

Abstract
Nuclear factor (NF)- κB is one of the most important transcription factors that plays a crucial role in the regulation of a wide spectrum of genes involved in modulating the cell cycle, apoptosis, cell growth, angiogenesis, inflammation and the tissue invasiveness of highly malignant cells. NF-κB activity has been found to be constitutively elevated in a number of human tumors from either a haematological or solid origin, such as melanomas. In several studies, NF-κB activation was shown to be an adverse prognostic factor, and in melanoma it was proposed as an event that promotes tumor progression. This study aimed to evaluate whether NF-κB activation in tumor tissues, assessed by the expression of the NF-κB p65 subunit, has an effect on the survival of melanoma patients. The expression of NF-κB was immunohistochemically investigated, and the correlation with survival was analyzed. Furthermore, the immunostaining for p53 and survivin was evaluated, and the relationship of these apoptotic and anti-apoptotic factors with NF-κB expression was analyzed. Kaplan-Meier analysis showed that patients with low levels of NF-κB in the nuclei of tumor cells had a significantly longer survival compared to those with high levels. Multivariate analysis confirmed the predictive value of nuclear NF-κB, showing that its expression maintains significance after the model was adjusted using clinicopathological factors. The results demonstrate the correlation of NF-κB p65 nuclear staining with the disease-specific 5-year survival of melanoma patients and suggest that nuclear NF-κB p65 may be promising as an early independent prognostic factor in patients with primary cutaneous melanoma.
Keywords: melanoma, nuclear factor-κB, p53, survivin, survival biomarkers
Introduction
Nuclear factor (NF)-κB is one of the most important transcription factors that plays an essential role in the regulation of the expression and function of a wide spectrum of genes involved in the modulation of the cell cycle, apoptosis, cell growth, angiogenesis, inflammation and tissue invasiveness of highly malignant cells (1–3). NF-κB is a homodimeric or heterodimeric complex comprising proteins of the Rel-family: p65 (RelA), RelB, c-Rel, p50 (NF-κB1) and p52 (NF-κB2), all of which contain a Rel homology domain in the N-terminal region that mediates dimerization and DNA binding (4). The most commonly detected dimers are p65/p50, p65/p65 and p50/p50. Due to the presence of a strong transcriptional activation domain, p65 is responsible for the majority of NF-κB transcriptional activity (5). Moreover, the p65/p50 heterodimer comprises the major activated form of NF-κB in numerous cell types (4). NF-κB proteins are expressed in most cells, but are normally sequestered in the cytoplasm through binding with the inhibitors of NF-κBs (IκBs) (6). A number of pathways of cell stimulation, such as pro-inflammatory, mutagenic and pro-apoptotic stimuli, lead to the activation of the IκB kinase complex which phosphorylates the IκBs, targeting them for ubiquitination and degradation by the 26S proteosome according to the canonical NF-κB signal transduction pathway (7,8). The released and activated NF-κB complexes are then free to translocate into the nucleus and engage transcriptional programs, binding to the consensus enhancer sequence on the promoters of target genes.
The activation of NF-κB, usually assessed by the presence of nuclear p65, has been observed in a number of human tumors from either a haematological or solid origin, such as melanomas (1). Enhanced nuclear translocation of p65/p50 was found in melanoma cell lines in comparison to normal melanocytes (2). Moreover, other in vitro and in vivo studies have shown that NF-κB activity is up-regulated in dysplastic nevi and lesions of human melanoma when compared to human nevi or melanocytes in normal skin (9–11). Largely due to the central role that NF-κB plays in suppressing apoptosis (7), NF-κB activation appears to promote melanoma progression (12–14). The anti-apoptotic mechanisms are essentially based on the ability of NF-κB to activate the transcription of genes that are able to suppress cell death, such as survivin (2), thus allowing the escape of tumor cells from apoptosis and enhancing their metastatic potential.
Survivin is a member of the inhibitor of apoptosis protein family (15), undetectable in most differentiated normal tissues, but strongly expressed in embryonic and fetal organs. It is implicated in cell division, prevention of apoptosis, cellular stress response and checkpoint mechanisms of genomic integrity (16). It is overexpressed in many human malignancies, and such overexpression is associated with poor prognosis (17–19). Transcription of the survivin gene is inhibited by the p53 tumor suppressor (20), essential in the regulation of cellular response to DNA damage. p53 regulates the expression of various genes that contribute to cell cycle arrest, DNA repair or apoptosis (21–26). Mutations of p53 occur in approximately 50% of cancer types and are generally associated with a worse prognosis as well as a higher resistance to treatment (27). Loss or mutation of p53, in addition to being a possible mechanism responsible for survivin overexpression, appears to directly or indirectly lead to NF-κB activation in melanoma cells (4,28–30).
In this study, the expression of NF-κB, survivin and p53 was immunohistochemically investigated, and the relationship among these factors was analyzed in primary cutaneous melanoma. Since further improvements in melanoma prognosis are likely to come from the development of novel molecular markers, the study aimed to evaluate the prognostic prediction of melanoma by NF-κB expression. The correlation between NF-κB expression and clinicopathological factors of patients was also examined.
Materials and methods
Samples
Archival tissue blocks of sporadic primary cutaneous melanoma were obtained from 70 patients. The patients underwent observation at the Oncologic Hospital ‘Businco’, Cagliari, Italy, and at the Department of Pathology, Cancer Center of Solca, Cuenca, Ecuador, between November 1995 and April 2008, and were selected for further study according to the following criteria: melanoma with vertical growth phase and complete clinical data including follow-up until July 2009. Lymph node status and the presence of metastases were verified by a clinical and pathological examination. This study included a total of 70 stage I–IV melanoma patients, whose clinicopathological characteristics are shown in Table I. The patients included 30 men and 40 women, ranging in age from 12 to 100 years (median 68). The anatomic location of the primary tumor included 18 tumors located in the head and neck, 13 in the trunk, 8 in the upper extremities and 31 in the lower extremities. According to Clark’s classification (31), 4 tumors were level II, 11 level III, 23 level IV and 32 level V. According to the American Joint Committee on Cancer (AJCC) staging system (32), 51 tumors were stages I–II and 19 were stages III–IV. Regarding tumor thickness, 17 tumors were classified as T1–T2 and 53 as T3–T4.
Table I.
Clinicopathological characteristics of 70 cutaneous melanoma patients.
| Characteristics | Patients (n=70) |
|---|---|
| Age at diagnosis (%) | |
| ≤68a years | 37 (53) |
| >68 years | 33 (47) |
| Gender (%) | |
| Men | 30 (43) |
| Women | 40 (57) |
| Tumor thickness (%) | |
| T1–T2 | 17 (24) |
| T3–T4 | 53 (76) |
| Clark level (%) | |
| II–III | 15 (21) |
| IV–V | 55 (79) |
| Stage (%) | |
| I–II | 51 (73) |
| III–IV | 19 (27) |
| Anatomic location (%) | |
| Head and neck | 18 (26) |
| Trunk | 13 (19) |
| Upper extremities | 8 (11) |
| Lower extremities | 31 (44) |
Median value.
Following surgical resection, each tumor was fixed in formalin and completely embedded in multiple paraffin blocks. The sections removed from the block with the largest tumor thickness were evaluated. Tumoral areas were identified on haematoxylin and eosin-stained sections and on adjacent sections immunohistochemically stained for melanoma-associated antigens, including S-100 protein, melan A and HMB-45. An independent histopathological analysis was performed by two pathologists (M.P. and J.U.) on separate occasions. The study protocol was approved by the Research Ethics Committee of our institutions, and informed consent was obtained from all of the patients involved in the study.
Immunohistochemistry
Serial microtome sections (5-μm) were treated for the immunohistochemical demonstration of NF-κB (p65), survivin, p53 and melanoma-associated antigens S-100, melan A and HMB-45, using the alkaline phosphatase-streptavidin method.
Antigen retrieval was performed by heating at 95°C for 40 min in 10 mM citrate buffer solution (pH 6.0), followed by gradual cooling for 20 min for the demonstration of NF-κB (p65), survivin, p53 and melan A, and by immersion in 0.1% trypsin solution in phosphate-buffered saline at 37°C for 5 or 10 min for the S-100 protein and the HMB-45 antigen, respectively. Non-specific binding was blocked with 10% normal goat or normal horse serum for 45 min. Mouse monoclonal antibodies to human p65 (clone F-6, 1:100 dilution; Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA), to human p53 (clone DO-7, 1:50 dilution; Dakopatts, Glostrup, Denmark), to human melan A (clone A103, 1:100 dilution; Dakopatts) and to human HMB-45 (clone HMB-45, 1:100 dilution; Dakopatts), rabbit polyclonal antibodies to recombinant human survivin protein (1:2,000 dilution; Novus Biologicals, Littleton, CO, USA) and to bovine S-100 protein (1:1,000 dilution; Dakopatts) were used as primary antisera. Biotinylated anti-mouse and anti-rabbit immunoglobulins G (1:800 and 1:200 dilution, respectively; Vector Laboratories, Burlingame, CA, USA) were used as secondary antisera. The sections were further incubated in alkaline phosphatase-streptavidin (1:1,000 dilution; Vector Laboratories) for 30 min at room temperature, reacted with Fast Red Substrate System (Dakopatts) and counterstained with Mayer’s haematoxylin. Sections from human prostate gland were used as positive controls for NF-κB, sections of rat testis as positive controls for survivin and sections of melanoma strongly expressing p53 as positive controls for p53. Negative controls were established by replacing the primary antibodies with normal serum.
Evaluation of immunoreactivity
To identify and count tumor cells positive for NF-κB in the nucleus, cytoplasm or both, the entire tumor of each case was microscopically examined through ×200 magnification fields with a 144-intersection point square reticulum (0.78 mm2) inserted in the eyepiece. The average of these counts per field was considered. Similarly, adjacent sections from the same samples were evaluated for survivin and p53 immunoreactivity.
The results were stratified according to a staining score that considered the percentage of positive cells as well as the staining intensity. Cases in which NF-κB cytoplasmic staining was detected in >10% of tumor cells with a moderate/strong staining intensity were considered to show NF-κB overexpression in the cytoplasm. Regarding NF-κB nuclear staining, >5% positive cells were used with staining intensity from weak to strong as a cut-off, indicating NF-κB nuclear immunopositivity. The samples were scored as positive for the expression of survivin or p53 when >10% of tumor cells showed a moderate/strong staining intensity; otherwise, the samples were scored as negative.
Statistical analysis
Data were computed with the Statistical Package for the Social Sciences (SPSS) 15.0 software. Correlations between NF-κB, survivin and p53 expression, and that of NF-κB expression with clinicopathological parameters of stages I–IV melanoma patients were assessed by Fisher’s exact or Pearson’s χ2 test.
Overall survival of patients with stages I and II melanoma (51 samples) was calculated from the date of histological diagnosis to the date the patients succumbed to melanoma or the last follow-up, until July 2009. Data on patients who succumbed to other causes were censored at the time of death. Survival curves were obtained using the Kaplan-Meier method, and comparisons were made using the log-rank test and adjusted for specific prognostic factors. The 95% confidence intervals (95% CI) for survival were calculated and reported. Multivariate analysis was performed using the Cox proportional hazard model. The tests used were two-sided. Differences were considered statistically significant at P≤0.05.
Results
Immunohistochemistry
Immunoreactivity for NF-κB was observed in the nuclear and cytoplasmic compartment of the tumor cells, with nuclear staining localized in a small amount of cells with respect to cytoplasmic staining. The immunostaining pattern in the nuclei and cytoplasm was heterogeneous, with the cell clusters showing a stronger staining intensity than other cells in some cases since in other samples positive cells were homogeneously distributed throughout the tumor with an intense immunostaining. In normal epidermis, NF-κB was expressed in a cytoplasmic as opposed to a nuclear pattern. In some cases, the percentage of positive cells decreased from the edge towards the central area of tumor nodules. A total of 29% of the samples showed immunoreactivity for NF-κB in the nuclei, and 54% exhibited NF-κB overexpression in the cytoplasm of tumor cells. An intense immunoreactivity adjacent to the nucleus, leaning against the nuclear envelope, was observed in a number of tumors (46%). A total of 50% of the samples showed this staining and/or the immunoreaction localized inside the nucleus and were scored as positive for nuclear NF-κB (Fig. 1).
Figure 1.
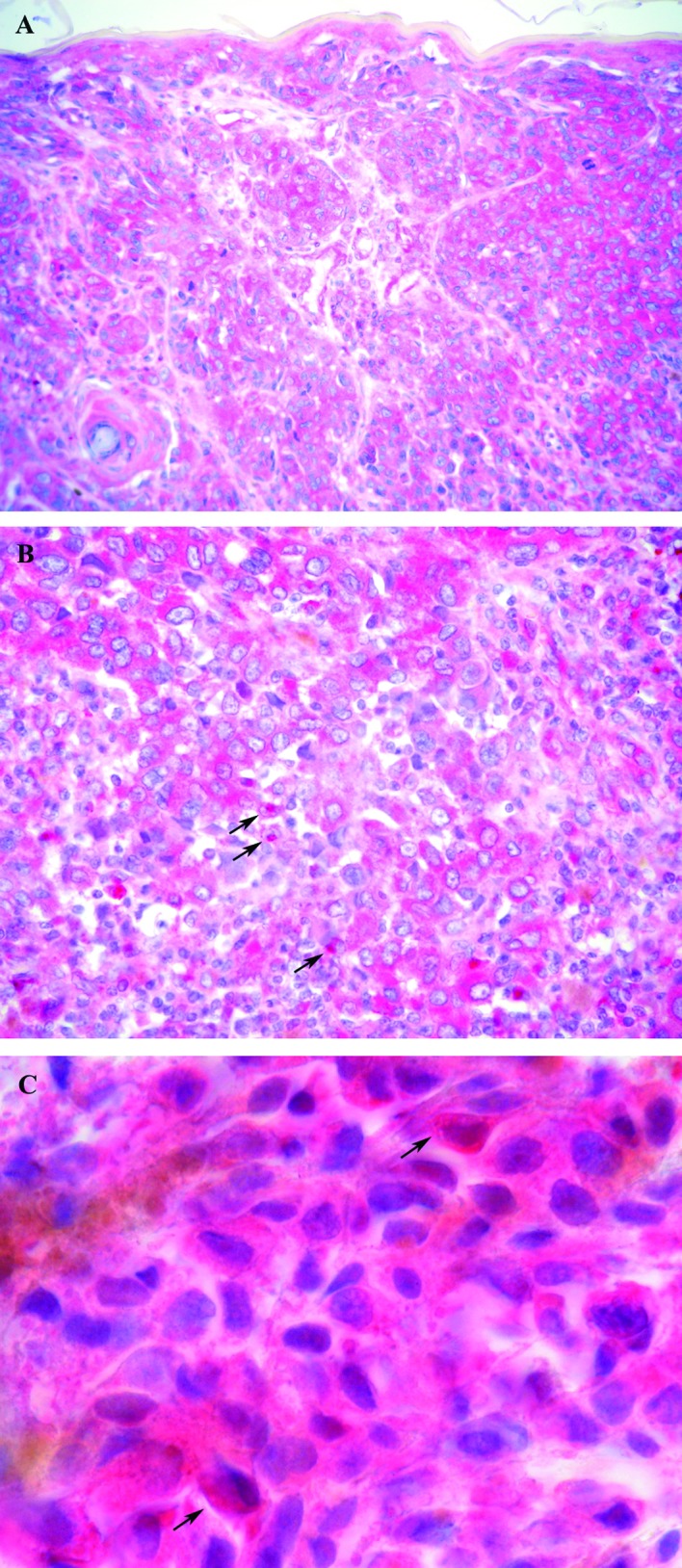
NF-κB immunohistochemical expression was observed in the cytoplasm (A and B), adjacent to the nucleus (B, arrows) and in the nuclei of tumor cells (C, arrows). Original magnification, ×200 (A); ×400 (B) and ×1,000 (C).
p53 expression, restricted to the nuclei of tumor cells, was found in 63% of cases. p53-positive cells were distributed homogeneously within the tumor with a staining intensity from moderate to strong. Positive cells were also observable in the basal epidermal layers (Fig. 2A).
Figure 2.
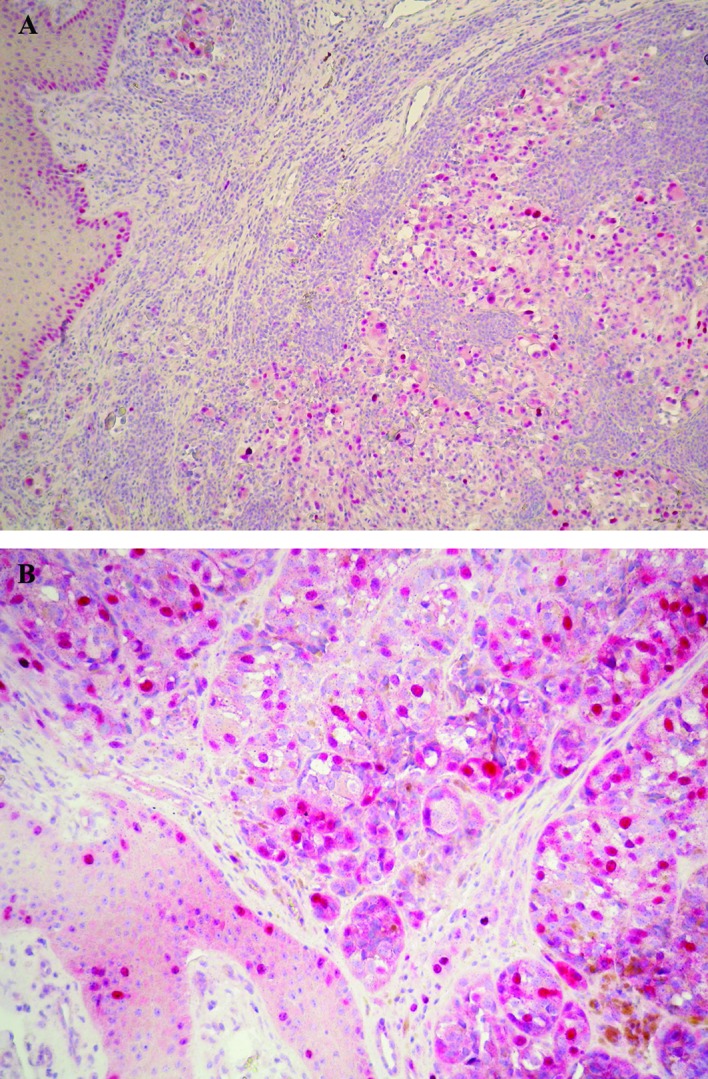
Strong nuclear immunoreactivity for p53 (A) and nuclear or cytoplasmic expression of survivin (B) were detected throughout the tumoral area and in the basal epidermal layers. Original magnification, ×100 (A) and ×200 (B).
Survivin immunoreaction was found in the nucleus and cytoplasm, with the nuclear staining being more intense. Survivin immunopositivity was noted in 59% of the cases in nuclear and/or cytoplasmic staining with positive cells distributed homogeneously throughout the tumor in some cases or with a heterogeneous staining pattern in other cases. A number of mitotic figures were intensely stained, and the epidermis surrounding the tumor also showed survivin-positive cells (Fig. 2B).
Statistical analysis
Positive immunoreactivity for NF-κB in the nuclei or cytoplasm of tumor cells was more frequent in cases positive for p53 and survivin. When analyzed by Fisher’s exact test, NF-κB positivity in the nuclei was significantly associated with p53 overexpression in tumor cells (P=0.025), but not with survivin immunoreactivity (P>0.05). Although tumors with NF-κB overexpression in the cytoplasm were positive for p53 and survivin, the differences were not statistically significant (P>0.05). When a combined nuclear and cytoplasmic NF-κB staining was evaluated, i.e., tumor cells were considered positive if they exhibited nuclear and/or a cytoplasmic immunoreaction, total NF-κB showed a statistical association with p53 (P=0.004) and survivin overexpression (P=0.045). Table II shows NF-κB expression in relation to p53 and survivin.
Table II.
NF-κB expression in relation to p53 and survivin.
| Nuclear NF-κB (no. of cases) | Cytoplasmic NF-κB (no. of cases) | Total NF-κB (no. of cases) | |||||||
|---|---|---|---|---|---|---|---|---|---|
|
|
|
|
|||||||
| Negative | Positive | P-valuea | Negative | Positive | P-valuea | Negative | Positive | P-valuea | |
| p53 | 0.025 | 0.143 | 0.004 | ||||||
| Negative | 18 | 8 | 15 | 11 | 15 | 11 | |||
| Positive | 17 | 27 | 17 | 27 | 9 | 35 | |||
| Survivin | 0.145 | 0.469 | 0.045 | ||||||
| Negative | 18 | 11 | 15 | 14 | 14 | 15 | |||
| Positive | 17 | 24 | 17 | 24 | 10 | 31 | |||
Fisher’s exact test.
Bold, significant p-values.
Kaplan-Meier univariate analysis showed that patients with low levels of NF-κB in the nuclei of tumor cells (nuclear NF-κB-negative) had a significantly longer survival compared to those with high levels (nuclear NF-κB-positive) (P=0.004; Fig. 3). The Kaplan-Meier estimates of overall survival probabilities at 60 months were 0.92 (95% CI 0.83–1.00) for patients with nuclear NF-κB-negative tumors (low nuclear NF-κB expression) and 0.45 (95% CI 0.24–0.66) for those with nuclear NF-κB-positive tumors (high expression) (Table III). Multivariate Cox regression analysis showed that the predictive value of nuclear NF-κB expression maintained significance after the model was adjusted using clinicopathological factors, such as age, gender, anatomic location, tumor thickness, Clark level and AJCC stage (P<0.05). Patient survival was also associated with total NF-κB expression (P=0.04) since the Kaplan-Meier estimates of overall survival probabilities at 60 months were 0.90 (95% CI 0.78–1.00) for patients with a low total NF-κB expression and 0.62 (95% CI 0.45–0.80) for those with a high expression (Table III). This predictive value maintained significance even after adjustment with the clinicopathological factors (P<0.05). In the correlation analysis, nuclear and total NF-κB positivity was significantly associated with advanced tumor stage (P=0.03 and 0.002, respectively), but not with other clinicopathological factors, such as gender, age, tumor thickness, Clark level and anatomic site (P>0.05).
Figure 3.
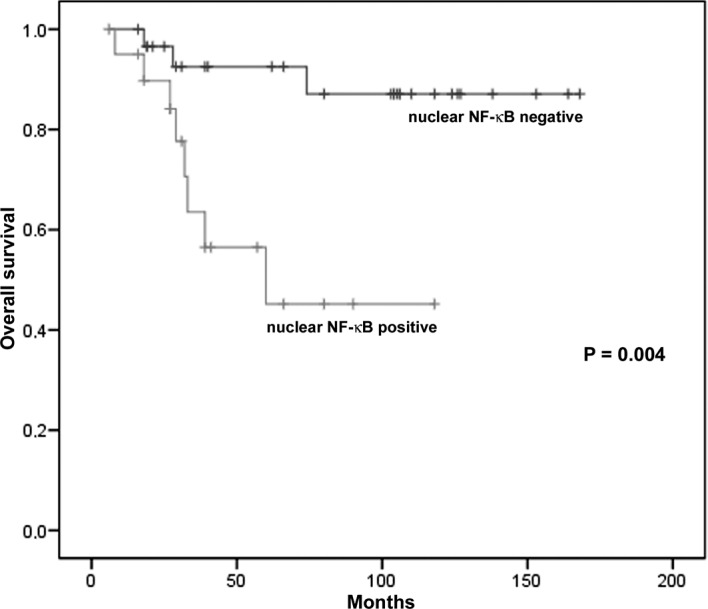
Kaplan-Meier overall survival analysis of stages I and II melanoma patients according to the expression of nuclear NF-κB.
Table III.
Univariate analysis of NF-κB expression for 5-year survival in melanoma patients.
| Variable | No. of patients | No. of events | 5-year survival | SEa | P-valueb |
|---|---|---|---|---|---|
| Nuclear NF-κB | 0.004 | ||||
| Negative | 30 | 3 | 0.92 | 0.05 | |
| Positive | 21 | 8 | 0.45 | 0.14 | |
| Total NF-κB | 0.040 | ||||
| Negative | 23 | 2 | 0.90 | 0.06 | |
| Positive | 28 | 9 | 0.62 | 0.10 |
Standard error;
Log-rank test.
Bold, significant p-values.
Discussion
NF-κB plays an important role in cancer development and progression. Numerous studies have demonstrated that NF-κB is activated in a variety of cancer types, including breast, lung, gastric, esophageal, pancreatic, prostate cancer (33–38) and melanoma (2,8).
Our study aimed to evaluate whether NF-κB expression in tumor tissues has an effect on the survival of melanoma patients. We used immunohistochemistry, a simple reproducible method for pathologists to conduct the investigation or for routine use in the estimation of NF-κB. Intense immunoreactivity was found adjacent to the nucleus, leaning against the nuclear envelope. This immunopositivity may reflect the presence of NF-κB dimers activated in the cytoplasm and travelling to the nucleus. For this reason, immunopositivity was considered to indicate NF-κB activation in the tumor tissue, together with nuclear staining and cytoplasmic overexpression. In particular, it was included in nuclear positivity as an index of NF-κB activation towards its transcriptional activity. The Kaplan-Meier univariate analysis showed that patients with low levels of NF-κB in the nuclei of tumor cells (nuclear NF-κB-negative) had a significantly longer survival compared to those with high levels (nuclear NF-κB-positive). In order to confirm the prognostic role of nuclear NF-κB expression, we conducted a multivariate Cox regression analysis for patient survival, including known prognostic factors such as age, gender, anatomic location, tumor thickness, Clark level and AJCC stage. The predictive value of nuclear NF-κB expression was found to maintain significance after the model was adjusted using the aforementioned clinicopathological factors, thereby confirming the value of nuclear NF-κB as an independent prognostic variable in this patient population. This result is consistent with previously reported immunohistochemical studies showing the association between high nuclear NF-κB p65 expression and progression-free survival of patients with prostate cancer (39,40) and metastatic serous ovarian carcinoma (41). In numerous other studies, NF-κB activation was universally verified to be an adverse prognostic factor (42), and in melanoma it was proposed as an event that promotes tumor progression (12–14). Our findings, which show that the nuclear staining of NF-κB p65 is correlated with disease-specific 5-year survival of melanoma patients, confirm that an increased nuclear expression of NF-κB p65 is a crucial activity during melanoma progression.
Inhibition of apoptosis is the most likely pathway through which NF-κB signaling promotes the development of cancer. Moreover, NF-κB activation is known to suppress apoptosis (7). NF-κB may facilitate invasion and metastasis by inducing the expression of molecules that are mediators of the migration of cancerous cells, crossing of vessel walls and invasion at sites of metastasis (2). NF-κB activity is involved in the regulation of angiogenesis, the process by which tumor cells promote neo-vascularization, an essential step for their growth and invasiveness. Vascular endothelial growth factor, the main member of the angiogenic factor family, is under the transcriptional regulation of NF-κB (43). It has been reported that tumor vascularity is an early requirement for melanoma progression and that NF-κB plays a mediating role between melanoma cells and tumor vasculature (44).
Statistical analysis showed that a positive immunoreaction for NF-κB in the nuclei or cytoplasm of tumor cells was more frequent in cases positive for p53 and survivin and that a complete NF-κB expression was statistically associated with p53 and survivin overexpression. Our results show the concomitant presence of activated forms of NF-κB and p53 overexpression in melanoma cells, an association that may be explained by the functional relationship between the two proteins, since they both play a central role in the control of proliferation and apoptosis. Normally, p53 is able to block the activity of other transcription factors, including NF-κB, in order to promote apoptosis. When p53 is mutated or has lost its activity, a condition detectable through its accumulation and pronounced immunohistochemical expression, its functional loss leads to the activation of NF-κB, observable as an enhanced nuclear localization, as noted in our melanoma samples. Moreover, p53 and NF-κB (p65) mutually inhibit the transcriptional activity, establishing a functional relationship between the two pathways (1). The concomitant expression of activated NF-κB and survivin is in agreement with the role of NF-κB in enhancing the expression of anti-apoptotic proteins, such as survivin, as a mechanism for protection from apoptosis in melanoma (2). Our findings confirm the presence of a constitutive activation of NF-κB in melanoma cells, showing in particular a significantly elevated expression of RelA (p65) and its increased nuclear translocation, suggesting NF-κB activation through the canonical pathway. Notably, the majority of the studies concerning the association between NF-κB and tumor development usually involved the p65 subunit (39–41).
The present study indicates that the interaction of NF-κB with p53 and survivin is a potential key signaling pathway in the process of melanoma pathogenesis. Moreover, it suggests that patients with tumors that are strongly positive for nuclear NF-κB p65 expression should be regarded as being at high risk of mortality, and that nuclear NF-κB p65 may be a promising early independent prognostic factor in patients with primary cutaneous melanoma. The evaluation of nuclear NF-κB expression, either alone or in combination with other routinely available clinical and histological prognostic markers, may be useful in improving the prediction of outcome of melanoma patients. NF-κB inhibition is considered to be promising in the fight against cancer, and many efforts are now concentrated on the ability to identify novel NF-κB targets specifically activated in tumors. For this reason, we believe that targeting NF-κB may be useful in developing a risk-adjusted approach to adjuvant therapies in melanoma patients.
Acknowledgements
This study was supported by grants from the Ministero Istruzione Università Ricerca – MIUR, Ministero Affari Esteri – MAE and Fondazione Banco di Sardegna. Particular thanks are due to Mrs. Itala Mosso and Mr. Massimo Annis for the expert technical assistance.
References
- 1.Pacifico F, Leonardi A. NF-κB in solid tumors. Biochem Pharmacol. 2006;72:1142–1152. doi: 10.1016/j.bcp.2006.07.032. [DOI] [PubMed] [Google Scholar]
- 2.Amiri KI, Richmond A. Role of nuclear factor-κB in melanoma. Cancer Metastasis Rev. 2005;24:301–313. doi: 10.1007/s10555-005-1579-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Meteoglu I, Erdogdu IH, Meydan N, Erkus M, Barutca S. NF-kappaB expression correlates with apoptosis and angiogenesis in clear cell renal cell carcinoma tissues. J Exp Clin Cancer Res. 2008;27:53. doi: 10.1186/1756-9966-27-53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Ueda Y, Richmond A. NF-κB activation in melanoma. Pigment Cell Res. 2006;19:112–124. doi: 10.1111/j.1600-0749.2006.00304.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Poser I, Bosserhoff AK. Transcription factors involved in development and progression of malignant melanoma. Histol Histopathol. 2004;19:173–188. doi: 10.14670/HH-19.173. [DOI] [PubMed] [Google Scholar]
- 6.Ghosh S, Karin M. Missing pieces in the NF-kappaB puzzle. Cell. 2002;109:S81–S96. doi: 10.1016/s0092-8674(02)00703-1. [DOI] [PubMed] [Google Scholar]
- 7.Naugler WE, Karin M. NF-κB and cancer-identifying targets and mechanisms. Curr Opin Genet Dev. 2008;18:19–26. doi: 10.1016/j.gde.2008.01.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Yang J, Pan WH, Clawson GA, Richmond A. Systemic targeting inhibitor of kappaB kinase inhibits melanoma tumor growth. Cancer Res. 2007;67:3127–3134. doi: 10.1158/0008-5472.CAN-06-3547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Dhawan P, Richmond A. A novel NF-kappa B-inducing kinase-MAPK signaling pathway up-regulates NF-kappa B activity in melanoma cells. J Biol Chem. 2002;277:7920–7928. doi: 10.1074/jbc.M112210200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.McNulty SE, Tohidian NB, Meyskens FL. RelA, p50 and inhibitor of kappa B alpha are elevated in human metastatic melanoma cells and respond aberrantly to ultraviolet light B. Pigment Cell Res. 2001;14:456–465. doi: 10.1034/j.1600-0749.2001.140606.x. [DOI] [PubMed] [Google Scholar]
- 11.McNulty SE, Del Rosario R, Cen D, Meyskens FL, Jr, Yang S. Comparative expression of NF-kappaB proteins in melanocytes of normal skin vs. benign intradermal naevus and human metastatic melanoma biopsies. Pigment Cell Res. 2004;17:173–180. doi: 10.1111/j.1600-0749.2004.00128.x. [DOI] [PubMed] [Google Scholar]
- 12.Huang S, DeGuzman A, Bucana CD, Fidler IJ. Level of interleukin-8 expression by metastatic human melanoma cells directly correlates with constitutive NF-kappaB activity. Cytokines Cell Mol Ther. 2000;6:9–17. doi: 10.1080/13684730050515868. [DOI] [PubMed] [Google Scholar]
- 13.Payne AS, Cornelius LA. The role of chemokines in melanoma tumor growth and metastasis. J Invest Dermatol. 2002;118:915–922. doi: 10.1046/j.1523-1747.2002.01725.x. [DOI] [PubMed] [Google Scholar]
- 14.Richmond A. NF-kappa B, chemokine gene transcription and tumour growth. Nat Rev Immunol. 2002;2:664–674. doi: 10.1038/nri887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Salvesen GS, Duckett CS. Apoptosis: IAP proteins: blocking the road to death’s road. Nat Rev Mol Cell Biol. 2002;3:401–410. doi: 10.1038/nrm830. [DOI] [PubMed] [Google Scholar]
- 16.Altieri DC. The case for survivin as a regulator of microtubule dynamics and cell-death decisions. Curr Opin Cell Biol. 2006;18:609–615. doi: 10.1016/j.ceb.2006.08.015. [DOI] [PubMed] [Google Scholar]
- 17.Altieri DC. Validating survivin as a cancer therapeutic target. Nat Rev Cancer. 2003;3:46–54. doi: 10.1038/nrc968. [DOI] [PubMed] [Google Scholar]
- 18.Zaffaroni N, Pennati M, Daidone MG. Survivin as a target for new anticancer interventions. J Cell Mol Med. 2005;9:360–372. doi: 10.1111/j.1582-4934.2005.tb00361.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Piras F, Perra MT, Murtas D, Minerba L, Floris C, Maxia C, Demurtas P, Ugalde J, Ribatti D, Sirigu P. Combinations of apoptosis and cell-cycle control biomarkers predict the outcome of human melanoma. Oncol Rep. 2008;20:271–277. [PubMed] [Google Scholar]
- 20.Mirza A, McGuirk M, Hockenberry TN. Human survivin is negatively regulated by wild-type p53 and participate in p53-dependent apoptotic pathway. Oncogene. 2002;21:2613–2622. doi: 10.1038/sj.onc.1205353. [DOI] [PubMed] [Google Scholar]
- 21.Mees C, Nemunaitis J, Senzer N. Transcription factors: their potential as targets for an individualized therapeutic approach to cancer. Cancer Gene Ther. 2009;16:103–112. doi: 10.1038/cgt.2008.73. [DOI] [PubMed] [Google Scholar]
- 22.Iyer NG, Chin SF, Ozdag H, Daigo Y, Hu DE, Cariati M, Brindle K, Aparicio S, Caldas C. p300 regulates p53-dependent apoptosis after DNA damage in colorectal cancer cells by modulation of PUMA/p21 levels. Proc Natl Acad Sci USA. 2004;101:7386–7391. doi: 10.1073/pnas.0401002101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Sirigu P, Piras F, Minerba L, Murtas D, Maxia C, Colombari R, Corbu A, Perra MT, Ugalde J. Prognostic prediction of the immunohistochemical expression of p16 and p53 in cutaneous melanoma: a comparison of two populations from different geographical regions. Eur J Histochem. 2006;50:191–198. [PubMed] [Google Scholar]
- 24.Ryan KM, O’Prey J, Vousden KH. Loss of nuclear factor-κB is tumor promoting but does not substitute for loss of p53. Cancer Res. 2004;64:4415–4418. doi: 10.1158/0008-5472.CAN-04-1474. [DOI] [PubMed] [Google Scholar]
- 25.Fridman JS, Lowe SW. Control of apoptosis by p53. Oncogene. 2003;22:9030–9040. doi: 10.1038/sj.onc.1207116. [DOI] [PubMed] [Google Scholar]
- 26.Zuckerman V, Wolyniec K, Sionov RV, Haupt S, Haupt Y. Tumour suppression by p53: the importance of apoptosis and cellular senescence. J Pathol. 2009;219:3–15. doi: 10.1002/path.2584. [DOI] [PubMed] [Google Scholar]
- 27.Olivier M, Hussain SP, Caron de Fromentel C, Hainaut P, Harris CC. TP53 mutation spectra and load: a tool for generating hypotheses on the etiology of cancer. IARC Sci Publ. 2004;157:247–270. [PubMed] [Google Scholar]
- 28.Culmsee C, Siewe J, Junker V, Retiounskaia M, Schwarz S, Camandola S, El-Metainy S, Behnke H, Mattson MP, Krieglstein J. Reciprocal inhibition of p53 and nuclear factor-kappaB transcriptional activities determines cell survival or death in neurons. J Neurosci. 2003;23:8586–8595. doi: 10.1523/JNEUROSCI.23-24-08586.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Wang J, Ouyang W, Li J, Wei L, Ma Q, Zhang Z, Tong Q, He J, Huang C. Loss of tumor suppressor p53 decreases PTEN expression and enhances signaling pathways leading to activation of activator protein 1 and nuclear factor-kappaB induced by UV radiation. Cancer Res. 2005;65:6601–6611. doi: 10.1158/0008-5472.CAN-04-4184. [DOI] [PubMed] [Google Scholar]
- 30.Shao J, Fujiwara T, Kadowaki Y, Fukazawa T, Waku T, Itoshima T, Yamatsuji T, Nishizaki M, Roth JA, Tanaka N. Overexpression of the wild-type p53 gene inhibits NF-kappaB activity and synergizes with aspirin to induce apoptosis in human colon cancer cells. Oncogene. 2000;19:726–736. doi: 10.1038/sj.onc.1203383. [DOI] [PubMed] [Google Scholar]
- 31.Clark WH, Jr, Elder DE, Guerry D, IV, Braitman LE, Trock BJ, Schultz D, Synnestvedt M, Halpern AC. Model predicting survival in stage I melanoma based on tumor progression. J Natl Cancer Inst. 1989;81:1893–1904. doi: 10.1093/jnci/81.24.1893. [DOI] [PubMed] [Google Scholar]
- 32.Greene FL, Page DL, Fleming ID, Fritz A, Balch CM, Haller DG, Morrow M, editors. American Joint Committee on Cancer Staging Manual. 6th edition. Springer; Philadelphia: 2002. [Google Scholar]
- 33.Montagut C, Tusquets I, Ferrer B, Corominas JM, Bellosillo B, Campas C, Suarez M, Fabregat X, Campo E, Gascon P, Serrano S, Fernandez PL, Rovira A, Albanell J. Activation of nuclear factor-kappa B is linked to resistance to neoadjuvant chemotherapy in breast cancer patients. Endocr Relat Cancer. 2006;13:607–616. doi: 10.1677/erc.1.01171. [DOI] [PubMed] [Google Scholar]
- 34.Zhang Z, Ma J, Li N, Sun N, Wang C. Expression of nuclear factor-kappaB and its clinical significance in non-small-cell lung cancer. Ann Thorac Surg. 2006;82:243–248. doi: 10.1016/j.athoracsur.2006.01.049. [DOI] [PubMed] [Google Scholar]
- 35.Levidou G, Korkolopoulou P, Nikiteas N, Tzanakis N, Thymara I, Saetta AA, Tsigris C, Rallis G, Vlasis K, Patsouris E. Expression of nuclear factor kappaB in human gastric carcinoma: relationship with I kappaB a and prognostic significance. Virchows Arch. 2007;450:519–527. doi: 10.1007/s00428-007-0396-5. [DOI] [PubMed] [Google Scholar]
- 36.Izzo JG, Malhotra U, Wu TT, Luthra R, Correa AM, Swisher SG, Hofstetter W, Chao KS, Hung MC, Ajani JA. Clinical biology of esophageal adenocarcinoma after surgery is influenced by nuclear factor-kappaB expression. Cancer Epidemiol Biomarkers Prev. 2007;16:1200–1205. doi: 10.1158/1055-9965.EPI-06-1083. [DOI] [PubMed] [Google Scholar]
- 37.Weichert W, Boehm M, Gekeler V, Bahra M, Langrehr J, Neuhaus P, Denkert C, Imre G, Weller C, Hofmann HP, Niesporek S, Jacob J, Dietel M, Scheidereit C, Kristiansen G. High expression of RelA/p65 is associated with activation of nuclear factor-kappaB-dependent signaling in pancreatic cancer and marks a patient population with poor prognosis. Br J Cancer. 2007;97:523–530. doi: 10.1038/sj.bjc.6603878. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Lessard L, Karakiewicz PI, Bellon-Gagnon P, Alam-Fahmy M, Ismail HA, Mes-Masson AM, Saad F. Nuclear localization of nuclear factor-kappaB p65 in primary prostate tumors is highly predictive of pelvic lymph node metastases. Clin Cancer Res. 2006;12:5741–5745. doi: 10.1158/1078-0432.CCR-06-0330. [DOI] [PubMed] [Google Scholar]
- 39.Fradet V, Lessard L, Bégin LR, Karakiewicz P, Masson AM, Saad F. Nuclear factor-kappaB nuclear localization is predictive of biochemical recurrence in patients with positive margin prostate cancer. Clin Cancer Res. 2004;10:8460–8464. doi: 10.1158/1078-0432.CCR-04-0764. [DOI] [PubMed] [Google Scholar]
- 40.Ross JS, Kallakury BV, Sheehan CE, Fisher HA, Kaufman RP, Jr, Kaur P, Gray K, Stringer B. Expression of nuclear factor-kappa B and I kappa B alpha proteins in prostatic adenocarcinomas: correlation of nuclear factor-kappa B immunoreactivity with disease recurrence. Clin Cancer Res. 2004;10:2466–2472. doi: 10.1158/1078-0432.ccr-0543-3. [DOI] [PubMed] [Google Scholar]
- 41.Kleinberg L, Dong HP, Holth A, Risberg B, Trope’ CG, Nesland JM, Flørenes VA, Davidson B. Cleaved caspase-3 and nuclear factor-kappaB p65 are prognostic factors in metastatic serous ovarian carcinoma. Hum Pathol. 2009;40:795–806. doi: 10.1016/j.humpath.2008.10.019. [DOI] [PubMed] [Google Scholar]
- 42.Liu X, Wang B, Ma X, Guo Y. NF-kappaB activation through the alternative pathway correlates with chemoresistance and poor survival in extranodal NK/T-cell lymphoma, nasal type. Jpn J Clin Oncol. 2009;39:418–424. doi: 10.1093/jjco/hyp037. [DOI] [PubMed] [Google Scholar]
- 43.Kiriakidis S, Andreakos E, Monaco C, Foxwell B, Feldmann M, Paleolog E. VEGF expression in human macrophages is NF-kappaB-dependent: studies using adenoviruses expressing the endogenous NF-kappaB inhibitor IkappaBalpha and a kinase-defective form of the IkappaB kinase 2. J Cell Sci. 2003;116:665–674. doi: 10.1242/jcs.00286. [DOI] [PubMed] [Google Scholar]
- 44.Kashani-Sabet M, Shaikh L, Miller JR, III, Nosrati M, Ferreira CM, Debs RJ, Sagebiel RW. NF-kappa B in the vascular progression of melanoma. J Clin Oncol. 2004;22:617–623. doi: 10.1200/JCO.2004.06.047. [DOI] [PubMed] [Google Scholar]