

Abstract
Microorganisms modify rates and mechanisms of chemical and physical weathering and clay growth, thus playing fundamental roles in soil and sediment formation. Because processes in soils are inherently complex and difficult to study, we employ a model based on the lichen–mineral system to identify the fundamental interactions. Fixed carbon released by the photosynthetic symbiont stimulates growth of fungi and other microorganisms. These microorganisms directly or indirectly induce mineral disaggregation, hydration, dissolution, and secondary mineral formation. Model polysaccharides were used to investigate direct mediation of mineral surface reactions by extracellular polymers. Polysaccharides can suppress or enhance rates of chemical weathering by up to three orders of magnitude, depending on the pH, mineral surface structure and composition, and organic functional groups. Mg, Mn, Fe, Al, and Si are redistributed into clays that strongly adsorb ions. Microbes contribute to dissolution of insoluble secondary phosphates, possibly via release of organic acids. These reactions significantly impact soil fertility. Below fungi–mineral interfaces, mineral surfaces are exposed to dissolved metabolic byproducts. Through this indirect process, microorganisms can accelerate mineral dissolution, leading to enhanced porosity and permeability and colonization by microbial communities.
Mineral Weathering, Microbes, and Geochemical Cycles
The Importance of Mineral Weathering.
Rocks at the Earth’s surface typically formed at high temperature and pressure. Exposure of the minerals to oxygenated solutions initiates chemical and physical reactions, resulting in mineral dissolution and crystallization of new phases, such as clays, that are more stable at Earth’s surface conditions (Fig. 1). Nanocrystalline products contribute abundant reactive surface area and thus can impact bioavailability of beneficial and toxic elements (Fig. 2). Weathering affects the compositions of ground water, river and lake water, and ultimately, of oceans (1). Resistant primary and secondary minerals are redistributed to form sediments and soils. Thus, weathering leads to a major geochemical fractionation near the Earth’s surface. Such reactions have occurred throughout geological time, shaping the compositions of the mantle, crust, hydrosphere, and atmosphere. Mineral weathering also directly impacts humans, affecting water quality, agriculture, architectural stability, landscape evolution, integrity of repositories for high level nuclear waste, and distribution of mineral resources.
Figure 1.

A granite weathering profile that passes from almost fresh rock (below) into soil. Characterization of the mineralogy and microbiology of samples from throughout this profile reveals chemical and physical changes that accompany soil formation.
Figure 2.

High-resolution transmission electron microscope (HRTEM) image of nanocrystalline material produced by chemical weathering.
Microbial Distributions in Natural Environments.
Microbes inhabit diverse environments at, and near, the Earth’s surface. Their potential to cause geochemical change is immense. Viable cells exist in extreme environments, from subzero temperatures in Antarctica (2–5) to above boiling temperatures in hot springs and hydrothermal vents (6). Microbes are essentially ubiquitous in sediments, and metabolically active cells have been discovered in rocks buried several kilometers below the Earth’s surface (6–14).
Of the three domains of life, the majority of microorganisms reported to date from soils and sediments are bacteria (11, 15–21). Archaea, minor members of many microbial populations, are especially important under more extreme conditions, such as those encountered in saline lakes (22) and very hot aqueous environments (22–24). Eukaryotes typically occur in more moderate environments, although some fungi and protists thrive at very low pH (refs. 25 and 27; K. J. Edwards, T. M. Gihring, and J.F.B., unpublished data). Recent surveys of subsurface aquifers show large microbial populations, ≈105–108 cells/cm3 (15, 28–30). Cell concentrations ranging from 103 to 109 cells/cm3 have been reported from soils, sediments, and natural waters (15, 31–33). Greater than 108 cells/cm2 of surface area occur on metal sulfide minerals (25–27).
Why Do Microbes Interact with Minerals?
Some biologically essential elements are readily available from natural waters (e.g., Ca, Si, carbonate needed for structural fabrication) (34), but a subset must be actively scavenged (e.g., Fe, K, P). All organisms need Fe, but the solubility of Fe in natural oxygenated near-surface waters is low (35). Some organisms synthesize Fe-specific complexing agents to improve Fe bioavailability (35–41). Other microbes utilize compounds that act as electron shuttles to improve accessibility to redox sites within minerals. Enzyme cofactors such as Mo, Cu, Zn, Mg, Fe, Cr, and Ni can be derived by dissolution of sulfide minerals and ferromagnesian silicates. Phosphorus, required for construction of DNA, RNA, ADP, ATP, phospholipids, and polyphosphates, is available by dissolution of minerals such as apatite [Ca5(PO4)3(F, Cl, OH)] and other typically less soluble secondary phosphates (42–44). A subset of organisms (lithotrophs) associate closely with minerals because they derive their metabolic energy from inorganic substrates (e.g., Mn+2, Fe+2, S, NH4+, and H2) (7, 45–48). Other microbes (heterotrophs) utilize organic material originating from photosynthetic or lithotrophic microorganisms, and compounds such as O2, NO3−, Fe+3, and SO4−2 serve as electron acceptors (45, 46).
Some interactions between cells, organic products, and minerals liberate ions from surfaces. This may be incidental (indirect consequences of growth) or under cellular control. Deciphering these interactions constitutes an important challenge for the future. Regardless of whether microbe–mineral interactions are directed or otherwise, microbial stimulation of mineral dissolution directly affects the fertility of agricultural and other soils.
Microbial Controls on Mineral Weathering Reactions.
Most geochemical research on mineral weathering has focused on inorganic aspects (49). These studies provide valuable information on chemical dissolution rates, mechanisms, and products. Results reveal the order of reactivity of silicate minerals and show how rates depend on temperature, pH, and mineral and solution compositions. The absolute values of rates remain in considerable dispute. Conversely, soil scientists and agronomists have long recognized the fundamental importance of microorganisms in soil development (50).
Understanding of near-surface systems requires integrated mineralogical, geochemical, and biological analysis. A variety of organic metabolic products can dramatically suppress or greatly accelerate rates of dissolution and secondary mineral formation (51–56). Energy generation by catalysis of redox reactions greatly affects element speciation (45, 46). The form of nitrogen is dramatically modified by microbial nitrogen fixation. Organic detritus greatly modifies the water retention capability and physical properties (porosity and permeability) of sediments and soils (57). Conversely, availability of metal oxides, sulfides, and hydroxides able to support lithotrophic growth, redox state, and pH dramatically controls microbial populations. Clay precipitation reactions may provide proton sources or sinks, and a variety of silicate minerals may serve as sources of metal reductive and oxidative power. In this paper, we discuss physical, chemical, and biochemical aspects of mineral dissolution and clay formation. Our approach is to analyze the lichen–mineral microcosm to identify the key factors responsible for microbe–mineral interactions and then to combine mineral and microbial characterization and experimentation to quantify the impact of these factors.
Incipient Weathering and Microbial Colonization.
In the initial stages of alteration, only rock surfaces exposed to air and water are colonized by biofilms. Because of space restrictions, incipient weathering at distance from the biofilm–mineral interface is predominantly inorganic. Inorganic reactions are restricted to sites of fluid access, typically at grain boundaries or in proximity to defects, and to reactions involving readily exchangeable ions (such as interlayer sites in layer silicates) (58–60). Transmission electron microscope investigations show that incipient silicate weathering involves surface hydration, surface recrystallization, and release of ions to solution (60, 61). The scales of elemental redistribution are small, and primary mineral chemical gradients are preserved (62). Surface energy minimization leads to secondary minerals (clays and nanocrystalline oxyhydroxides) in highly specific orientations with respect to the parent mineral (Fig. 3). Thus, mineral surfaces are often coated by clays that largely fill space created by dissolution (63), leaving little room for microbial occupancy. Only after rock density is significantly reduced by dissolution does sufficient (micron-scale) space develop for microbial colonization to proceed.
Figure 3.

High-resolution transmission electron microscope (HRTEM) image showing clays reaction products formed during early weathering of pyroxene develop in highly specific orientations with respect to the parent structure.
Dissolution of reactive phases (e.g., reduced minerals such as sulfides, olivine, and soluble solids such as glass) may facilitate microbial colonization. Metal sulfides, minor components of most crustal rocks (e.g., granites, basalts, metamorphic rocks), may be the first minerals to undergo chemical weathering, leading to generation of acid by the reaction
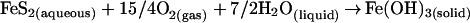 |
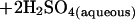 |
The rate of bacterial oxidation of ferrous iron released from pyrite surfaces is up to one million times faster than the inorganic oxidation rate at low pH (64). Because Fe3+ is the predominant pyrite oxidant at low pH, acid generation rates in natural environments are largely determined by microbial activity (65, 66). If sulfide mineral weathering in rocks is microbially mediated, sulfide dissolution can lead directly to colonization by lithotrophic species. Sulfuric acid generated by sulfide dissolution accelerates dissolution of surrounding silicates, increasing porosity available for microbial access and colonization.
What Can We Learn from the Lichen–Mineral Microcosm?
A wide variety of microorganisms colonize mineral surfaces, among the most familiar of which are lichens. Although classically described as symbiotic associations between photosynthetic microorganisms and fungi (67, 68), lichens are actually extremely complex microbial communities. A mass of fungal hyphae, or thallus, composes the majority of any lichen. Photosynthetic microorganisms lie just beneath the upper surface. Although these are typically green algae, other photosynthetic microbes such as diatoms and cyanobacteria occur. This upper zone is a region of carbon transfer, in the form of sugars, between the photosynthetic assemblage and the fungal partner. Lower light levels preclude photosynthesis deeper within the lichen thallus, but other prokaryotes reside among the fungal hyphae. Little is known regarding the biodiversity of this nonphotosynthetic assemblage, and the role these organisms play in nutrient transfer from substratum to fungus is a fertile area for research.
Lichens accelerate the degradation of minerals by physical and chemical methods and are ideal microcosms in which to study microbially mediated mineral weathering. Their extent is easily defined and sampled, and a limited assemblage of minerals grading from fresh to weathered is usually present. Ideally, mineral surfaces of known age (e.g., tombstones, building surfaces, or quarry faces) are studied, permitting field-based weathering rate determinations. Fungal hyphae, perhaps acting in concert with physical weathering mechanisms such as freeze-thaw, penetrate mineral cleavages and grain boundaries, leading to accumulation of substratum-derived mineral fragments within the lower thallus (Fig. 4). The intact organomineral interface from lichen-encrusted boulders in a 90-year-old rock quarry (52) was characterized by transmission electron microscopy. Mineral surfaces in microbially colonized regions are coated in complex mixtures of high molecular weight polymers, clays, and oxyhydroxides. Cells are attached to mineral surface and mineral weathering proceeds via polymer-mediated dissolution/transport/recrystallization. Chemical weathering is accelerated relative to uncolonized surfaces.
Figure 4.

Optical micrograph showing a cross section through the lichen–feldspar interface. Photosynthetic microorganisms (pm) exist within the upper levels of a mass of fungal hyphae (h). A fungal fruiting body (a) is present in this image. Fungal hyphae contribute to physical weathering by penetrating feldspar cleavages and grain boundaries to expose the interior of crystals to microbial colonization.
A zone model for microbially mediated mineral weathering has been developed (Fig. 5) based on correlation of different styles of silicate mineral weathering with pore size-controlled microbial distributions (53). In brief, Zone 1 consists of the upper lichen thallus and is devoid of weathering of substratum-derived mineral particles. Zone 2 is a region of extreme mineral weathering, characterized by direct contact between cells, extracellular polymers, associated compounds, and mineral surfaces. Complex, nanometer-scale mixture of clay products, organic polymers, and primary minerals are common (Fig. 6). In some cases, mineral formation on cell surfaces preserves relict cell shapes. Unlike essentially inorganic weathering, the secondary mineral orientations are less controlled by the primary mineral structure and are more determined by the distribution of polymers onto which they nucleate. Zone 3 weathering reactions, although accelerated by microbial products, are not mediated by direct microbial contact. Orientations of secondary minerals are more commonly determined by orientations of the primary minerals, as for essentially inorganic reactions. Unweathered minerals and minerals undergoing early, predominantly inorganic reactions comprise Zone 4 (Fig. 5).
Figure 5.

Zone model cartoon illustrating mineral weathering occurring in zones that are impacted by microbes to different degrees and in different ways. Zone 4 includes unweathered rock and rock incipiently weathered by inorganic reactions. Zone 3 is where reactions are accelerated by dissolved organic molecules (predominantly acids) but cells are in direct contact with reacting mineral surfaces. Zone 2 is the area of direct contact between microbes, organic products, including polymers, and mineral surfaces. Zone 1 is where photosynthetic members of the symbiosis generate fixed carbon and where crystalline lichen acids precipitate.
Figure 6.

This energy-filtered transmission electron microscope (EFTEM) zero loss image reveals that complex mixtures of organic polymers (p) and clay minerals (c) exist at the lichen–microbe interface.
Application of Insights from the Lichen–Mineral Interface to Soils
Microbial Populations in Soils.
Microbial populations increase in abundance and diversity as rock is weathered and transformed to soil. In parallel, the microstructure and chemical complexity of the system increases, making high-resolution studies of processes occurring in soils extremely difficult. For this reason, the lichen–rock interface is a preferable system for study. Insights from the lichen system then must be tested for their applicability to soils.
A diverse community, comprised of extremely high numbers of symbiotic and nonsymbiotic microorganisms, inhabit the soil zone around the roots of vascular plants. Although microbial concentrations in soils are very high, ≈109 cells/cm3, only ≈1–10% of the total, are alive (29). Thus, viability may ultimately prove to be a more important parameter than diversity or overall cell counts for mineral weathering studies.
The assemblage of plant roots and their associated microflora and fauna in intimate contact with soil particles is termed the rhizosphere. Because many studies reveal the importance of microbe–mineral interactions to plant nutrition (69), it is critical to understand the rates and mechanisms of mineral transformations occurring in the rhizosphere.
Which Microorganisms Are Involved?
Microbial populations in soils are very diverse. One gram of soil can contain anywhere from hundreds to many thousands of species of microorganisms (16–21, 70, 71). Furthermore, the level of diversity from agricultural soils reported in molecular biological studies is astonishing.
Identification of individual microbial cells in natural samples minerals remained challenging until recently. Traditional microbial identification techniques relied on culturing and isolation of microbial cells from a natural sample and then characterization of microbial isolates via a series of biochemical tests. This approach gives very biased population data because, although a large fraction of the visible cells are viable, only 0.01–10% of microorganisms are culturable by using current methods. Although these techniques yield some information on microbial populations and diversity, all information on the spatial distribution of organisms is lost. Recent breakthroughs in molecular biological approaches have provided powerful approaches to study the distribution and diversity of microbial cells in situ (71, 72, 73).
The basis of the molecular approach to microbial population analysis is to extract, amplify, purify, and sequence DNA, typically the ribosomal DNA, from a natural microbial population and then to identify individual organisms by comparison between the obtained sequences and sequences from known organisms. Regardless of whether an organism has been identified previously, differences between its sequence and those of known organisms can be used to place the species onto a phylogenetic tree (71–73). Once the sequence has been obtained, a DNA-specific, fluorescently labeled probe can be used to identify individual cells in environmental samples or in laboratory cultures (25–27, 71, 73, 74). These probes are designed so that they will bind only to ribosomal RNA of organisms with complementary sequence (71, 73). Under appropriate experimental conditions, the degree of probe specificity (e.g., binds to all bacteria versus binds to Thiobacillus caldus) is determined by choice of probe sequence (e.g., an oligonucleotide common to all bacteria versus one common to only the target species). Microscopic visualization using multiple probes allows direct in situ characterization of the microbial population at the species (or higher) level.
Combined DNA sequencing and probing has been used successfully in a number of natural environments (25–27, 73, 74). In some cases, it has been possible to quantify the proportions of bacteria, archaea, and eukaryotes and to correlate abundances with geochemical conditions. This has provided new insights into interconnections between physical, chemical, and biological components of natural systems.
Analysis of DNA from environmental samples and characterization of environmental samples using probes constructed with DNA sequences of interest are powerful new approaches for geochemical studies. These tools have not yet been deployed widely but should allow future researchers to evaluate interrelationships between microbial ecology and geochemical and physical parameters (25). Ultimately, these data can be correlated with metabolic information to infer how and why microbes interact with mineral surfaces in natural environments.
DNA has been extracted and sequenced from several hundred soil microorganisms (16–21, 70). Most of the sequences were from bacteria, but many did not correspond to any of the known species. Several could not be classified into any of the larger known bacteria groups. Recently, archaea have been consistently reported from the rhizosphere. The distribution and importance of these organisms remain unclear.
Experimental Quantification of Processes Occurring in Lichen Zones: Insights for Rhizosphere Processes.
Plant roots excrete a combination of sugars, organic acids, and amino acids. It is believed that plant roots exude carbohydrates to encourage and sustain the associated microbial community. This large-scale carbon transfer from photosynthesizer to a nonphotosynthetic microbial community is analogous to Zone 1 in Barker and Banfield’s (53) microbial weathering model (Fig. 5).
The plant–microbe symbiosis is not the only similarity between the rhizosphere and lichen communities. Within soils, some microorganisms live in solution, but most are attached to mineral surfaces where they can directly affect mineral reactions (75, 76). Attached cells, largely fungi and bacteria, may impact mineral dissolution, precipitation, and clay hydration reactions in a manner analogous to that seen in Zone 2, at the lichen–mineral interface (Fig. 5).
Physical restrictions placed on microbial distribution by pore size also apply to rhizosphere environments. Consequently, Zone 3 (Fig. 5) reactions are expected in soils as well. Although only the outer surface of a given mineral grain may be colonized, interior surfaces may show enhanced dissolution because of organic acids of plant and microbial origin. Elevated carbonic acid levels may result from fungal and bacterial degradation of organic matter (77, 78). Several to several hundred micromolar concentrations of oxalate, acetate, lactate, formate, pyruvate, propionate, malate, succinate, citrate, isocitrate, and aconitate have been detected in rhizosphere soil (79–81).
The fertility of numerous soils is limited by the abundance of bioavailable phosphate. In incipiently weathered rocks, microbial growth also may be phosphorus-limited because the abundance of organophosphates is low. Phosphorus limitation can develop during weathering because of either phosphorus adsorption onto secondary iron oxyhydroxide surfaces or binding of phosphorus into insoluble secondary minerals.
During the initial stages of weathering, apatite is replaced by chemically or microbially precipitated secondary phosphate minerals containing iron (strengite), aluminum (variscite), and lanthanides (e.g., rhabdophane and florencite; Fig. 7). Secondary lanthanide phosphates persist until at least one-third of the initial mineral constituents are removed by dissolution (82). These phosphates are completely solubilized in the soil zone (83). DNA staining, in combination with high-resolution scanning electron microscope observations and unpublished experimental data, suggest that solubilization of secondary phosphates results from microbial colonization (Fig. 8). Microbes apparently respond to phosphate limitation by locating themselves at sites of inorganic phosphorus release or by excretion of organic acids or other complexing compounds to actively dissolve the otherwise insoluble secondary phosphate minerals (5, 43, 84). Further work is needed to determine which organic compounds are important. The effectiveness of likely compounds should be tested experimentally. Analysis of organic constituents in phosphorus-stressed cultures could be used to verify the microbial response. Identification of the mechanisms involved and microorganisms responsible for release of phosphorus from resistant secondary phosphates may provide new strategies for enhancement of productivity of low fertility soils (e.g., how to enhance the effectiveness of phosphate fertilizers and to manage soil fertility through manipulation of microbial populations).
Figure 7.

Secondary phosphate minerals formed on the surface of apatite during the early stages of weathering. These insoluble phases bind P in a relatively nonbioavailable form.
Figure 8.

Microorganisms colonize surfaces of secondary minerals in pits formed by apatite dissolution. Microbial processes mobilize phosphorus from insoluble secondary phases.
Dissolution of inorganic phosphates may localize microbial activity and stimulate weathering of adjacent minerals. For example, it has been shown (85) that microbially mediated feldspar dissolution rates increase when feldspars contain apatite inclusions. Because feldspar dissolution rates are strongly pH-dependent, it is probable that the results can be attributed to release of acidic microbial byproducts.
Mineral dissolution studies with cultures of bacteria and fungi show a dramatic increase in dissolution rates of feldspar, biotite (Fig. 9a), quartz, apatite, and other minerals (54, 86–93). In experiments with bacteria, mica, and feldspar, there is a direct correlation between microbial organic ligand production and increased release of Si, Al, and Fe (88–90). Although these experiments replicate natural systems to some extent, they are inherently complex because of the combination of acidity effects, ion-complexing effects, and growth media effects. Thus, it is difficult to distinguish the contributing factors.
Figure 9.

(Left) Fe released from biotite to solution in three experiments, two of which used bacterial cultures (B0428 and B0665) and one that was a control experiment. Significant enhancement of biotite dissolution rate is observed. (Right) Results of a feldspar dissolution experiment (pH 4.0) using undifferentiated and medium molecular weight polysaccharides demonstrate enhanced feldspar dissolution by two to three orders of magnitude under some conditions.
Lichen–mineral interface characterization studies suggest that microbial colonization can dramatically impact both the rates and the mechanisms of silicate mineral weathering reactions via direct and indirect processes. To compare the effectiveness of organic–mineral interactions that depend on contact between microbes and mineral surfaces with those interactions that only involve soluble compounds, it is necessary to develop appropriately simplified experimental models.
Acid production is the most basic mechanism by which microbes affect weathering reactions. We infer that this is largely responsible for enhanced reactivity in Zone 3 (Fig. 5). Solutions of organic acids in concentrations comparable to or slightly higher than ground water show increase in dissolution rates of less than one order of magnitude (56, 57). These relatively small effects may hide much larger responses in natural systems in which local microenvironments may be characterized by very high acid concentrations because of cell proximity.
It is possible to demonstrate directly that microbes can cause low pH microenvironments at mineral surfaces. The pH values can be measured by using microelectrodes (94) or, in even smaller volumes, by using confocal-based techniques. For example, pH values of 3–4 were detected in proximity to bacterial cells within cleavages in biotite when the bulk solution pH was 7.0 (88). As acidity increases, below pH = 5, the rates of silicate mineral dissolution increase by a factor of aH+n (95). Lowering pH to 3–4 corresponds to a 10- to 1,000-fold increase in dissolution rate. The mechanism of the reaction changes as well. Typically, elements such as Fe and Al are relatively insoluble at neutral pH and form secondary Fe-hydroxides and vermiculite or smectite-like silicate clays. However, as acidity increases, Fe and Al solubility and mobility increase, probably leading to formation of different secondary minerals, such as the aluminosilicates kaolinite and halloysite.
In addition to inorganic acid production, microbes also can catalyze mineral weathering rates by production of organic ligands. Ligands can complex with ions on the mineral surface and can weaken metal–oxygen bonds. Alternatively, ligands indirectly affect reactions by forming complexes with ions in solution, thereby decreasing solution saturation state. Experimental studies using relatively dilute solutions of compounds such as oxalic acid, citric acid, pyruvate, α-ketoglutarate, acetate, propionate, lactate, etc. have shown rate enhancements for silicate dissolution of up to one order of magnitude (96–101). The effect is somewhat similar to that of acid production because organic ligands affect silicate mineral dissolution stoichiometry by complexing with, and increasing the solubility of, less soluble major ions such as Al and Fe (96, 102, 103).
The transformation of biotite to vermiculite with the release of the interlayer K is perhaps one of the most important biologically mediated geochemical reactions occurring in the rhizosphere (104–106). Uptake of K+ by microbial cells and plant roots lowers solution saturation state, thereby indirectly promoting the weathering. However, plants and associated ectomycorrhizal populations have more aggressive methods for weathering sheet silicates. The extensive transformation of biotite to vermiculite has been attributed to root-induced pH decreases in the rhizosphere and acid dissolution of the mica structure (104). In a similar experimental study (93), plant roots and ectomycorrhizal fungi actively produced oxalate and increased biotite weathering when they were stressed for K and Mg. Plant roots and associated microbial populations also physically disrupt sheet silicates, exposing new surface area for chemical alteration (107).
It has been shown that microorganisms produce intermediate molecular weight organic compounds that are important in iron transport to the cell surface (36, 37). Under conditions of Fe limitation, microbes produce siderophores that have a very high affinity for Fe+3. Siderophores form very strong stable bidentate complexes with octahedrally coordinated Fe+3 as well as with other major elements in minerals, e.g., Al and Si. Several hundred siderophores from microbial cultures and natural environments have been isolated and identified, most of which have either hydroxamate or catecholate functional groups. Estimates of siderophore concentrations in soil microenvironments range from 10 to 2,000 mg/liter (38, 39). Mineral weathering experiments with naturally occurring siderophores show that these compounds can accelerate the rate of Fe-oxide and silicate mineral dissolution by about one order of magnitude and thereby dramatically impact Fe cycling in soils (35, 38, 39, 41).
In addition to producing acids and low and intermediate molecular weight organic ligands, microbes also produce high molecular weight polymers. Observations of naturally weathered minerals associated with lichens show that, in the direct zone, these polymers coat surfaces. Polymers can affect mineral weathering reactions by several processes (52, 53). Slime layers are ≈99% water, so they can increase the contact time between water and the mineral surface. Polymers also increase the diffusion of ions away from the mineral surface. However, these extracellular polymers may have more direct chemical effects on mineral weathering as well.
The effect of high molecular weight organic molecules on mineral dissolution rates was quantified by using dissolution experiments involving model extracellular polymers. The model polysaccharides (alginates) varied in the ratio of constituent mannuronic and guluronic acids. Mannuronic and guluronic acids differ in their effect because of different concentrations and orientations of carboxylic acids functional groups. Like low molecular weight organic acids, polysaccharides can increase the extent of mineral weathering (Fig. 9b), presumably by complexing with ions in solution, thereby lowering solution saturation state (51, 108).
Microbially induced metal binding and mineralization is important in the rhizosphere as well as at the lichen–mineral interface. Cell surfaces and associated polymers provide effective nucleation sites for secondary silicate mineral precipitation. Some rocks contain high abundances of minerals enriched in potentially toxic elements (e.g., As, Cd, Cu, U). Microorganisms react to high metal concentrations in a variety of ways. One biochemical response to toxic substances is production of extracellular polymers that bind and effectively immobilize the compound and, in some cases, biomineralization. For example, microbes immobilize U by intra- and extracellular precipitation of secondary minerals (109, 110). Alternatively, some microbes exposed to arsenic employ mobile genetic structures (plasmids) to manufacture proteins that (although counterintuitive) reduce the less toxic As5+ to more toxic As3+ and secrete it from their cells (111). In other cases, microbes can completely volatilize the toxic element (e.g., by formation of methyl-mercury compounds) (111). Thus, microbial responses to toxic metals can dramatically change metal abundances and elemental speciation.
Applications of Microbe-Mineral Interaction Studies for the 21st Century.
Among the many challenges for the 21st century are questions concerning how (i) to maintain agricultural soil fertility to ensure food production for a still-growing world population; (ii) to prevent further environmental damage; and (iii) to invent cost-effective ways to remediate existing contamination of soil, sediments, and water. Improved knowledge of how microorganisms interact with their environments and contribute to geochemical transformations will be critical to these endeavors.
We have established above that microbes are important agents of physical and chemical change in natural systems. In fact, global-scale models verify that chemical weathering reactions directly impact climate. It is widely accepted that, early in Earth’s history, the atmosphere was dominated by CO2 and that oxygen concentrations only increased with the evolution of efficient photosynthetic microbial populations. Early inorganic rock weathering resulted in accumulation of Ca and Mg in ocean waters, leading to precipitation of Ca, Mg-carbonates and draw down of CO2. The rates of chemical weathering of Ca-silicates, Mg-silicates, and Ca, Mg-silicates determine the rate of supply of Ca and Mg to oceans and thus affect the magnitude of this critical feedback mechanism (112–116). Mineral dissolution rates are important inputs into global climate models (112–115). However, the impact of microbial processes on weathering reactions over geological time are unclear, and the relevant rates remain in dispute. If microorganisms significantly affect mineral dissolution rates, then the evolution of the biosphere (dominated by microorganisms for the majority of time), atmosphere, hydrosphere, and lithosphere have been closely coupled (117). Much can be learned about the long term response of the Earth system through quantifying the microbial–mineralogical feedback mechanisms. Thus, a challenge for the next century will be to develop more rigorous and comprehensive models for climate change.
Climate change implies altered patterns of rainfall and temperature and thus, modification of the processes occurring in surficial materials, including soils. Prediction of the consequence of climate change requires a rather comprehensive understanding of the interrelationships between mineralogy, soil chemistry, microbial populations, rainfall, and temperature. Such understanding will benefit from investigations of soil-forming mineral reactions occurring under diverse climatic and hydrological conditions. Analyses must include detailed characterization of primary minerals and their weathering products, a census of microorganisms and determination of their patterns of distribution, and contextural information about soil physics and chemistry.
It should be possible to increase the rates of soil development and optimize methods used to maintain or increase soil fertility through knowledge about microbial populations and how microbes cooperate to affect mineral dissolution, degradation of organic compounds, immobilization of ions, precipitation of minerals, and change in solution chemistry. For example, identification of microbial species capable of solubilizing secondary phosphates may provide new insights for improved fertilizer efficiency (e.g., simultaneous addition of microorganisms or microbial compounds with phosphate fertilizers).
Understanding of microbial metal tolerance and of metal resistance strategies should lead to new approaches to environmental cleanup. Understanding how microbes affect the fate of contaminants in the natural environment will be critical for waste treatment and site remediation. An example of current regulatory interest is the long term, near-site containment of nuclides released from geological repositories. Challenges such as this demand sophisticated models that include all factors, inorganic and biological. Interdisciplinary study of microbe-mineral-solution interactions are required for model development.
As we learn about the remaining 99–99.9% of organisms we currently know essentially nothing about, new species with new capabilities will be discovered. These may be vitally important sources of new compounds of tremendous technological importance. The discovery of hyperthermophilic microorganisms by Thomas Brock (University of Wisconsin, Madison, WI) (26) led to the identification of a thermally stable enzyme, taq polymerase, which has revolutionized the PCR step of DNA analysis. It is difficult to predict what the future will hold, but the discovery of enzymes capable of carrying out redox reactions at silicate and oxide mineral surfaces as well as in solutions may be of some importance for enhancement of soil fertility, economic metal extraction, and materials science.
The global-scale balance between inorganic and microbial process is unclear, and the quantitative effects of microbes on geochemical cycles are essentially a mystery. At present, we are identifying specific processes that may be important and are quantifying them via laboratory studies. Ecological aspects of microbially dominated systems, including the symbioses and competition, largely remain a challenge for the future. New understanding of Earth’s surface processes should come from detailed and quantitative studies that utilize current molecular biological and biochemical approaches in concert with high-resolution mineralogical and geochemical analyses.
Acknowledgments
The authors acknowledge the editorial assistance of Dr. J. V. Smith (University of Chicago) and thank Dr. R. A. Eggleton (Australian National University), K. Edwards (University of Wisconsin, Madison, WI), T. Gihring (University of Wisconsin, Madison, WI), and Dr. P. Bond (University of Wisconsin, Madison, WI) for their contributions to this work. This research was supported by Grants EAR-9317082 and EAR-9706382 from the National Science Foundation and Grant DE-FG02-93ER14328 from the Department of Energy.
References
- 1.Banfield J F, Nealson K H. Rev Mineral. 1997;35:1. [Google Scholar]
- 2.Vincent W F, James M R. Biodivers Conserv. 1996;5:1451. [Google Scholar]
- 3.Vishniac H S. Biodivers Conserv. 1996;5:1356. [Google Scholar]
- 4.Broady P A. Biodivers Conserv. 1996;5:1307. [Google Scholar]
- 5.Johnston C G, Vestal J R. Microb Ecol. 1993;25:305. doi: 10.1007/BF00171895. [DOI] [PubMed] [Google Scholar]
- 6.Kerr R A. Science. 1997;276:703. [PubMed] [Google Scholar]
- 7.Stevens T O, McKinley J P. Science. 1995;270:450. [Google Scholar]
- 8.Stevens T O, McKinley J P, Fredrickson J K. Microb Ecol. 1993;25:35. doi: 10.1007/BF00182128. [DOI] [PubMed] [Google Scholar]
- 9.Onstott T C, Tseng H Y, Phelps T J, Colwell F S. Abstr Programs Geol Soc Am. 1995;27:6. , 312 (abstr.). [Google Scholar]
- 10.Phelps T J, Raione E G, White D C, Fliermans C B. Geomicrobiol J. 1989;7:79. [Google Scholar]
- 11.Balkwill D, Boone D R, Colwell F S, Griffin T, Kieft T L, Lehman R M, McKinley J P, Nierzwicki-Bauer S, Onstott T C, Tseng H Y, et al. Eos Trans Am Geophys Union. 1994;75:34. [Google Scholar]
- 12.Ghiorse W C. Science. 1997;275:789. [Google Scholar]
- 13.Fredrickson J K, Onstott T C. Sci Am. 1996;275:68. doi: 10.1038/scientificamerican1096-68. [DOI] [PubMed] [Google Scholar]
- 14.Fyfe W S. Science. 1996;273:448. [Google Scholar]
- 15.Balkwill D J. Geomicrobiol J. 1989;7:33. [Google Scholar]
- 16.Borneman J, Skroch P W, O’Sullivan K M, Palus J A, Rumjanek N G, Jansen J L, Nienhuis J, Triplett E W. Appl Environ Microbiol. 1996;62:1935. doi: 10.1128/aem.62.6.1935-1943.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Borneman J, Triplett E W. Appl Environ Microbiol. 1997;63:2647. doi: 10.1128/aem.63.7.2647-2653.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Kennedy A C, Gewin V L. Soil Sci. 1997;162:607. [Google Scholar]
- 19.Torsvik V, Sorheim R, Goksoyr J. J Ind Microbiol Biotechnol. 1996;17:170. [Google Scholar]
- 20.Rheims H, Rainey F A, Stackebrandt E. J Ind Microbiol Biotechnol. 1996;17:159. [Google Scholar]
- 21.Franzmann P D. Biodivers Conserv. 1996;5:1295. [Google Scholar]
- 22.Zimmer C. Discover. 1995;16:30. [Google Scholar]
- 23.Barns S M, Fundyga R E, Jeffries M W. Proc Natl Acad Sci USA. 1994;91:1609. doi: 10.1073/pnas.91.5.1609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Barns S M, Nierzwicki-Bauer S A. Rev Mineral. 1997;35:35. [Google Scholar]
- 25.Edwards, K. J., Goebel, B. M., Rodgers, T. M., Schrenk, M. O., Gihring, T. M., Cardona, M. M., Hu, B., McGuire, M. M., Hamers R. J., Pace, N. R., et al. (1999) Geomicrobiol. J., in press.
- 26.Brock T D. Thermophilic Microorganisms and Life at High Temperatures. New York: Springer; 1978. [Google Scholar]
- 27.Edwards K J, Schrenk M O, Hamers R J, Banfield J F. Am Mineral. 1998;83:1444. [Google Scholar]
- 28.Sinclair J L, Ghiorse W C. Geomicrobiol J. 1989;7:15. [Google Scholar]
- 29.Bone T L, Balkwill D L. Microb Ecol. 1988;16:49. doi: 10.1007/BF02097404. [DOI] [PubMed] [Google Scholar]
- 30.Fredrickson J K, Garland T R, Hicks R J, Thomas J M, Li S W, McFadden K M. Geomicrobiol J. 1989;7:53. [Google Scholar]
- 31.Albrechtsen H-J, Winding A. Microb Ecol. 1992;23:303. doi: 10.1007/BF00164102. [DOI] [PubMed] [Google Scholar]
- 32.Kampfer P, Steiof M, Dott W. Microb Ecol. 1991;21:227. doi: 10.1007/BF02539156. [DOI] [PubMed] [Google Scholar]
- 33.Hicks R J, Fredrickson J K. Geomicrobiol J. 1989;7:67. [Google Scholar]
- 34.de Vrind-de Jong E W, de Vrind J P M. Rev Mineral. 1997;35:267. [Google Scholar]
- 35.Stone A T. Rev Mineral. 1997;35:309. [Google Scholar]
- 36.Neilands J B. Annu Rev Biochem. 1981;50:715. doi: 10.1146/annurev.bi.50.070181.003435. [DOI] [PubMed] [Google Scholar]
- 37.Neilands J B. Annu Rev Microbiol. 1982;36:285. doi: 10.1146/annurev.mi.36.100182.001441. [DOI] [PubMed] [Google Scholar]
- 38.Hersman L, Maurice P, Sposito G. Chem Geol. 1996;132:25. [Google Scholar]
- 39.Hersman L, Lloyd T, Sposito G. Geochim Cosmochim Acta. 1995;59:3327. [Google Scholar]
- 40.Ochs M, Brunner I, Stumm W, Cosovic B. Water Air Soil Pollution. 1993;68:213. [Google Scholar]
- 41.Watteau F, Berthelin J. J Soil Biol. 1994;30:1. [Google Scholar]
- 42.Jansson M. Microb Ecol. 1987;14:81. doi: 10.1007/BF02011573. [DOI] [PubMed] [Google Scholar]
- 43.Leyval C, Berthelin J. Plant Soil. 1989;117:103. [Google Scholar]
- 44.Cumming J R. Tree Physiol. 1993;13:173. doi: 10.1093/treephys/13.2.173. [DOI] [PubMed] [Google Scholar]
- 45.Chapelle F H. Ground Water Microbiology and Geochemistry. New York: Wiley; 1993. [Google Scholar]
- 46.Lovely D R, Chapelle F H. Rev Geophys. 1995;33:365. [Google Scholar]
- 47.Ehrlich H L. Chem Geol. 1996;132:1. [Google Scholar]
- 48.Berthelin J. In: Microbial Geochemistry. Krumbein W E, editor. Oxford: Blackwell Scientific; 1983. p. 223. [Google Scholar]
- 49.White A F, Brantley S L. Rev Mineral. 1995;31:1. [Google Scholar]
- 50.Huang P M, Schnitzer M, editors. Interactions of Soil Minerals with Natural Organics and Microbes. Madison, WI: Soil Science Society of America; 1986. [Google Scholar]
- 51.Welch, S. A., Barker, W. W. & Banfield, J. F. (1999) Geochim. Cosmochim. Acta, in press.
- 52.Barker W W, Banfield J F. Chem Geol. 1996;132:55. [Google Scholar]
- 53.Barker W W, Banfield J F. Geomicrobiol J. 1998;15:223. [Google Scholar]
- 54.Ullman W J, Kirchman D L, Welch S A, Vandevivere P. Chem Geol. 1996;132:11. [Google Scholar]
- 55.Fortin D, Ferris F G, Beveridge T J. Rev Mineral. 1997;35:161. [Google Scholar]
- 56.Drever J I. Geochim Cosmochim Acta. 1994;58:2325. [Google Scholar]
- 57.Drever J I, Vance G F. In: The Role of Organic Acids in Geological Processes. Lewan M D, Pittman E D, editors. New York: Springer; 1994. p. 138. [Google Scholar]
- 58.Banfield J F, Eggleton R A. Clays Clay Miner. 1988;36:47. [Google Scholar]
- 59.Banfield J F, Eggleton R A. Clays Clay Miner. 1990;38:77. [Google Scholar]
- 60.Hochella M, Banfield J F. Rev Mineral. 1995;30:353. [Google Scholar]
- 61.Banfield J F, Veblen D R, Jones B F. Contrib Mineral Petrol. 1990;106:110. [Google Scholar]
- 62.Banfield J F, Barker W W. Geochim Cosmochim Acta. 1994;58:1419. [Google Scholar]
- 63.Banfield J F, Jones B J, Veblen D R. Geoochim Cosmochim Acta. 1991;55:2781. [Google Scholar]
- 64.Singer P C, Stumm W. Science. 1970;167:1121. doi: 10.1126/science.167.3921.1121. [DOI] [PubMed] [Google Scholar]
- 65.Norstrom D K, Southam G. Rev Mineral. 1997;35:360. [Google Scholar]
- 66.Nordstrom, D. K. & Alpers, N. (1999) Rev. Econ. Geol.6, in press.
- 67.Hale M E. The Biology of Lichens. London: Edward Arnold; 1983. p. 180. [Google Scholar]
- 68.Nash T H. Lichen Biology. New York: Cambridge Univ. Press; 1996. p. 303. [Google Scholar]
- 69.Jungk A O. Plant Roots: The Hidden Half. New York: Dekker; 1996. p. 529. [Google Scholar]
- 70.Bintrim S B, Donohue T J, Handelsman J, Roberts G P, Goodman R M. Proc Natl Acad Sci USA. 1997;94:277. doi: 10.1073/pnas.94.1.277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Amann R, Snaider J, Wagner M. J Bacteriol. 1996;178:3496. doi: 10.1128/jb.178.12.3496-3500.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Pace R N, Stahl D A, Lane D J, Olsen G J. Adv Microbiol Ecol. 1986;9:1. [Google Scholar]
- 73.Amman R I, Ludwig W, Schleifer K-H. Microbiol Rev. 1995;59:143. doi: 10.1128/mr.59.1.143-169.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Schrenk M O, Edwards K J, Goodman R M, Hamers R J, Banfield J F. Science. 1998;279:1519. doi: 10.1126/science.279.5356.1519. [DOI] [PubMed] [Google Scholar]
- 75.Holm P E, Nielsen P H, Albrechtsen H-J, Christensen T A. Appl Environ Microbiol. 1992;58:3020. doi: 10.1128/aem.58.9.3020-3026.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Hazen T C, Jimenez L, de Victoria G L. Microb Ecol. 1991;22:293. doi: 10.1007/BF02540231. [DOI] [PubMed] [Google Scholar]
- 77.Chapelle F H, Zelibor J L, Jr, Grimes D Y, Knobel L L. Water Resources Res. 1987;23:1625. [Google Scholar]
- 78.Chapelle F H, Lovely D R. Appl Environ Microbiol. 1990;56:1865. doi: 10.1128/aem.56.6.1865-1874.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Fox T R, Comerford N B. Soil Sci Soc Am J. 1990;54:1139. [Google Scholar]
- 80.Shen Y, Strîm L, Jînsson J, Tyler G. Soil Biol Biochem. 1996;28:1163. [Google Scholar]
- 81.Grierson P F. Plant Soil. 1992;144:259. [Google Scholar]
- 82.Banfield J F, Eggleton R A. Clays Clay Miner. 1989;37:113. [Google Scholar]
- 83.Taunton A E, Welch S A, Santelli C M, Fournelle J, Banfield J F. Geological Society of America Meeting: Abstracts with Programs. Boulder, CO: Geological Society of America; 1998. , A304. [Google Scholar]
- 84.Illmer P A, Barbato A, Schinner F. Soil Biol Biochem. 1995;27:265. [Google Scholar]
- 85.Rodgers J R, Bennett P C, Hiebert F K. Geological Society of America Meeting: Abstracts with Programs. Boulder, CO: Geological Society of America; 1998. , A305. [Google Scholar]
- 86.Barker W W, Welch S A, Banfield J F. Rev Mineral. 1997;35:391. [Google Scholar]
- 87.Berthelin J, Belgy G. Geoderma. 1979;21:297. [Google Scholar]
- 88.Barker W W, Welch S A, Chu S, Banfield J F. Am Mineral. 1998;83:1551. [Google Scholar]
- 89.Welch, S. A. & Ullman W. J (1999) Chem. Geol., in press.
- 90.Vandevivere P, Welch S A, Ullman W J, Kirchman D L. Microb Ecol. 1994;27:241. doi: 10.1007/BF00182408. [DOI] [PubMed] [Google Scholar]
- 91.Thorseth I H, Furnes H, Tumyr O. Chem Geol. 1995;119:139. [Google Scholar]
- 92.Callot G, Maurette M, Pottier L, Dubois A. Nature (London) 1987;328:147. [Google Scholar]
- 93.Paris F, Bottom B, Lapeyrie F. Plant Soil. 1996;179:141. [Google Scholar]
- 94.Parasuraman C S. Dissertation. Newark: Univ. of Delaware; 1995. [Google Scholar]
- 95.Blum A, Lasaga A C. Nature (London) 1988;31:431. [Google Scholar]
- 96.Welch S A, Ullman W J. Geochim Cosmochim Acta. 1993;57:2725. [Google Scholar]
- 97.Welch S A, Ullman W J. Geochim Cosmochim Acta. 1996;60:2939. [Google Scholar]
- 98.Stillings L L, Drever J I, Brantley S L, Sun Y, Oxburgh R. Chem Geol. 1996;132:79. [Google Scholar]
- 99.Bennett P C, Melcer M E, Siegel D I, Hassett J P. Geochim Cosmochim Acta. 1988;52:1521. [Google Scholar]
- 100.Hansley P L. J Sediment Petrol. 1987;57:666. [Google Scholar]
- 101.Manley E P, Evans L J. Soil Sci. 1986;141:106. [Google Scholar]
- 102.Wogelius R A, Walther J V. Geochim Cosmochim Acta. 1991;55:943. [Google Scholar]
- 103.Antweiler R C, Drever J I. Geochim Cosmochim Acta. 1983;47:623. [Google Scholar]
- 104.Hinsinger P, Elsass F, Jaillard B, Robert M. J Soil Sci. 1993;44:535. [Google Scholar]
- 105.Hinsinger P, Jaillard B. J Soil Sci. 1993;44:525. [Google Scholar]
- 106.Berthelin J, Leyval C. Plant Soil. 1982;68:369. [Google Scholar]
- 107.April R, Keller D. Biogeochemistry. 1990;9:11. [Google Scholar]
- 108.Welch S A, Vandevivere P. Geomicrobiol J. 1995;12:227. [Google Scholar]
- 109.Macaskie L E, Yong P, Doyle T C, Roig M G, Diaz M, Manzano T. Biotechnol Bioeng. 1997;53:100. doi: 10.1002/(SICI)1097-0290(19970105)53:1<100::AID-BIT13>3.0.CO;2-S. [DOI] [PubMed] [Google Scholar]
- 110.Jeong B Y, Hawes C, Bonthrone K M, Macaskie L E. Microbiology. 1997;143:2497. doi: 10.1099/00221287-143-7-2497. [DOI] [PubMed] [Google Scholar]
- 111.Silver S. Rev Mineral. 1997;35:345. [Google Scholar]
- 112.Walker C G, Hays P B, Kasting J F. J Geophys Res. 1981;86:9776. [Google Scholar]
- 113.Berner R A, Lasaga A C, Garrels R M. Am J Sci. 1983;283:641. [Google Scholar]
- 114.Volk T. Am J Sci. 1987;287:763. [Google Scholar]
- 115.Brady P V. J Geophys Res. 1991;96:18101. [Google Scholar]
- 116.Brady P V, Carroll S A. Geochim Cosmochim Acta. 1994;58:1853. [Google Scholar]
- 117.Schwartzman D W, Volk T. Paleogeogr Paleoclimatol Paleoecol. 1991;90:357. [Google Scholar]