

Abstract
Temozolomide (TMZ) is standard chemotherapy for glioblastoma multiforme (GBM). Intratumoral hypoxia is common in GBM and may be associated with the development of TMZ resistance. Oxygen therapy has previously been reported to potentiate the effect of chemotherapy in cancer. In this study, we investigated whether hyperoxia can enhance the TMZ-induced cytotoxicity of human GBM cells, and whether and how it would resensitize TMZ-resistant GBM cells to TMZ. TMZ-sensitive human GBM cells (D54-S and U87-S) were treated with TMZ to develop isogenic subclones of TMZ-resistant cells (D54-R and U87-R). All cell lines were then exposed to different oxygen levels (1, 21, 40, or 80 %), with or without concomitant TMZ treatment, before assessment of cell cytotoxicity and morphology. Cell death and survival pathways elicited by TMZ and/or hyperoxia were elucidated by western blotting. Our results showed that TMZ sensitivity of both chemo-sensitive and resistant cells was enhanced significantly under hyperoxia. At the cell line-specific optimum oxygen concentration (D54-R, 80 %; U87-R, 40 %), resistant cells had the same response to TMZ as the parent chemosensitive cells under normoxia via the caspase-dependent pathway. Both TMZ and hyperoxia were associated with increased phosphorylation of ERK p44/42 MAPK (Erk1/2), but to a lesser extent in D54-R cells, suggesting that Erk1/2 activity may be involved in regulation of hyperoxia and TMZ-mediated cell death. Overall, hyperoxia enhanced TMZ toxicity in GBM cells by induction of apoptosis, possibly via MAPK-related pathways. Induced hyperoxia is a potentially promising approach for treatment of TMZ-resistant GBM.
Keywords: Glioma, Temozolomide, Chemoresistance, Hypoxia, Apoptosis, MAPK
Introduction
Glioblastoma multiforme (GBM) is a highly malignant primary brain tumor characterized by rapid growth, invasiveness, early recurrence, and resistance to conventional therapy. Overall prognosis is poor; median survival is approximately 12 months [1]. The current regimen for treatment of GBM after maximum surgical resection includes concomitant chemo-radiation using temozolomide (TMZ), followed by six cycles of adjuvant TMZ [2]. Although TMZ can significantly prolong survival, many patients continue to suffer from recurrent disease, because of de-novo or acquired drug resistance [3, 4]. The mechanism underlying TMZ resistance is incompletely understood [5], and, because TMZ is now standard therapy and forms the control arm in clinical trials of other novel agents, understanding and overcoming TMZ resistance is urgent, and is an active area of investigation [6].
The biological effects of hyperoxia have been widely studied for a variety of neurological conditions, for example carbon monoxide poisoning, traumatic brain injury, and ischemic stroke [7–9]. The potential beneficial effects of hyperoxia have been attributed to increased oxygenation within the plasma and brain tissues which can potentially stabilize intracranial pressure, prevent blood–brain barrier disruption, suppress neutrophil–endothelial adhesion, and reduce edema [8, 10]. On the other hand, hyperoxia may activate apoptosis by induction of reactive oxygen species (ROS). Hyperoxia may also interfere with expression of cytokines, growth factors, and transcription factors whereby survival or cell-death pathways are affected to different degrees [11]. Its therapeutic role, therefore, remains controversial.
Intra-tumoral hypoxia is common in GBM, and hypoxic cancer cells are known to be more resistant to radiation or cytotoxic drugs [12, 13]. Conversely, hyperoxia has been shown to potentiate the effect of these treatments by enhancing chemocytotoxicity in vitro [14] and neovascularization in vivo [15]. Although hyperoxia has previously been described for control of tumor growth and progression in glioma, its potential application as an adjunct to chemotherapy has not been investigated [16, 17]. In this study we investigated whether and how different concentrations of environmental oxygen would affect TMZ toxicity in chemosensitive and chemoresistant GBM cells. The hypothesis was that hyperoxia would enhance the effect of TMZ especially in TMZ-resistant GBM cells.
Materials and methods
TMZ-resistant cells and treatment
We have previously described the development of isogenic subclones of TMZ-resistant GBM cells by means of chronic TMZ exposure [18]. Briefly, human GBM cell lines, D54-MG (Duke University Medical Center, USA) and U87-MG (American Type Culture Collection, Manassas, VA, USA), were cultured in Dulbecco’s modified Eagle’s medium (DMEM)/F12 (1:1) and minimum essential medium (MEM)-α, respectively. These were supplemented with 10 % heat inactivated fetal bovine serum (Gibco; Invitrogen, Grand Island, NY, USA). Parent TMZ-sensitive cells (designated D54-S and U87-S) were initially exposed to 100 μM TMZ (Temodal; Schering–Plough, Whitehouse Station, NJ, USA) for two weeks and then continuously to the IC50 of TMZ for 12 months. The TMZ-resistant subclones (D54-R and U87-R) so produced were isolated and maintained in low dose (100 μM) TMZ.
Oxygen treatment
TMZ-sensitive (D54-S and U87-S) and resistant (D54-R and U87-R) cells were exposed to different degrees of oxygen saturation under normobaric conditions (hypoxia, 1 %; normoxia, 21 %; hyperoxia 40 or 80 %), with or without concomitant TMZ for 72 h. Cells were then examined for in-vitro chemosensitivity, apoptosis-related and survival-related protein changes.
Clonogenic assay
Cells were seeded at a density of 500 cells/well on six-well plates. After incubation for 24 h they were treated with TMZ (250, 500, 1,000, or 2,000 μM) for 48 h. Cells without TMZ challenge were used as control cells. Cells were then rinsed with fresh medium and left to form colonies after incubation for 14 days. Colonies were then stained with crystal violet (5 g/L; Sigma–Aldrich, Saint Louis, MO, USA). Percentage inhibition was calculated as the number of colonies formed under the treatment conditions relative to the control. Error bars represented standard deviations from three independent experiments conducted in triplicate. **P < 0.01 and *P < 0.05, versus control.
Cytotoxicity assay
Cytotoxicity was measured by sulforhodamine B (SRB) assay (Sigma–Aldrich). Cells were seeded in 96-well plates (5,000 cells/well) and cultured in the absence or presence of TMZ (250, 500, 1,000, 2,000, and 3,000 μM) for 96 h. Determination of cell density was based on cellular protein content as measured by absorbance (OD) at 490 nm. The percentages of viable cells relative to the controls (cells without previous TMZ treatment) were calculated and plotted. The IC50 values were calculated by derivation of the best-fit line, by use of three independent experiments performed in triplicate.
Western blot
Total protein lysates (30 μg) were separated by 12 % SDS-PAGE using a Mini-Protean electrophoresis cell (Bio-Rad Laboratories, Hercules, CA, USA), and then transferred to a PVDF membrane (0.22 µm; Millipore, Bedford, MA, USA) by use of a Mini Trans-Blot cell (Bio-Rad Laboratories). After blocking, the membrane was probed with one of the following primary antibodies at 1:1,000 dilution for 1 h: rabbit monoclonal antibodies against caspase 3, bax, bcl-2, total p44/42 MAP Kinase (Erk 1/2), and phosphor-p44/42 MAP kinase (Erk1/2) (Thr202/Tyr204) (all from Cell Signaling Technology, Danvers, MA, USA). HRP-conjugated secondary antibodies at 1:10,000 were then used for incubation for 1 h. Immunoreactivity signals were amplified by use of the ECL Plus western blotting detection system (GE Biosciences, Buckinghamshire, UK).
Results
TMZ-resistant GBM cells
After exposing the parent D54-S and U87-S cells to TMZ for 10 months, isogenic TMZ-resistant subclones (D54-R and U87-R) were developed. Clonogenic formation assays were performed after TMZ re-challenge. Compared with the resistant cell lines (D54-R and U87-R), reduced clonogenic formation was observed for D54-S and U87-S 14 days after TMZ treatment (Fig. 1a). Colony formation was significantly inhibited (P < 0.05) in the TMZ-sensitive cell lines upon further challenge with different concentrations of TMZ (250 500, 1,000, and 2,000 μM) (Fig. 1b). On the basis of the percentages of colonies formed relative to the control, IC50 for D54-R and U87-R cells was increased sixfold (IC50 = 2,861.6 μM; P < 0.05) and threefold (IC50 = 1,731.74 μM; P < 0.05) compared with the parent control cells (D54-S, IC50 = 480.8 μM and U87-S, IC50 = 590.1 μM). The results indicated that we had successfully generated two isogenic TMZ-resistant GBM subclones.
Fig. 1.
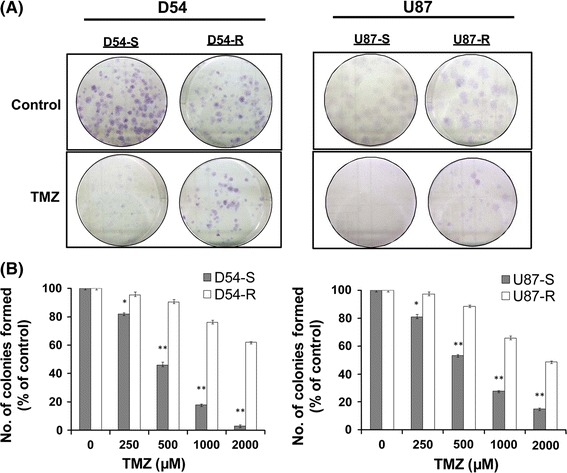
TMZ-resistant D54-R and U87-R cells. (a) Representative dishes from clonogenic assay of the four cells 14 days after TMZ treatment. (b) Clonogenic percentage survival of TMZ-sensitive (D54-S and U87-S) and TMZ-resistant (D54-R and U87-R) cells after rechallenge with TMZ. Each experiment was repeated three times and performed in triplicate. Columns, mean results from triplicate assay; bars, SD; *P < 0.05 and **P < 0.01
Hyperoxia induced cell death synergistically with TMZ
Seeded cells were incubated for 72 h at three oxygen levels (hypoxia, 1 %; normoxia, 21 %; and hyperoxia, 40 or 80 %) with or without concomitant TMZ. In the absence of TMZ, survival of both chemosensitive (D54-S and U87-S) and chemoresistant (D54-R and U87-R) cells increased under hypoxia (1 %), and a mild decrease was observed under cell type-dependent optimum hyperoxic conditions (D54-80 % and U87-40 %) (Fig. 2). With concomitant TMZ given at the IC50 for the parent cells, hyperoxia caused a marked decrease in cell survival, especially for the TMZ-resistant cells. These findings were suggestive of a synergistic effect between hyperoxia and TMZ in inducing cell death. Furthermore, under light microscopy, cells cultured under hyperoxia had altered morphology with increased pseudopod formation (Fig. 2).
Fig. 2.
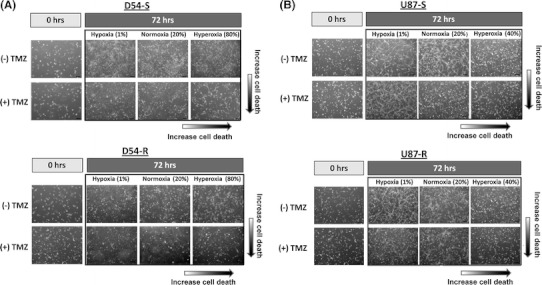
Morphology changed after 72 h treatment at different oxygen saturation with or without concomitant TMZ. a D54, b U87 cells
Hyperoxia enhances chemosensitivity of TMZ-resistant GBM cells
In-vitro chemosensitivity assay showed that cell survival was affected by the ambient oxygen concentration. Figure 3a, b shows that hypoxia had a survival benefit for both sensitive (D54-S and U87-S) and resistant (D54-R and U87-R) cells against TMZ, whereas hyperoxia led to growth inhibition. Figure 3c shows the corresponding IC50 values and their respective ratios compared with the IC50 values under normoxia. Overall, IC50 were highest for both TMZ-sensitive and TMZ-resistant cell lines under hypoxia. Compared with cells under normoxia, the IC50 for D54-S and D54-R cells increased 1.9-fold and 1.4-fold, respectively, under hypoxia. Similarly, the IC50 for U87-S and U87-R cells increased 1.4-fold and 1.1-fold, respectively. Hyperoxia had the opposite effect, resulting in a rapid reduction in cell growth. At 40 % hyperoxia, 1.3 and 2-fold decreases in IC50 were observed for D54-S and U87-S cells, respectively. Similar inhibitory effects were also observed for D54-R and U87-R cells, for which 1.1 and 2.4-fold decreases, respectively, were observed.
Fig. 3.
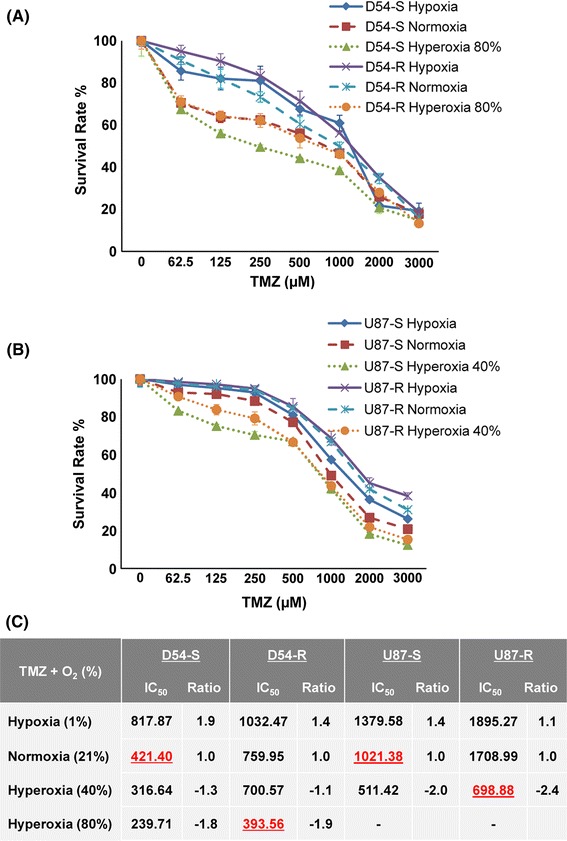
Survival curves for a D54 and b U87 cells under different oxygen saturation conditions. c Alteration of TMZ IC50 values and their respective ratio to normoxia under different oxygen saturation conditions for TMZ-sensitive (D54-S and U87-S) and TMZ-resistant (D54-R and U87-R) cells
This phenomenon of hyperoxia-induced growth inhibition was cell line-dependent (Fig. 3c). D54 cells were less sensitive to 40 % oxygen than U87 cells, as was apparent from the lower ratio compared with normoxia for both sensitive and resistant cells. At 80 % hyperoxia, 1.8 and 1.9-fold decreases in IC50 were observed for D54-S and D54-R cells, respectively, whereas most of the U87 cells died (results not shown). Moreover, hyperoxia was found to ameliorate TMZ resistance in both D54-R and U87-R cells, with a decreases in IC50 from 759.95 μM (D54-R) and 1708.99 μM (U87-R) under 21 % O2, to 393.56 μM (D54-R under 80 % O2) and 698.88 μM (U87-R under 40 % O2), respectively. These were lower than that for the parent cell lines under normoxic conditions. Our findings indicate that the response of GBM to TMZ was significantly affected by environmental oxygen saturation. Hyperoxia may potentiate the anti-tumor effect of TMZ, whereas hypoxia may reduce TMZ sensitivity.
Hyperoxia-induced cell death through caspase-dependent pathways
We then examined if hyperoxia-induced cell death was dependent on pro-apoptotic pathways. Mediators of apoptosis were examined for their altered protein expression under different treatment conditions. Treatment with TMZ or exposure to hyperoxia alone was associated with upregulation of the pro-apoptotic protein Bax, and almost unaltered, if not reduced, expression of the anti-apoptotic protein Bcl-2 (Fig. 4). These resulted in an increased Bax:Bcl-2 ratio, hence cell death. The most significant response was observed after combining TMZ with hyperoxia, which resulted in an increase in caspase 3 activity, known to be the “executioner” of apoptosis. Our findings suggested that the synergistic effects of TMZ and hyperoxia were at least partially mediated via caspase-dependent apoptosis. Interestingly, for D54-R and U87-R cells activation of apoptosis was of similar extent after exposure to hyperoxia with or without concomitant TMZ. The result suggested that hyperoxia alone may be potent enough for attenuation of TMZ resistance.
Fig. 4.
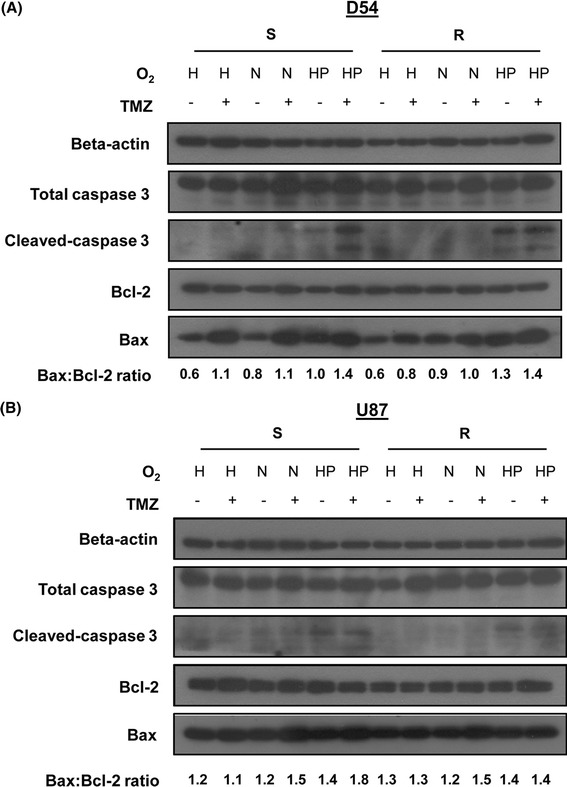
Western blotting shows upregulation of apoptotic mediators. Activation of caspase 3 and increased Bax:Bcl-2 ratio are positively associated with increased oxygen saturation and concomitant TMZ. a D54 and b U87. HP, hyperoxia; N, normoxia; H, hypoxia; S, TMZ-sensitive GBM cells; R, TMZ-resistant GBM cells
Hyperoxia activated the ERK p44/42 MAPK (Erk1/2) signal-transduction pathway
Cellular stress, for example DNA damage, and oxidative stress are known to activate cascades of protein kinases that participate in signal transduction. The MAPK pathway is one of these signaling pathways. We therefore investigated whether activation of ERK1/2 was involved in GBM’s responses to hyperoxia and TMZ. We immunoblotted the total and phosphorylated forms of Erk1/2 proteins under different treatment conditions (Fig. 5). We found upregulation of phosphor-Erk1/2 protein in both the TMZ-sensitive (D54-S and U87-S) and TMZ-resistant (D54-R and U87-R) cells after hyperoxia or TMZ treatment, indicating that both stressful stimuli could activate the Erk1/2 signal transduction pathway. Moreover, Erk1/2 activity was higher in D54-S and U87-S cells than in D54-R and U87-R cells after 72 h, suggesting that TMZ-resistant GBM cells may be less responsive to oxidative and DNA damage stressors. We then investigated the effect of combined hyperoxia–TMZ treatment. For both D54-S and U87-S cells, phosphor-Erk1/2 expression increased in an oxygen-dependent manner when coupled with TMZ treatment (Fig. 5). The findings suggested the MAPK pathway may be important in the hyperoxia-induced and TMZ-induced signal transduction which mediates other death signaling pathways.
Fig. 5.
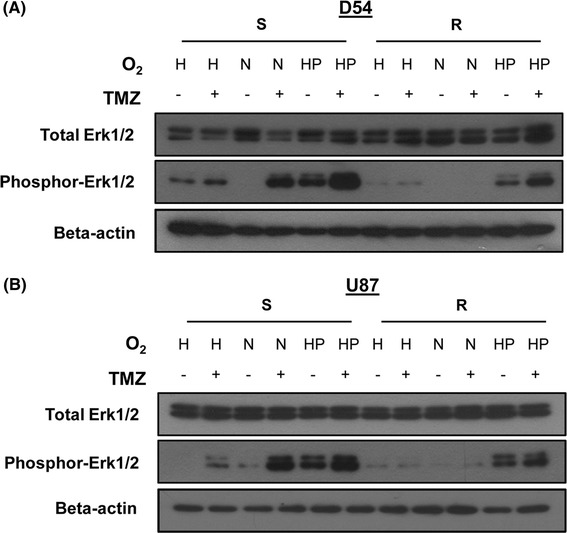
Activation of the Erk1/2 MAPK signal transduction pathway after exposure to hyperoxia and TMZ treatment. Increased phosphorylation of Erk1/2 proteins under hyperoxia and concomitant TMZ conditions as demonstrated by western blotting. a D54 and b U87
Discussion
TMZ, an imidazotetrazine derivative of the alkylating agent dacarbazine, is currently standard therapy for GBM. TMZ is well-tolerated and is therapeutically beneficial because of its ability to methylate DNA at the N-7 or O-6 positions of guanine residues. Despite the positive effect of TMZ on survival, treatment failure because of drug resistance is a significant clinical issue. Several mechanisms underlying TMZ resistance in GBM have been described, including the expression of the repair enzyme O-6-methylguanine-DNA-methyltransferase (MGMT) [19, 20], loss of p53 function [21], selection of pre-existing TMZ-resistant cells [3], mitochondrial adaptive response [22], and dysregulation of glucose transporters and drug metabolism [23]. At present, effective strategies for treatment of TMZ-resistant GBM are lacking. Use of O-6-benzylguanine (O6-BG), an inhibitor of MGMT, has been tested in a phase II trial, with mixed outcomes [24].
GBM is characterized by poorly organized vasculature, hypoxic areas, and necrosis. Hypoxia in tumors is generally associated with increased aggressiveness and resistance to chemotherapy and radiation [12]. In GBM, hypoxia is associated with increased expression of stem-cell markers, and formation of neurospheres and other drug-resistant phenotypes [25]. Pastolatto et al. reported high expression of MGMT (a DNA repair enzyme in GBM stem cells) that was positively correlated with intra-tumoral hypoxia and TMZ resistance [26]. In an attempt to overcome TMZ resistance, inhibition of hypoxia-inducible factors (HIFs), a key regulator of cellular responses to hypoxia, has been conceived as a therapeutic approach for enhancement of chemosensitivity [27, 28]. Inhibitors of HIFs have been studied as chemotherapy adjuncts with therapeutic agents, for example angiogenesis inhibitors and cytotoxic drugs [29, 30]. Shen et al. reported the robust anti-tumor effects of combining HIF-1α inhibition with TMZ in D54 GBM cells [31].
We have adopted a different approach to modulation of hypoxia-mediated chemoresistance—use of induced hyperoxia. Hyperoxia has previously been shown to inhibit cell proliferation [32] and potentiate the effect of chemotherapy by enhancement of cytotoxicity or neovascularization in cancers [33]. Its use as a chemotherapy adjunct has been described for a variety of conditions, for example ovarian [15], breast [34], bone [35], prostate [36], and lung cancers [37]. For brain tumors, hyperoxia in the form of hyperbaric oxygen therapy (HBO) has been shown to increase oxygen level both within and around glioma tissues in patients [38]. Hyperoxia may also promote a reversion from anaerobic metabolism to non-tumorigenic oxidative metabolism [39]. Reported studies on HBO for brain tumors focused mainly on its role as a radiation sensitizer [40, 41] and in the prevention of radiation-induced brain injury [42]. The potential use of hyerpoxia as a chemotherapy adjunct remains unexplored. Stuhr et al. reported reduced tumor growth, reduced vascular density, and changes in gene expression after normobaric and moderate HBO therapy but without concomitant TMZ [16].
As far as we are aware, ours is the first study using hyperoxia as an adjuvant to TMZ therapy for GBM. With an in-vitro model, we demonstrated that normobaric hyperoxia could enhance TMZ cytotoxicity whereas hypoxia had survival benefits. Furthermore, TMZ-resistant cells could regain the same chemo-sensitivity toward TMZ as their parent TMZ-sensitive cells. The potential clinical implication is that hyperoxia may be used to attenuate TMZ resistance in GBM. Regarding the underlying mechanism, our findings suggested that hyperoxia-induced apoptosis was likely to be caspase-dependent. Caspase activation is the irreversible onset of apoptosis [43], and several studies have also described caspase activation as one of the crucial steps in hyperoxia-induced cell death [44, 45]. Our study provides additional information about this phenomenon in brain tumor cells, in that caspase 3 activation was enhanced synergistically under combined hyperoxia–TMZ treatment. Similar changes in Bax:Bcl-2 ratio were also indicative of the importance of pro and anti-apoptotic proteins [46].
We also demonstrated that Erk1/2 MAPK may be critically involved in regulating hyperoxia-induced cell death [47]. The latter may be regulated by a variety of factors [43], for example activation of the protein kinases that participate in signal transduction [48]. Although it is generally believed to be a survival mediator for protecting cells against cell death, Erk1/2 MAPK has been shown to mediate hyperoxic cell death [49, 50]. In our study, the increased phosphorylation of Erk1/2 after exposure to concomitant hyperoxia–TMZ treatment was suggestive of a potentiating effect on cell death. Taken together, our findings indicate that apoptosis and MAPK pathways may be intimately involved in a signaling network under hyperoxia in GBM. The exact mechanism by which this could modulate hyerpoxia-mediated and TMZ-mediated cell death deserves further studies.
This is the first study to investigate the effect of hyperoxia as an adjunct to TMZ in GBM. Combined hyperoxia can significantly potentiate the anti-tumor effect of TMZ. Hyperoxia may attenuate TMZ resistance in GBM via MAPK signaling and apoptotic pathways. Our findings lay the foundation for further in-vivo studies, and have potentially important implications in the treatment of GBM and other cancers in which chemoresistance has a major effect on treatment outcome.
Acknowledgments
This work was supported by small project grant from the University of Hong Kong (project code: 201007176020).
Conflict of interest
The authors have no conflict to report.
Open Access
This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
Abbreviation
- GBM
Glioblastoma multiforme
- HBO
Hyperbaric oxygen
- IC50
Half-maximum inhibitory concentration
- ROS
Reactive oxygen species
- SRB
Sulforhodamine B
- TMZ
Temozolomide
References
- 1.Norden AD, Wen PY. Glioma therapy in adults. Neurologist. 2006;12:279–292. doi: 10.1097/01.nrl.0000250928.26044.47. [DOI] [PubMed] [Google Scholar]
- 2.Stupp R, Hegi ME, Mason WP, van den Bent MJ, Taphoorn MJ, Janzer RC, Ludwin SK, Allgeier A, Fisher B, Belanger K, Hau P, Brandes AA, Gijtenbeek J, Marosi C, Vecht CJ, Mokhtari K, Wesseling P, Villa S, Eisenhauer E, Gorlia T, Weller M, Lacombe D, Cairncross JG, Mirimanoff RO. Effects of radiotherapy with concomitant and adjuvant temozolomide versus radiotherapy alone on survival in glioblastoma in a randomised phase III study: 5-year analysis of the EORTC-NCIC trial. Lancet Oncol. 2009;10:459–466. doi: 10.1016/S1470-2045(09)70025-7. [DOI] [PubMed] [Google Scholar]
- 3.Auger N, Thillet J, Wanherdrick K, Idbaih A, Legrier ME, Dutrillaux B, Sanson M, Poupon MF. Genetic alterations associated with acquired temozolomide resistance in SNB-19, a human glioma cell line. Mol Cancer Ther. 2006;5:2182–2192. doi: 10.1158/1535-7163.MCT-05-0428. [DOI] [PubMed] [Google Scholar]
- 4.de Saldanha da Gama Fischer J, Costa Carvalho P, da Fonseca CO, Liao L, Degrave WM, da Gloria da Costa Carvalho M, Yates JR, Domont GB (2011) Chemo-resistant protein expression pattern of glioblastoma cells (A172) to perillyl alcohol. J Proteome Res 10:153–160 [DOI] [PMC free article] [PubMed]
- 5.Chakravarti A, Palanichamy K. Overcoming therapeutic resistance in malignant gliomas: current practices and future directions. Cancer Treat Res. 2008;139:173–189. [PubMed] [Google Scholar]
- 6.Chinot OL, de La Motte Rouge T, Moore N, Zeaiter A, Das A, Phillips H, Modrusan Z, Cloughesy T. AVAglio: phase 3 trial of bevacizumab plus temozolomide and radiotherapy in newly diagnosed glioblastoma multiforme. Adv Ther. 2011;28:334–340. doi: 10.1007/s12325-011-0007-3. [DOI] [PubMed] [Google Scholar]
- 7.Singhal AB. A review of oxygen therapy in ischemic stroke. Neurol Res. 2007;29:173–183. doi: 10.1179/016164107X181815. [DOI] [PubMed] [Google Scholar]
- 8.Kumaria A, Tolias CM. Normobaric hyperoxia therapy for traumatic brain injury and stroke: a review. Br J Neurosurg. 2009;23:576–584. doi: 10.3109/02688690903050352. [DOI] [PubMed] [Google Scholar]
- 9.Tolias CM, Kumaria A, Bullock MR (2009) Hyperoxia and traumatic brain injury. J Neurosurg 110:607–609; author reply 609–611 [DOI] [PubMed]
- 10.Tatarkova Z, Engler I, Calkovska A, Mokra D, Drgova A, Hodas P, Lehotsky J, Dobrota D, Kaplan P. Effect of long-term normobaric hyperoxia on oxidative stress in mitochondria of the guinea pig brain. Neurochem Res. 2011;36:1475–1481. doi: 10.1007/s11064-011-0473-7. [DOI] [PubMed] [Google Scholar]
- 11.Konsavage WM, Zhang L, Wu Y, Shenberger JS. Hyperoxia-induced activation of the integrated stress response in the newborn rat lung. Am J Physiol Lung Cell Mol Physiol. 2012;302:L27–L35. doi: 10.1152/ajplung.00174.2011. [DOI] [PubMed] [Google Scholar]
- 12.Amberger-Murphy V. Hypoxia helps glioma to fight therapy. Curr Cancer Drug Targets. 2009;9:381–390. doi: 10.2174/156800909788166637. [DOI] [PubMed] [Google Scholar]
- 13.Brat DJ, Mapstone TB. Malignant glioma physiology: cellular response to hypoxia and its role in tumor progression. Ann Intern Med. 2003;138:659–668. doi: 10.7326/0003-4819-138-8-200304150-00014. [DOI] [PubMed] [Google Scholar]
- 14.Stuhr LE, Iversen VV, Straume O, Maehle BO, Reed RK. Hyperbaric oxygen alone or combined with 5-FU attenuates growth of DMBA-induced rat mammary tumors. Cancer Lett. 2004;210:35–40. doi: 10.1016/j.canlet.2004.02.012. [DOI] [PubMed] [Google Scholar]
- 15.Alagoz T, Buller RE, Anderson B, Terrell KL, Squatrito RC, Niemann TH, Tatman DJ, Jebson P. Evaluation of hyperbaric oxygen as a chemosensitizer in the treatment of epithelial ovarian cancer in xenografts in mice. Cancer. 1995;75:2313–2322. doi: 10.1002/1097-0142(19950501)75:9<2313::AID-CNCR2820750921>3.0.CO;2-6. [DOI] [PubMed] [Google Scholar]
- 16.Stuhr LE, Raa A, Oyan AM, Kalland KH, Sakariassen PO, Petersen K, Bjerkvig R, Reed RK. Hyperoxia retards growth and induces apoptosis, changes in vascular density and gene expression in transplanted gliomas in nude rats. J Neurooncol. 2007;85:191–202. doi: 10.1007/s11060-007-9407-2. [DOI] [PubMed] [Google Scholar]
- 17.D’Agostino DP, Olson JE, Dean JB. Acute hyperoxia increases lipid peroxidation and induces plasma membrane blebbing in human U87 glioblastoma cells. Neuroscience. 2009;159:1011–1022. doi: 10.1016/j.neuroscience.2009.01.062. [DOI] [PubMed] [Google Scholar]
- 18.Sun S, Wong TS, Zhang XQ, Pu JK, Lee NP, Day PJ, Ng GK, Lui WM, Leung GK. Protein alterations associated with temozolomide resistance in subclones of human glioblastoma cell lines. J Neurooncol. 2012;107:89–100. doi: 10.1007/s11060-011-0729-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Esteller M, Hamilton SR, Burger PC, Baylin SB, Herman JG. Inactivation of the DNA repair gene O6-methylguanine-DNA methyltransferase by promoter hypermethylation is a common event in primary human neoplasia. Cancer Res. 1999;59:793–797. [PubMed] [Google Scholar]
- 20.Hegi ME, Diserens AC, Gorlia T, Hamou MF, de Tribolet N, Weller M, Kros JM, Hainfellner JA, Mason W, Mariani L, Bromberg JE, Hau P, Mirimanoff RO, Cairncross JG, Janzer RC, Stupp R. MGMT gene silencing and benefit from temozolomide in glioblastoma. N Engl J Med. 2005;352:997–1003. doi: 10.1056/NEJMoa043331. [DOI] [PubMed] [Google Scholar]
- 21.Bocangel DB, Finkelstein S, Schold SC, Bhakat KK, Mitra S, Kokkinakis DM. Multifaceted resistance of gliomas to temozolomide. Clin Cancer Res. 2002;8:2725–2734. [PubMed] [Google Scholar]
- 22.Oliva CR, Nozell SE, Diers A, McClugage SG, 3rd, Sarkaria JN, Markert JM, Darley-Usmar VM, Bailey SM, Gillespie GY, Landar A, Griguer CE. Acquisition of temozolomide chemoresistance in gliomas leads to remodeling of mitochondrial electron transport chain. J Biol Chem. 2010;285:39759–39767. doi: 10.1074/jbc.M110.147504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Le Calve B, Rynkowski M, Le Mercier M, Bruyere C, Lonez C, Gras T, Haibe-Kains B, Bontempi G, Decaestecker C, Ruysschaert JM, Kiss R, Lefranc F. Long-term in vitro treatment of human glioblastoma cells with temozolomide increases resistance in vivo through up-regulation of GLUT transporter and aldo-keto reductase enzyme AKR1C expression. Neoplasia. 2010;12:727–739. doi: 10.1593/neo.10526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Quinn JA, Jiang SX, Reardon DA, Desjardins A, Vredenburgh JJ, Rich JN, Gururangan S, Friedman AH, Bigner DD, Sampson JH, McLendon RE, Herndon JE, 2nd, Walker A, Friedman HS. Phase II trial of temozolomide plus o6-benzylguanine in adults with recurrent, temozolomide-resistant malignant glioma. J Clin Oncol. 2009;27:1262–1267. doi: 10.1200/JCO.2008.18.8417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Bar EE, Lin A, Mahairaki V, Matsui W, Eberhart CG. Hypoxia increases the expression of stem-cell markers and promotes clonogenicity in glioblastoma neurospheres. Am J Pathol. 2010;177:1491–1502. doi: 10.2353/ajpath.2010.091021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Pistollato F, Abbadi S, Rampazzo E, Persano L, Della Puppa A, Frasson C, Sarto E, Scienza R, D’Avella D, Basso G. Intratumoral hypoxic gradient drives stem cells distribution and MGMT expression in glioblastoma. Stem Cells. 2010;28:851–862. doi: 10.1002/stem.518. [DOI] [PubMed] [Google Scholar]
- 27.Li L, Lin X, Staver M, Shoemaker A, Semizarov D, Fesik SW, Shen Y. Evaluating hypoxia-inducible factor-1alpha as a cancer therapeutic target via inducible RNA interference in vivo. Cancer Res. 2005;65:7249–7258. doi: 10.1158/0008-5472.CAN-04-4426. [DOI] [PubMed] [Google Scholar]
- 28.Semenza GL. Targeting HIF-1 for cancer therapy. Nat Rev Cancer. 2003;3:721–732. doi: 10.1038/nrc1187. [DOI] [PubMed] [Google Scholar]
- 29.Lu Y, Li X, Lu H, Fan Z. 1, 9-Pyrazoloanthrones downregulate HIF-1alpha and sensitize cancer cells to cetuximab-mediated anti-EGFR therapy. PLoS One. 2010;5:e15823. doi: 10.1371/journal.pone.0015823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Pourgholami MH, Cai ZY, Badar S, Wangoo K, Poruchynsky MS, Morris DL. Potent inhibition of tumoral hypoxia-inducible factor 1alpha by albendazole. BMC Cancer. 2010;10:143. doi: 10.1186/1471-2407-10-143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Li L, Lin X, Shoemaker AR, Albert DH, Fesik SW, Shen Y. Hypoxia-inducible factor-1 inhibition in combination with temozolomide treatment exhibits robust antitumor efficacy in vivo. Clin Cancer Res. 2006;12:4747–4754. doi: 10.1158/1078-0432.CCR-05-2842. [DOI] [PubMed] [Google Scholar]
- 32.Granowitz EV, Tonomura N, Benson RM, Katz DM, Band V, Makari-Judson GP, Osborne BA. Hyperbaric oxygen inhibits benign and malignant human mammary epithelial cell proliferation. Anticancer Res. 2005;25:3833–3842. [PubMed] [Google Scholar]
- 33.Takiguchi N, Saito N, Nunomura M, Kouda K, Oda K, Furuyama N, Nakajima N. Use of 5-FU plus hyperbaric oxygen for treating malignant tumors: evaluation of antitumor effect and measurement of 5-FU in individual organs. Cancer Chemother Pharmacol. 2001;47:11–14. doi: 10.1007/s002800000190. [DOI] [PubMed] [Google Scholar]
- 34.Moen I, Tronstad KJ, Kolmannskog O, Salvesen GS, Reed RK, Stuhr LE. Hyperoxia increases the uptake of 5-fluorouracil in mammary tumors independently of changes in interstitial fluid pressure and tumor stroma. BMC Cancer. 2009;9:446. doi: 10.1186/1471-2407-9-446. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Kawasoe Y, Yokouchi M, Ueno Y, Iwaya H, Yoshida H, Komiya S. Hyperbaric oxygen as a chemotherapy adjuvant in the treatment of osteosarcoma. Oncol Rep. 2009;22:1045–1050. doi: 10.3892/or_00000534. [DOI] [PubMed] [Google Scholar]
- 36.Kalns J, Krock L, Piepmeier E., Jr The effect of hyperbaric oxygen on growth and chemosensitivity of metastatic prostate cancer. Anticancer Res. 1998;18:363–367. [PubMed] [Google Scholar]
- 37.Petre PM, Baciewicz FA Jr, Tigan S, Spears JR (2003) Hyperbaric oxygen as a chemotherapy adjuvant in the treatment of metastatic lung tumors in a rat model. J Thorac Cardiovasc Surg 125:85–95; discussion 95 [DOI] [PubMed]
- 38.Beppu T, Kamada K, Yoshida Y, Arai H, Ogasawara K, Ogawa A. Change of oxygen pressure in glioblastoma tissue under various conditions. J Neurooncol. 2002;58:47–52. doi: 10.1023/A:1015832726054. [DOI] [PubMed] [Google Scholar]
- 39.Moen I, Oyan AM, Kalland KH, Tronstad KJ, Akslen LA, Chekenya M, Sakariassen PO, Reed RK, Stuhr LE. Hyperoxic treatment induces mesenchymal-to-epithelial transition in a rat adenocarcinoma model. PLoS One. 2009;4:e6381. doi: 10.1371/journal.pone.0006381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Al-Waili NS, Butler GJ. A combination of radiotherapy, nitric oxide and a hyperoxygenation sensitizing protocol for brain malignant tumor treatment. Med Hypotheses. 2007;68:528–537. doi: 10.1016/j.mehy.2005.12.054. [DOI] [PubMed] [Google Scholar]
- 41.Beppu T, Kamada K, Nakamura R, Oikawa H, Takeda M, Fukuda T, Arai H, Ogasawara K, Ogawa A. A phase II study of radiotherapy after hyperbaric oxygenation combined with interferon-beta and nimustine hydrochloride to treat supratentorial malignant gliomas. J Neurooncol. 2003;61:161–170. doi: 10.1023/A:1022169107872. [DOI] [PubMed] [Google Scholar]
- 42.Perez-Espejo MA, Garcia-Fernandez R, Tobarra-Gonzalez BM, Palma-Copete JD, Gonzalez-Lopez A, De la Fuente-Munoz I, Salinas-Ramos J, Felipe-Murcia M, Martinez-Lage JF, Fernandez-Perez J, Romero JM. Usefulness of hyperbaric oxygen in the treatment of radionecrosis and symptomatic brain edema after LINAC radiosurgery. Neurocirugia (Astur) 2009;20:449–453. [PubMed] [Google Scholar]
- 43.Zaher TE, Miller EJ, Morrow DM, Javdan M, Mantell LL. Hyperoxia-induced signal transduction pathways in pulmonary epithelial cells. Free Radic Biol Med. 2007;42:897–908. doi: 10.1016/j.freeradbiomed.2007.01.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Zhang XF, Foda HD. Pulmonary apoptosis and necrosis in hyperoxia-induced acute mouse lung injury. Zhonghua Jie He He Hu Xi Za Zhi. 2004;27:465–468. [PubMed] [Google Scholar]
- 45.Tateda K, Deng JC, Moore TA, Newstead MW, Paine R, 3rd, Kobayashi N, Yamaguchi K, Standiford TJ. Hyperoxia mediates acute lung injury and increased lethality in murine Legionella pneumonia: the role of apoptosis. J Immunol. 2003;170:4209–4216. doi: 10.4049/jimmunol.170.8.4209. [DOI] [PubMed] [Google Scholar]
- 46.O’Reilly MA, Staversky RJ, Huyck HL, Watkins RH, LoMonaco MB, D’Angio CT, Baggs RB, Maniscalco WM, Pryhuber GS. Bcl-2 family gene expression during severe hyperoxia induced lung injury. Lab Invest. 2000;80:1845–1854. doi: 10.1038/labinvest.3780195. [DOI] [PubMed] [Google Scholar]
- 47.Romashko J, 3rd, Horowitz S, Franek WR, Palaia T, Miller EJ, Lin A, Birrer MJ, Scott W, Mantell LL. MAPK pathways mediate hyperoxia-induced oncotic cell death in lung epithelial cells. Free Radic Biol Med. 2003;35:978–993. doi: 10.1016/S0891-5849(03)00494-5. [DOI] [PubMed] [Google Scholar]
- 48.Seger R, Krebs EG. The MAPK signaling cascade. FASEB J. 1995;9:726–735. [PubMed] [Google Scholar]
- 49.Buckley S, Driscoll B, Barsky L, Weinberg K, Anderson K, Warburton D. ERK activation protects against DNA damage and apoptosis in hyperoxic rat AEC2. Am J Physiol. 1999;277:L159–L166. doi: 10.1152/ajplung.1999.277.1.L159. [DOI] [PubMed] [Google Scholar]
- 50.Petrache I, Choi ME, Otterbein LE, Chin BY, Mantell LL, Horowitz S, Choi AM. Mitogen-activated protein kinase pathway mediates hyperoxia-induced apoptosis in cultured macrophage cells. Am J Physiol. 1999;277:L589–L595. doi: 10.1152/ajplung.1999.277.3.L589. [DOI] [PubMed] [Google Scholar]