.01.007. [DOI] [PubMed] [Google Scholar]
- Kratz E, Eimon PM, Mukhyala K, Stern H, Zha J, Strasser A, Hart R, Ashkenazi A. Functional characterization of the Bcl-2 gene family in the zebrafish. Cell Death Differ. 2006;13:1631–1640. doi: 10.1038/sj.cdd.4402016. [DOI] [PubMed] [Google Scholar]
- Leonard WJ, O'Shea JJ. Jaks and STATs: biological implications. Annu Rev Immunol. 1998;16:293–322. doi: 10.1146/annurev.immunol.16.1.293. [DOI] [PubMed] [Google Scholar]
- Liang J, Burgess SM. Gross and fine dissection of inner ear sensory epithelia in adult zebrafish (Danio rerio) J Vis Exp. 2009;27:1211. doi: 10.3791/1211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lim DJ. Ultrastructural cochlear changes following acoustic hyperstimulation and ototoxicity. Ann Otol Rhinol Laryngol. 1976;85:740–751. doi: 10.1177/000348947608500604. [DOI] [PubMed] [Google Scholar]
- Liu H, Ma Y, Cole SM, Zander C, Chen KH, Karras J, Pope RM. Serine phosphorylation of STAT3 is essential for Mcl-1 expression and macrophage survival. Blood. 2003;102:344–352. doi: 10.1182/blood-2002-11-3396. [DOI] [PubMed] [Google Scholar]
- Lombarte A, Yan HY, Popper AN, Chang JS, Platt C. Damage and regeneration of hair cell ciliary bundles in a fish ear following treatment with gentamicin. Hear Res. 1993;64:166–174. doi: 10.1016/0378-5955(93)90002-i. [DOI] [PubMed] [Google Scholar]
- Ma EY, Rubel EW, Raible DW. Notch signaling regulates the extent of hair cell regeneration in the zebrafish lateral line. J Neurosci. 2008;28:2261–2273. doi: 10.1523/JNEUROSCI.4372-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McGill TJ, Schuknecht HF. Human cochlear changes in noise induced hearing loss. Laryngoscope. 1976;86:1293–1302. doi: 10.1288/00005537-197609000-00001. [DOI] [PubMed] [Google Scholar]
- McHenry MJ, Feitl KE, Strother JA, Van Trump WJ. Larval zebrafish rapidly sense the water flow of a predator's strike. Biol Lett. 2009;5:477–479. doi: 10.1098/rsbl.2009.0048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Millimaki BB, Sweet EM, Dhason MS, Riley BB. Zebrafish atoh1 genes: classic proneural activity in the inner ear and regulation by Fgf and Notch. Development. 2007;134:295–305. doi: 10.1242/dev.02734. [DOI] [PubMed] [Google Scholar]
- Montgomery J, Carton G, Voigt R, Baker C, Diebel C. Sensory processing of water currents by fishes. Philos Trans R Soc Lond B Biol Sci. 2000;355:1325–1327. doi: 10.1098/rstb.2000.0693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morrissy AS, Morin RD, Delaney A, Zeng T, McDonald H, Jones S, Zhao Y, Hirst M, Marra MA. Next-generation tag sequencing for cancer gene expression profiling. Genome Res. 2009;19:1825–1835. doi: 10.1101/gr.094482.109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nicolson T. The genetics of hearing and balance in zebrafish. Annu Rev Genet. 2005;39:9–22. doi: 10.1146/annurev.genet.39.073003.105049. [DOI] [PubMed] [Google Scholar]
- Oates AC, Wollberg P, Pratt SJ, Paw BH, Johnson SL, Ho RK, Postlethwait JH, Zon LI, Wilks AF. Zebrafish stat3 is expressed in restricted tissues during embryogenesis and stat rescues cytokine signaling in a STAT1-deficient human cell line. Dev Dyn. 1999;215:352–370. doi: 10.1002/(SICI)1097-0177(199908)215:4<352::AID-AJA7>3.0.CO;2-J. [DOI] [PubMed] [Google Scholar]
- Oxtoby E, Jowett T. Cloning of the zebrafish krox-20 gene (krx-20) and its expression during hindbrain development. Nucleic Acids Res. 1993;21:1087–1095. doi: 10.1093/nar/21.5.1087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parinov S, Kondrichin I, Korzh V, Emelyanov A. Tol2 transposon-mediated enhancer trap to identify developmentally regulated zebrafish genes in vivo. Dev Dyn. 2004;231:449–459. doi: 10.1002/dvdy.20157. [DOI] [PubMed] [Google Scholar]
- Qin HR, Kim HJ, Kim JY, Hurt EM, Klarmann GJ, Kawasaki BT, Duhagon Serrat MA, Farrar WL. Activation of signal transducer and activator of transcription 3 through a phosphomimetic Serine 727 promotes prostate tumorigenesis independent of Tyrosine 705 phosphorylation. Cancer Res. 2008;68:7736–7741. doi: 10.1158/0008-5472.CAN-08-1125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qin Z, Barthel LK, Raymond PA. Genetic evidence for shared mechanisms of epimorphic regeneration in zebrafish. Proc Natl Acad Sci U S A. 2009;106:9310–9315. doi: 10.1073/pnas.0811186106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raz R, Lee CK, Cannizzaro LA, d'Eustachio P, Levy DE. Essential role of STAT3 for embryonic stem cell pluripotency. Proc Natl Acad Sci U S A. 1999;96:2846–2851. doi: 10.1073/pnas.96.6.2846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Riehle KJ, Campbell JS, McMahan RS, Johnson MM, Beyer RP, Bammler TK, Fausto N. Regulation of liver regeneration and hepatocarcinogenesis by suppressor of cytokine signaling 3. J Exp Med. 2008;205:91–103. doi: 10.1084/jem.20070820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roberson DW, Rubel EW. Cell division in the gerbil cochlea after acoustic trauma. Am J Otol. 1994;15:28–34. [PubMed] [Google Scholar]
- Sano S, Itami S, Takeda K, Tarutani M, Yamaguchi Y, Miura H, Yoshikawa K, Akira S, Takeda J. Keratinocyte-specific ablation of Stat3 exhibits impaired skin remodeling, but does not affect skin morphogenesis. EMBO J. 1999;18:4657–4668. doi: 10.1093/emboj/18.17.4657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schebesta M, Lien C, Engel FB, Keating MT. Transcriptional profiling of caudal fin regeneration in zebrafish. ScientificWorldJournal. 2006;6(Suppl 1):38–54. doi: 10.1100/tsw.2006.326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schuck JB, Smith ME. Cell proliferation follows acoustically-induced hair cell bundle loss in the zebrafish saccule. Hear Res. 2009;253:67–76. doi: 10.1016/j.heares.2009.03.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sepich DS, Calmelet C, Kiskowski M, Solnica-Krezel L. Initiation of convergence and extension movements of lateral mesoderm during zebrafish gastrulation. Dev Dyn. 2005;234:279–292. doi: 10.1002/dvdy.20507. [DOI] [PubMed] [Google Scholar]
- Siddiquee K, Zhang S, Guida WC, Blaskovich MA, Greedy B, Lawrence HR, Yip ML, Jove R, McLaughlin MM, Lawrence NJ, Sebti SM, Turkson J. Selective chemical probe inhibitor of Stat3, identified through structure-based virtual screening, induces antitumor activity. Proc Natl Acad Sci U S A. 2007;104:7391–7396. doi: 10.1073/pnas.0609757104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith ME, Coffin AB, Miller DL, Popper AN. Anatomical and functional recovery of the goldfish (Carassius auratus) ear following noise exposure. J Exp Biol. 2006;209:4193–4202. doi: 10.1242/jeb.02490. [DOI] [PubMed] [Google Scholar]
- Soucek S, Michaels L, Frohlich A. Evidence for hair cell degeneration as the primary lesion in hearing loss of the elderly. J Otolaryngol. 1986;15:175–183. [PubMed] [Google Scholar]
- Stone JS, Rubel EW. Delta1 expression during avian hair cell regeneration. Development. 1999;126:961–973. doi: 10.1242/dev.126.5.961. [DOI] [PubMed] [Google Scholar]
- Stone JS, Cotanche DA. Hair cell regeneration in the avian auditory epithelium. Int J Dev Biol. 2007;51:633–647. doi: 10.1387/ijdb.072408js. [DOI] [PubMed] [Google Scholar]
- Taub R. Liver regeneration: from myth to mechanism. Nat Rev Mol Cell Biol. 2004;5:836–847. doi: 10.1038/nrm1489. [DOI] [PubMed] [Google Scholar]
- Thisse B, Thisse C. Fast release clones: a high throughput expression analysis. ZFIN Direct Data Submission. 2004 Available at: http://zfin.org/cgi-bin/webdriver?MIval=aa-pubview2.apg&OID=ZDB-PUB-040907-1. [Google Scholar]
- Torbenson M, Yang SQ, Liu HZ, Huang J, Gage W, Diehl AM. STAT-3 overexpression and p21 up-regulation accompany impaired regeneration of fatty livers. Am J Pathol. 2002;161:155–161. doi: 10.1016/S0002-9440(10)64167-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vollrath MA, Kwan KY, Corey DP. The micromachinery of mechanotransduction in hair cells. Annu Rev Neurosci. 2007;30:339–365. doi: 10.1146/annurev.neuro.29.051605.112917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang H, Park O, Lafdil F, Shen K, Horiguchi N, Yin S, Fu XY, Kunos G, Gao B. Interplay of hepatic and myeloid signal transducer and activator of transcription 3 in facilitating liver regeneration via tempering innate immunity. Hepatology. 2010;51:1354–1362. doi: 10.1002/hep.23430. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wen Z, Zhong Z, Darnell JE., Jr Maximal activation of transcription by statl and stat3 requires both tyrosine and serine phosphorylation. Cell. 1995;82:241–250. doi: 10.1016/0092-8674(95)90311-9. [DOI] [PubMed] [Google Scholar]
- Westerfield M. The zebrafish book: a guide to the laboratory use of the zebrafish (Danio Rerio) Ed 4. Eugene, OR: University of Oregon; 2000. [Google Scholar]
- Williams JA, Holder N. Cell turnover in neuromasts of zebrafish larvae. Hear Res. 2000;143:171–181. doi: 10.1016/s0378-5955(00)00039-3. [DOI] [PubMed] [Google Scholar]
- Woods C, Montcouquiol M, Kelley MW. Math1 regulates development of the sensory epithelium in the mammalian cochlea. Nat Neurosci. 2004;7:1310–1318. doi: 10.1038/nn1349. [DOI] [PubMed] [Google Scholar]
- Wüstefeld T, Rakemann T, Kubicka S, Manns MP, Trautwein C. Hyperstimulation with Interleukin 6 inhibits cell cycle progression after hepatectomy in mice. Hepatology. 2000;32:514–522. doi: 10.1053/jhep.2000.16604. [DOI] [PubMed] [Google Scholar]
- Xiao T, Roeser T, Staub W, Baier H. A GFP-based genetic screen reveals mutations that disrupt the architecture of the zebrafish retinotectal projection. Development. 2005;132:2955–2967. doi: 10.1242/dev.01861. [DOI] [PubMed] [Google Scholar]
- Yamashita S, Miyagi C, Carmany-Rampey A, Shimizu T, Fujii R, Schier AF, Hirano T. Stat3 controls cell movements during zebrafish gastrulation. Dev Cell. 2002;2:363–375. doi: 10.1016/s1534-5807(02)00126-0. [DOI] [PubMed] [Google Scholar]
- Yoshimura A. Regulation of cytokine signaling by the SOCS and Spred family proteins. Keio J Med. 2009;58:73–83. doi: 10.2302/kjm.58.73. [DOI] [PubMed] [Google Scholar]
- Yu H, Pardoll D, Jove R. STATs in cancer inflammation and immunity: a leading role for STAT3. Nat Rev Cancer. 2009;9:798–809. doi: 10.1038/nrc2734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu BM, Ishida Y, Robinson GW, Pacher-Zavisin M, Yoshimura A, Murphy PM, Hennighausen L. SOCS3 negatively regulates the gp130-STAT3 pathway in mouse skin wound healing. J Invest Dermatol. 2008;128:1821–1829. doi: 10.1038/sj.jid.5701224. [DOI] [PubMed] [Google Scholar]