

Highlights
► First demonstration of post-mitotic fusion of macrophages infected with Candida albicans. ► Post-mitotic fusion was attributed to C. albicans hyphae but not yeast form cells. ► Post-mitotic fusion may inhibit macrophage proliferation and the formation of new uninfected phagocytes.
Keywords: Candida albicans, Macrophage, Phagocytosis, Mitosis, Innate immunology
Abstract
The pathogenicity of the opportunistic human fungal pathogen Candida albicans depends on its ability to inhibit effective destruction by host phagocytes. Using live cell video microscopy, we show here for the first time that C. albicans inhibits cell division in macrophages undergoing mitosis. Inhibition of macrophage cell division is dependent on the ability of C. albicans to form hyphae, as it is rarely observed following phagocytosis of UV-killed or morphogenesis-defective mutant Candida. Interestingly, failed cell division following phagocytosis of hyphal C. albicans is surprisingly common, and leads to the formation of large multinuclear macrophages. This raises question as to whether inhibition of macrophage cell division is another virulence attribute of C. albicans or enables host macrophages to contain the pathogen.
1. Introduction
Candida albicans is a major life-threatening human fungal pathogen. Host defence against systemic infection relies mainly on phagocytosis of Candida by cells of the innate immune system (Gow et al., 2012). Our recent work has studied C. albicans-macrophage interactions using phagocytosis assays and live cell video microscopy coupled with sophisticated image analysis tools (McKenzie et al., 2010; Lewis et al., 2012). Such studies reveal dynamic aspects of the host-pathogen interaction that are not evident from studies based on fixed time point analyses. We have shown, for example, that C. albicans can rarely use non-lytic expulsion as a means to escape from macrophages, leaving both pathogen and host cell viable (Bain et al., 2012) and provided detailed information on how hyphal formation within macrophages leads to macrophage lysis and C. albicans escape (McKenzie et al., 2010). Here we show that infection of macrophages with C. albicans can result in failure of macrophages to complete mitosis. Instead of separating into two individual cells, daughter phagocytes remain together and can fuse to reform a single large macrophage. This intriguing and surprisingly common phenomenon may represent yet another virulence attribute of C. albicans.
2. Description of observation
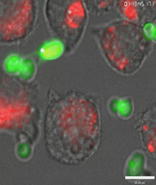
Live cell video microscopy was used to study the dynamics of macrophage mitosis. Mitosis of J774.1 macrophages was observed frequently in the absence of C. albicans. During the 6 h observation period 30.8 ± 5.2% (mean ± s.e.) of macrophages underwent mitosis, and of these only 0.9 ± 0.9% failed to divide into two separate daughter cells. Video 1 shows an example of a J774.1 macrophage that successfully underwent mitosis. When macrophages were cultured with live C. albicans, 29.5 ± 5.7% of macrophages initiated mitosis; however, 35.9 ± 6.1% of 147 mitosis events examined resulted in failed cell division. In Video 2 the macrophage ingests multiple C. albicans, which form hyphae within the macrophage. The host macrophage initiated mitosis, but instead of completely separating, the daughter cells remained fused together. In all cases of failed cell separation, C. albicans hyphae were observed spanning both daughter cells. Macrophages did not persist in their attempt to undergo mitosis indefinitely. Instead macrophages fused back together on average 44.6 ± 3.3 min after initiation of cytokinesis. It is noteworthy that phagocytosis could still occur whilst macrophages were attempting to undergo mitosis.
Macrophages infected with the C. albicans glycosylation mutants, mnt1Δmnt2Δ, mns1Δ and mnn4Δ also underwent frequent post-mitotic fusion. For example, 26.5 ± 3.2% of 134 mitosis events examined failed to complete cell division when J774.1s were cultured with the mnt1Δmnt2Δ O-glycosylation mutant In contrast, when macrophages were cultured with UV-killed C. albicans 23.8% of macrophages underwent mitosis and of these, only 0.7 ± 0.4% of 143 attempts examined resulted in failed cell division. Mitosis was always successful when J774.1 macrophages were cultured with the hyphal morphogenetic-defective mutant efg1Δ. The majority of failed cell divisions observed during this study involved phagocytes that had taken up hyphal C. albicans cells, although it was also rarely seen in macrophages infected with clumps of C. albicans in the yeast phase morphology.
Mitotically active macrophages were often clustered in specific regions of cultures. Video 3 shows an example of four macrophages in close proximity failing to divide after initiating mitosis. One of the macrophages (B) is ruptured prior to completion of mitosis, whereas three other macrophages initiated mitosis, but the daughter cells remained fused together rather than separating into individual cells. All examples of post-mitotic fusion of daughter cells shown in Video 3 involved macrophages infected with hyphal C. albicans.
3. Discussion
This is the first report of inhibition of cell division in macrophages cultured with C. albicans to date. Inhibition of cell division has previously been reported with the human fungal pathogens Cryptococcus neoformans and Candida krusei (Luo et al., 2008; García-Rodas et al., 2011). Interestingly, infection with C. neoformans has also been shown to promote cell cycle progression as a strategy to spread infection (Luo et al., 2012). Failure to complete cell division in macrophages infected with hyphal C. albicans may occur due to difficulties in dividing large cargos, such as hyphae, between daughter cells and our observations suggest that hyphal size may be an important factor in this process. C. albicans hyphae may impede spindle formation or prevent the actin ring from contracting and pinching the macrophage into separate daughter cells during cytokinesis. Some microbial pathogens are known to interfere with cell cycle progression, including Clamydia trachomatis, which can selectively block cytokinesis (Greene and Zhong, 2003). Likewise, C. albicans may either have evolved a specific mechanism to prevent macrophage replication or interfere with this process indirectly due to the enlargement and distension of the phagosome. The observation that C. albicans hyphae spanned both daughter cells in all cases of failed cell separation implies mechanical rather than active inhibition of cytokinesis. Interestingly, non-lytic expulsion of hyphal C. albicans has been observed immediately prior to a host macrophage undergoing mitosis (Bain et al., 2012). It is possible that, in this instance, expulsion of C. albicans may have enabled mitosis and cytokinesis to proceed normally.
Macrophages are pivotal components of the innate immune response to infection with C. albicans and mitosis of tissue-derived macrophages plays an important role in macrophage proliferation in infected tissues. Failure of cell division following mitosis induction may inhibit macrophage proliferation and the formation of new uninfected macrophages. Thus, the ability of C. albicans to impede daughter cell separation may be yet another mechanism employed by C. albicans to aid its survival. An alternative explanation is that this process benefits the host, in that macrophages may sense that they carry dividing fungal cells and inhibit division to prevent spreading of the infection. Interestingly, we observed failed cell division most frequently in clusters of macrophages exposed to and infected with multiple C. albicans hyphae. One may speculate that this may result in macrophage giant cell formation that may help suppress growth of large sized microbial cells. Failed cell division is likely to have profound consequences on the host response to infection with C. albicans and future studies are needed to establish the mechanism by which phagocytosis of fungal cells inhibits macrophage mitosis.
4. Methods
C. albicans strain CAI4-CIp10 (NGY152) was grown in SC-Ura medium at 30oC, as described previously (Lewis et al., 2012). For phagocytosis assays (McPhillips and Erwig, 2009; Mora-Montes et al., 2012), 1 × 106 macrophages in supplemented DMEM medium (Lonza, Slough, UK) were seeded onto glass based Iwaki dishes (VWR, Leistershire, UK) and cultured overnight at 37 °C with 5% CO2. During experiments, medium was replaced with supplemented CO2-independent medium containing 1 μM LysoTracker Red (Invitrogen, Paisley, UK). C. albicans were stained with 1 mg/ml FITC (Sigma, Dorset, UK) in 0.05 M carbonate–bicarbonate buffer (pH 9.6) for 10 min at 20 °C in the dark, washed three times, resuspended in PBS, and added to macrophages at a 3:1 ratio. Video microscopy was conducted at 37 °C with a DeltaVision Core microscope (Applied Precision, Washington, USA) and images were captured at 1 min intervals for 6 h using an EMCCD camera. Movies were visualised and edited using Volocity 5.0 imaging analysis software. Means and standard errors were calculated.
Acknowledgments
L.P.E. is a Scottish Senior Clinical Fellow and acknowledges the support of the Chief Scientist Office (SCD/03). This work was funded by Wellcome Trust Project Grant to L.P.E. 089930. N.A.R.G. was funded by a Wellcome Trust Programme Grant (080088) and an equipment Grant (075470) (for DeltaVision), and by a FP7-2007-2013 Grant (HEALTH-F2-2010-260338-ALLFUN).
Footnotes
Supplementary data associated with this article can be found, in the online version, at http://dx.doi.org/10.1016/j.fgb.2012.05.007.
Appendix A. Supplementary material
{kind=link}
{kind=link}
References
- Bain J.M., Lewis L.E., Okai B., Quinn J., Gow N.A.R., Erwig L.-P. Non-lytic expulsion/exocytosis of Candida albicans from macrophages. Fung. Genet Biol. 2012 doi: 10.1016/j.fgb.2012.01.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- García-Rodas R., González-Camacho F., Rodríguez-Tudela J.L., Cuenca-Estrella M., Zaragoza O. The interaction between Candida krusei and murine macrophages results in multiple outcomes, including intracellular survival and escape from killing. Infect. Immun. 2011;79:2136–2144. doi: 10.1128/IAI.00044-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gow N.A.R., van de Veerdonk F.L., Brown A.J.P., Netea M.G. Candida albicans morphogenesis and host defence: discriminating invasion from colonization. Nat. Rev. Microbiol. 2012 doi: 10.1038/nrmicro2711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greene W., Zhong G. Inhibition of host cell cytokinesis by Chlamydia trachomatis infection. J. Infect. 2003;47:45–51. doi: 10.1016/s0163-4453(03)00039-2. [DOI] [PubMed] [Google Scholar]
- Lewis L.E., Bain J.M., Lowes C., Gillespie C., Rudkin F.M., Gow N.A.R., Erwig L.-P. Stage specific assessment of Candida albicans phagocytosis by macrophages identifies cell wall composition and morphogenesis as key determinants. PLoS Pathog. 2012;8(3):e1002578. doi: 10.1371/journal.ppat.1002578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luo Y., Alvarez M., Xia L., Casadevall A. The outcome of phagocytic cell division with infectious cargo depends on single phagosome formation. PLoS One. 2008;3:e3219. doi: 10.1371/journal.pone.0003219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luo Y., Tucker S.C., Casadevall A. Fc- and complement-receptor activation stimulates cell cycle progression of macrophage cells from G1 to S. J. Immunol. 2012;174:7226–7233. doi: 10.4049/jimmunol.174.11.7226. [DOI] [PubMed] [Google Scholar]
- McKenzie C.G., Koser U., Lewis L.E., Bain J.M., Mora-Montes H.M., Barker R.N. Contribution of Candida albicans cell wall components to recognition by and escape from murine macrophages. Infect. Immun. 2010;78:1650–1658. doi: 10.1128/IAI.00001-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McPhillips K.A., Erwig L.P. Assessment of apoptotic cell phagocytosis by macrophages. Methods Mol. Biol. 2009;559:247–256. doi: 10.1007/978-1-60327-017-5_17. [DOI] [PubMed] [Google Scholar]
- Mora-Montes H.M., McKenzie C., Bain J.M., Lewis L.E., Erwig L.P., Gow N.A.R. Interactions between macrophages and cell wall oligosaccharides of Candida albicans. Methods Mol. Biol. 2012;845:247–260. doi: 10.1007/978-1-61779-539-8_16. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.