

Abstract
Host genetic factors may predict the outcome and treatment response in hepatitis C virus (HCV) infection. Very recently, three landmark genome-wide association studies identified single nucleotide polymorphisms near the interleukin 28B (IL28B) region which were more frequent in responders to treatment. IL28B encodes interferon (IFN)λ3, a type III IFN involved in host antiviral immunity. Favourable variants of the two most widely studied IL28B polymorphisms, rs12979860 and rs8099917, are strong pretreatment predictors of early viral clearance and sustained viral response in patients with genotype 1 HCV infection. Further investigations have implicated IL28B in the development of chronic HCV infection versus spontaneous resolution of acute infection and suggest that IL28B may be a key factor involved in host immunity against HCV. This paper presents an overview about the biological activity and clinical applications of IL28B, summarizing the available data on its impact on HCV infection. Moreover, the potential usefulness of IFNλ in the treatment and natural history of this disease is also discussed.
1. Introduction
More than 20 years after the discovery of the hepatitisC virus (HCV) in 1989, it is now well established that HCV infection affects all countries, leading to a major global health problem that requires widespread active interventions for its prevention and control [1]. Prevalence data estimate that 130–170 million persons, or 2-3% of the world population are infected with HCV [2]. HCV infection is characterized by two distinct outcomes: in about 30% of cases, innate and adaptive immune responses achieve a permanent control of infection, referred to as spontaneous HCV clearance; however, most frequently, the host immune responses fail and chronic infection is established [3]. Chronic hepatitis C is highly heterogeneous in clinical presentation and outcomes. This heterogeneity may be dependent on virus genotype but is also largely related to host factors that have been clearly proven to affect the severity and rapidity of disease progression [4]. The successful eradication of HCV in chronically infected patients, defined as a sustained virological response (SVR), is associated with a reduced risk of disease progression. Currently, pegylated interferon (PEG-IFN) plus ribavirin (RBV) is considered the standard of care for chronic hepatitis C, but the rate of SVR is around 50% in patients with HCV genotype 1, the most common genotype [5, 6]. Because PEG-IFN/RBV therapy is costly and often accompanied by several adverse effects, pre-treatment predictions of those patients who are unlikely to benefit from this regimen enables ineffective treatment to be avoided. Viral load and viral genotypes and the stage of liver disease strongly predict the response to HCV treatment [7, 8]. Moreover, host genetic differences may influence the response to HCV treatment. Recently, through a genome-wide association study (GWAS) of patients infected by genotype 1 HCV, it has been reported that single nucleotide polymorphisms (SNPs) linked to the cytokine IFNλ3 (also known as IL28B) are strongly associated with a response to PEG-IFN/RBV therapy [9–12]. There has subsequently been rapidly increasing data regarding the significance of the IL28B polymorphism not only in response to therapy but also in spontaneous clearance of acute HCV infection. Clinically available tests then made IL28B genotype testing become part of the standard of care. Genetic analysis of the host may thus predict which patients are more likely to respond to treatment, taking into account that IL28B genotype is only one of many factors that can influence response rates to PEG-IFN/RBV therapy in HCV infection and should be interpreted in the context of other clinical factors predicting SVR.
2. The IFNλ Family
IFNλ was identified during the recent years and classified as a new group, type III IFN. The IFNλ gene family is composed of three distinct genes: IFNλ1 (IL29), IFNλ2 (IL28A), and IFNλ3 (IL28B) [13, 14]. The members of this IFN family interact through unique receptors that are distinct from type I (IFNα/β) and type II (IFNγ) IFN receptors. The receptor for type III IFN is composed of the unique IFNλR1 chain, also called IL28Rα and the IL10R2 chain, which is shared with IL10, IL22, and IL26 receptor complexes (Figure 1) [15]. IL28A, IL28B, and IL29 are clustered together on chromosome 19 (19q13.13 region) and are coexpressed together with other type I IFNs (IFNα and IFNβ) by virus-infected cells [13, 14]. IFNλRs are expressed at variable levels on most cell types, and they mediate signalling resulting in induction of many of the same genes that are induced by signalling through IFNα/β Rs [13]. Very interestingly, hepatocytes from liver biopsy specimens have a high IFNλ receptor expression (IL28Rα), while no expression is evidenced on fibroblasts, endothelial cells, adipocytes, or primary central nervous system cells [16–18].
Figure 1.
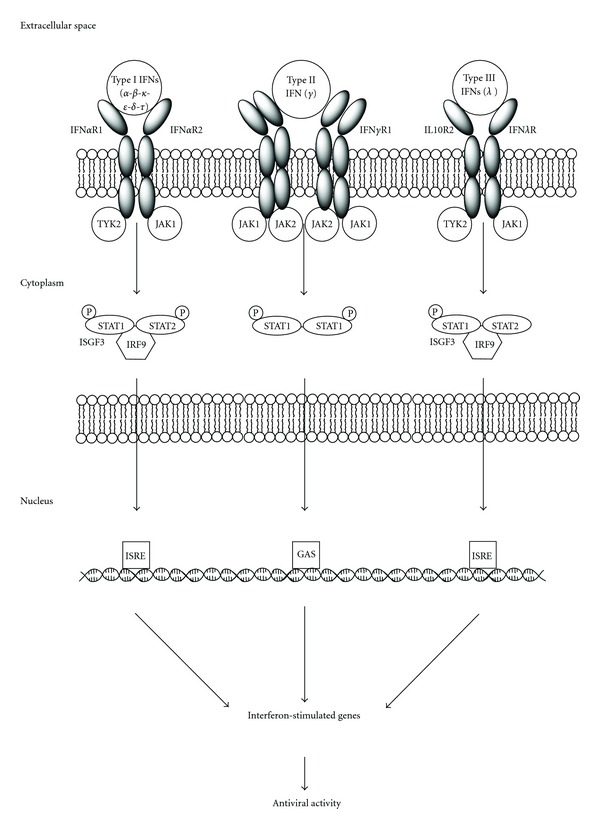
Types I, II, and III IFN receptors and their downstream signalling pathways.
Signalling through type I (IFNα/β) or type III (IFNλ) IFN receptor complexes results in the formation of a transcription factor complex known as IFN-stimulated gene factor 3 (ISGF3), which consists of three proteins: STAT1, STAT2 and IRF-9 (also known as ISGF3γ or p48). After it is fully assembled, ISGF3 translocates to the nucleus where it binds to IFN-stimulated response elements (ISREs) in the promoters of various IFN-stimulated genes classically associated with the antiviral phenotype, including OAS1, MX1, IRF7, and EIF2AK2 [double-stranded RNA-activated protein kinase (PKR)] [19]. The proteins encoded by these genes mediate the antiviral activity induced by the IFNs [20]. As a result, the downstream biological activities induced by either IFNα or IFNλ are very similar, including induction of antiviral and antiproliferative activity in many cell types.
There are currently very limited available data comparing the different biological activity of the three IFNλs. Biological activity ultimately depends on the receptor cytoplasmic domains, which are not related, and these may trigger overlapping but different biological functions. This, in addition to the pattern of receptor distribution among different cell types, means that the different IFNλs are functionally distinct [21]. The effects of IFNλs on immune cell function appear to be complex and diverse, while the antiviral effects are clearer. In fact, IFNλ-induced antiviral activity has been demonstrated against many different viruses but has also been shown to inhibit in vitro replication of HBV and HCV [22–26]. In particular, it has been demonstrated that IFNλ3 inhibits HCV replication in three independent HCV models by the JAK-STAT pathway [27]. The first use of IFNλ in clinical setting has started for hepatitis C: pegylated rHu IFNλ (PEG-IFNλ) has now been evaluated in two phase 1 clinical trials [28, 29]. The initial phase 1A study was designed to evaluate the safety, tolerability, pharmacokinetic, and pharmacodynamic activity of a single dose of PEG-IFNλ in healthy volunteers; a few participants developed reversible, dose-related increases in liver transaminases, but PEG-IFNλ did not induce fever, fatigue, or any overt haematological changes [28]. The phase 1B study has been conducted in patients with chronic genotype 1 HCV infection, mostly non responders to PEG-IFN/RBV therapy; PEG-IFNλ induced significant decreases in the levels of HCV, was well tolerated, and did not induce any significant haematological toxicities such as neutropenia, thrombocytopenia, or anemia [29]. However, this study lacks a direct comparison between IFNλ and IFNα and the influence of viral and patient genotypes, and it is not clear whether the antiviral effects are mediated directly or through the stimulation of immune cells or both. Since the presumed effects of IFNλ on viral clearance observed in the GWAS are mediated through treatment with exogenous IFNα, it is conceivable that although IFNλ may be less potent than IFNα as a direct antiviral, taken together these cytokines may have an additive effect [24].
3. The Effect of IFNλ3 Polymorphism on IFNλ Biology
IL28B/IFNλ3 may harbour the ability to induce potent innate antiviral responses in vitro via signalling through the IL28 receptor complex, whose expression has been verified on a variety of cells, including lymphocytes [24, 30, 31]. Moreover, studies performed in vivo showed that IL28B/IFNλ3 had the potential to induce helper T-cell type 1-biased adaptive cellular immune responses [32]. IFNλ3 has significant influence on antigen-specific CD8+ T-cell function, especially in regards to cytotoxicity, since it is a potent effector of the immune system with special emphasis on CD8+ T-cell killing functions [32].
IFNλ3 is to date the only member of type III interferons with genetic variations associated with differential expression profiles for downstream genes involved in the immune response and outcome for a disease that exhibits symptoms of dysregulation of the immune response. In 2009, three genome-wide association studies reported an association between SVR and two SNPs located near the gene region encoding IFNλ3 (IL28B; rs12980275 and rs8099917) in HCV-infected patients treated with PEG-IFN/RBV combination therapy [9–11]. Further subsequent studies well established that the response to IFNα or the natural clearance of HCV infection is dependent on SNPs, upstream of IFNλ3, which could be used as biomarkers to help determine the treatment outcome [21]. However, the exact mechanisms by which IFNλ3 polymorphisms are identified in the GWAS affect immune function or exert specific antiviral effects in HCV-infected patients are still unclear.
In 2010, two studies performed on HCV-infected patients revealed that both the SNPs of IL28B linked with SVR were strongly associated with lower hepatic expression of interferon-stimulated genes (ISGs) [33, 34]. These results imply that IFNλ3 polymorphism may explain the relationship between hepatic ISG expression and HCV treatment outcome but do not reveal any specific mechanism. However, the increase of infected hepatocyte death rate for specific IFNλ3 polymorphism suggests that an immune-mediated mechanism may be responsible [35].
Further investigations reported some relationships between IL28B genetic variation and allergic disease in children and susceptibility to develop hepatocellular carcinoma in HBV-infected patients [36, 37]. However, even though screening of these polymorphisms and functional studies would be useful to clinical practice for identifying groups at high risk and might help to modify the design of surveillance programs, these studies do not provide any mechanism that explains this association.
In summary, the identity of the functional variants underlying the observed associations is still unknown, and further studies are encouraged to clarify the biological impact of IFNλ3 polymorphism.
4. The Impact of IFNλ3 Polymorphisms on HCV Infection
4.1. rs12979860 Polymorphism
The SNP on chromosome 19q13 (rs12979860) strongly associated with SVR in genotype 1 HCV-infected subjects, identified by the study of Ge et al., results in three possible genotypes: the C/C genotype was associated with 2.5 or greater rate (depending on ethnicity) of SVR compared to the T/T genotype, and the C allele was overrepresented in a random multiethnic population as compared to the chronically infected study cohort, raising the possibility that the C allele may favour spontaneous clearance of HCV [11]. According to this report, the rs12979860 polymorphism also may explain much of the difference in response between different population groups: in fact, the genotype leading to better response presents with greater frequency in European than in African populations [11]. Moreover, the C allele was found associated with more subjective appetite, energy, and sleep complaints, as well as lower serum triglycerides and higher serum LDL cholesterol but less hepatic steatosis [38–40]. This latter report was confirmed by a subsequent study, which showed a lower steatosis severity grade in HCV-infected patients with C/C genotype [41]. This association was then observed in HCV genotypes 2, 3, and 4, and also in HCV/HIV-infected patients even though prior non-responders [42–46]. The C allele also appears to affect positively early viral kinetics in patients with chronic hepatitis C receiving interferon-free treatment [47, 48]. However, the C allele is associated with more pronounced liver histopathology damage in patients chronically infected with HCV genotype 3, which may be secondary to higher viral load but has no impact among genotype 2 infected patients, implying that IL28B may differentially regulate the course of genotype 2 and 3 infection [49, 50]. In an analysis of HIV-infected patients with acute hepatitis C, the C/C genotype was associated with higher serum levels of hepatitis C virus RNA, and lower γGT and CD4 cell count, but not significantly associated with treatment response rates, suggesting that its effects would be different in HIV-infected patients with chronic and acute hepatitis C [51]. The C/C genotype predicts SVR in chronically coinfected patients, but it is also associated with higher all-cause mortality [52, 53]. When this variant was genotyped in HCV cohorts comprised of individuals who spontaneously cleared the virus or had persistent infection, the C/C genotype strongly enhanced resolution of HCV infection amongst individuals of both European and African ancestry, showing that IL28B plays a determinant role in natural clearance of HCV and spontaneous resolution of HCV infection [54]. This report was also observed in women affected by acute hepatitis C [55]. However, rs12979860 homozygosity is not associated with resistance to HCV infection in exposed uninfected patients [56]. Studying the impact of donor and recipient genotypes of IL28B rs12979860C>T SNP on hepatitis C virus (HCV) liver graft reinfection revealed a dominant but not exclusive impact of the donor rather than the recipient genetic background on the natural course and treatment outcome [57]. Interestingly, the risk of developing posttransplant diabetes mellitus is significantly increased in recipients carrying the IL28B rs12979860C>T SNP [58].
4.2. rs8099917 Polymorphism
The study by Rauch et al. demonstrated that the rs8099917 minor allele (T/G or T/T) was associated with progression to chronic HCV infection and also with failure to respond to therapy, with the strongest effects in patients with HCV genotype 1, 2, or 4 [12, 59]. However, an analysis performed on Taiwanese patients with a lower daily viral production rate than Western patients, demonstrated that the T/T genotype may contribute to the increased viral clearance rate and better virological responses in these patients [60]. Even though there are no studies considering multiethnic cohorts, a recent meta-analysis evidenced that rs8099917 T/T had slight predictive value in Asian patients [61]. A further study reported that combination analyses of SNP of rs8099917 in recipient and donor tissues and mutations in HCV RNA allowed prediction of SVR to PEG-IFN/RBV therapy in patients with recurrent HCV infection after orthotopic liver transplantation [62]. Another study revealed the high prevalence of the rs8099917 G allele in HCV/HIV-1-coinfected patients as well as its strong association with treatment failure in HCV genotype 1-infected patients [63]. The rs8099917 T/T genotype is associated with higher levels of apoB-100 and LDL cholesterol in genotype 1-HCV-infected patients [64].
4.3. Combined IFN3λ Polymorphisms
Both favourable genotypes for rs12979860 (C/C) and rs8099917 (T/T) were associated with spontaneous HCV clearance, possibly interacting in synergy with female sex [65]. On the other side, IL28B variants associated with poor response to interferon therapy may predict slower fibrosis progression, especially in patients infected with non-1 HCV genotypes [66].
Concerning HCV genotype 3-infected subjects, IL28B polymorphisms are associated with RVR but not SVR to PEG-IFN therapy [67]; other studies showed that IL28B polymorphisms are strongly associated with the first phase viral decline during PEG-IFN/RBV therapy of chronic HCV infection, irrespective of HCV genotype, and in genotype 1–4 HIV/HCV-coinfected patients [66, 68, 69]. These reports suggest that IL28B polymorphisms could play a role in blocking the production or release of virions in the first phase viral decline [70]. Treatment outcome in HCV-infected patients may be also influenced by viral polymorphisms within the viral core and NS5A proteins, even though it has been clearly demonstrated that IL28B polymorphisms and HCV core amino acid 70 substitutions contribute independently to an SVR to PEG-INF/RBV therapy [71]. Another independent predictor of RVR and final therapeutic outcome is IFNγ inducible protein-10 (IP-10), and the concomitant assessment of pretreatment IP-10 and IL28B-related SNPs has been proposed to improve response or spontaneous clearance prediction [72–74].
When ten SNPs of IL28B were simultaneously analysed in treatment-naïve patients with genotype 1-HCV chronic infection who received PEG-IFN/RBV, rs12979860 resulted as the critical predictor for SVR even in patients without RVR [75]. A further study demonstrated that SNPs rs8099917 and rs12979860 used alone may be useful for predicting the outcome of HCV treatment, and in a rational and cost-effective approach, determination of only one of these two SNPs is sufficient for predicting SVR. Because of the highest predictive SVR associated with rs12979860 C/C compared with the rs8099917 T/T, rs12979860 determination alone is sufficient for predicting interferon response [76, 77], even in HIV/HCV-coinfected patients [78]. However, there is evidence that a significant proportion of heterozygous carriers of the rs12979860 T nonresponder allele can profit with respect to SVR prediction by further determination of the rs8099917 SNPs [79].
When the combined effect of variants of IL28B with human leukocyte antigen C (HLA-C) and its ligands the killer immunoglobulin-like receptors (KIRs), which have previously been implicated in HCV viral control, was studied, prediction of HCV treatment response improved, supporting a role for natural killer (NK) cell activation in PEG-IFN/RBV treatment-induced clearance, partially mediated by IL28B [80]. Nevertheless, there are conflicting reports on the concept that IFNλ3 might directly act on NK cells to modify their functional activity [81, 82].
4.4. IFN3λ Polymorphisms and Directly Acting Antivirals (DAAs)
HCV therapy has been recently updated by the approval of two DAAs against the NS3/4A serine protease for use in genotype 1, the ketoamide inhibitors boceprevir and telaprevir [83, 84]. The impact of IFN3λ polymorphisms on therapy outcome has been evaluated in several phase 3 clinical trials of boceprevir and telaprevir when used in conjunction with PEG-IFN/RBV. The predictive factors of SVR to a regimen of PEG-IFN/RBV/telaprevir therapy was evaluated in 72 Japanese adults infected with HCV genotype 1, showing rs8099917 T/T as significant determinant of SVR [85]. In the Phase 3 ADVANCE trial, the addition of telaprevir to PEG-IFN/RBV improved SVR across all rs12979860 genotypes [86]. The retrospective analyses from the SPRINT II and RESPOND-2 trials suggest that patients with the rs12979860 C/C genotype are highly likely to be treated for just 6 months with PEG-IFN/RBV/boceprevir [87].
Several other classes of DAAs are under development, and it is expected that they will increase SVR rates and decrease the required duration of therapy [88]. The role of IL28B variations in predicting response to triple-therapy regimens including DAAs different from boceprevir and telaprevir has very recently been investigated, but it is difficult to draw firm conclusions owing to the small number of patients in some groups, and additional research is required before final conclusions can be drawn [89]. Finally, the rs12979860 C/C genotype presented a favourable influence on early viral kinetics during treatment with an interferon-free DAA regimen (mericitabine plus danoprevir), even though the importance of IL28B genotype on response to future interferon-free combination DAA regimens remains to be determined with analyses of larger and longer duration studies [47] (refer to the Table 1 for a comprehensive summary of the effects).
Table 1.
Summary of the impact of different IL28B polymorphisms on HCV infection in several conditions (SVR: sustained virological response; RVR: rapid virological response).
| Polymorphism | Genotype | Impact on HCV | Impact on HCV/HIV | Impact on liver graft reinfection |
|---|---|---|---|---|
| rs12979860 | C/C | (i) Higher SVR (genotypes 1, 2, 3, 4) [11, 41–45] (ii) Spontaneous clearance [11] (iii) Early viral kinetics [46, 47] |
(i) Higher SVR [51] (ii) No influence on acute HCV infection [50] (iii) Higher all-cause mortality [52, 53] |
(i) Natural course and treatment outcome dependent on donor rather than recipient genetic background [54] (ii) Higher frequency of posttransplant diabetes mellitus [55] |
|
| ||||
| rs8099917 | T/G or T/T | (i) Treatment failure (genotypes 1, 2, 4) [12, 56] (ii) Increased viral clearance and virological response in Taiwanese patients [57] |
(i) SVR in patients with recurrent HCV infection [58] | |
| G/G | (i) High prevalence [59] (ii) Treatment failure (genotype 1) [60] |
|||
|
| ||||
| Combined | C/C and T/T | (i) Spontaneous clearance [61] (ii) Early viral kinetics [62, 64, 65] (iii) RVR but not SVR (genotype 3) [63] |
(i) Early viral kinetics (genotypes 1, 4) [65] | |
5. Conclusion
The IFNλ3 gene polymorphism closely associates with the natural course and treatment response of chronic hepatitis C in different populations, irrespective of HCV genotype, thus it can be considered an important predictive pretreatment factor. The differential global distribution of IFNλ3 SNPs may explain the observed clinical differences between ethnic groups. The identification of these polymorphisms will provide support for clinical decision making in current standard care. However, in genotype 2/3 patients who achieve RVR or in treatment initiated in acute HCV, IFNλ3 SNPs testing may have less clinical utility. This raises questions about justifying the cost of incorporating IFNλ3 SNPs testing in routine practice in such patients. Furthermore, the effect of the IL28B genotype is not absolute, and it should not be used as a criterion for denying therapy when unfavourable.
Future studies are needed to establish the role of IFNλ3 genotype using direct antivirals, which rapidly reduce the viral load and may therefore lower the influence of IL28B genotyping in predicting SVR. Further functional studies of IFNλs and the significant SNPs should be investigated to improve the positive predictive value using the point mutation analysis of the targeted polymorphisms. For applying a practical tailor-made therapy, it is also necessary to reveal the cause of exceptional cases that do not follow the IL28B genotyping.
Acknowledgments
This work was supported by the Fondazione Banca del Monte di Foggia, Italy.
References
- 1.Lavanchy D. Evolving epidemiology of hepatitis C virus. Clinical Microbiology and Infection. 2011;17(2):107–115. doi: 10.1111/j.1469-0691.2010.03432.x. [DOI] [PubMed] [Google Scholar]
- 2.Shepard CW, Finelli L, Alter MJ. Global epidemiology of hepatitis C virus infection. Lancet Infectious Diseases. 2005;5(9):558–567. doi: 10.1016/S1473-3099(05)70216-4. [DOI] [PubMed] [Google Scholar]
- 3.Rauch A, Rohrbach J, Bochud PY. The recent breakthroughs in the understanding of host genomics in hepatitis C. European Journal of Clinical Investigation. 2010;40(10):950–959. doi: 10.1111/j.1365-2362.2010.02337.x. [DOI] [PubMed] [Google Scholar]
- 4.Alberti A, Vario A, Ferrari A, Pistis R. Review article: chronic hepatitis C—natural history and cofactors. Alimentary Pharmacology and Therapeutics. 2005;22(supplement 2):74–78. doi: 10.1111/j.1365-2036.2005.02602.x. [DOI] [PubMed] [Google Scholar]
- 5.Manns MP, Cornberg M, Wedemeyer H. Current and future treatment of hepatitis C. Indian Journal of Gastroenterology. 2001;20(supplement 1):C47–C51. [PubMed] [Google Scholar]
- 6.Fried MW, Shiffman ML, Reddy KR, et al. Peginterferon alfa-2a plus ribavirin for chronic hepatitis C virus infection. The New England Journal of Medicine. 2002;347(13):975–982. doi: 10.1056/NEJMoa020047. [DOI] [PubMed] [Google Scholar]
- 7.Muir AJ, Bornstein JD, Killenberg PG. Peginterferon alfa-2b and ribavirin for the treatment of chronic hepatitis C in blacks and non-Hispanic whites. The New England Journal of Medicine. 2004;350(22):2265–2271. doi: 10.1056/NEJMoa032502. [DOI] [PubMed] [Google Scholar]
- 8.Kau A, Vermehren J, Sarrazin C. Treatment predictors of a sustained virologic response in hepatitis B and C. Journal of Hepatology. 2008;49(4):634–651. doi: 10.1016/j.jhep.2008.07.013. [DOI] [PubMed] [Google Scholar]
- 9.Tanaka Y, Nishida N, Sugiyama M, et al. Genome-wide association of IL28B with response to pegylated interferon-α and ribavirin therapy for chronic hepatitis C. Nature Genetics. 2009;41(10):1105–1109. doi: 10.1038/ng.449. [DOI] [PubMed] [Google Scholar]
- 10.Suppiah V, Moldovan M, Ahlenstiel G, et al. IL28B is associated with response to chronic hepatitis C interferon-α and ribavirin therapy. Nature Genetics. 2009;41(10):1100–1104. doi: 10.1038/ng.447. [DOI] [PubMed] [Google Scholar]
- 11.Ge D, Fellay J, Thompson AJ, et al. Genetic variation in IL28B predicts hepatitis C treatment-induced viral clearance. Nature. 2009;461(7262):399–401. doi: 10.1038/nature08309. [DOI] [PubMed] [Google Scholar]
- 12.Rauch A, Kutalik Z, Descombes P, et al. Genetic variation in IL28B is associated with chronic hepatitis C and treatment failure: a genome-wide association study. Gastroenterology. 2010;138(4):1338–1345. doi: 10.1053/j.gastro.2009.12.056. [DOI] [PubMed] [Google Scholar]
- 13.Kotenko SV, Gallagher G, Baurin VV, et al. IFN-λs mediate antiviral protection through a distinct class II cytokine receptor complex. Nature Immunology. 2003;4(1):69–77. doi: 10.1038/ni875. [DOI] [PubMed] [Google Scholar]
- 14.Sheppard P, Kindsvogel W, Xu W, et al. IL-28, IL-29 and their class II cytokine receptor IL-28R. Nature Immunology. 2003;4(1):63–68. doi: 10.1038/ni873. [DOI] [PubMed] [Google Scholar]
- 15.Lasfar A, Abushahba W, Balan M, Cohen-Solal KA. Interferon lambda: a new sword in cancer immunotherapy. Clinical and Developmental Immunology. 2011;2011:11 pages. doi: 10.1155/2011/349575.349575 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Witte K, Gruetz G, Volk HD, et al. Despite IFN- receptor expression, blood immune cells, but not keratinocytes or melanocytes, have an impaired response to type III interferons: Implications for therapeutic applications of these cytokines. Genes and Immunity. 2009;10(8):702–714. doi: 10.1038/gene.2009.72. [DOI] [PubMed] [Google Scholar]
- 17.Pagliaccetti NE, Robek MD. Interferon-λ in HCV infection and therapy. Viruses. 2010;2(8):1589–1602. doi: 10.3390/v2081589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Rodríguez JL, Sandoval J, Serviddio G, et al. Id2 leaves the chromatin of the E2F4-p130-controlled c-myc promoter during hepatocyte priming for liver regeneration. Biochemical Journal. 2006;398(3):431–437. doi: 10.1042/BJ20060380. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Donnelly RP, Dickensheets H, O’Brien TR. Interferon-lambda and therapy for chronic hepatitis C virus infection. Trends in Immunology. 2011;32(9):443–450. doi: 10.1016/j.it.2011.07.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Burke JD, Fish EN. Antiviral strategies: the present and beyond. Current Molecular Pharmacology. 2009;2(1):32–39. doi: 10.2174/1874467210902010032. [DOI] [PubMed] [Google Scholar]
- 21.Kelly C, Klenerman P, Barnes E. Interferon lambdas: the next cytokine storm. Gut. 2011;60(9):1284–1293. doi: 10.1136/gut.2010.222976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Coccia EM, Severa M, Giacomini E, et al. Viral infection and toll-like receptor agonists induce a differential expression of type I and λ interferons in humans plasmacytoid and monocyte-derived dendritic cells. European Journal of Immunology. 2004;34(3):796–805. doi: 10.1002/eji.200324610. [DOI] [PubMed] [Google Scholar]
- 23.Robek MD, Boyd BS, Chisari FV. Lambda interferon inhibits hepatitis B and C virus replication. Journal of Virology. 2005;79(6):3851–3854. doi: 10.1128/JVI.79.6.3851-3854.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ank N, West H, Bartholdy C, Eriksson K, Thomsen AR, Paludan SR. Lambda interferon (IFN-λ), a type III IFN, is induced by viruses and IFNs and displays potent antiviral activity against select virus infections in vivo. Journal of Virology. 2006;80(9):4501–4509. doi: 10.1128/JVI.80.9.4501-4509.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Doyle SE, Schreckhise H, Khuu-Duong K, et al. Interleukin-29 uses a type 1 interferon-like program to promote antiviral responses in human hepatocytes. Journal of Hepatology. 2006;44(4):896–906. doi: 10.1002/hep.21312. [DOI] [PubMed] [Google Scholar]
- 26.Marcello T, Grakoui A, Barba-Spaeth G, et al. Interferons α and λ inhibit hepatitis C virus replication with distinct signal transduction and gene regulation kinetics. Gastroenterology. 2006;131(6):1887–1898. doi: 10.1053/j.gastro.2006.09.052. [DOI] [PubMed] [Google Scholar]
- 27.Zhang L, Jilg N, Shao RX, et al. IL28B inhibits hepatitis C virus replication through the JAK-STAT pathway. Journal of Hepatology. 2011;55(2):289–298. doi: 10.1016/j.jhep.2010.11.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Ramos EL. Preclinical and clinical development of pegylated interferon-lambda 1 in chronic hepatitis C. Journal of Interferon and Cytokine Research. 2010;30(8):591–595. doi: 10.1089/jir.2010.0066. [DOI] [PubMed] [Google Scholar]
- 29.Muir AJ, Shiffman ML, Zaman A, et al. Phase 1b study of pegylated interferon lambda 1 with or without ribavirin in patients with chronic genotype 1 hepatitis C virus infection. Journal of Hepatology. 2010;52(3):822–832. doi: 10.1002/hep.23743. [DOI] [PubMed] [Google Scholar]
- 30.Siebler J, Wirtz S, Weigmann B, et al. IL-28A is a key regulator of T-cell-mediated liver injury via the T-box transcription factor T-bet. Gastroenterology. 2007;132(1):358–371. doi: 10.1053/j.gastro.2006.10.028. [DOI] [PubMed] [Google Scholar]
- 31.Dellgren C, Gad HH, Hamming OJ, Melchjorsen J, Hartmann R. Human interferon-λ3 is a potent member of the type III interferon family. Genes and Immunity. 2009;10(2):125–131. doi: 10.1038/gene.2008.87. [DOI] [PubMed] [Google Scholar]
- 32.Morrow MP, Pankhong P, Laddy DJ, et al. Comparative ability of IL-12 and IL-28B to regulate Treg populations and enhance adaptive cellular immunity. Blood. 2009;113(23):5868–5877. doi: 10.1182/blood-2008-11-190520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Honda M, Sakai A, Yamashita T, et al. Hepatic ISG expression is associated with genetic variation in interleukin 28B and the outcome of IFN therapy for chronic hepatitis C. Gastroenterology. 2010;139(2):499–509. doi: 10.1053/j.gastro.2010.04.049. [DOI] [PubMed] [Google Scholar]
- 34.Urban TJ, Thompson AJ, Bradrick SS, et al. IL28B genotype is associated with differential expression of intrahepatic interferon-stimulated genes in patients with chronic hepatitis C. Journal of Hepatology. 2010;52(6):1888–1896. doi: 10.1002/hep.23912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Scott J, Holte S, Urban T, et al. IL28B genotype effects during early treatment with peginterferon and ribavirin in difficult-to-treat hepatitis C virus infection. Journal of Infectious Diseases. 2011;204(3):419–425. doi: 10.1093/infdis/jir264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Gaudieri S, Lucas M, Lucas A, et al. Genetic variations in IL28B and allergic disease in children. PLoS ONE. 2012;7(1, article e30607) doi: 10.1371/journal.pone.0030607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Ren S, Lu J, Du X, et al. Genetic variation in IL28B is associated with the development of hepatitis B-related hepatocellular carcinoma. doi: 10.1007/s00262-012-1203-y. Cancer Immunology, Immunotherapy. In press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Lotrich FE, Loftis JM, Ferrell RE, Rabinovitz M, Hauser P. IL28B polymorphism is associated with both side effects and clearance of hepatitis C during interferon-alpha therapy. Journal of Interferon and Cytokine Research. 2011;31(3):331–336. doi: 10.1089/jir.2010.0074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Li JH, Lao XQ, Tillmann HL, et al. Interferon-lambda genotype and low serum low-density lipoprotein cholesterol levels in patients with chronic hepatitis C infection. Journal of Hepatology. 2010;51(6):1904–1911. doi: 10.1002/hep.23592. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Tillmann HL, Patel K, Muir AJ, et al. Beneficial IL28B genotype associated with lower frequency of hepatic steatosis in patients with chronic hepatitis C. Journal of Hepatology. 2011;55(6):1195–1200. doi: 10.1016/j.jhep.2011.03.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Clark PJ, Thompson AJ, Zhu Q, et al. The association of genetic variants with hepatic steatosis in patients with genotype 1 chronic hepatitis C infection. doi: 10.1007/s10620-012-2171-y. Digestive Diseases and Sciences. In press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Mangia A, Thompson AJ, Santoro R, et al. An IL28B polymorphism determines treatment response of hepatitis C virus genotype 2 or 3 patients who do not achieve a rapid virologic response. Gastroenterology. 2010;139(3):821–827. doi: 10.1053/j.gastro.2010.05.079. [DOI] [PubMed] [Google Scholar]
- 43.Pineda JA, Caruz A, Rivero A, et al. Prediction of response to pegylated interferon plus ribavirin by IL28B gene variation in patients coinfected with HIV and hepatitis C virus. Clinical Infectious Diseases. 2010;51(7):788–795. doi: 10.1086/656235. [DOI] [PubMed] [Google Scholar]
- 44.Sarrazin C, Susser S, Doehring A, et al. Importance of IL28B gene polymorphisms in hepatitis C virus genotype 2 and 3 infected patients. Journal of Hepatology. 2011;54(3):415–421. doi: 10.1016/j.jhep.2010.07.041. [DOI] [PubMed] [Google Scholar]
- 45.Labarga P, Barreiro P, Mira JA, et al. Impact of IL28B polymorphisms on response to peginterferon plus ribavirin in HIV-HCV coinfected patients with prior non-response or relapse. doi: 10.1097/QAD.0b013e3283471d83. AIDS. In press. [DOI] [PubMed] [Google Scholar]
- 46.Asselah T, De MS, Broet P, et al. IL28B polymorphism is associated with treatment response in patients with genotype 4 chronic hepatitis C. Journal of Hepatology. 2012;56(3):527–532. doi: 10.1016/j.jhep.2011.09.008. [DOI] [PubMed] [Google Scholar]
- 47.Chu TW, Kulkarni R, Gane EJ, et al. Effect of IL28B genotype on early viral kinetics during interferon-free treatment of patients with chronic hepatitis C. Gastroenterology. 2012;142(4):790–795. doi: 10.1053/j.gastro.2011.12.057. [DOI] [PubMed] [Google Scholar]
- 48.Naggie S, Osinusi A, Katsounas A, et al. Dysregulation of innate immunity in HCV genotype 1 IL28B unfavorable genotype patients: impaired viral kinetics and therapeutic response. doi: 10.1002/hep.25647. Journal of Hepatology. In press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Rembeck K, Alsio A, Christensen PB, et al. Impact of IL28B-related single nucleotide polymorphisms on liver histopathology in chronic hepatitis C genotype 2 and 3. PLoS ONE. 2012;7(1, article e29370) doi: 10.1371/journal.pone.0029370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Rembeck K, Westin J, Lindh M, Hellstrand K, Norkrans G, Lagging M. Association between IL28B-related genetic variants and liver histopathology differs between hcv genotypes. doi: 10.1002/hep.25598. Journal of Hepatology. In press. [DOI] [PubMed] [Google Scholar]
- 51.Nattermann J, Vogel M, Nischalke HD, et al. Genetic variation in IL28B and treatment-induced clearance of hepatitis C virus in HIV-positive patients with acute and chronic hepatitis C. Journal of Infectious Diseases. 2011;203(5):595–601. doi: 10.1093/infdis/jiq098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Clausen LN, Astvad K, Ladelund S, Larsen MV, Schonning K, Benfield T. Hepatitis C viral load, genotype 3 and IL28B CC genotype predict mortality in HIV and hepatitis C coinfected individuals. doi: 10.1097/QAD.0b013e3283553581. AIDS. In press. [DOI] [PubMed] [Google Scholar]
- 53.Parczewski M, Bander D, Leszczyszyn-Pynka M, Urbanska A, Socha L, Boron-Kaczmarska A. IL28B CC genotype is associated with higher all-cause mortality in antiretroviral-treated HIV-infected patients. doi: 10.1089/AID.2011.0354. AIDS. In press. [DOI] [PubMed] [Google Scholar]
- 54.Thomas DL, Thio CL, Martin MP, et al. Genetic variation in IL28B and spontaneous clearance of hepatitis C virus. Nature. 2009;461(7265):798–801. doi: 10.1038/nature08463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Tillmann HL, Thompson AJ, Patel K, et al. A polymorphism near IL28B is associated with spontaneous clearance of acute hepatitis C virus and jaundice. Gastroenterology. 2010;139(5):1586–1592. doi: 10.1053/j.gastro.2010.07.005. [DOI] [PubMed] [Google Scholar]
- 56.Knapp S, Warshow U, Ho KMA, et al. A polymorphism in IL28B distinguishes exposed, uninfected individuals from spontaneous resolvers of HCV infection. Gastroenterology. 2011;141(1):320–325. doi: 10.1053/j.gastro.2011.04.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Lange CM, Moradpour D, Doehring A, et al. Impact of donor and recipient IL28B rs12979860 genotypes on hepatitis C virus liver graft reinfection. Journal of Hepatology. 2011;55(2):322–327. doi: 10.1016/j.jhep.2010.10.037. [DOI] [PubMed] [Google Scholar]
- 58.Veldt BJ, Duarte-Rojo A, Thompson AJ, et al. Recipient IL28B polymorphism is an important independent predictor of posttransplant diabetes mellitus in liver transplant patients with chronic hepatitis C. American Journal of Transplantation. 2012;12(3):737–744. doi: 10.1111/j.1600-6143.2011.03843.x. [DOI] [PubMed] [Google Scholar]
- 59.Kawaoka T, Hayes CN, Ohishi W, et al. Predictive value of the IL28B polymorphism on the effect of interferon therapy in chronic hepatitis C patients with genotypes 2a and 2b. Journal of Hepatology. 2011;54(3):408–414. doi: 10.1016/j.jhep.2010.07.032. [DOI] [PubMed] [Google Scholar]
- 60.Hsu CS, Hsu SJ, Chen HC, et al. Association of IL28B gene variations with mathematical modeling of viral kinetics in chronic hepatitis C patients with IFN plus ribavirin therapy. Proceedings of the National Academy of Sciences of the United States of America. 2011;108(9):3719–3724. doi: 10.1073/pnas.1100349108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Chen Y, Xu HX, Wang LJ, Liu XX, Mahato RI, Zhao YR. Meta-analysis: IL28B polymorphisms predict sustained viral response in HCV patients treated with pegylated interferon-alpha and ribavirin. Alimentary Pharmacology & Therapeutics. 2012;36(2):91–103. doi: 10.1111/j.1365-2036.2012.05131.x. [DOI] [PubMed] [Google Scholar]
- 62.Fukuhara T, Taketomi A, Motomura T, et al. Variants in IL28B in liver recipients and donors correlate with response to peg-interferon and ribavirin therapy for recurrent hepatitis C. Gastroenterology. 2010;139(5):1577–1585. doi: 10.1053/j.gastro.2010.07.058. [DOI] [PubMed] [Google Scholar]
- 63.Aparicio E, Parera M, Franco S, et al. IL28B SNP rs8099917 is strongly associated with pegylated interferon-α and ribavirin therapy treatment failure in HCV/HIV-1 coinfected patients. PLoS ONE. 2010;5(10, article e13771) doi: 10.1371/journal.pone.0013771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Aizawa Y, Yohizawa K, Aida Y, Ishiguro H, Abe H, Tsubota A. Genotype rs8099917 near the IL28B gene and amino acid substitution at position 70 in the core region of the hepatitis C virus are determinants of serum apolipoprotein B-100 concentration in chronic hepatitis C. Molecular and Cellular Biochemistry. 2012;360(1-2):9–14. doi: 10.1007/s11010-011-1037-5. [DOI] [PubMed] [Google Scholar]
- 65.van den Berg CH, Grady BP, Schinkel J, et al. Female sex and IL28B, a synergism for spontaneous viral clearance in hepatitis C virus (HCV) seroconverters from a community-based cohort. PLoS ONE. 2011;6(11, article e27555) doi: 10.1371/journal.pone.0027555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Bochud PY, Bibert S, Kutalik Z, et al. IL28B alleles associated with poor hepatitis C virus (HCV) clearance protect against inflammation and fibrosis in patients infected with non-1 HCV genotypes. Journal of Hepatology. 2012;55(2):384–394. doi: 10.1002/hep.24678. [DOI] [PubMed] [Google Scholar]
- 67.Moghaddam A, Melum E, Reinton N, et al. IL28B genetic variation and treatment response in patients with hepatitis C virus genotype 3 infection. Journal of Hepatology. 2011;53(3):746–754. doi: 10.1002/hep.24154. [DOI] [PubMed] [Google Scholar]
- 68.Arends JE, Fransen JH, Hoepelman AI, van Baarle D. Association between IL28B polymorphisms and first-phase viral load decrease in chronic hepatitis C virus-infected patients treated with peginterferon alfa-2b/ribavirin. International Journal of Antimicrobial Agents. 2011;38(6):538–539. doi: 10.1016/j.ijantimicag.2011.08.010. [DOI] [PubMed] [Google Scholar]
- 69.Rivero-Juarez A, Camacho EA, Perez-Camacho I, et al. Association between the IL28B genotype and hepatitis C viral kinetics in the early days of treatment with pegylated interferon plus ribavirin in HIV/HCV co-infected patients with genotype 1 or 4. Journal of Antimicrobial Chemotherapy. 2012;67(1):202–205. doi: 10.1093/jac/dkr439. [DOI] [PubMed] [Google Scholar]
- 70.Dahari H, Perelson AS. Hepatitis C virus kinetics in chimeric mice during antiviral therapy. Journal of Hepatology. 2007;46(6):2048–2049. doi: 10.1002/hep.21798. [DOI] [PubMed] [Google Scholar]
- 71.Hayes CN, Kobayashi M, Akuta N, et al. HCV substitutions and IL28B polymorphisms on outcome of peg-interferon plus ribavirin combination therapy. Gut. 2011;60(2):261–267. doi: 10.1136/gut.2010.223495. [DOI] [PubMed] [Google Scholar]
- 72.Lagging M, Askarieh G, Negro F, et al. Response prediction in chronic hepatitis c by assessment of IP-10 and IL28B-related single nucleotide polymorphisms. PLoS ONE. 2011;6(2, article e17232) doi: 10.1371/journal.pone.0017232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Fattovich G, Covolo L, Bibert S, et al. IL28B polymorphisms, IP-10 and viral load predict virological response to therapy in chronic hepatitis C. Alimentary Pharmacology and Therapeutics. 2011;33(10):1162–1172. doi: 10.1111/j.1365-2036.2011.04635.x. [DOI] [PubMed] [Google Scholar]
- 74.Beinhardt S, Aberle JH, Strasser M, et al. Serum level of IP-10 increases predictive value of IL28B polymorphisms for spontaneous clearance of acute HCV infection. Gastroenterology. 2012;142(1):78–85. doi: 10.1053/j.gastro.2011.09.039. [DOI] [PubMed] [Google Scholar]
- 75.Lin CY, Chen JY, Lin TN, et al. IL28B SNP rs12979860 is a critical predictor for on-treatment and sustained virologic response in patients with hepatitis C virus genotype-1 infection. PLoS ONE. 2011;6(3, article e18322) doi: 10.1371/journal.pone.0018322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Halfon P, Bourliere M, Ouzan D, et al. A single IL28B genotype SNP rs12979860 determination predicts treatment response in patients with chronic hepatitis C Genotype 1 virus. European Journal of Gastroenterology & Hepatology. 2011;23(10):931–935. doi: 10.1097/MEG.0b013e328349d0ef. [DOI] [PubMed] [Google Scholar]
- 77.Li S, Hu P, Zhang QQ, et al. Single nucleotide polymorphisms of the IL28B and sustained virologic response of patients with chronic hepatitis C to PEG-interferon/ribavirin therapy: a meta-analysis. Hepatitis Monthly. 2011;11(3):163–172. [PMC free article] [PubMed] [Google Scholar]
- 78.de Castellarnau M, Aparicio E, Parera M, et al. Deciphering the interleukin 28B variants that better predict response to pegylated interferon-alpha and ribavirin therapy in HCV/HIV-1 coinfected patients. PLoS ONE. 2012;7(2, article e31016) doi: 10.1371/journal.pone.0031016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Fischer J, Bohm S, Scholz M, et al. Combined effects of different IL28B gene variants onthe outcome of dual combination therapy in chronic HCV type 1 infection. doi: 10.1002/hep.25582. Journal of Hepatology. In press. [DOI] [PubMed] [Google Scholar]
- 80.Suppiah V, Gaudieri S, Armstrong NJ, et al. IL28B, HLA-C, and KIR variants additively predict response to therapy in chronic hepatitis C virus infection in a European Cohort: a cross-sectional study. PLoS Medicine. 2011;8(9, article e1001092) doi: 10.1371/journal.pmed.1001092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Dring MM, Morrison MH, McSharry BP, et al. Innate immune genes synergize to predict increased risk of chronic disease in hepatitis C virus infection. Proceedings of the National Academy of Sciences of the United States of America. 2011;108(14):5736–5741. doi: 10.1073/pnas.1016358108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Kramer B, Eisenhardt M, Glassner A, et al. Do lambda-IFNs IL28A and IL28B act on human natural killer cells? Proceedings of the National Academy of Sciences of the United States of America. 2011;108(34):E519–E520. doi: 10.1073/pnas.1108850108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Bacon BR, Gordon SC, Lawitz E, et al. Boceprevir for previously treated chronic HCV genotype 1 infection. The New England Journal of Medicine. 2011;364(13):1207–1217. doi: 10.1056/NEJMoa1009482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Zeuzem S, Andreone P, Pol S, et al. Telaprevir for retreatment of HCV infection. The New England Journal of Medicine. 2011;364(25):2417–2428. doi: 10.1056/NEJMoa1013086. [DOI] [PubMed] [Google Scholar]
- 85.Akuta N, Suzuki F, Hirakawa M, et al. Amino acid substitution in hepatitis C virus core region and genetic variation near the interleukin 28B gene predict viral response to telaprevir with peginterferon and ribavirin. Journal of Hepatology. 2010;52(2):421–429. doi: 10.1002/hep.23690. [DOI] [PubMed] [Google Scholar]
- 86.Jacobson IM, McHutchison JG, Dusheiko G, et al. Telaprevir for previously untreated chronic hepatitis C virus infection. The New England Journal of Medicine. 2011;364(25):2405–2416. doi: 10.1056/NEJMoa1012912. [DOI] [PubMed] [Google Scholar]
- 87.Poordad F, Bronowicki JP, Gordon SC, et al. IL28B polymorphism predicts virologic response in patients with hepatitis C genotype 1 treated with boceprevir (Boc) combination therapy. Journal of Hepatology. 2011;54(supplement 1):p. S6. [Google Scholar]
- 88.Welsch C, Jesudian A, Zeuzem S, Jacobson I. New direct-acting antiviral agents for the treatment of hepatitis C virus infection and perspectives. Gut. 2012;61(supplement 1):i36–i46. doi: 10.1136/gutjnl-2012-302144. [DOI] [PubMed] [Google Scholar]
- 89.Lange CM, Zeuzem S. IL28B single nucleotide polymorphisms in the treatment of hepatitis C. Journal of Hepatology. 2011;55(3):692–701. doi: 10.1016/j.jhep.2011.03.006. [DOI] [PubMed] [Google Scholar]