

Abstract
Background
The Yellow Cat Member of the Cedar Mountain Formation (Early Cretaceous, Barremian?) of Utah has yielded a rich dinosaur fauna, including the basal therizinosauroid theropod Falcarius utahensis at its base. Recent excavation uncovered a new possible therizinosauroid taxon from a higher stratigraphic level in the Cedar Mountain Formation than F. utahensis.
Methodology/Principal Findings
Here we describe a fragmentary skeleton of the new theropod and perform a phylogenetic analysis to determine its phylogenetic position. The skeleton includes fragments of vertebrae, a scapula, forelimb and hindlimb bones, and an ischium. It also includes several well-preserved manual unguals. Manual and pedal morphology show that the specimen is distinct from other theropods from the Cedar Mountain Formation and from previously described therizinosauroids. It is here named as the holotype of a new genus and species, Martharaptor greenriverensis. Phylogenetic analysis places M. greenriverensis within Therizinosauroidea as the sister taxon to Alxasaurus + Therizinosauridae, although support for this placement is weak.
Conclusions/Significance
The new specimen adds to the known dinosaurian fauna of the Yellow Cat Member of the Cedar Mountain Formation. If the phylogenetic placement is correct, it also adds to the known diversity of Therizinosauroidea.
Introduction
The Cedar Mountain Formation (Lower Cretaceous) of Utah, USA, preserves a rich theropod paleofauna. The paleofauna of the Ruby Ranch Member includes a large carnosaur similar to Acrocanthosaurus [1]. The paleofauna of the lower sequence of the Yellow Cat Member includes the basal therizinosauroid Falcarius utahensis [2]–[4], the troodontid Geminiraptor suarezarum [5], an unnamed velociraptorine dromaeosaurid [6], and the dromaeosaurine dromaeosaurid Yurgovuchia doellingi [6]. The paleofauna of the upper sequence of the Yellow Cat Member includes the coelurosaur Nedcolbertia justinhofmanni [7], the dromaeosaurine dromaeosaurid Utahraptor ostrommaysorum [8], and an unnamed eudromaeosaur [6].
Here we describe a new theropod specimen from the upper Yellow Cat Member.
The specimen is from the Hayden-Corbett Quarry, Utah state Loc. # Gr.287v, approximately eight miles southeast of the city of Green River, Utah. The specimen is a possible therizinosauroid.
The clade Therizinosauroidea is part of the clade Coelurosauria within the dinosaurian clade Theropoda. Therizinosauroids are known from the Lower and Upper Cretaceous of Asia and North America [9]–[23]. Therizinosauroids range in size from the approximately collie-sized Beipiaosaurus inexpectus [9] to the gigantic Therizinosaurus cheloniformis, which towered over contemporaneous tyrannosaurids [24]. They exhibit dental specializations for herbivory [2], [9], as do several other coelurosaurian groups [23], [25].
One possible therizinosauroid, Eshanosaurus deguchiianus, has been reported from the Lower Jurassic of China [26]. However, the specimen consists only of a single dentary bone, and of its 11 putatively therizinosauroid character states that were reexamined in a recent study [27], all but three (high tooth count, tooth roots wider than crowns, morphology of lateral dentary ridge) are also known in Sauropodomorpha [27]. If the specimen is therizinosauroid, it establishes a ghost lineage ∼65 million years long for Therizinosauroidea [27] and ∼35 million years long for Coelurosauria. We therefore urge caution in the interpretation of E. deguchiianus as a therizinosauroid until more of its anatomy is discovered.
Methods
Phylogenetic Analysis
We entered data from the new specimen into an updated version (Appendix S1, S2) of a phylogenetic data matrix of Coelurosauria from a recent study [6]. Updates consist of corrections to character 212 (wide distal expansion of scapula) for several therizinosauroid OTUs. Phylogenetic analysis was performed with PAUP 4.0 for Windows [28]. A heuristic search with 1000 random addition-sequence replicates was performed, with no limit to “maxtrees.”
Nomenclatural Acts
The electronic version of this document does not represent a published work according to the International Code of Zoological Nomenclature (ICZN), and hence the nomenclatural acts contained in the electronic version are not available under that Code from the electronic edition. Therefore, a separate edition of this document was produced by a method that assures numerous identical and durable copies, and those copies were simultaneously obtainable (from the publication date noted on the first page of this article) for the purpose of providing a public and permanent scientific record, in accordance with Article 8.1 of the Code. The separate print-only edition is available on request from PLoS by sending a request to PLoS ONE, 1160 Battery Street, Suite 100, San Francisco, CA 94111, USA along with a check for $10 (to cover printing and postage) payable to “Public Library of Science”.
In addition, this published work and the nomenclatural acts it contains have been registered in ZooBank, the proposed online registration system for the ICZN. The ZooBank LSIDs (Life Science Identifiers) can be resolved and the associated information viewed through any standard web browser by appending the LSID to the prefix “http://zoobank.org/”. The LSID for this publication is: urn:lsid:zoobank.org:pub:12E26311-C182-40D0-8F6F-7FE4B343B261.
Results
Systematic Paleontology
Systematic hierarchy.
Dinosauria Owen, 1841 [29]
Saurischia Seeley, 1887 [30]
Theropoda Marsh, 1881 [31]
Coelurosauria von Huene, 1914 [32]
Therizinosauroidea Maleev, 1954 [33]
Martharaptor gen nov.
urn:lsid:zoobank.org:act:9F589210-60D8-4971-95A5-6ACE4D0BA361
Martharaptor greenriverensis sp. nov.
urn:lsid:zoobank.org:act:5065191E-53CD-4B48-8C95-8DC7E405F9CB
Holotype
The holotype specimen is UMNH VP 21400 (Natural History Museum of Utah, Salt Lake City, Utah).
Etymology
The species name refers to the city of Green River in Emery County, Utah. The genus name honors Martha Hayden, who co-discovered the site and has served as the assistant to three successive state paleontologists of Utah over a period of about 25 years.
Geological setting
The Hayden-Corbett Site (Gr287v) is at the top of an approximately 20 cm thick sandstone, interpreted to represent a crevasse splay deposit, in the Yellow Cat Member of the Cedar Mountain Formation approximately 8 miles southeast of Green River, Utah in the immediate vicinity of the Suarez and Crystal Geyser sites, both of which are bone beds yielding abundant specimens of the basal therizinosauroid Falcarius. A gravelly, calcareous sandstone often hosting a stromatolitic layer at its top that divides the Yellow Cat Member into an upper and lower sequence [34], [35] has been informally designated the caprock in this area. The Hayden-Corbett site is above this regional marker bed, whereas both the Suarez and Crystal Geyser Quarries are below the caprock (Fig. 1). The upper Yellow Cat Member has been dated at about 124 Ma (early Aptian) based on detrital zircons [36].
Figure 1. Stratigraphy of the Hayden-Corbett Site (Gr287v).

Stratigraphic section of the basal Cedar Mountain Formation of eastern Utah, showing that Martharaptor is from a bed stratigraphically higher than those yielding Falcarius.
The bones of the holotype of M. greenriverensis were found disarticulated. However, there is no indication that more than one individual is present in the sample. All the skeletal material, including the surface “float” that drew attention to the skeleton, was found in an area of less than two square meters, and most of it is from an excavated mudstone block that is less than one square meter in lateral area and less than 0.2 m deep. There is no duplication of elements, the sizes of the bones are consistent with their having come from a single individual, and there are no morphological indicators that multiple taxa are present. We are therefore confident that the material can all be assigned to a single individual.
Diagnosis
Theropod dinosaur with the following combination of character states: cervical prezygapophyses not flexed; cranial dorsal vertebrae with hypapophyses and a single pair of pneumatopores; manual unguals without proximodorsal lips and with prominent flexor tubercles and strong curvature; manual unguals in which total length perpendicular to the articular facet is subequal to total height parallel to the articular facet; ungual of manual digit III nearly as large as that of digit II; distal end of scapula expanded; proximal end of ischium laterally compressed; metatarsal I proximally attenuated and distally reduced in transverse width relative to the other metatarsals; all metatarsals distally non-ginglymoid; fourth metatarsal distally attenuated immediately proximal to condyles; pedal unguals laterally compressed and strongly curved; first pedal ungual smaller than the others.
No other theropod dinosaur exhibits this combination of character states. However, because of the fragmentary nature of the specimen, it is important to be specific about how this combination of character states distinguishes the specimen as a new taxon. Therefore, below we show how these character states distinguish the new specimen from other theropods of the Cedar Mountain Formation and from previously described therizinosauroids.
Martharaptor greenriverensis can be distinguished from the Cedar Mountain Formation coelurosaur Nedcolbertia justinhofmanni by differences in manual and pedal morphology. In N. justinhofmanni the manual unguals are nearly straight, and the flexor tubercle of the first manual ungual is strongly pendant and approximately half the height of the articular facet. The proximal surface of metatarsal II is subtriangular, and that of metatarsal III is a craniocaudally elongate rectangle [7]. In contrast, the manual unguals of M. greenriverensis are strongly curved, the proximal surface of metatarsal II is nearly square, and the proximal surface of metatarsal III is approximately as wide transversely at it is long craniocaudally.
In the known material of M. greenriverensis there are no preserved bones in common with the troodontid Geminiraptor suarezarum [5], the dromaeosaurids Utahraptor ostrommaysorum [8] and Yurgovuchia doellingi [6], or the unnamed dromaeosaurids from the Cedar Mountain Formation [6]. However, M. greenriverensis lacks character states that are present in troodontids and dromaeosaurids, such as flexed cervical prezygapophyses, a distally unexpanded scapula, and strongly pendant flexor tubercles on the manual unguals.
M. greenriverensis can be distinguished from Falcarius utahensis, a therizinosauroid from the Cedar Mountain Formation, by the morphology of its manual unguals and fourth metatarsal. In F. utahensis the shape of the first manual ungual is markedly different from the others [2], whereas in M. greenriverensis all manual unguals resemble each other in shape (Fig. 2). The shaft of the third manual ungual is more gracile in F. utahensis [2] and is taller (deeper in the dorso-palmar dimension) in M. greenriverensis (Fig. 2). In M. greenriverensis metatarsal IV is more strongly beveled distally than it is in Falcarius [3].
Figure 2. Outlines of manual unguals I (left), II (middle), and III (right) of Martharaptor greenriverensis and two other basal therizinosauroids.
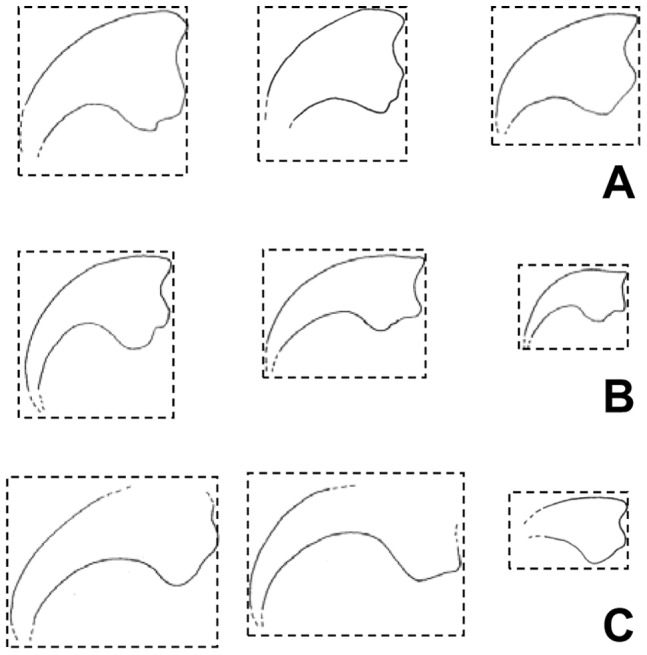
(A)–Martharaptor greenriverensis. (B)–Falcarius utahensis, after Zanno [3] (C)–Beipiaosaurus inexpectus, drawn from photos by the senior author from a cast of IVPP (Institute of Vertebrate Paleontology and Paleoanthropology, Beijing, China) V 11559. Outlines are to scale within sub-figure A and within sub-figure C but not within sub-figure B or between sub-figures. Note that M. greenriverensis differs from F. utahensis and B. inexpectus in overall ungual shape and flexor tubercle morphology, and differs from B. inexpectus in the relative size of the ungual of digit III. The size of the ungual of digit III relative to the others cannot be determined for F. utahensis, because no known specimen preserves all three unguals.
Other therizinosauroids differ from M. greenriverensis in the following ways. In Beipiaosaurus inexpectus the manual unguals are more gracile and the third manual is much smaller than the others [9], whereas in M. greenriverensis the manual unguals are deep and the third ungual is only slightly smaller than the others (Fig. 2). In Alxasaurus elesitaiensis and members of Therizinosauridae other than Nothronychus the dorsal surface of each manual ungual arches above the level of the proximal articular facet [12], [19], [21], whereas such is not the case in M. greenriverensis. In members of Therizinosauridae the proximal surface of metatarsal I is robust and articulates with the tarsus [16], [20], [23], whereas that of M. greenriverensis is attenuated and exhibits a lateral facet for articulation with the medial side of metatarsal II. In members of Therizinosauridae the distal surface of metatarsal IV is perpendicular to the shaft [16], [20], [23], whereas it is beveled in M. greenriverensis.
Specimen Description
Preserved bones of M. greenriverensis are described below. Table 1 lists the measurements of these bones.
Table 1. Measurements of bones of the holotype of Martharaptor greenriverensis.
| Bone | Figure | Dimension | Measurement |
| Cervical neural arch | 3A | Total length | 72.70 |
| Prezygapophysis: transverse width | 13.76 | ||
| Cranial dorsal vertebra | 3B–E | Centrum: length | 62.07 |
| Centrum: transverse width | 20.5 | ||
| Centrum: height of cranial face | 41.47* | ||
| Centrum: height of caudal face | 45.77 | ||
| Distal caudal centrum | 3F, G | Length | 32.41* |
| Height | 17.6* | ||
| Presumed ulna | 3H | Length | 159* |
| Diameter | 40.36 | ||
| Radius | 3I | Length | 118.11* |
| Shaft: diameter at proximalend, in plane of flattened distaltip | 22.39 | ||
| Shaft: diameter at proximalend, perpendicular to plane offlattened distal tip | 22.98 | ||
| Distal tip: greatest breadth | 56.88 | ||
| Distal tip: width perpendicular to greatest breadth | 10.74 | ||
| Scapula | 3J, K | Greatest length of proximal fragment | 150.03* |
| Shaft: width | 32.01 | ||
| Length of distal fragment | 186.3* | ||
| Ischium | 3L | Length | 98.65* |
| Possible distal pubis | 3M | Greatest length | 104.18* |
| Width perpendicular to greatest length | 5019* | ||
| Presumed metacarpal I | 4A | Length | 19.15* |
| Distal transverse width | 23.3 | ||
| Presumed manual phalanx I-1 | 4B | Length | 22.40* |
| Proximal height | 18.95 | ||
| Proximal transverse width | 19.12 | ||
| Penultimate manual phalanx | 4C | Length | 25.4* |
| Distal height | 20.1 | ||
| Distal transverse width | 18.23 | ||
| Penultimate manual phalanx | 4D | Length | 25.48* |
| Distal height | 16.23* | ||
| Distal transverse width | 15.95 | ||
| Manual phalanx | 4E | Length | 18.77* |
| Distal height | 18.22 | ||
| Distal transverse width | 19.06 | ||
| Manual phalanx | 4F | Length | 22.09* |
| Distal height | 14.38* | ||
| Distal transverse width | 14.96* | ||
| Manual ungual (digit I) | 4G | Height of articular facet | 43.07 |
| Height of proximal end including flexor tubercle | 43.07 | ||
| Length perpendicular to articular facet | 55.09* | ||
| Greatest transverse width | 15.93 | ||
| Manual ungual (digit I) | 4H | Length approximately perpendicular to articular facet | 33.25* |
| Height approximately parallel to articular facet | 28.58* | ||
| Manual ungual (digit II) | 4I | Height of articular facet | 28.76 |
| Height of proximal end including flexor tubercle | 37.93 | ||
| Length perpendicular to articular facet | 44.44* | ||
| Greatest transverse width | 16.18 | ||
| Manual ungual (digit II) | 4J | Height of articular facet | 29.09 |
| Height of proximal end including flexor tubercle | 36.53 | ||
| Length of proximal fragment perpendicular to articular facet | 40.04* | ||
| Greatest transverse width | 15.90 | ||
| Greatest length of distal fragment | 18.25* | ||
| Manual ungual (digit III) | 4K | Height of articular facet | 24.55 |
| Height of proximal end including flexor tubercle | 36.21 | ||
| Length perpendicular to articular facet | 49.95* | ||
| Greatest transverse width | 14.03 | ||
| Manual ungual (digit III) | 4L | Height of articular facet | 24.92 |
| Height of proximal end including flexor tubercle | 35.65 | ||
| Length perpendicular to articular facet | 51.05* | ||
| Greatest transverse width | 9.56 | ||
| Metatarsal I | 5A | Greatest length | 46.41* |
| Distal width | 19.09 | ||
| Distal depth | 15.03 | ||
| Metatarsal II | 5B | Length | 55.42* |
| Proximal transverse width | 31.22 | ||
| Proximal depth | 30.89 | ||
| Metatarsal II | 5D | Length | 37.76* |
| Distal transverse width | 30.72 | ||
| Distal depth | 38.73 | ||
| Presumed metatarsal III | 5C | Length | 23.22* |
| Proximal transverse width | 25.22* | ||
| Proximal depth | 29.98 | ||
| Metatarsal IV | 5E | Length | 50.48* |
| Distal transverse width | 29.24* | ||
| Distal depth | 28.07* | ||
| Pedal phalanx | 6A | Length | 20.62* |
| Proximal transverse width | 27.84 | ||
| Proximal depth | 24.52 | ||
| Pedal phalanx | 6B | Length | 31.14* |
| Proximal transverse width | 29.03 | ||
| Proximal depth | 23.32 | ||
| Pedal phalanx | 6C | Length | 27.20.41* |
| Proximal transverse width | 25.17* | ||
| Proximal depth | 27.51 | ||
| Pedal phalanx | 6D | Length | 41.98* |
| Proximal transverse width | 23.98* | ||
| Proximal depth | 30.89 | ||
| Pedal phalanx | 6E | Length | 32.21* |
| Proximal transverse width | 26.15 | ||
| Proximal depth | 32.88 | ||
| Pedal phalanx | 6F | Length | 46.17* |
| Proximal transverse width | 25.43* | ||
| Proximal depth | 19.71* | ||
| Pedal phalanx | 6G | Length | 30.54* |
| Distal transverse width | 25.81 | ||
| Distal depth | 20.64 | ||
| Pedal phalanx | 6H | Length | 25.15* |
| Distal transverse width | 27.37 | ||
| Distal depth | 18.71* | ||
| Pedal phalanx | 6I | Length | 25.86* |
| Proximal transverse width | 28.59 | ||
| Proximal depth | 25.09 | ||
| Pedal phalanx | 6J | Length | 16.87* |
| Proximal transverse width | 23.45 | ||
| Proximal depth | 20.51* | ||
| Pedal phalanx | 6K | Length | 27.67* |
| Distal transverse width | 25.61* | ||
| Distal depth | 20.00* | ||
| Pedal ungual (digit I) | 6L | Length, approximately perpendicular to articular facet | 29.88* |
| Height approximately parallel to articular facet | 26.95* | ||
| Pedal ungual | 6M | Height of articular facet | 26.75 |
| Height of proximal end including flexor tubercle | 29.01* | ||
| Length perpendicular to articular facet | 26.99* | ||
| Greatest transverse width | 13.63 |
Measurements are in mm. For cases in which the total measurement cannot be given because part of the bone is missing, the measurement of the preserved portion is given and marked with an asterisk. For pedal bones, “depth” refers to the dorsoplantar dimension.
A partial neural arch from a cervical vertebra preserves the left zygapophyses and the lateral lamina connecting them (Fig. 3A). The prezygapophysis is not flexed, but little else can be discerned on this fragment.
Figure 3. Vertebrae, scapula, forelimb bones, and pelvic bones of Martharaptor greenriverensis (UMNH VP 21400).

(A)–Partial cervical neural arch, dorsal view. (B–E)–Cranial dorsal centrum in cranial (B), caudal (C), right lateral (D), and left lateral (E) views. (F–G)–Distal caudal centrum in lateral (F) and ventral (G) views. (H)–Possible ulna. (I)–Possible radius. (J–K)–Left scapula in lateral (J) and medial (K) views. (L)–Proximal end of ischium. (M)–Possible distal end of pubis Scale bar = 50 mm. acr = acromium process, ar = acetabular rim, gl = glenoid, hyp = hypapophysis, poz = postzygapophysis, pnp = pneumatopore, prz = prezygapophysis.
The centrum of one cranial dorsal vertebra is preserved (Fig. 3B–E), and it has been transversely flattened post-mortem. The cranial and caudal articular surfaces of the centrum are shallowly concave. The relatively size of the small hypapophysis at the cranial end of the centrum suggests that this vertebra is the first or last hypapophysis-bearing vertebra in the series. The proportions of the centrum suggest that it is more likely the first than the last such vertebra, upon comparison with Falcarius [4] and the oviraptorosaur Nomingia [37]. A distinct pneumatopore is present immediately caudodorsal to the hypapophysis.
The ventral part of another centrum with a nearly flat ventral surface is relatively small, suggesting that it is from the tail (Fig. 3F–G).
The distal end of a bone is interpreted as a radius (Fig. 3I). The distal end is incomplete, but the preserved portion is expanded and flattened, as is usual in a derived coelurosaurian radius. The shaft is round in cross-section.
The partial shaft of another long bone is interpreted as an ulna (Fig. 3H). It is round in cross-section and internally hollow and has been split lengthwise down the middle. It appears slightly bowed, but this may be an optical illusion created by its pattern of missing bone chunks. Its diameter is larger than that of the presumed radius, which is consistent with a theropod ulna.
The distal tip of a manual element, interpreted as metacarpal I from the right hand, is preserved (Fig. 4A). It is ginglymoid and asymmetrical, with the medial condyle extending further distally than the lateral condyle.
Figure 4. Manual bones of Martharaptor greenriverensis (UMNH VP 21400).

(A)–Presumed metacarpal I. (B)–Presumed phalanx I-1. (C)–Penultimate phalanx. (D) Penultimate phalanx. (E)–Unidentified phalanx. (F)–Unidentified phalanx. (G)–Ungual of digit I. (H)–Ungual of digit I. (I)–Ungual of digit II. (J)–Ungual of digit II. (K)–Ungual of digit III. (L)–Ungual of digit III. Scale bar = 50 mm. Numbers on sub-figures refer to proximal (1), distal (2), dorsal (3), palmar (4), and side (5) views; for side views, whether the side is medial or lateral cannot be determined.
The proximal tip of a manual element that may be phalanx I-1 is preserved (Fig. 4B). It fits tightly with the presumed right first metacarpal. The distal tips of four other non-ungual manual phalanges are preserved, one much smaller than the other two (Fig. 4C–F). All three are distally ginglymoid and symmetrical. Two are penultimate phalanges, as indicated by the close spacing of the distal condyles; these two phalanges exhibit a tight fit with manual unguals I and II.
Five nearly complete manual ungual phalanges are preserved, each with its tip broken off (Fig. 4G–L). None of the manual unguals of M. greenriverensis has the proximodorsal “lip” that is found on the manual unguals of some oviraptorosaurian and paravian coelurosaurs [38]. All of the manual unguals of the specimen are similar in overall size, but one is 13–20% taller proximally than the others (Fig. 4G). It is therefore most likely from the first finger. Two of the other nearly complete unguals have a flexor tubercle that is more bulbous than that of the others. Their articular facets are smaller than those of the other unguals, so they are most likely from the third finger. The other two unguals are therefore likely from the second finger. An isolated piece of an ungual from near the tip appears to be from the second finger. A piece of the palmar curve of another ungual matches the curvature of the largest ungual and is therefore probably from the first finger of the other hand. One of the unguals of digit II is missing a piece from the middle, so that the proximal and distal portions are separated by a gap (Fig. 4J).
The left scapula is preserved, with a piece missing from the middle of its shaft (Fig. 3J–K). Its distal end is moderately expanded, as in the therizinosauroids Beipiaosaurus [11] and Suzhousaurus [14]. In Falcarius and other therizinosauroids, breakage prevents assessment of whether a distal expansion is present [2], [20], [21], [23]. The acromion area juts out prominently, like a short hatchet head. Its shape resembles that of the preserved portion of the acromion of Falcarius [4]. As in Falcarius [4] and Beipiaosaurus [11], there is no extension of the glenoid onto the lateral surface of the scapula.
The proximal end of the ischium is preserved (Fig. 3L). It resembles its counterpart in Falcarius [4]. The preserved part of the bone is laterally compressed. Part of the acetabular rim and the cranial edge of the proximal shaft are preserved.
The flattened, expanded end of a bone with a shaft is preserved (Fig. 3). Enough of the shaft is broken off to make it difficult to determine whether the shaft is round or flattened in cross-section. The expanded end is broken into a shape that resembles the deltopectoral crest of a proximal humerus, but this is an artifact. The diameter of the shaft is similar to that of the presumed radius and less than that of the presumed ulna. This is inconsistent with a humerus, the shaft of which is larger in diameter than a radius or ulna. It is possible that this is the distal end of a pubis, in which case the expanded end is a partial pubic boot resembling that of Falcarius [4] more than those of other therizinosauroids [15], [23].
The first metatarsal of the left foot is missing its proximal tip (Fig. 5A). As in most other theropods, including the basal therizinosauroids Falcarius [4] and Beipiaosaurus [9], the proximal end tapers. This differs from the condition in Therizinosauridae, in which the proximal end is robust and articulates with the tarsus [16], [20], [23]. In M. greenriverensis the medial surface of metatarsal I exhibits a distinct facet for articulation with the lateral surface of metatarsal II. The condyles are round in medial and lateral view, and the medial condyle extends slightly further distally than the lateral condyle. The articular surface for phalanx I-1 extends onto the dorsal surface of the metatarsal.
Figure 5. Metatarsals of Martharaptor greenriverensis (UMNH VP 21400).

(A)–Left metatarsal I. (B)–Left metatarsal II. (C)–Presumed left metatarsal III. (D)–Right metatarsal II. (E)–Right metatarsal IV. Scale bar = 50 mm. Numbers on sub-figures refer to proximal (1), distal (2), dorsal (3), plantar (4), medial (5), and lateral (6) views.
The preserved proximal end of the left second metatarsal has a nearly square proximal surface (Fig. 5B). Only the distal end of the right second metatarsal is preserved (fig. 5D). It is a rounded knob, with two distinct condyles only on the plantar surface. Of those, the lateral condyle is much more bulbous than the medial condyle.
The proximal end of another metatarsal (Fig. 5C) exhibits good fit with the proximal end of right metatarsal II, but with so little of the bone preserved it is difficult to be certain that it is the right third metatarsal.
The distal end of the right fourth metatarsal (Fig. 5E) has much of its distal surface eroded, but enough is preserved to tell that the distal end is not ginglymoid and that two distinct condyles are present only on the plantar side. Of these the lateral condyle is more bulbous than the medial condyle. The medial condyle projects much further distally than the lateral condyle, producing a striking asymmetry and a beveling that is much stronger than that of metatarsal II. A small bit of the shaft is preserved, just enough to show that it was constricted distally and that its distal end was not in contact with metatarsal III, both of which character states it shares with Falcarius [4] and the oviraptorosaur Chirostenotes (CMN [Canadian Museum of Nature, Ottawa, Ontario] FV 8538).
Several partial pedal phalanges are preserved (Fig. 6). Three are proximal ends. In each of the three the articular surface is concave and lacks a vertical ridge, which suggests that they represent phalanx 1 of their respective toes. In each of the three the flattened plantar surface suggests a phalanx rather than a metatarsal. In all three the transverse width is consistent with phalanx II-1 or IV-1, too large to be phalanx I-1, and too small to be phalanx III-1, given the size of metatarsal I and distal metatarsal II. The distal ends of three ginglymoid pedal phalanges show slight asymmetry between the condyles. For two of the three, the corresponding proximal end of another phalanx is present that exhibits good fit (see caption to Fig. 6). The proximal ends of two more pedal phalanges are present.
Figure 6. Pedal phalanges of Martharaptor greenriverensis (UMNH VP 21400).

(A–K)–Unidentified phalanges. (L)–Ungual of digit I. (M)–Ungual of unidentified digit (II, III, or IV). Scale bar = 50 mm. Numbers on sub-figures refer to proximal (1), distal (2), dorsal (3), plantar (4), and side (5) views; for side views, whether the side is medial or lateral cannot be determined. Phalanx H articulates well with phalanx J, and phalanx G articulates well with phalanx I.
Only one pedal phalanx preserves its whole length (Fig. 6F). Its dorsal surface and the proximal and distal ends are eroded. Its small size in comparison with the other pedal elements is consistent with its having come from toe IV.
Two pedal unguals are present (Fig. 6L–M); both are laterally compressed and strongly curved, as in other therizinosauroids [12], [18], [22] and unlike other non-paravian theropods. Only the proximal extremity of one is preserved. The other is considerably smaller and exhibits a size consistent with that of the ungual of the first toe. Its tip and its proximal extremity are missing. Its curvature is stronger than that of any of the manual unguals.
In addition to the material described above, the sample includes many bone shards and tiny fragments that are too incomplete to identify.
Phylogenetic Analysis
The phylogenetic analysis found 1444 trees of 1303 steps. For these trees the consistency index is 0.3761, the homoplasy index is 0.6239, the retention index is 0.8136, and the rescaled consistency index is 0.3059. The strict consensus tree places M. greenriverensis within Therizinosauroidea as the sister taxon to Alxasaurus + Therizinosauridae (Fig. 7). Outside Therizinosauroidea the topology of the tree is identical to that found by the study that used the previous draft of the present data matrix [6] and nearly identical to those found by several other recent phylogenetic studies of Coelurosauria [23], [39]–[41]. A decay analysis found low decay indices (Bremer values) of only 1 or 2 for therizinosauroid clades (Fig. 7). This is likely due to the large amount of missing data, a phenomenon that lowers decay indices, in Martharaptor. Decay indices outside Therizinosauroidea are identical to those from the previous study [6].
Figure 7. Phylogenetic position of Martharaptor greenriverensis within Coelurosauria, as found by this study.

Genus names in green are Asian therizinosauroids, and those in red are North American therizinosauroids. Numbers at therizinosauroid clades indicate decay indices (Bremer values).
Synapomorphies of Therizinosauroidea or its sub-clades that are demonstrably present on Martharaptor include: distal scapular expansion; ungual of finger III approximately the same size as that of finger II; and strongly curved hallucal ungual. A full list of synapomorphies of the therizinosauroid clades, as found by this analysis, is given in Appendix S3.
Discussion
Martharaptor greenriverensis adds to the known dinosaurian fauna of the Cedar Mountain Formation. If it is truly therizinosauroid, it also adds to the known diversity of Therizinosauroidea. A greater amount of known skeletal material would yield greater confidence in the phylogenetic placement of Martharaptor, and we acknowledge the possibility that a future phylogenetic study with more skeletal material could recover a different phylogenetic placement for the taxon. However, the known material exhibits no character states that are inconsistent with basal therizinosauroid status. Also, the strong curvature and lateral compression of the pedal unguals is unknown in non-therizinosauroid theropods outside Paraves, and the material lacks typical paravian traits such as flexed cervical prezygapophyses, strongly pendant flexor tubercles on all manual unguals, and ginglymoid metatarsals.
Supporting Information
Character List for Phylogenetic Analysis of Coelurosauria.
(DOC)
Phylogenetic Data Matrix for Phylogenetic Analysis of Coelurosauria.
(DOC)
Synapomorphies of clades within Therizinosauroidea.
(DOC)
Acknowledgments
Martha Hayden and Cari Corbett (North Carolina State University) co-discovered the site. The site was collected under BLM paleontological permit # UT-S-04-011. Volunteers from the Utah Friends of Paleontology helped excavate the site together with Lindsay Zanno and Robert Bakker. Lindsay Zanno and two anonymous reviewers provided input that improved this manuscript.
Funding Statement
No current external funding sources for this study.
References
- 1. Harris JD (1998) Large, Early Cretaceous theropods in North America. N Mex Mus Nat Hist Sci Bull 14: 225–228. [Google Scholar]
- 2. Kirkland JI, Zanno LE, Sampson SD, Clark JM, DeBlieux DD (2005) A primitive therizinosauroid dinosaur from the Early Cretaceous of Utah. Nature 435: 84–87. [DOI] [PubMed] [Google Scholar]
- 3. Zanno LE (2006) The pectoral girdle and forelimb of the primitive therizinosauroid Falcarius utahensis (Theropoda, Maniraptora): analyzing evolutionary trends within Therizinosauroidea. J Vertebr Paleontol 26: 636–650. [Google Scholar]
- 4. Zanno LE (2010) Osteology of Falcarius utahensis (Dinosauria: Theropoda): characterizing the anatomy of basal therizinosaurs. Zool J Linn Soc-Lond 158: 196–230. [Google Scholar]
- 5. Senter P, Kirkland JI, Bird J, Bartlett JA (2011) A new troodontid theropod dinosaur from the Lower Cretaceous of Utah. PLoS ONE 5(12 e14329): 1–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Senter P, Kirkland JI, DeBlieux DD, Madsen S, Toth N (2012) New dromaeosaurids (Dinosauria: Theropoda) from the Lower Cretaceous of Utah, and the evolution of the dromaeosaurid tail. PLoS ONE 7(5 e36790): 1–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Kirkland JI, Britt BB, Whittle CH, Madsen SK, Burge DL (1998) A small coelurosaurian theropod from the Yellow Cat Member of the Cedar Mountain Formation (Lower Cretaceous, Barremian) of eastern Utah. N Mex Mus Nat Hist Sci Bull 14: 239–248. [Google Scholar]
- 8. Kirkland JI, Burge D, Gaston R (1993) A large dromaeosaur (Theropoda) from the Lower Cretaceous of eastern Utah. Hunteria 2(10): 1–16. [Google Scholar]
- 9. Xu X, Tang Z, Wang X (1999) A therizinosauroid dinosaur with integumentary structures from China. Nature 399: 350–354. [Google Scholar]
- 10. Xu X, Cheng Y, Wang X, Chang C (2003) Pygostyle-like structure from Beipiaosaurus (Theropoda, Therizinosauroidea) from the Lower Cretaceous Yixian Formation of Liaoning, China. Acta Geol Sin 77: 294–298. [Google Scholar]
- 11. Xu X, Zheng X, You H (2009) A new feather type in a nonavian theropod and the early evolution of feathers. Proc Nat Acad Sci USA 106: 832–834. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Russell DA, Dong Z (1993) The affinities of a new theropod from the Alxa Desert, Inner Mongolia, People’s Republic of China. Can J Ea Sci 30: 2107–2127. [Google Scholar]
- 13.Dong Z (1979) [The Cretaceous dinosaur fossils in southern China]. In Institute of Vertebrate Paleontology and Paleoanthropology, editors. [Mesozoic and Cenozoic Red Beds in Southern China]. Beijing, Nanjing Institute of Paleontological Science Press. p. 342–345. [in Chinese].
- 14. Li D, Peng C, You H, Lamanna MC, Harris JD, et al. (2007) A large therizinosauroid (Dinosauria: Theropoda) from the Early Cretaceous of northwestern China. Acta Geol Sin 81: 539–549. [Google Scholar]
- 15. Li D, You H, Zhang J (2008) A new specimen of Suzhousaurus megatherioides (Dinosauria: Therizinosauroidea) from the Early Cretaceous of northwestern China. Can J Ea Sci 45: 769–779. [Google Scholar]
- 16.Perle A (1979) [Segnosauridae–a new family of theropods from the Late Cretaceous of Mongolia]. Sovm Sov-Mong Paleontol Eksped Tr 8: 45–55. [in Russian].
- 17. Barsbold R, Perle A (1980) Segnosauria, a new infraorder of carnivorous dinosaurs. Acta Palaeontol Pol 25: 187–195. [Google Scholar]
- 18.Perle A (1981) [A new segnosaurid from the Upper Cretaceous of Mongolia]. Sovm Sov-Mong Paleontol Eksp Tr 15: 50–59. [In Russian].
- 19. Xu X, Zhang X, Sereno P, Zhao X, Kuang X, et al. (2002) A new therizinosauroid (Dinosauria, Theropoda) from the Upper Cretaceous Iren Dabasu Formation of Nei Mongol. Vertebr PalAs 40: 228–240. [Google Scholar]
- 20. Zhang X, Xu X, Zhao X, Sereno P, Kuang X, et al. (2001) A long-necked therizinosauroid dinosaur from the Upper Cretaceous Iren Dabasu Formation of Nei Mongol, People’s Republic of China. Vertebr PalAs 39: 282–290. [Google Scholar]
- 21.Barsbold R (1976) [New information on Therizinosaurus (Therizinosauridae, Theropoda)]. Sovm Sov-Mong Paleontol Eksp Tr 3: 76–92. [in Russian].
- 22. Kirkland JI, Wolfe DG (2001) First definitive therizinosauroid (Dinosauria: Theropoda) from North America. J Vertebr Paleontol 21: 410–414. [Google Scholar]
- 23. Zanno LE, Gillette DD, Albright LB, Titus AL (2009) A new North American therizinosauroid and the role of herbivory in “predatory” dinosaur evolution. Proc Roy Soc B 276: 3505–3511. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Senter P, Robins JR (2010) Hip heights of the gigantic theropod dinosaurs Deinocheirus mirificus and Therizinosaurus cheloniformis, and implications for museum mounting and paleoecology. Bull Gunma Mus Nat Hist 14: 1–10. [Google Scholar]
- 25. Zanno LE, Makovicky PJ (2011) Herbivorous ecomorphology and specialization patterns in theropod dinosaur evolution. Proc Nat Acad Sci USA 108: 232–237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Xu X, Zhao X, Clark JM (2001) A new therizinosaur from the Lower Jurassic Lower Lufeng Formation of Yunnan, China. J Vertebr Paleontol 21: 477–483. [Google Scholar]
- 27. Barrett PM (2009) The affinities of the enigmatic dinosaur Eshanosaurus deguchiianus from the Early Jurassic of Yunnan Province, People’s Republic of China. Palaeontology 52: 681–688. [Google Scholar]
- 28.Swofford DL (2001) PAUP*. Phylogenetic Analysis Using Parsimony (*and Other Methods). Version 4.0b10 for 32-Bit Microsoft Windows. Sunderland, Massachusetts, Sinauer Associates.
- 29.Owen R (1841) Report on British fossil reptiles, part II. In: Report of the eleventh meeting of the British Association for the Advancement of Science. London: John Murray. 60–204.
- 30. Seeley (1887) On the classification of the fossil animals commonly named Dinosauria. P R Soc London 63: 165–171. [Google Scholar]
- 31. Marsh OC (1881) Principal characters of American Jurassic dinosaurs. Part V. Am J Sci 21: 417–423. [Google Scholar]
- 32. von Huene F (1914) Das natürliche System der Saurischia. Centralbl Min Geol Paläontol Abt B 1914: 154–158. [Google Scholar]
- 33.Maleev EA (1954) [New turtle-like reptile in Mongolia]. Dokl Akad Nauk SSSR 87: 131–134 [in Russian].
- 34. Kirkland JI, Madsen SK (2007) The Lower Cretaceous Cedar Mountain Formation, eastern Utah: The view up an always interesting learning curve, in Lund W.R. (ed.) Field Guide to Geological excursions in southern Utah, Geological Society of America Rocky Mountain Section 2007 Annual Meeting, Grand Junction Geological Society, Ut Geol Assoc Publ. 35: 1–108 (CD-ROM). [Google Scholar]
- 35. Suarez MB, Suarez CA, Kirkland JI, Gonzalas LA, Grandstaff DE (2007) Sedimentology, stratigraphy, and depositional environment of the Crystal Geyser Dinosaur Quarry, east-central Utah: Palaios. 22: 513–527. [Google Scholar]
- 36.Greenhalgh BW, Britt BB (2007) Stratigraphy and sedimentology of the Morrison-Cedar Mountain boundary, east-central Utah. In: Willis GC, Hylland MD, Clark DL, Chidsey TC Jr, editors. Central Utah: diverse geology of a dynamic landscape. Salt Lake City: Utah Geological Association. 81–100.
- 37. Barsbold R, Osmólska H, Watabe M, Currie PJ, Tsogbaatar K (2000) A new oviraptorosaur (Dinosauria, Theropoda) from Mongolia: the first dinosaur with a pygostyle. Acta Palaeontol Pol 45: 97–106. [Google Scholar]
- 38. Senter P (2007) A method for distinguishing dromaeosaurid manual unguals from pedal “sickle claws.”. Bull Gunma Mus Nat Hist 11: 1–6. [Google Scholar]
- 39. Zhang F, Zhou Z, Xu X, Wang X, Sullivan C (2008) A bizarre Jurassic maniraptoran from China with elongate ribbon-like feathers. Nature 455: 1105–1108. [DOI] [PubMed] [Google Scholar]
- 40. Hu D, Hou L, Zhang L, Xu X (2009) A pre-Archaeopteryx troodontid theropod from China with long feathers on the metatarsus. Nature 461: 640–643. [DOI] [PubMed] [Google Scholar]
- 41. Choiniere JN, Xu X, Clark JM, Forster CA, Guo Y, et al. (2010) A basal alvarezsauroid theropod from the early Late Jurassic of Xinjiang, China. Science 327: 571–574. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Character List for Phylogenetic Analysis of Coelurosauria.
(DOC)
Phylogenetic Data Matrix for Phylogenetic Analysis of Coelurosauria.
(DOC)
Synapomorphies of clades within Therizinosauroidea.
(DOC)