

Abstract
We previously demonstrated that vitamin B6 suppresses tumorigenesis in the colon of mice and exerts an anti-inflammatory effect through the inhibition of NF-κB activation. As these effects resemble the pharmacological properties of thiazolidinedione (TZD), a synthetic peroxisome proliferator-activated receptor-γ (PPARγ) ligand, this study was designed to examine the effect of vitamin B6 on the activation of PPARγ and adipogenesis in 3T3-L1 adipocyte cells. Pyridoxal 5′-phosphate (PLP), one of the vitamin B6 derivatives, was shown to promote adipogenesis in the 3T3-L1 adipocytes. In addition, PLP specifically induced mRNA expression of PPARγ target genes in the 3T3-L1 adipocytes and enhanced the lipid accumulation and adipocyte fatty acid-binding protein (aP2) mRNA expression in NIH3T3 cells stably expressing PPARγ. Furthermore, the administration of vitamin B6 increased the expression of aP2 mRNA in mouse adipose tissues. Collectively, these observations suggest a novel function of vitamin B6 as an activator for PPARγ, which may contribute to the anti-tumor and anti-inflammatory effects of vitamin B6.
Keywords: vitamin B6, 3T3-L1, peroxisome proliferator-activated receptor-γ, adipocytes
Introduction
Vitamin B6 is well known as a water-soluble vitamin essential for normal growth, development and metabolism. Natural vitamin B6 consists of six interconvertible compounds: pyridoxine, pyridoxal, pyridoxamine and their phosphorylated forms, pyridoxine 5′-phosphate, pyridoxal 5′-phosphate (PLP) and pyridoxamine 5′-phosphate. In particular, PLP is a biologically active form of vitamin B6 and functions as a cofactor for numerous enzymes involved in amino acid and cellular metabolisms (1). Vitamin B6 has been classically classified as a coenzyme; however, recent reports have prompted us to consider other physiological functions for vitamin B6. Vitamin B6 has been reported to play an important protective role against several types of diseases (2–4). We previously demonstrated a preventive effect of dietary vitamin B6 in moderate dosage against tumorigenesis in the colon of mice which were administered azoxymethane (AOM) (5). Further studies have demonstrated that the anti-tumorigenesis effect of vitamin B6 in the colon of AOM-injected mice may be mediated, in part, via the suppression of cell hyperproliferation, inducible nitric oxide synthase (iNOS) expression and oxidative stress (5,6). Matsubara et al found an anti-angiogenic effect of vitamin B6 in an ex vivo serum-free matrix culture using rat aortic ring models (7). Furthermore, our recent study revealed an anti-inflammatory effect of vitamin B6, showing that treatment with vitamin B6 inhibits lipopolysaccharide (LPS)-induced iNOS and cyclooxygenase-2 (COX2) expression in RAW 264.7 cells through suppression of NF-κB activation, the pro-inflammatory transcription factor (8). In addition to this in vitro experiment, dietary vitamin B6 inhibited nitric oxide (NO) production in response to LPS administration in vivo (8). Since angiogenesis and inflammation have been considered to play critical roles in the pathogenesis of colon cancer, the anti-angiogenic and anti-inflammatory effects of vitamin B6 may elucidate the mechanisms underlying the anti-tumor properties of vitamin B6.
Peroxisome proliferator-activated receptor-γ (PPARγ) is one such protein that may explain, in part, the anti-tumor behavior of vitamin B6. PPARγ, one of three PPAR subtypes (PPARα, PPARγ and PPARδ/β), is a ligand-dependent transcription factor that belongs to the nuclear hormone receptor superfamily (9). PPARγ is found highly expressed in adipose tissue, and to a lesser extent in the colon and immune system (10). PPARγ enhances adipocyte-specific gene expression via the formation of a heterodimeric DNA-binding complex with the retinoid X receptor. To date, overexpression and knockout studies with mice strongly suggest that PPARγ is an essential transcription factor in adipogenesis in vitro and in vivo (11,12). Furthermore, the most extensively employed insulin-sensitizing drugs, thiazolidinedione derivatives (TZDs), such as troglitazone, pioglitazone and rosiglitazone, have been found to possess a high affinity for PPARγ (13), suggesting that the pharmacological actions of TZDs are mediated through PPARγ activation in adipocytes. It has been reported that improvement in insulin resistance by PPARγ activation is due to an increase in the number of differentiating adipocytes, which promotes adipose-secreted hormone expression and, eventually, glucose homeostasis (9). Furthermore, several substances including the prostaglandin J2 derivative are potent natural ligands of PPARγ (14) which control a variety of physiological functions through the PPARγ signaling pathway.
Activation of PPARγ via TZDs has been found to elicit both anti-neoplastic and anti-inflammatory effects. Therefore, PPARγ is expected to be a pharmacological target for several types of cancers and cardiovascular diseases (15,16). In colon carcinogenesis, troglitazone, one of the TZDs, has been shown to inhibit tumor growth (17) and to reduce the number of aberrant crypt foci, which are precursor lesions of colon cancer in the colon of mice administered AOM (18,19). In addition, recent reports have demonstrated that the anti-atherosclerotic effect of TZDs is partially explained by the anti-inflammatory functions of PPARγ, and that TZDs inhibit the expression of various inflammatory proteins including iNOS and COX2 in macrophages. To date, several mechanisms underlying the anti-inflammatory effects of PPARγ ligands have been proposed, including the inhibition of NF-κB activity (20).
Kawada et al previously tested the effects of vitamins and their analogues on the terminal differentiation of 3T3-L1 cells and found that vitamin B6 derivatives regulated adipogenesis of 3T3-L1 adipocyte cells (21). As the pharmacological properties of TZDs appear to be similar to the biological effects of vitamin B6, we hypothesized that vitamin B6 regulates PPARγ activation similar to TZDs. To examine this hypothesis, we investigated whether vitamin B6 affects adipogenesis in 3T3-L1 adipocytes, which are able to be efficiently differentiated in the presence of TZDs and support PPARγ target gene mRNA expression. The present study shows that treatment with PLP, one of the vitamin B6 derivatives, accelerates adipogenesis in 3T3-L1 adipocytes and that PLP up-regulates the expression of PPARγ target genes in NIH3T3 cells transfected with PPARγ. Furthermore, an acute administration of vitamin B6 was shown to enhance the expression of a PPARγ target gene in vivo. These observations suggest that vitamin B6 plays an important role as an activator of PPARγ in vitro and in vivo.
Materials and methods
Chemical reagents
Pyridoxal hydrochloride, PLP and pyridoxine hydrochloride were obtained from Nacalai Tesque (Osaka, Japan). T-174 [5-[2-(naphthalenylmethyl)-5-benzoxazolyl]-methyl]-2,4-thiazolidinedione), a specific ligand for PPARγ, was kindly provided by Tanabe Mitsubishi Pharma Co. (Osaka, Japan).
Cell culture
Mouse 3T3-L1 preadipocytes, NIH3T3 cells and Phoenix 293 cells were cultured in a maintenance medium (10% fetal calf serum, 100 U/ml penicillin and 100 μg/ml streptomycin in Dulbecco’s modified Eagle’s medium at 37°C in 5% CO2/95% air under a humidified condition. High titer retroviruses harboring PPARγ were produced in Phoenix 293 cells and used to infect NIH3T3 cells, as reported previously (22).
Adipocyte differentiation and Oil-Red-O staining
For differentiation assays, confluent 3T3-L1 cells were treated with differentiation medium [maintenance medium plus 0.5 mM 3-isobutyl-1-methylxanthine (IBMX)], 5 μg/ml insulin, and 1 μM dexamethasone (DEX) and incubated for 2 days. Differentiation medium was then replaced with adipocyte growth medium (maintenance medium supplemented with 5 μg/ml insulin) with or without PLP, which was refreshed every 2 days. Differentiated 3T3-L1 cells were fixed with 4% buffered paraformaldehyde for 15 min. A stock solution of 0.3% Oil-Red-O (Sigma) in isopropanol (w/v) was diluted 6:4 prepared in water, filtered and added to the fixed cells. Cells were washed twice in phosphate-buffered saline (PBS) and photographed.
Glycerol-3-phosphate dehydrogenase activity
Differentiated 3T3-L1 cells were washed twice with ice-cold PBS and suspended in 25 mM Tris-HCl, pH 7.5, 1 mM EDTA, and 1 mM phenylmethylsulfonyl fluoride, 1 mg/ml pepstatin, and 1 mg/ml leupeptin. Membranes were disrupted by sonication, and supernatants were collected following centrifugation at 13,000 × g for 10 min. The reaction was initiated by the addition of supernatants to a standard mixture containing 100 mM triethanolamine/HCl buffer (pH 7.5), 2.5 mM EDTA, 0.12 mM β-nicotinamide adenine dinucleotide disodium salt (NADH), 0.2 mM dihydroxyacetone-phosphate and 0.1 mM β-mercaptoethanol. The change in absorbance at 340 nm was measured using a spectrophotometer at 25°C. One unit of enzyme activity was determined to correspond to the oxidation of 1 nmol of NADH/min.
Experimental animals
Male ICR mice (5-weeks old) (Charles River Japan Inc., Japan) were housed in groups of 6 animals in plastic cages in a room with controlled temperature (24°C) and a 12-h light/dark cycle. The animals were provided with free access to the diet and water and were maintained according to the ‘Guide for the Care and Use of Laboratory Animals’ established by Hiroshima University. The experimental diet (vitamin B6-free) consisted of the following components (in g/kg diet): α-corn starch, 402; casein, 200; sucrose, 200; corn oil, 100; cellulose, 50; AIN-93G salt mixture, 35; AIN-93 vitamin mixture (pyridoxine-free), 10; L-cystine, 3. The experimental feeding period was 2 weeks, and the mice were divided into 3 groups of 8 mice each. The animals received an oral administration of pyridoxine hydrochloride (200 or 500 mg/kg body weight) or physiological saline. Six hours after oral administration, epididymal fat pads were removed.
Northern blot hybridization
Total RNA from differentiated 3T3-L1 cells was isolated using Isogen (Nippon Gene). Total RNA from mouse adipose tissues was isolated using RNeasy lipid tissue kit (Qiagen). Total RNA (10 μg) was subjected to Northern blot hybridization. Briefly, RNA was fractionated in 1% agarose gel containing 0.66 M formaldehyde and 0.02 M MOPS (pH 7.0). Fractionated RNAs were transferred onto a Hybond-N+ nylon filter (GE Healthcare) by capillary blotting and then cross-linked by ultraviolet irradiation. cDNA fragments of mouse adipocyte fatty acid-binding protein (aP2) and mouse glycerol kinase (GyK) were amplified using specific primers. Primers were as follows: aP2 forward primer, 5′-GAAGACAGCTCCTCCTCGAAGGTT-3′; aP2 reverse primer, 5′-GGAAGTCACGCCTTTCATAACA-3′; Gyk forward primer, 5′-TGGTGTCAGCAACCAGAGGGA-3′; Gyk reverse primer, 5′-GGCCATAGATCTCAGAAGAAC-3′. 32P-labeled cDNA fragments encoding aP2, GyK, human β-actin and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) were used for Northern blot hybridization as probes. Hybridization was performed in PerfectHyb (Toyobo) at 65°C for 20 h. The membrane was finally washed with 0.2X SSC and 0.1% SDS at 65°C for 30 min, and the hybridization signals were analyzed using the BAS system (Fuji Film).
Statistical analyses
Values are presented as the means ± SE. Statistical significance among the means was estimated at P<0.05 according to the Student’s t-test.
Results
The effect of vitamin B6 on adipogenesis in 3T3-L1 adipocytes
To investigate the effect of vitamin B6 on adipogenesis, 3T3-L1 preadipocytes, which are well characterized as an in vitro model of adipocyte differentiation were used. 3T3-L1 cells differentiated into mature adipocytes upon exposure to a hormonal stimulus (0.5 mM IBMX, 5 μg/ml insulin and 1 μM DEX). After a 2-day incubation with the hormonal mixture, 3T3-L1 cells were cultured in a medium containing vitamin B6 for 12 days and then subjected to Oil-Red-O staining for lipid droplet visualization. As shown in Fig. 1A, 3T3-L1 cells treated with PLP accumulated larger and a greater number of lipid droplets than the control cells. Since glycerol-3-phosphate dehydrogenase (GPDH) occupies a central position in the pathway of triglyceride synthesis, GPDH enzyme activity was measured as an adipocyte differentiation marker. In agreement with the Oil-Red-O staining, treatment with 100 μM PLP resulted in a significant increase (by 60%) in GPDH enzyme activity compared to untreated cells (Fig. 1B).
Figure 1.
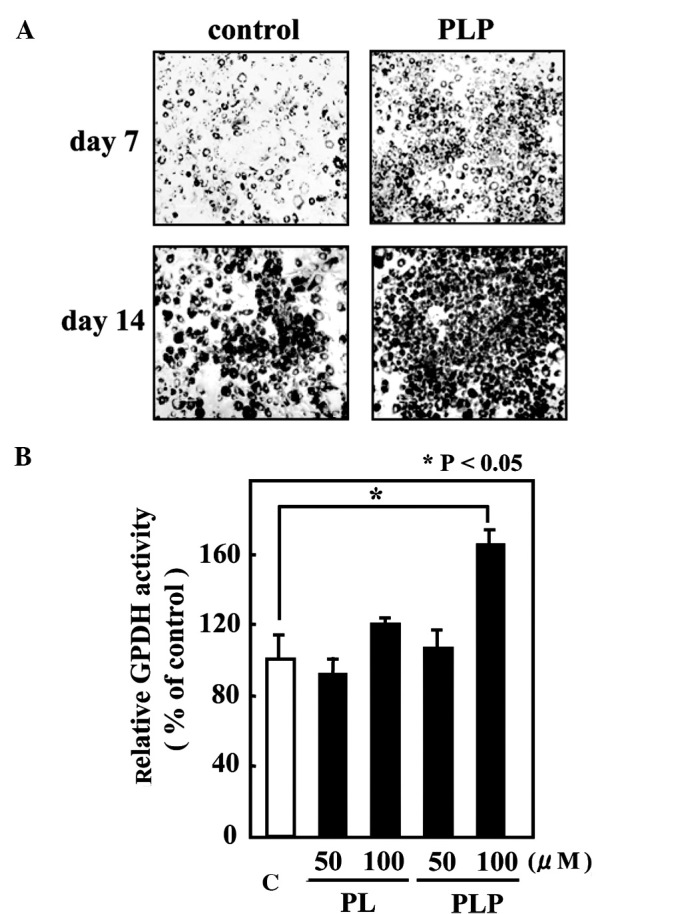
The effect of vitamin B6 on adipogenesis in 3T3-L1 adipocytes. (A) 3T3-L1 adipocytes were treated with 100 μM PLP for 12 days (from day 2–14). Intracellular lipid droplets were visualized by Oil-Red-O staining. (B) GPDH activity was determined after a 12-day cultivation with vitamin B6 at the indicated concentrations. The GPDH activity of untreated cells was set at 100%, and the relative activities were presented as fold induction compared to that of the untreated cells (C). Values are the mean ± SE (n=5). *P<0.05 vs. control.
PLP induces the expression of PPARγ target genes in 3T3-L1 adipocytes
As described above, PLP promoted adipogenesis in 3T3-L1 cells. Subsequently, mRNA expression of PPARγ target genes was investigated in the differentiated 3T3-L1 adipocytes. On day 6 after differentiation was induced with the hormonal stimulus, 3T3-L1 cells were treated with either a vehicle control (DMSO), 10 µM T-174 or 100 µM PLP for 24 or 48 h. After 24 h, the addition of 10 µM T-174 resulted in an increase in the mRNA expression levels of PPARγ target genes, aP2 and Gyk (Fig. 2). Although the addition of 100 μM PLP for 24 h had no influence on the mRNA expression levels of PPARγ target genes, PLP treatment for 48 h demonstrated 1.6- and 4.3-fold increases in the mRNA expression levels of aP2 and Gyk genes, respectively (Fig. 2).
Figure 2.
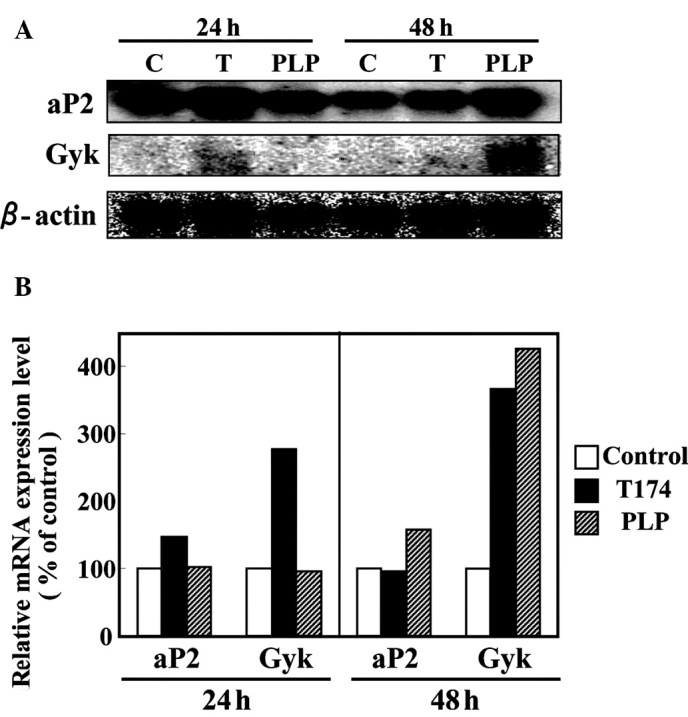
PLP induces the mRNA expression of PPARγ target genes in 3T3-L1 adipocytes. On day 6 after induction of differentiation, 3T3-L1 adipocytes were treated with 10 µM T-174 (T) or 100 µM PLP for 24 or 48 h. (A) Total RNA (10 µg) from 3T3-L1 cells was prepared and subjected to Northern blot analyses. (B) The radioactivity in each band was quantified using an image analyzer and normalized to that of β-actin mRNA.
PLP induces the expression of a PPARγ target gene in NIH3T3 cells transfected with PPARγ
PLP was shown to up-regulate mRNA expression in PPARγ target genes in differentiated 3T3-L1 adipocytes. To examine whether PLP activates PPARγ-dependent transcription in cultured cells, retroviruses harboring PPARγ were used to infect NIH3T3 cells. In PPARγ-transfected NIH3T3 cells, PLP enhanced aP2 mRNA expression and lipid accumulation (Fig. 3). Taken together, these results showed that PLP activates PPARγ-dependent transcription in cultured cells.
Figure 3.

PLP enhances lipid accumulation and aP2 mRNA expression in NIH3T3 cells stably trasnfected with PPARγ. Retroviruses harboring PPARγ were used to infect NIH3T3 cells. (A) NIH3T3 cells stably expressing PPARγ (NIH3T3/PPARγ) were treated with 250 µM PLP for 96 h. Intracellular lipid droplets were visualized by Oil-Red-O staining. (B) NIH3T3/PPARγ cells were treated with 100 µM or 250 µM PLP for 48 or 96 h. Total RNA (10 µg) from NIH3T3 cells was prepared and subjected to Northern blot analyses.
The effect of acute administration of vitamin B6 on expression of a PPARγ target gene in mouse adipose tissues
The present study demonstrated that PLP promotes adipogenesis in 3T3-L1 cells and the mRNA expression of PPARγ target genes in differentiated 3T3-L1 adipocytes. Therefore, we aimed to ascertain whether vitamin B6 regulates the expression of a PPARγ target gene in vivo and to examine the effect of vitamin B6 on the transactivation activity of PPARγ in vivo. ICR mice were fed a vitamin B6-free diet for 2 weeks, and the animals subsequently received an oral administration of pyridoxine hydrochloride (either 200 or 500 mg/kg body weight). Six hours after oral administration, total RNA was isolated from the epididymal fat pads. The administration of vitamin B6 significantly increased aP2 mRNA level in the adipose tissues in a dose-dependent manner (Fig. 4).
Figure 4.
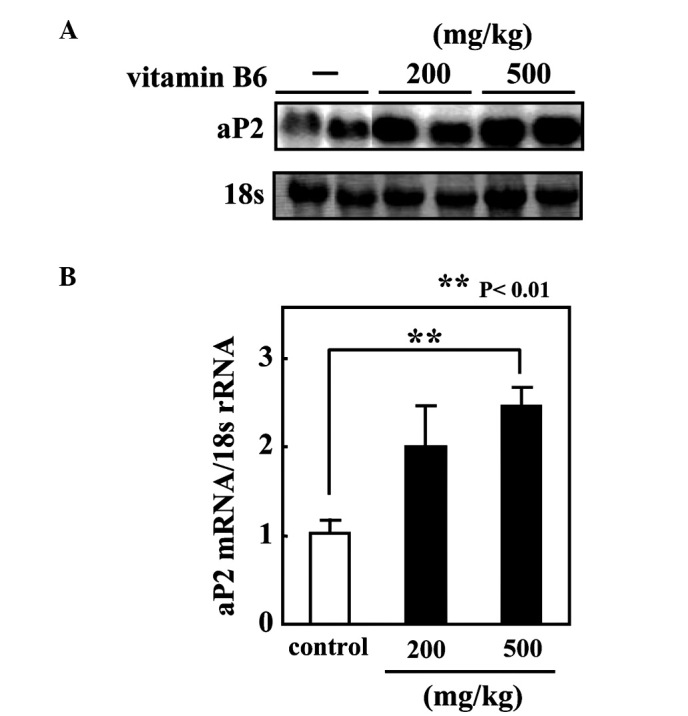
The effect of the acute administration of vitamin B6 on aP2 mRNA expression in mouse adipose tissues. (A) Mice were fed a vitamin B6-free diet for 2 weeks and then received an oral administration of pyridoxine hydrochloride (200 or 500 mg/kg body weight) or physiological saline as control. After a 6-h administration of pyridoxine hydrochloride, 10 μg of total RNA from epididymal fat pads was isolated and subjected to Northern blot analysis. (B) The radioactivity in each band was quantified using an image analyzer and normalized to that of 18S. Each value relative to the mRNA level of the control mice represents the mean ± SE. (n=4). **P<0.01 vs. control.
Discussion
In the present study, PLP, one of the vitamin B6 derivatives, was found to promote adipogenesis in 3T3-L1 cells and up-regulate PPARγ-dependent gene expression in vitro and in vivo. To address whether the effect of PLP on adipogenesis is dependent on the transactivation of PPARγ, NIH3T3 cells stably infected with retroviruses encoding PPARγ were constructed. NIH3T3 cells, which are not committed to adipocytes, have been well studied and show a similar phenotype to 3T3-L1 cells when PPARγ is expressed. In fact, adipocyte differentiation of PPARγ-infected cells was induced in response to T-174 treatment (data not shown), indicating that NIH3T3 cells stably expressing PPARγ are well suited for monitoring adipogenesis simply for PPARγ activation capabilities. In this stable cell line, PLP was shown to promote lipid accumulation, strongly suggesting that PLP stimulates PPARγ transactivation.
Two molecular mechanisms by which PLP affects the expression of PPARγ target genes in a direct or indirect manner were considered for study. First, as it was previously reported that thiazolidine compounds are produced by condensation of aminothiols, such as cysteine, with PLP under a physiological condition (23), PLP may act as a PPARγ ligand by conversion to thiazolidine compounds. Indeed, PLP treatment alone was shown to require more time to induce mRNA expression of PPARγ target genes compared to T-174 treatment. In general, PPARγ ligands are well characterized as lipophilic substances, which contain long-chain polyunsaturated fatty acids and fatty acid metabolites, such as 15-deoxy-Δ12,14-prostaglandin J2. PPARγ is thought to have a larger ligand-binding pocket compared to that of other nuclear receptors (24), which may allow PLP-derived compounds to act as ligands. However, observations in this study raise a critical question concerning the selectivity of PLP for PPARγ target genes. As shown in Fig. 2, PLP was shown to up-regulate aP2 and Gyk mRNA expression, but not lipoprotein lipase (LPL) (data not shown), which is also a well-known PPARγ target gene (25), in 3T3-L1 cells. There is increasing evidence that transcriptional regulation varies even among the PPARγ-responsive genes via distinct mechanisms (26–28), and, in fact, several isoprenols were found to induce the expression of aP2 mRNA, but not LPL mRNA in 3T3-L1 cells (29), suggesting that PLP may act as a partial (or weak) agonist similar to the isoprenols.
Second, PLP may influence the interaction of cofactors with PPARγ, as it has been proposed that ligand-induced transcriptions of PPARγ target genes are mediated via the recruitment of distinct cofactors (26–28). Recently, Huq et al found an important role for vitamin B6 in gene regulation by direct PLP conjugation to receptor interacting protein 140 (RIP140) (30). RIP140 was originally identified as a ligand-dependent co-repressor which binds to estrogen receptors, and, to date, has been shown to interact with and repress a number of other nuclear receptors, including thyroid hormone receptors and estrogen-related receptors (31). Leonardsson et al reported that RIP140-null mice show defects in fat accumulation and energy expenditure in adipose tissues, suggesting an important role of RIP140 in regulating the balance between energy storage and energy expenditure (32). Furthermore, in RIP140-null adipocytes, energy expenditure was reportedly elevated with high expression levels of uncoupling protein 1 (Ucp1) and carnitine palmitoyltransferase 1b mRNAs (33). Debevec et al proposed that the absence of RIP140 leads to the recruitment of PPARγ, together with PPARα and estrogen receptor α, to the UCP1 enhancer to allow activation of Ucp1 gene transcription (34). These observations prompted us to further consider the physiological significance of the conjugation between PLP and RIP140 for PPARγ activation in adipocytes.
In summary, these observations presented here show that vitamin B6 up-regulates the expression of PPARγ target genes in vitro and in vivo. This study indicates a novel physiological function of vitamin B6 which may be, in part, mediated by PPARγ activation. The anti-tumor and anti-inflammatory effects of vitamin B6, furthermore, may relate to PPARγ activation. Further study is necessary to elucidate the molecular mechanism underlying the action of vitamin B6 on PPARγ.
Acknowledgments
We are grateful to Drs Yasutomi Kamei and Junko Mizukami for their technical advice. This study was supported, in part, by a Grant-in-Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science and Technology of Japan.
Abbreviations:
- PPARγ,
peroxisome proliferator-activated receptor-γ;
- TZD,
thiazolidinedione;
- PLP,
pyridoxal 5′-phosphate;
- PBS,
phosphate-buffered saline
References
- 1.Moon WH, Kirksey A. Cellular growth during prenatal and early postnatal periods in progeny of pyridoxine-deficient rats. J Nutr. 1973;103:123–133. doi: 10.1093/jn/103.1.123. [DOI] [PubMed] [Google Scholar]
- 2.Haynes WG. Hyperhomocysteinemia, vascular function and atherosclerosis: effects of vitamins. Cardiovasc Drugs Ther. 2002;16:391–399. doi: 10.1023/a:1022130217463. [DOI] [PubMed] [Google Scholar]
- 3.Cattaneo M, Lombardi R, Lecchi A, Bucciarelli P, Mannucci PM. Low plasma levels of vitamin B(6) are independently associated with a heightened risk of deep-vein thrombosis. Circulation. 2001;104:2442–2446. doi: 10.1161/hc4501.098925. [DOI] [PubMed] [Google Scholar]
- 4.Onorato JM, Jenkins AJ, Thorpe SR, Baynes JW. Pyridoxamine, an inhibitor of advanced glycation reactions, also inhibits advanced lipoxidation reactions. Mechanism of action of pyridoxamine. J Biol Chem. 2000;275:21177–21184. doi: 10.1074/jbc.M003263200. [DOI] [PubMed] [Google Scholar]
- 5.Komatsu SI, Watanabe H, Oka T, Tsuge H, Nii H, Kato N. Vitamin B-6-supplemented diets compared with a low vitamin B-6 diet suppress azoxymethane-induced colon tumorigenesis in mice by reducing cell proliferation. J Nutr. 2001;131:2204–2207. doi: 10.1093/jn/131.8.2204. [DOI] [PubMed] [Google Scholar]
- 6.Komatsu S, Watanabe H, Oka T, Tsuge H, Kat N. Dietary vitamin B6 suppresses colon tumorigenesis, 8-hydroxyguanosine, 4-hydroxynonenal, and inducible nitric oxide synthase protein in azoxymethane-treated mice. J Nutr Sci Vitaminol. 2002;48:65–68. doi: 10.3177/jnsv.48.65. [DOI] [PubMed] [Google Scholar]
- 7.Matsubara K, Mori M, Matsuura Y, Kato N. Pyridoxal 5′-phosphate and pyridoxal inhibit angiogenesis in serum-free rat aortic ring assay. Int J Mol Med. 2001;8:505–508. doi: 10.3892/ijmm.8.5.505. [DOI] [PubMed] [Google Scholar]
- 8.Yanaka N, Koyama TA, Komatsu S, Nakamura E, Kanda M, Kato N. Vitamin B6 suppresses NF-κB activation in LPS-stimulated mouse macrophages. Int J Mol Med. 2005;16:1071–1075. [PubMed] [Google Scholar]
- 9.Kersten S, Desvergne B, Wahli W. Roles of PPARs in health and disease. Nature. 2000;405:421–424. doi: 10.1038/35013000. [DOI] [PubMed] [Google Scholar]
- 10.Braissant O, Foufelle F, Scotto C, Dauca M, Wahli W. Differential expression of peroxisome proliferator-activated receptors (PPARs): tissue distribution of PPAR-alpha, -beta, and -gamma in the adult rat. Endocrinology. 1996;137:354–366. doi: 10.1210/endo.137.1.8536636. [DOI] [PubMed] [Google Scholar]
- 11.Chawla A, Schwarz EJ, Dimaculangan DD, Lazar MA. Peroxisome proliferator-activated receptor (PPAR) gamma: adipose-predominant expression and induction early in adipocyte differentiation. Endocrinology. 1994;135:798–800. doi: 10.1210/endo.135.2.8033830. [DOI] [PubMed] [Google Scholar]
- 12.Tontonoz P, Hu E, Graves RA, Budavari AI, Spiegelman BM. mPPAR gamma 2: tissue-specific regulator of an adipocyte enhancer. Genes Dev. 1994;8:1224–1234. doi: 10.1101/gad.8.10.1224. [DOI] [PubMed] [Google Scholar]
- 13.Lehmann JM, Moore LB, Smith-Oliver TA, Wilkison WO, Willson TM, Kliewer SA. An antidiabetic thiazolidinedione is a high affinity ligand for peroxisome proliferator-activated receptor gamma (PPAR gamma) J Biol Chem. 1995;270:12953–12956. doi: 10.1074/jbc.270.22.12953. [DOI] [PubMed] [Google Scholar]
- 14.Forman BM, Tontonoz P, Chen J, Brun RP, Spiegelman BM, Evans RM. 15-Deoxy-delta 12, 14-prostaglandin J2 is a ligand for the adipocyte determination factor PPAR gamma. Cell. 1995;83:803–812. doi: 10.1016/0092-8674(95)90193-0. [DOI] [PubMed] [Google Scholar]
- 15.Grommes C, Landreth GE, Heneka MT. Antineoplastic effects of peroxisome proliferator-activated receptor gamma agonists. Lancet Oncol. 2004;5:419–429. doi: 10.1016/S1470-2045(04)01509-8. [DOI] [PubMed] [Google Scholar]
- 16.Sarraf P, Mueller E, Jones D, King FJ, DeAngelo DJ, Partridge JB, Holden SA, Chen LB, Singer S, Fletcher C, Spiegelman BM. Differentiation and reversal of malignant changes in colon cancer through PPARgamma. Nat Med. 1998;4:1046–1052. doi: 10.1038/2030. [DOI] [PubMed] [Google Scholar]
- 17.Tanaka T, Kohno H, Yoshitani S, Takashima S, Okumura A, Murakami A, Hosokawa M. Ligands for peroxisome proliferator-activated receptors alpha and gamma inhibit chemically induced colitis and formation of aberrant crypt foci in rats. Cancer Res. 2001;61:2424–2428. [PubMed] [Google Scholar]
- 18.Osawa E, Nakajima A, Wada K, Ishimine S, Fujisawa N, Kawamori T, Matsuhashi N, Kadowaki T, Ochiai M, Sekihara H, Nakagama H. Peroxisome proliferator-activated receptor gamma ligands suppress colon carcinogenesis induced by azoxymethane in mice. Gastroenterology. 2003;124:361–367. doi: 10.1053/gast.2003.50067. [DOI] [PubMed] [Google Scholar]
- 19.Welch JS, Ricote M, Akiyama TE, Gonzalez FJ, Glass CK. PPARgamma and PPARdelta negatively regulate specific subsets of lipopolysaccharide and IFN-gamma target genes in macrophages. Proc Natl Acad Sci USA. 2003;100:6712–6717. doi: 10.1073/pnas.1031789100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Appel S, Mirakaj V, Bringmann A, Weck MM, Grunebach F, Brossart P. PPAR-gamma agonists inhibit toll-like receptor-mediated activation of dendritic cells via the MAP kinase and NF-kappaB pathways. Blood. 2005;106:3888–3894. doi: 10.1182/blood-2004-12-4709. [DOI] [PubMed] [Google Scholar]
- 21.Kawada T, Aoki N, Kamei Y, Maeshige K, Nishiu S, Sugimoto E. Comparative investigation of vitamins and their analogues on terminal differentiation, from preadipocytes to adipocytes, of 3T3-L1 cells. Comp Biochem Physiol A. 1990;96:323–326. doi: 10.1016/0300-9629(90)90699-s. [DOI] [PubMed] [Google Scholar]
- 22.Yanaka N, Nogusa Y, Fujioka Y, Yamashita Y, Kato N. Involvement of membrane protein GDE2 in retinoic acid-induced neurite formation in Neuro2A cells. FEBS Lett. 2007;581:712–718. doi: 10.1016/j.febslet.2007.01.035. [DOI] [PubMed] [Google Scholar]
- 23.Terzuoli L, Leoncini R, Pagani R, Guerranti R, Vannoni D, Ponticelli F, Marinello E. Some chemical properties and biological role of thiazolidine compounds. Life Sci. 1998;63:1251–1267. doi: 10.1016/s0024-3205(98)00387-7. [DOI] [PubMed] [Google Scholar]
- 24.Uppenberg J, Svensson C, Jaki M, Bertilsson G, Jendeberg L, Berkenstam A. Crystal structure of the ligand binding domain of the human nuclear receptor PPARgamma. J Biol Chem. 1998;273:31108–31112. doi: 10.1074/jbc.273.47.31108. [DOI] [PubMed] [Google Scholar]
- 25.Schoonjans K, Peinado-Onsurbe J, Lefebvre AM, Heyman RA, Briggs M, Deeb S, Staels B, Auwerx J. PPARalpha and PPARgamma activators direct a distinct tissue-specific transcriptional response via a PPRE in the lipoprotein lipase gene. EMBO J. 1996;15:5336–5348. [PMC free article] [PubMed] [Google Scholar]
- 26.Kodera Y, Takeyama K, Murayama A, Suzawa M, Masuhiro Y, Kato S. Ligand type-specific interactions of peroxisome proliferator-activated receptor gamma with transcriptional coactivators. J Biol Chem. 2000;275:33201–33204. doi: 10.1074/jbc.C000517200. [DOI] [PubMed] [Google Scholar]
- 27.Robinson CE, Wu X, Nawaz Z, Onate SA, Gimble JM. A corepressor and chicken ovalbumin upstream promoter transcriptional factor proteins modulate peroxisome proliferator-activated receptor-gamma2/retinoid X receptor alpha-activated transcription from the murine lipoprotein lipase promoter. Endocrinology. 1990;140:1586–1593. doi: 10.1210/endo.140.4.6653. [DOI] [PubMed] [Google Scholar]
- 28.Guan HP, Ishizuka T, Chui PC, Lehrke M, Lazar MA. Corepressors selectively control the transcriptional activity of PPARgamma in adipocytes. Genes Dev. 2005;19:453–461. doi: 10.1101/gad.1263305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Takahashi N, Kawada T, Goto T, Yamamoto T, Taimatsu A, Matsui N, Kimura K, Saito M, Hosokawa M, Miyashita K, Fushiki T. Dual action of isoprenols from herbal medicines on both PPARgamma and PPARalpha in 3T3-L1 adipocytes and HepG2 hepatocytes. FEBS Lett. 2002;514:315–322. doi: 10.1016/s0014-5793(02)02390-6. [DOI] [PubMed] [Google Scholar]
- 30.Huq MD, Tsai NP, Lin YP, Higgins L, Wei LN. Vitamin B6 conjugation to nuclear corepressor RIP140 and its role in gene regulation. Nat Chem Biol. 2007;3:161–165. doi: 10.1038/nchembio861. [DOI] [PubMed] [Google Scholar]
- 31.Fritah A, Christian M, Parker MG. The metabolic coregulator RIP140: an update. Am J Physiol endocrinol Metab. 2010;299:E335–E340. doi: 10.1152/ajpendo.00243.2010. [DOI] [PubMed] [Google Scholar]
- 32.Leonardsson G, Steel JH, Christian M, Pocock V, Milligan S, Bell J, So PW, Medina-Gomez G, Vidal-Puig A, White R, Parker MG. Nuclear receptor corepressor RIP140 regulates fat accumulation. Proc Natl Acad Sci USA. 2004;101:8437–8442. doi: 10.1073/pnas.0401013101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Christian M, Kiskinis E, Debevec D, Leonardsson G, White R, Parker MG. RIP140-targeted repression of gene expression in adipocytes. Mol Cell Biol. 2005;25:9383–9391. doi: 10.1128/MCB.25.21.9383-9391.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Debevec D, Christian M, Morganstein D, Seth A, Herzog B, Parker M, White R. Receptor interacting protein 140 regulates expression of uncoupling protein 1 in adipocytes through specific peroxisome proliferator activated receptor isoforms and estrogen-related receptor alpha. Mol Endocrinol. 2007;21:1581–1592. doi: 10.1210/me.2007-0103. [DOI] [PMC free article] [PubMed] [Google Scholar]