

Abstract
Transient receptor potential vanilloid subfamily member 1 (TRPV1) is activated by capsaicin, acid, and heat and mediates pain through peripheral nerves. In the tongue, TRPV1 expression has been reported also in the epithelium. This indicates a possibility that sensation is first received by the epithelium. However, how nerves receive sensations from the epithelium remains unclear. To clarify the anatomical basis of this interaction, we performed immunohistochemical studies in the rodent tongue to detect TRPV1 and calcitonin gene-related peptide (CGRP), a neural marker. Strong expression of TRPV1 in the epithelium was observed and was restricted to the apex of the tongue. Double immunohistochemical staining revealed that CGRP-expressing nerve terminals were in close apposition to the strongly TRPV1-expressing epithelium of fungiform papilla in the apex of rodent tongues. These results suggest that the TRPV1-expressing epithelium monitors the oral environment and acquired information may then be conducted to the adjacent CGRP-expressing terminals.
Key words: TRPV1, tongue, peripheral nerve, epithelium, burning mouth syndrome, capsaicin.
Introduction
The tongue, as a sensory organ, plays an important role in taste reception. It has been reported that capsaicin, the active substance in chili peppers contributes to the pungent taste and causes burning sensation or burning pain.1 It is believed that the capsaicin receptor, transient receptor potential vanilloid subfamily member 1 (TRPV1), is expressed in the tongue, which mediates burning sensation or burning pain through communication from the peripheral nervous system to the central nervous system.2,3 TRPV1 is activated not only by capsaicin, but also by acid and heat (>43°C) and is expressed in the trigeminal ganglion and spinal ganglion, which have unmyelinated C-fibers. Therefore, it is considered that the physiology of reception and transmission through C-fibers related to pain and heat corresponds to the site of expression.2,3
Expression of TRPV1 in the peripheral nervous system has already been reported. TRPV1 is expressed in the main organs of the whole body, the trigeminal nervous system, skin, digestive organs (stomach and intestines), and respiratory organs (trachea and lungs).4–7 Oral TRPV1 expressions have been reported not only in tongue peripheral nerves,1,8–10 but also in tongue and palatine epithelium.8,10 Based on these findings, it has been hypothesized that sensation is first received by TRPV1 expressed in the epithelium and then transmitted to peripheral nerves. The importance of the epithelium, particularly in the palatal mucosa has recently drawn attention.10,11 TRPV1 epithelial expressions suggest that the epithelium may be a novel sensory detector of temperature and chemicals in the oral environment.10,11 TRPV1-immunoreactive (ir) epithelial cells are more prevalent than TRPV1-ir nerves in the surface layer of the dorsum of the tongue.12 Therefore, it is possible that the sensory and transmission systems exist in the surface of the tongue as well, transmitting the sensations mediated by TRPV1 through the epithelium facing to the oral cavity to the peripheral nervous system. However, the anatomical interaction between the TRPV1-ir epithelium and the peripheral nerves, which receive the sensation from the epithelium, remains unclear. In immunohistochemical studies, one author reported TRPV1-ir images in the tongue epithelium of healthy subjects,1 whereas the other reported that TRPV1 was not expressed in the epithelium of both healthy subjects and burning mouth syndrome patients.8 The patient is characterized by oral burning sensation or oral burning pain.8 Thus, a consensus on the expression of TRPV1 in the epithelium of the human tongue has not been established. It has been reported that administration of capsaicin to the tongue influences expression of calcitonin gene-related peptide (CGRP) in peripheral nerves.13 CGRP-ir nerves are distributed around the epithelium, lingual papilla, and blood vessels of the tongue and the tongue is rich in CGRP-ir nerves.13 Because it induces vasodilatation, it is associated with headaches and the promotion of histamine diffusion, which causes inflammation.14,15 There is a possibility of capsaicin sensitivity in CGRP-ir nerves. Because of the distribution of nerves in the tongue, CGRP-ir nerves may be among the peripheral nerves related to pain reception and sensory information transmission from the TRPV1-ir epithelium. The purpose of this study was to determine the neuroanatomical basis of signal transmission from the epithelial cells to the peripheral nervous system.
Materials and Methods
Animals
A total of 14 adult male rodents were used in this study, 7 Wistar rats and 7 Imprinting Control Region (ICR) mice. Research was performed in accordance with the guidelines of the official Japanese regulations for research on animals, according to institutionally approved protocols (the Nippon Dental University, approval No.08–02). The animals were perfused with 4% paraformaldehyde in 0.1 M phosphate-buffered saline after being deeply anesthetized using Nembutal as previously described.16
Single immunohistochemistry for TRPV1 and CGRP
Tongues were dissected carefully at the posterior portion of the tongue corpus or at the border of the root. Additionally, one rat trigeminal ganglion was removed from the skull and used for antibody adsorption test as described later. The tissues were frozen rapidly by immersioning in cold n-hexane and cut transversely or sagittally at a thickness of 30 µm using a cryostat. The sagittal sections were serially cut and collected. In these sections, one section was stained using hematoxylin and eosin solution for general cytoarchitecture observation (Figure 1). For TRPV1 and CGRP immunohistochemistry, we conducted as previously described procedure.16 In briefly, the sections were processed for single immunoperoxidase staining after blocking by using normal goat serum. These were incubated with primary antibodies for 1–2 days at 4°C or room temperature (rabbit polyclonal anti-TRPV1 antibody: 1:50, PC420 Calbiochem, formerly Ab-1 Oncogene;10 mouse monoclonal anti-TRPV1 antibody: 1:3000, MAB0290 Abnova; and rabbit polyclonal anti-CGRP antibody: 1:1000, T-4032 Peninsula Lab). Finally, sections were incubated with biotinylated anti-rabbit or anti-mouse secondary goat antibodies (1:200, Vector Laboratories) and visualized by using avidin-biotin complex conjugated horseradish peroxidase (ABC Elite kits, Vector Laboratories) with 3, 3′-diaminobenzidine tetrahydrochloride (DAB) containing 0.5% nickel ammonium sulfate (NAS). Routine controls for immunostaining were carried out by omitting the three primary antibodies in the all experiments. For positive control, we confirmed TRPV-1 expression in the rat tongue using a rabbit polyclonal anti-TRPV1 antibody (PC420 Calbiochem, formerly Ab-1 Oncogene10), which is used in a first report of TRPV-1 expression in epithelium.10 The specificity of the rabbit polyclonal anti-TRPV1 antibody and anti-CGRP antibody were described previously in rat tongue and trigeminal ganglion.10,17,18 On the other hands, to assess the specificity of the mouse monoclonal anti-TRPV1 antibody (MAB0290 Abnova), we demonstrated the following adsorption test. After preincubation of the anti-TRPV1 antibody with 10 µg/mL of the TRPV1 carboxy-terminal peptide (P14113 neuromics), immunoreactivity for the TRPV1 was abolished in the rat tongue and trigeminal ganglion as shown in Figure 2.
Figure 1.
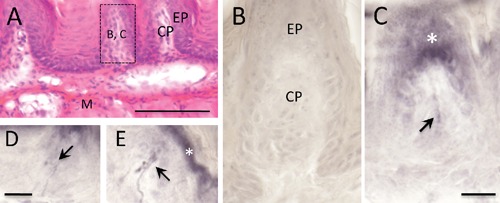
Single immunoperoxidase staining of TRPV1 and a hematoxylin and eosin staining (HE) in the mouse tongue. A) Sagittal section of HE staining in an anterior region of the tongue; a boxed region is approximately corresponding to panels B and C; B) control section by omitting the primary antibody for TRPV1 immunoperoxidase staining; C–E) TRPV1 immunoreactive (ir) nerve fibers are observed (arrows); white asterisks indicate strongly TRPV1-ir epithelium of filiform papilla. CP, connective tissue; EP, epithelium; M, muscle layer. Scale bars: A) 100 µm; B–E) 20 µm.
Figure 2.
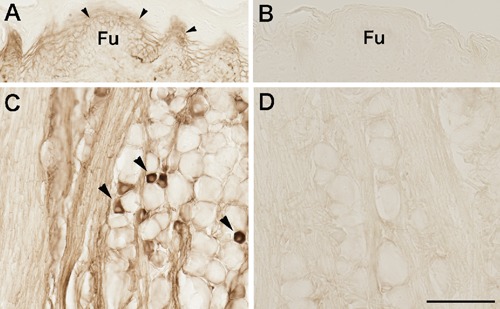
TRPV1 immunoreactive (ir) lingual epithelia (arrowheads in panel A) and trigeminal ganglion neurons (arrowheads in panel C). After the adsorption of primary TRPV1 antibody, no TRPV1-ir epithelia and neurons were detected (B and D). Fu, fungiform papilla. Scale bars: 100 µm.
Double immunohistochemistry for TRPV1 and CGRP
TRPV1 and CGRP double immunoperoxidase staining was performed as previously described.19 After CGRP single immunoperoxidase staining was performed using the rabbit polyclonal antibody as described above, the stained sections were incubated again in mouse monoclonal anti-TRPV1 antibody for 1–2 days at 4°C or room temperature. The sections were incubated with biotinylated anti-mouse secondary goat antibody and visualized by DAB without NAS. For TRPV1 and CGRP double immunofluorescence staining, sections were incubated in a mixture of two primary antibodies, mouse monoclonal anti-TRPV1 antibody (1:3000) and rabbit polyclonal anti-CGRP antibody (1:1000). The sections were then incubated in a mixture of two secondary antibodies, Alexa Fluor 488-conjugated anti-mouse and Alexa Fluor 594-conjugated anti-rabbit polyclonal goat antibodies (1:200, Molecular Probes, Life Technologies Japan Ltd., Tokyo, Japan).
Images manipulation and mapping preparation
Photographs were taken as digital images by a digital camera equipped with a microscope (FSX-100, Olympus Corp., Tokyo, Japan). Images were merged and/or adjusted for brightness to match the real image using standard image software (Adobe Photoshop CS4, Adobe). Using the TRPV1 and CGRP double immunoperoxidase staining sections, we mapped anteroposterior distribution of the TRPV1-ir epithelium and CGRP-ir nerve terminals using a light microscope with a camera lucida drawing apparatus (BX51, Olympus).
Results
TRPV1 distributions
We observed TRPV1 localization in tongue epithelial cells and nerve terminals (Figure 1: mouse case). TRPV1-ir terminals were clearly observed in connective tissue beneath the papilla epithelium (Figure 1, D–E). The papilla epithelium was also strongly positive for TRPV1 (Figure 1: mouse case; Figures 3, 4 and 5: rat case). TRPV1-ir terminals were sparsely distributed in the connective tissue in contrast to the strongly TRPV1-ir epithelium. Portions of the strongly TRPV1-ir epithelium were observed on top of the papilla (Figure 1C, Figures 3, 4 and 5). These observations were corresponding to a previous report describing TRPV1 expression in the rat tongue epithelium and nerve fibers.10 This confirmed that two anti-TRPV1 antibodies used in this study recognized appropriate TRPV1 distributions in the rodent tongue.
Figure 3.
Double immunoperoxidase staining of TRPV1 and CGRP in the rat tongue. A) Clear TRPV1 immunoreactive (ir) epithelium (arrows) are observed in apex region (left side of the panel A) of the tongue; note pale immunoreactivity of the TRPV1 in posterior region (right side of the panel A), double arrows indicate an approximate intersectional region; the double arrows correspond to double arrows in the Figure 4 A. B) Higher magnification of apex region; arrows showing TRPV1-ir epithelium; arrowheads showing CGRP-ir terminals, small black dot like structure, around TRPV1-ir epithelium of the fungiform papilla (see also Figures 4 and 5); C) higher magnification of posterior region; weakly TRPV1-ir is observed (arrows); D) control section by omitting the primary antibodies. Fi, filiform papilla; Fu, fungiform papilla. Scale bars: A) 100 µm; B–D) 50 µm.
Figure 4.
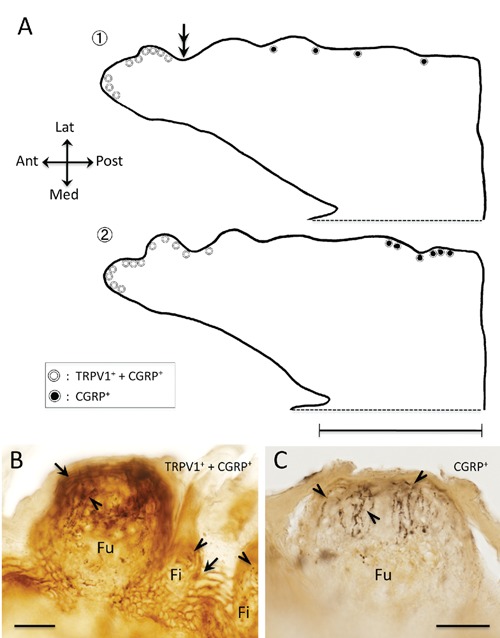
Distributions of TRPV1 immunoreactive (ir) and CGRP-ir in the fungiform papilla in the rat tongue. A) Line drawings of sagittal section of the tongue; the two line drawings are reconstructed from total 71 sections, the section no. 1 consists 35 sections and no. 2 consists 36 sections respectively, and depicted approximately 2 mm extent from midline to lateral; white and black circles indicating the fungiform papilla distribution on the tongue; white circles, strongly TRPV1-ir epithelium (arrow in panel B) and CGRP-ir terminals (arrowheads in panel B) distributing around the epithelium; black circles, CGRP-ir terminals (arrowheads in panel C) distributing around the epithelium, which is not TRPV1-ir epithelium (C); note concentration of white circles in apex region; double arrows indicate an approximate posterior border of the concentration and correspond to double arrows indicating in the Figure 3A; B,C) photographs of the two cases of fungiform papilla, B corresponding to the white circle and C corresponding to the black circle respectively. Ant, anterior; Fi, filiform papilla; Fu, fungiform papilla; Lat, lateral; Med, medial; Post, posterior. Scale bars: A) 5 mm; B,C) 30 µm.
Figure 5.
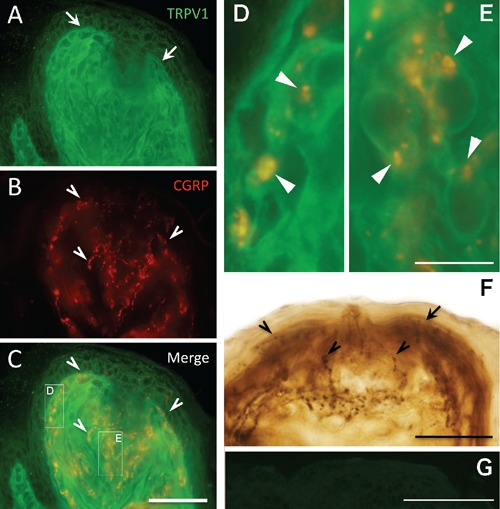
Double immunostaining of TRPV1 and CGRP in the fungiform papilla of rat tongue. A–C) Double immunofluorescence staining of TRPV1 (A) and CGRP (B) in the fungiform papilla; C) merged image of panels A and B; arrows showing TRPV1 immunoreactive (ir) epithelium in green; arrowheads showing CGRP-ir terminals in red; boxed regions (D and E) correspond to panels D and E. D, E) Higher magnification of boxed regions in panel C; note CGRP-ir terminals (arrowheads) appositions around the TRPV1-ir epithelium; F) double immunoperoxidase staining of TRPV1 and CGRP in apical of the fungiform papilla; note CGRP-ir terminals (arrowheads) distribute around strongly TRPV1-ir epithelium (arrows) in apical region of the papilla; G) control section by omitting the primary antibody for TRPV1 immunofluorescence staining. Scale bars: AC) 50 µm; D, E) 10 µm; F) 30 µm; G) 100 µm.
CGRP-ir nerve terminals in tongue papilla
CGRP-ir fibers were found in deep connective tissue areas that run into connective tissue beneath the papilla epithelium (Figure 6 A: rat case). The fiber terminated in the epithelium of filiform and fungiform papillae (Figure 6 B, C). Some CGRP-ir terminals were found densely localized at the apical edge of the papilla epithelium (Figure 6 B, C). In addition to the epithelial distributions, CGRP-ir fibers were observed around blood vessels (Figure 6 D, E) and in muscle fibers (data not shown).
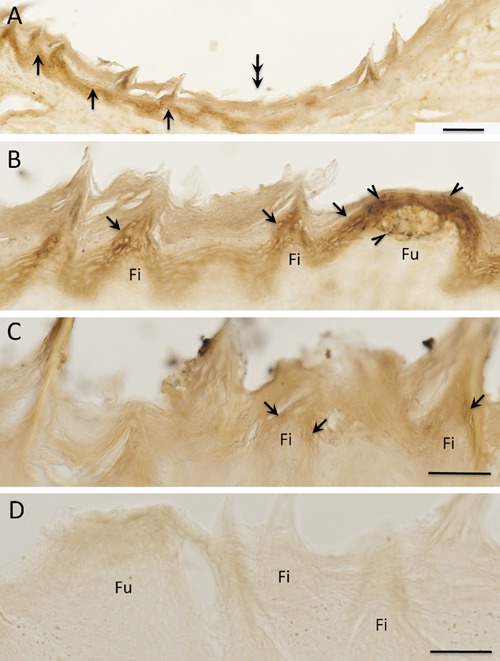
Figure 6.
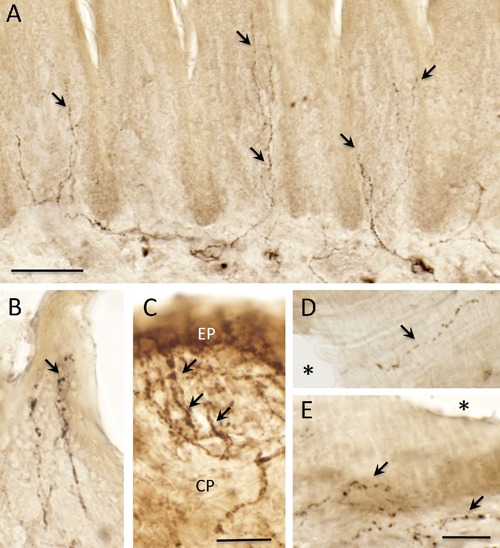
Single immunostaining of CGRP in the rat tongue. A–C) CGRP immunoreactive (ir) nerve fibers and terminals (arrows) in tongue papilla; filiform papilla (A and B); fungiform papillae (C); note dense CGRP-ir terminals in the fungiform papillae; D, E) CGRP-ir terminals are seen around blood vessel walls (arrows); asterisks indicate lumen of the blood vessel. CP, connective tissue; EP, epithelium. Scale bars: A) 50 µm; B–E) 25 µm.
Topographical organization of TRPV1 and CGRP in fungiform papilla
Upon double immunostaining, TRPV1 and CGRP displayed a notable anteroposterior distribution in the TRPV1-ir epithelium. Strong TRPV1 immunoreactivity was observed in the epithelium at the apex of the tongue in the sagittal sections (Figure 3). In the apex region, dense CGRP-ir terminals were co-localized with the TRPV1-ir epithelium, particularly in the fungiform papilla (Figures 4 and 5). We observed no clear co-localization of TRPV1 and CGRP in fungiform papilla in the posterior area of the tongue (Figure 4). Co-localization was confirmed by double immunofluorescence staining (Figure 5). Fluorescence images revealed apposition of CGRP-ir terminals to TRPV1-ir epithelial cells in the fungiform papilla in the apex region (Figure 5 A–E).
Discussion
We found strong expression of TRPV1 in the epithelial cells, particularly at the apex of the tongue. While TRPV1-ir nerve terminals were sparsely observed in comparison to previous studies.9,10 The sparse distributions of TRPV1-ir nerve terminals might be caused by fixation using 4% paraformaldehyde. There is a report that TRPV1 immunoreactivity is affected by formaldehyde concentration in the rat dorsal root ganglia.20 They resulted that decreasing formaldehyde concentration and increasing picric acid concentration elevated TRPV1 immunoreactivity. According to the report, the percentage of dorsal root ganglia neurons labeling for TRPV1 were 15% and 77% using 4% and 0.25% formaldehyde respectively.
A previous study claimed that TRPV1 epithelial expression was a novel observation and conspicuous when compared to TRPV1 expression in nerves.10 However, our findings of the restricted anterior distribution of TRPV1-ir epithelial cells in the rodent tongue have not been previously reported. In spite of fixative condition, which might not be optimized for detection TRPV1 in nerve, we could observe the clear restricted labeling of TRPV1-ir epithelial cells. This expression of the TRPV1 in apex of tongue implies consistency with the functional role of discriminating between prey and hazardous molecules by perception of heat and acid. In other words, the TRPV1-ir epithelium could serve as a barrier of entry to the oral cavity. As a point of comparison, the TRPV1 distribution pattern observed in this study is similar to a human case in which TRPV1 expression was determined by tongue biopsy. One author observed TRPV1 epithelial expression in the apex region of normal human tissue.1 In addition, the apex region of tongue is known to be one of the most common regions of burning pain in patients with burning mouth syndrome (BMS).1 BMS is characterized by oral burning sensation or oral burning pain in the absence of any organic abnormal findings in the oral mucous membrane. The pain appears in the tongue, the lips, and the hard palate and sometimes in the buccal mucosa and the pharynx.21,22 It is often diagnosed as glossodynia in Japan because the pain appears particularly in the tongue. Currently, glossodynia is included within the BMS diagnosis rubric. Hormonal changes associated with menopause, psychological factors, sensory disorders, and neuropathic mechanisms have been considered as causal factors of BMS; however, etiology and effective therapeutic strategies have remained elusive.22,23 Expression of TRPV1 may be an important factor in pain reception in the tongue of a patient with BMS.8 Some patients with BMS complain of pain during eating. In these patients, irritants such as chili peppers and hot food increase the pain. Use of surface anesthetics or Rikko-san, an oriental herbal medicine with local anesthetic activity, often relieves pain in the patients.24 TRPV1 in the tongue is considered to be associated with the BMS pain. Collectively, these reports indicate that the anterior distribution of TRPV1 may be functionally significant for perception of burning pain in normal conditions and/or BMS. To clarify the hypothesized functional organization of the tongue epithelium in humans, further investigation of TRPV1 expression in the whole tongue with anteroposterior direction mapping (as demonstrated in this study) is required.
CGRP-ir nerve terminals were observed widely in the tongue, including the epithelium and muscle, and around blood vessels. In papil- la epithelium, rich CGRP-ir terminals were observed in the fungiform papilla. Apposition of CGRP-ir terminals in the epithelium to the TRPV1-ir apical epithelium of the apex region were also evident. This suggests a possibility that TRPV1-ir epithelium receives ligands and information flow may be conducted to adjacent CGRP-ir terminals. Concerning potential epithelial cell and nerve interaction, the skin epithelium cell membrane makes contact, membrane-membrane apposition, with nerve fiber membranes to allow interaction between the epithelium and nerves.23,25 In TRPV2, another member of the TRPV subfamily that is activated by high temperature (>52°C) and is expressed in gingival epithelium cells, the epithelial cell membrane apposition with nerves was observed by transmission electron microscopy in rats.26 Previous anatomical researches have demonstrated tissue architecture for oral epithelium interaction with nerves. Recently, fungiform papilla epithelium has been shown to expresses vesicular glutamate transporter 1 in rats.27 This strongly suggests that fungiform papilla epithelium releases glutamate. In turn, nerve terminals around fungiform papilla epithelium express one of the glutamate receptors, GluR2/3.28 Therefore, tongue papilla epithelium can use glutamate as a transmitter and conduct signals to nerve terminals at least in the rat fungiform papilla. The two reports support the possibility that apposition of TRPV1-ir epithelium and CGRP-ir nerves in this study may allow functional interaction in physiological conditions.
Combining our findings of the possible interaction between the epithelium and nerves with aging and hormonal status may be key to understanding BMS. Endocrine status at menopause has been considered as a potential indicator of BMS etiology.22,29 In female rats (10–12 months of age), CGRP expression in the dorsal root ganglion was influenced by estrogen and progesterone.30 In human skin epithelium, TRPV1 expression also increased with aging.7 Thus, hormonal status affected CGRP expression in nerves and aging affected TRPV1 expression in the epithelium.
To clarify the hypothesized interaction between the TRPV1-ir epithelium and CGRP-ir nerves, further investigations are required, particularly those that utilize a polysynthetic approach.
Acknowledgments:
the authors would like to thank the members of the Department of Anatomy at NDU for their general cooperation.
References
- 1.Tachibana T, Chiba H. Relationship between glossodynia and capsaicin sensitive sensory neurons. [Article in Japanese] J Jpn Stomatol Soc. 2006;55:167–78. [Google Scholar]
- 2.Tominaga M, Tominaga T. Structure and function of TRPV. Pflugers Arch-Eur J Physiol. 2005;451:143–50. doi: 10.1007/s00424-005-1457-8. [DOI] [PubMed] [Google Scholar]
- 3.William D, Willis JR. The role of TRPV1 receptors in pain evoked by noxious thermal and chemical stimuli. Exp Brain Res. 2009;196:5–11. doi: 10.1007/s00221-009-1760-2. [DOI] [PubMed] [Google Scholar]
- 4.Denda M, Fuziwara S, Inoue K, Denda S, Akamatsu H, Tomitaka A, et al. Immunoreactivity of VR1 on epidermal keratinocyte. Biochem Biophys Res Commun. 2001;285:1250–2. doi: 10.1006/bbrc.2001.5299. [DOI] [PubMed] [Google Scholar]
- 5.Funakoshi K, Nakano M, Atobe Y, Goris RC, Kadota T, Yazama F. Differential development of TRPV1-expressing sensory nerves in peripheral organs. Cell Tissue Res. 2006;323:27–41. doi: 10.1007/s00441-005-0013-3. [DOI] [PubMed] [Google Scholar]
- 6.Lumpkin EA, Caterina MJ. Mechanisms of sensory transduction in the skin. Nature. 2007;445:856–65. doi: 10.1038/nature05662. [DOI] [PubMed] [Google Scholar]
- 7.Lee YM, Kim YK, Chung JH. Increased expression of TRPV1 channel in intrinsically aged and photoaged human skin in vivo. Exp Dermatol. 2009;18:431–6. doi: 10.1111/j.1600-0625.2008.00806.x. [DOI] [PubMed] [Google Scholar]
- 8.Yilmaz Z, Renton T, Yiangou Y, Zakrzewska J, Chessell IP, Bountra C, et al. Burning mouth syndrome as a trigeminal small fiber neuropathy: Increased heat and capsaicin receptor TRPV1 in nerve fibers correlates with pain score. J Clin Neurosci. 2007;14:864–71. doi: 10.1016/j.jocn.2006.09.002. [DOI] [PubMed] [Google Scholar]
- 9.Ishida Y, Ugawa S, Ueda T, Murakami S, Shimada S. Vanilloid receptor subtype-1 (VR1) is specifically localized to taste papillae. Brain Res Mol Brain Res. 2002;107:17–22. doi: 10.1016/s0169-328x(02)00441-2. [DOI] [PubMed] [Google Scholar]
- 10.Kido MA, Muroya H, Yamaza T, Terada Y, Tanaka T. Vanilloid receptor expression in the rat tongue and palatine. J Dent Res. 2002;82:393–7. doi: 10.1177/154405910308200513. [DOI] [PubMed] [Google Scholar]
- 11.Wang B, Danjo A, Kajiya H, Okabe K, Kido MA. Oral epithelial cells are activated via TRP channels. J Dent Res. 2011;90:163–7. doi: 10.1177/0022034510385459. [DOI] [PubMed] [Google Scholar]
- 12.Kawashima M, Imura K, Miwa Y, Sato I. Distribution patterns of the TRPV1 in the mouse tongue. [Article in Japanese] Acta Anatomica Nipponica. 2010;85:147–147. [Google Scholar]
- 13.Terenghi G, Polak JM, Rodrigo J, Mulderry PK, Bloom SR. Calcitonin gene-related peptide immunoreactive nerves in the tongue, epiglottis and pharynx of the rat: occurrence, distribution and origin. Brain Res. 1986;365:1–14. doi: 10.1016/0006-8993(86)90716-x. [DOI] [PubMed] [Google Scholar]
- 14.Benemei S, Nicoletti P, Capone JA, Geppetti J. Pain pharmacology in migraine: focus on CGRP and receptors. Neurol Sci. 2007;28:89–93. doi: 10.1007/s10072-007-0757-5. [DOI] [PubMed] [Google Scholar]
- 15.Shimizu T, Suzuki N. Neurotransmitter related to migraine headache. Clinic All-Round. 2007;56:983–7. [Google Scholar]
- 16.Imura K, Sato I. Novel localization of tenascin-X in adult mouse leptomeninges and choroid plexus. Ann Anat. 2008;190:324–8. doi: 10.1016/j.aanat.2008.04.003. [DOI] [PubMed] [Google Scholar]
- 17.Yamamoto Y, Hatakeyama T, Taniguchi K. Immunohisto chemical colocalization of TREK-1, TREK-2 and TRAAK with TRP channels in the trigeminal ganglion cells. Neurosci Lett. 2009;454:129–33. doi: 10.1016/j.neulet.2009.02.069. [DOI] [PubMed] [Google Scholar]
- 18.Kosaras B, Jakubowski M, Kainz V, Burstein R. Sensory innervation of the calvarial bones of the mouse. J Comp Neurol. 2009;515:331–48. doi: 10.1002/cne.22049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Imura K, Rockland KS. Long-range interneurons within the medial pulvinar nucleus of macaque monkeys. J Comp Neurol. 2006;498:649–66. doi: 10.1002/cne.21085. [DOI] [PubMed] [Google Scholar]
- 20.Hoffman EM, Schechter R, Miller KE. Fixative composition alters distributions of immunoreactivity for glutaminase and two markers of nociceptive neurons, Nav1.8 and TRPV1, in the rat dorsal root ganglion. J Histochem Cytochem. 2010;58:329–44. doi: 10.1369/jhc.2009.954008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Silverman IS. The burning mouth syndrome. J Dent Ass Afr. 1975;130:163–6. [PubMed] [Google Scholar]
- 22.Lauria G, Majorana A, Borgna M, Lombardi R, Penza P, Padovani A, et al. Trigeminal small-fiber sensory neuropathy causes burning mouth syndrome. Pain. 2005;115:332–7. doi: 10.1016/j.pain.2005.03.028. [DOI] [PubMed] [Google Scholar]
- 23.Suzuki T. Recent progress in sensory mechanism. Bull Tokyo Dent Coll. 2007;48:1–7. doi: 10.2209/tdcpublication.48.1. [DOI] [PubMed] [Google Scholar]
- 24.Takahashi T, Kawashima M, Shibaji T, Suzuki N, Umino M, Shimada M. A case of burning mouth syndrome effectively treated with Rikkosan. J Jpn Dent Soc Anesthesiol. 2007;35:392–3. [Google Scholar]
- 25.Hilliges M, Wang L, Johansson O. Ultrastructural evidence for nerve fibers within all vital layers of the human epidermis. J Invest Dermatol. 1995;104:134–7. doi: 10.1111/1523-1747.ep12613631. [DOI] [PubMed] [Google Scholar]
- 26.Shimohira D, Kido MA, Danjo A, Takao T, Wang B, Zhang JQ, et al. TRPV2 expression in rat oral mucosa. Histochem Cell Biol. 2009;132:423–33. doi: 10.1007/s00418-009-0616-y. [DOI] [PubMed] [Google Scholar]
- 27.Braud A, Boucher Y, Zerari-Mailly F. Vesicular glutamate transporters localization in the rat lingual papillae. Neuroreport. 2010;21:64–67. doi: 10.1097/WNR.0b013e3283341bd0. [DOI] [PubMed] [Google Scholar]
- 28.Caicedo A, Zucchi B, Pereira E, Roper SD. Rat gustatory neurons on the geniculate ganglion express glutamate receptor subunits. Chem Senses. 2004;29:463–71. doi: 10.1093/chemse/bjh048. [DOI] [PubMed] [Google Scholar]
- 29.Suzuki N. Clinical experiences of intractable pain in oral and maxillofacial regions. J Jpn Dent Soc Anesthesiol. 2007;35:323–9. [Google Scholar]
- 30.Gangula PR, Chauhan M, Reed L, Yallampalli C. Age-related changes in dorsal root ganglia, circulating and vascular calcitonin gene-related peptide (CGRP) concentrations in female rats: effect of female sex steroid hormones. Neurosci Lett. 2009;454:118–23. doi: 10.1016/j.neulet.2009.02.068. [DOI] [PMC free article] [PubMed] [Google Scholar]