

Abstract
Elderly patients face the problems of morbidity and mortality due to age-mediated disabilities. The purpose of the present study was to investigate the expression of thrombospondin-1 (TSP-1) and transforming growth factor-β (TGF-β) in aging mice, and its probable mechanism in the pathological changes of aging myocardium. The aging model group (AM) comprised 30-month-old mice, while the control group comprised 2-month-old mice. The pathological changes were explored by H&E staining, and the contents of superoxide dismulase (SOD) and malondialdehyde (MDA) in the hearts were determined by xanthine oxidation or TBA colorimetry. TSP-1 and TGF-β expression in the left ventricular myocardium was also measured by immunohistochemistry. The results showed that the activities of SOD decreased and the MDA content increased markedly in the hearts of the AM group compared to the control group. H&E staining showed that the control group myocardial cells lined up in order with clear structure and stained equably, while the AM group myocardial cells lined up in disorder with an augmented cell body and the appearance of many granules and interstitial fibrosis. Compared to the control group, in the hearts of the AM group, TSP-1 and TGF-β protein expression in myocardial cells showed a significant increase (P<0.01). TSP-1 and TGF-β expression increased in the myocardium, which may be related to pathological changes of age-related heart diseases, such as hypertrophy, fibrosis of myocardial cells and microvessel dissepiment thickening.
Keywords: aging, myocardium, thrombospondin-1, transforming growth factor-β
Introduction
With the projected increase in the elderly population and the prevalence of age-related cardiovascular disabilities worldwide, more and more elderly patients face the problems of morbidity and mortality due to age-mediated disabilities. Studies have found that the aging heart has a diminished functional and adaptive reserve capacity, an increased susceptibility to incur damage and a limited practical ability for regeneration (1). Specific age-related alterations in the myocardium cannot be directly ascribed to the loss or reduced number of myocytes, coronary atherosclerosis or vasospasm, scarring or other defined cardiac abnormalities. The development of interstitial fibrosis may also explain the altered cardiac function of the aged myocardium (2), and aging of human atrial myocardium is also accompanied by a decrease in nerve plexuses and an increase in fibrosis (3). However, the exact underlying mechanism is not yet fully understood.
Different cardiac pathology appears to be caused by a common fibrotic process. Recent studies indicate that dilated, ischaemic and hypertrophic cardiomyopathies are all associated with increased levels of transforming growth factor-β (TGF-β) 1, and TGF-β1 has now been suggested to play a major role in valvular disease and arrhythmia, particularly atrial fibrillation (4). Age-dependent changes, such as increased activation of the renin-angiotensin-aldosterone system (RAAS) (5) and an increase in oxidative stress (6), can be potent fibrogenic activators of TGF-β. After its extracellular activation, TGF-β binds to the specific receptors TGF-β-R1 and -R2 mediating phosphorylation of transcription signaling molecules (p-smads) and subsequent transcription of target genes (7).
Thrombospondin-1 (TSP-1) is the founding member of a family of matricellular glycoproteins (8). While predominantly in vitro studies have assigned diverse functions, such as phagocytosis, adhesion, chemotaxis, proliferation and migration of cells to TSP-1, it has also been identified as an activator of the TGF-β-procytokine complex. It is thought that binding of TSP-1 to the latent complex induces a conformational rearrangement of the latency associate peptide (LAP) in such a manner as to prevent the LAP from conferring latency on the mature domain of TGF-β (9). Several studies have previously determined that TSP-1 expression was greatly enhanced and exacerbated the development of glomerulosclerosis and tubulointerstitial fibrosis in the aging kidney (10). However, further studies concerning TSP-1 and TGF-β expression in the heart of aging mouse models are necessary.
In the present study, we employed 30-month-old mice to represent the aging mouse model. The aim of the present study was to confirm changes in TSP-1 and TGF-β expression in the aging model and to explore the pathogenesis of interstitial fibrosis in the aging heart, in order to provide evidence for a new clinical treatment for the vulnerable senescent myocardium.
Materials and methods
Animal and sample preparation
Twenty 1-month-old male Kunming mice with an initial body weight of 20–22 g were purchased from the Experiment Animal Center of Zhejiang University and divided into two groups: 10 mice were fed for 30 months as the aging mouse model group (AM) and another 10 were fed for 2 months as the control group. All mice were housed under a 12-h light/dark cycle and provided with standard diet and water. All procedures were conducted with the approval of the Zhejiang University Ethics and Animal Care Committee (under NIH policies) and according to institutional guidelines for the experimental use of animals.
Both groups of mice were sacrificed by dislocation of the cervical vertebra. Their thoracic cavities were then opened. The heart of each mouse was extracted, immediately fixed in precooling saline and blood was removed and rinsed clean with saline. Five mice per group were prepared for tissue homogenization. Quantum satis heart tissue was accurately weighed, cut into small pieces, placed into a glass tissue homogenizer and precooling saline was added for preparation of the tissue homogenate, which was centrifugalized. The supernatant was then used for determination of superoxide dismulase (SOD) and malondialdehyde (MDA). The remaining 5 mice per group were prepared for paraffin sections. The hearts were fixed in 4% paraformaldehyde for 4 h, then placed in 30% phosphate-buffered sucrose until the tissue sank. After embedding in paraffin, the hearts were sectioned on a rotary microtome and 8-μm-thick sections were cut on a freezing microtome through coronary planes of the heart.
Measurement of SOD activity and MDA content in the heart
The contents of SOD and MDA in the hearts were determined by xanthine oxidation or TBA colorimetry. They were measured according to the methods described in the instructions in the SOD and MDA kits using chemicals purchased from the Nanjing Jiancheng Bioengineering Institute.
Histopathological examination and immunohistochemical staining
Paraffin section samples of the heart from each mouse were stained with H&E, and then histopathological changes in the hearts were examined under an optical microscope. Immunohistochemistry was used to localize the TSP-1 and TGF-β. The ABC system was used with 3,3’ diaminobenzidine hydrochloride (DAB) as the chromagen. Briefly, heart sections were deparaffinized, washed in 0.01 M phosphate-buffered saline (PBS; pH 7.4) three times, each for 3 min, and treated with 3% H2O2 for 10 min at room temperature to block endogenous peroxidases.
The sections were subsequently washed with PBS three times, each for 3 min, and incubated overnight at 4°C in the primary antibody (monoclonal TSP-1 antibody or monoclonal TGF-β antibody; Santa Cruz Biotechnology); the dilution of the primary antibody was 1:100. The next day, the sections were incubated in a biotinylated secondary antibody (diluted to 1:200 in PBS) and subsequently in an avidin HRP solution. Immunolabeling was visualized with 0.05% DAB plus 0.3% H2O2 in PBS, and positive staining was shown as a yellow color. The sections were then counterstained with diluted hematoxylin, dehydrated through ethanol and xylene before being placed on coverslips with Permount. As a negative control, PBS was used instead of the primary antibody.
Image analysis and statistics
The heart sections were examined at x400 with UTHSCSA Image Tools 3.0 (University of Texas Medical School at San Antonio, TX, USA). The number of TSP-1- and TGF-β-positive cells was determined. Statistical analysis was performed using the SPSS statistical software program. Values were expressed as the mean ± SD. The significance of the difference was calculated by the two-tailed Student’s t test. P-values <0.05 are considered to denote statistical significance differences.
Results
General appearance
The two groups of mice showed no significant difference in weight and behaviour, and exhibited normal reactions and smooth hairs before modeling. However, after feeding for 6 weeks, compared to the control group, the AM group showed symptoms of senescence, such as decreased weight, yellow, loose and dim hairs, obtuse behaviour and reaction, and polyuria. The results corroborate previous studies (6–8).
SOD activity and MDA content in the heart
The SOD activity decreased and the MDA content increased markedly in the hearts of the AM group mice compared to the control group (P<0.05, Fig. 1).
Figure 1.
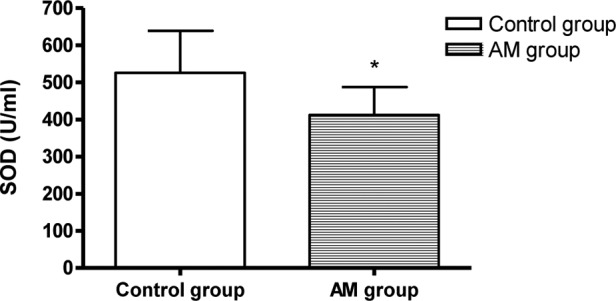
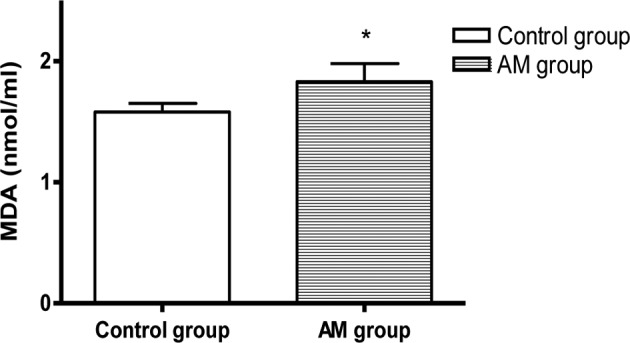
Comparison of SOD and MDA levels in hearts of the AM and control groups. *P<0.05 vs. control.
Histopathological changes in the hearts
H&E staining showed that myocardial cells of the control group lined up in order with clear structure and stained equably, while myocardial cells of the AM group lined up in disorder with an augmented cell body, uneven pigmentation, and the appearance of many granules and interstitial fibrosis (Fig. 2).
Figure 2.

H&E staining in the myocardium of the two groups. (A) Control group; (B) AM group.
TSP-1 and TGF-β expression and localization
Compared to the control group, in the hearts of the AM group, the numbers of TSP-1 or TGF-β-positive-immunostained myocardial cells were significantly increased (P<0.05, Figs. 3–5).
Figure 3.

TSP-1 immunohistochemical staining in the myocardium of the two groups. (A) Control group; (B) AM group.
Figure 5.

Comparison of the quantitative analysis of the number of TGF-β- and TSP-1-positive-immunostained myocardial cells in the AM and the control group. *P<0.01 vs. the control group.
Discussion
The aging heart is associated with suppressed inflammation, delayed phagocytosis of dead cardiomyocytes, and markedly diminished collagen deposition following myocardial infarction, due to a blunted response of fibroblasts to fibrogenic growth factors (2). The present study revealed that the SOD activity decreased and the MDA content increased markedly in the hearts of the AM group, demonstrating that the classical features of an aging mouse model were induced in the 30-month-old mice.
TGF-β is a group of potent regulatory cytokines involved in many biological processes. Cell culture studies have indicated that TGF-β inhibits mitotic growth of cardiomyocytes, and stimulates hypertrophic growth, fibrosis and re-expression of the fetal isoforms of myofibrillar protein genes (11). Experimental studies have demonstrated that age-related cardiac fibrosis and atherosclerosis are strongly associated with myocardial extracellular matrix (ECM) (12), and TGF-β is known to play a significant role in fibrotic cardiac remodeling and multiple factors altered in the aging heart. Fibrogenic pathways of TGF-β are involved in the regulation of the balance between matrix-degrading matrix metalloproteinases (MMPs) and their inhibitors (TIMPs) (13), mediating a sequential increase in connective tissue growth factor synthesis (14), which is a downstream target of the TGF-β/Smad pathway (7). These regulators may induce age-dependent changes in this proteolytic system and unusually increased deposition of ECM. In the present study, we found that TGF-β expression in myocardial cells of aging mice was significantly increased; it stimulated the proliferation of interstitial fibroblasts, aggravated fibrosis and produced decreased compliance in the aging heart.
TSP-1 is a multifunctional ECM glycoprotein and acts as a potent inhibitor of angiogenesis in vitro and in vivo. Studies have demonstrated that overexpression of TSP-1 in the aging kidney is correlated with glomerulosclerosis and tubulointerstitial fibrosis. TGF-β is secreted as a latent complex of the LAP and the mature domain, which must be activated for TGF-β to signal. Previous studies have identified TSP-1 as a physiological activator of TGF-β in vitro and in vivo. TSP-1 utilizes a two-step mechanism to activate latent TGF-β (15,16). Recent research has implied that there is a mutual activation between TGF-β and TSP-1. Studies of other pathological models proved that TGF-β upregulates TSP-1 synthesis (16), and TGF-β is involved in the fibrogenic process in hepatic stem cells through upregulation of fibronectin expression (17,18). The results of our present study showed that the aging heart model (30-month-old mice) led to the upregulation of TSP-1 and TGF-β protein expression in the myocardium, which suggest that TSP-1 and TGF-β play a role in the development of age-related cardiac fibrosis.
Our findings revealed quantitative changes in TSP-1 and TGF-β in the myocardium in 30-month-old mice. This indicates that the TSP-1–TGF-β axis may positively participate in the process of age-related myocardial fibrosis. Further study is required to identify the exact underlying mechanism.
Figure 4.

TGF-β immunohistochemical staining in the myocardium of the two groups. (A) Control group; (B) AM group.
Acknowledgments
This study was supported by Projects of the Zhejiang Education Bureau.
References
- 1.Jahangir A, Sagar S, Terzic A. Aging and cardioprotection. J Appl Physiol. 2007;103:2120–2128. doi: 10.1152/japplphysiol.00647.2007. [DOI] [PubMed] [Google Scholar]
- 2.Chen W, Frangogiannis NG. The role of inflammatory and fibrogenic pathways in heart failure associated with aging. Heart Fail Rev. 2010;15:415–422. doi: 10.1007/s10741-010-9161-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Burkauskiene A, Mackiewicz Z, Virtanen I, et al. Age-related changes in myocardial nerve and collagen networks of the auricle of the right atrium. Acta Cardiol. 2006;61:513–518. doi: 10.2143/AC.61.5.2017765. [DOI] [PubMed] [Google Scholar]
- 4.Khan R, Sheppard R. Fibrosis in heart disease: understanding the role of transforming growth factor-beta in cardiomyopathy, valvular disease and arrhythmia. Immunology. 2006;118:10–24. doi: 10.1111/j.1365-2567.2006.02336.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Ishihata A, Katano Y. Role of angiotensin II and endothelin-1 receptors in aging-related functional changes in rat cardiovascular system. Ann NY Acad Sci. 2006;1067:173–181. doi: 10.1196/annals.1354.021. [DOI] [PubMed] [Google Scholar]
- 6.Wang M, Zhang J, Walker S, et al. Involvement of NADPH oxidase in age-associated cardiac remodeling. J Mol Cell Cardiol. 2010;48:765–772. doi: 10.1016/j.yjmcc.2010.01.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Zhu HJ, Burgess AW. Regulation of transforming growth factor-beta signaling. Mol Cell Biol Res Commun. 2001;4:321–330. doi: 10.1006/mcbr.2001.0301. [DOI] [PubMed] [Google Scholar]
- 8.Iruela-Arispe ML. Regulation of thrombospondin1 by extracellular proteases. Curr Drug Targets. 2008;9:863–868. doi: 10.2174/138945008785909365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Schultz-Cherry S, Ribeiro S, Gentry L, et al. Thrombospondin binds and activates the small and large forms of latent transforming growth factor-beta in a chemically defined system. J Biol Chem. 1994;269:26775–26782. [PubMed] [Google Scholar]
- 10.Kang DH, Anderson S, Kim YG, et al. Impaired angiogenesis in the aging kidney: vascular endothelial growth factor and thrombospondin-1 in renal disease. Am J Kidney Dis. 2001;37:601–611. doi: 10.1053/ajkd.2001.22087. [DOI] [PubMed] [Google Scholar]
- 11.Bonnema DD, Webb CS, Pennington WR, et al. Effects of age on plasma matrix metalloproteinases (MMPs) and tissue inhibitor of metalloproteinases (TIMPs) J Card Fail. 2007;13:530–540. doi: 10.1016/j.cardfail.2007.04.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Edwards DR, Leco KJ, Beaudry PP, et al. Differential effects of transforming growth factor-beta 1 on the expression of matrix metalloproteinases and tissue inhibitors of metalloproteinases in young and old human fibroblasts. Exp Gerontol. 1996;131:207–223. doi: 10.1016/0531-5565(95)02010-1. [DOI] [PubMed] [Google Scholar]
- 13.Beggs ML, Nagarajan R, Taylor-Jones JM, et al. Alterations in the TGF beta signaling pathway in myogenic progenitors with age. Aging Cell. 2004;3:353–361. doi: 10.1111/j.1474-9728.2004.00135.x. [DOI] [PubMed] [Google Scholar]
- 14.Young GD, Murphy-Ullrich JE. The tryptophan-rich motifs of the thrombospondin type 1 repeats bind VLAL motifs in the latent transforming growth factor-beta complex. J Biol Chem. 2004;279:47633–47642. doi: 10.1074/jbc.M404918200. [DOI] [PubMed] [Google Scholar]
- 15.Zhang XM, Shen F, Xv ZY, et al. Expression changes of thrombospondin-1 and neuropeptide Y in myocardium of STZ-induced rats. Int J Cardiol. 2005;105:192–197. doi: 10.1016/j.ijcard.2004.12.065. [DOI] [PubMed] [Google Scholar]
- 16.Zhang XM, Shi PH, Cao SH, et al. Expression changes of transforming growth factor-beta1 and thrombospondin-1 in cavernous tissues of diabetic rats. Urol Int. 2010;84:221–225. doi: 10.1159/000277602. [DOI] [PubMed] [Google Scholar]
- 17.Cui W, Jin HB, Li ZW. Mechanism of the transforming growth factor-beta induction of fibronectin expression in hepatic stem-like cells. Braz J Med Biol Res. 2010;43:36–42. doi: 10.1590/s0100-879x2009007500017. [DOI] [PubMed] [Google Scholar]
- 18.Ren M, Wang B, Zhang J, et al. Smad2 and Smad3 as mediators of the response of adventitialfibroblasts induced by transforming growth factor β1. Mol Med Rep. 2011;4:561–567. doi: 10.3892/mmr.2011.458. [DOI] [PubMed] [Google Scholar]