

Abstract
Antibiotic resistance is a major challenge to modern medicine. Intraspecies and interspecies dissemination of antibiotic resistance genes among bacteria can occur through horizontal gene transfer. Competence-mediated gene transfer has been reported to contribute to the spread of antibiotic resistance genes in Streptococcus pneumoniae. Induction of the competence regulon is mediated by a 17-amino acid peptide pheromone called the competence stimulating peptide (CSP). Thus, synthetic analogs that competitively inhibit CSPs may reduce horizontal gene transfer. We performed saturated alanine scanning mutagenesis and other amino acid substitutions on CSP1 to screen for analogs that disable genetic transformation in S. pneumoniae. Substitution of the glutamate residue at the first position created analogs that could competitively inhibit CSP1-mediated competence development in a concentration-dependent manner. Additional substitutions of the negatively-charged glutamate residue with amino acids of different charge, acidity and hydrophobicity, as well as enantiomeric D-glutamate, generated analogs that efficiently outcompeted CSP1, suggesting the importance of negative charge and enantiomericity of the first glutamate residue for the function of CSP1. Collectively, these results indicate that glutamate residue at the first position is important for the ability of CSP1 to induce ComD, but is dispensable for the peptide to bind the receptor. Furthermore, these results demonstrate the potential applicability of competitive CSP analogs to control horizontal transfer of antibiotic resistance genes in S. pneumoniae.
Introduction
Streptococcus pneumoniae is a major cause of numerous diseases worldwide, including pneumonia, otitis media, meningitis, bacteremia and sepsis [1]. In recent decades, antibiotic resistant S. pneumoniae have been increasingly isolated in clinical settings [2]–[8]. Genetic transformation, which occurs when S. pneumoniae enters the competent state, contributes to the transfer and acquisition of antibiotic resistance genes [9]–[11]. The competence regulon is activated when CSP binds to its membrane-associated histidine kinase receptor ComD, which phosphorylates the response regulator ComE [12]–[15]. In turn, ComE activates the transcription of genes in the competence regulon, including the transcription of ComX, a competence specific sigma factor. ComX positively regulates the transcription of genes encoding effectors for DNA uptake and recombination [16]. Interestingly, we and others have shown that the competence regulon is also important for virulence during acute pneumonia and bacteremia models of mouse infection [17]–[22].
DNA sequence analysis predicts the existence of six distinct CSP subtypes, with an overwhelming majority of S. pneumoniae strains producing CSP1 or CSP2 [23], [24], corresponding to two variants of ComD, namely ComD1 and ComD2 [25]. Competence in ComD1 strains could be induced more efficiently with the “compatible” CSP1. In contrast, ComD2 strains are more sensitive to induction by the “compatible” CSP2 [26], [27].
Alanine scanning was previously used to identify amino acid residues that are important for the activity of CSP1 [28]. However, the author did not test the ability of these analogs to competitively inhibit CSP1. Most recently, by using amino acid substitution and deletion, we have identified synthetic analogs of CSP1 and CSP2 that competitively inhibit CSPs’ ability to induce the competence regulon, to control pneumococcal pneumonia and to reduce horizontal gene transfer of an antibiotic resistance gene during infection [27]. In this study, we performed a saturated alanine scanning mutagenesis of CSP1, as well as substitution of amino acids based on differing charges, acidity, hydrophobicity and enantiomericity, and examined the ability of these analogs to inhibit the development of competence.
Results
Saturated Alanine Mutagenesis Screen Identifies CSP1 Analogs That are Unable to Induce comX Expression and Genetic Transformation
We performed a saturated alanine scanning mutagenesis on CSP1 to identify additional synthetic analogs that would outcompete CSP1 and attenuate competence development and genetic transformation (Fig. 1). The ability of CSP1 and its analogs to induce competence in the wild-type S. pneumoniae strain D39 was compared by monitoring the: (i) promoter activity of comX, (ii) transformation frequency of rpsL gene that confers resistance to streptomycin [29]. Induction of ComX indicates that S. pneumoniae cells have entered the competent state for genetic transformation. Activation of the comX gene promoter was monitored by assaying ß-galactosidase activity in pneumococcal strain D39pcomX::lacZ [27]. The ability of several CSP1 analogs to activate the expression of comX, including CSP1-E1A, CSP1-R3A, CSP1-F7A, CSP1-F8A and CSP1-F11A, were attenuated by 95.5, 100, 95, 92.5, 95%, respectively (Fig. 2A). Because ComX regulates DNA uptake and transformation, we examined the ability of all analogs to induce genetic transformation. Consistent with their reduced ability to induce the expression of comX, CSP1-E1A, CSP1-R3A, CSP1-F7A, CSP1-F8A and CSP1-F11A were attenuated in their ability to induce genetic transformation, by 99.9, 100, 77.1, 68.9 and 84.5%, respectively (Fig. 2B).
Figure 1. Amino acid sequences of CSP1 and its analogs.

Analogs were synthesized based on the sequence of CSP1 by alanine substitutions.
Figure 2. Analyses of the ability of CSP1 analogs to induce comX expression and genetic transformation.

Pneumococcal cells were incubated with CSP1 or with each analog at a final concentration of 100 ng/ml. The ability of each peptide to induce the competence regulon was measured by induction of the comX expression using the ß-galactosidase assays in D39pcomX::lacZ cells (A) or by genetic transformation using the rpsL gene in D39 cells (B). Transformants were selected on THB agar containing 100 µg/ml streptomycin. Experiments were performed in triplicates and repeated three times. The means ± SD of one typical experiment are shown.
CSP1-E1A is An Analog that Competitively Inhibits CSP1
Because of their inability to induce comX expression and genetic transformation, we next determined whether these five analogs could competitively inhibit wild type CSP1. Analogs CSP1-R3A, CSP1-F7A, CSP1-F8A and CSP1-F11A failed to inhibit CSP1-induced comX expression (Fig. 3A), suggesting that these analogs were incapable of binding to ComD1 receptor. In contrast, CSP1-E1A, with glutamate residue at the first position substituted with alanine, competitively inhibited CSP1 in a concentration-dependent manner. The inductive activity of 100 ng CSP1 on comX expression was inhibited by 69, 86.7, 94 and 97% in the presence of 100, 200, 400 or 800 ng of CSP1-E1A, respectively (Fig. 3A). Similarly, CSP1-mediated genetic transformation was inhibited by 81.6, 94.4, 98.5 and 99.8% in the presence of indicated concentrations of CSP1-E1A, respectively (Fig. 3B). These results suggest that among the five CSP1 analogs with defective ability to activate the competence regulon, only CSP1-E1A is uniquely capable of competitively inhibiting CSP1.
Figure 3. CSP1-E1A is uniquely able to inhibit the expression of comX and genetic transformation.

(A) CSP-E1A competitively inhibits CSP1-mediated induction of comX in a dose dependent manner. D39pcomX::lacZ cells were exposed to 100 ng/ml of CSP1 alone or simultaneously with increasing concentrations of each CSP1 analog. The activity of the comX gene promoter was measured by β-galactosidase activity. (B) CSP-E1A competitively inhibits CSP1-mediated genetic transformation. Genetic transformation was performed using 30 µg/ml of the D39 genomic DNA containing the rpsL gene. Transformants were selected on THB agar containing 100 µg/ml streptomycin. Experiments were performed in triplicates and repeated three times. The means ± SD of one typical experiment are shown.
Negative Charge and Enantiomericity of the First Glutamate Residue are Important for the Activity of CSP1
Glutamate is polar and negatively charged. Substitution of glutamate with alanine (nonpolar, neutral) at the first position generates a strong competitive inhibitor of CSP1 (Figure 3). This observation suggests that the negative charge of glutamate is important for the activity of CSP1, but dispensable for the binding to ComD1. To determine the importance of negative charge on glutamate to the activity of CSP1, we examined five additional synthetic analogs (Fig. 4A), by substituting the glutamate with glutamine (CSP1-E1Q, polar, neutral), leucine (CSP1-E1L, non-polar, neutral), aspartic acid (CSP1-E1D, polar, negatively-charged), and arginine (CSP1-E1R, polar, positively-charged), and an enantiomeric analog using the D-glutamate (CSP1-D-E1, polar, negatively-charged) (Fig. 4A), for their ability to competitively inhibit CSP1. All five new analogs were attenuated in their ability to induce the expression of comX (Fig. 4B). Importantly, the analog CSP1-E1D, in which the glutamate residue is substituted with another polar and negatively-charged amino acid aspartate, retained 35.6% of the wild-type CSP1 activity to induce the expression of comX (Fig. 4A), confirming that negative charge of the glutamate residue is important for the activity of CSP1. Concurring with this hypothesis, we showed that CSP1-E1R (polar, positive-charged arginine) was most attenuated, with the levels of comX induction and genetic transformation reduced to 4.6% and 0% (800 ng/ml), respectively (Fig. 4A–B). In addition, CSP1-E1L and CSP1-E1Q were also able to induce comX expression and genetic transformation, but to lesser degrees than CSP1-E1D. Finally, substitution of L-glutamate in the first position of CSP1 with its enantiomeric isomer D-glutamate also impaired its ability to induce comX expression and genetic transformation (Fig. 4A–B). Collectively, these results suggest that negative charge and structural conformation in the N-terminus of CSP1 are important for its activity.
Figure 4. The negative charge and enantiomericity of glutamate residue in the first position is important for the activity of CSP1.
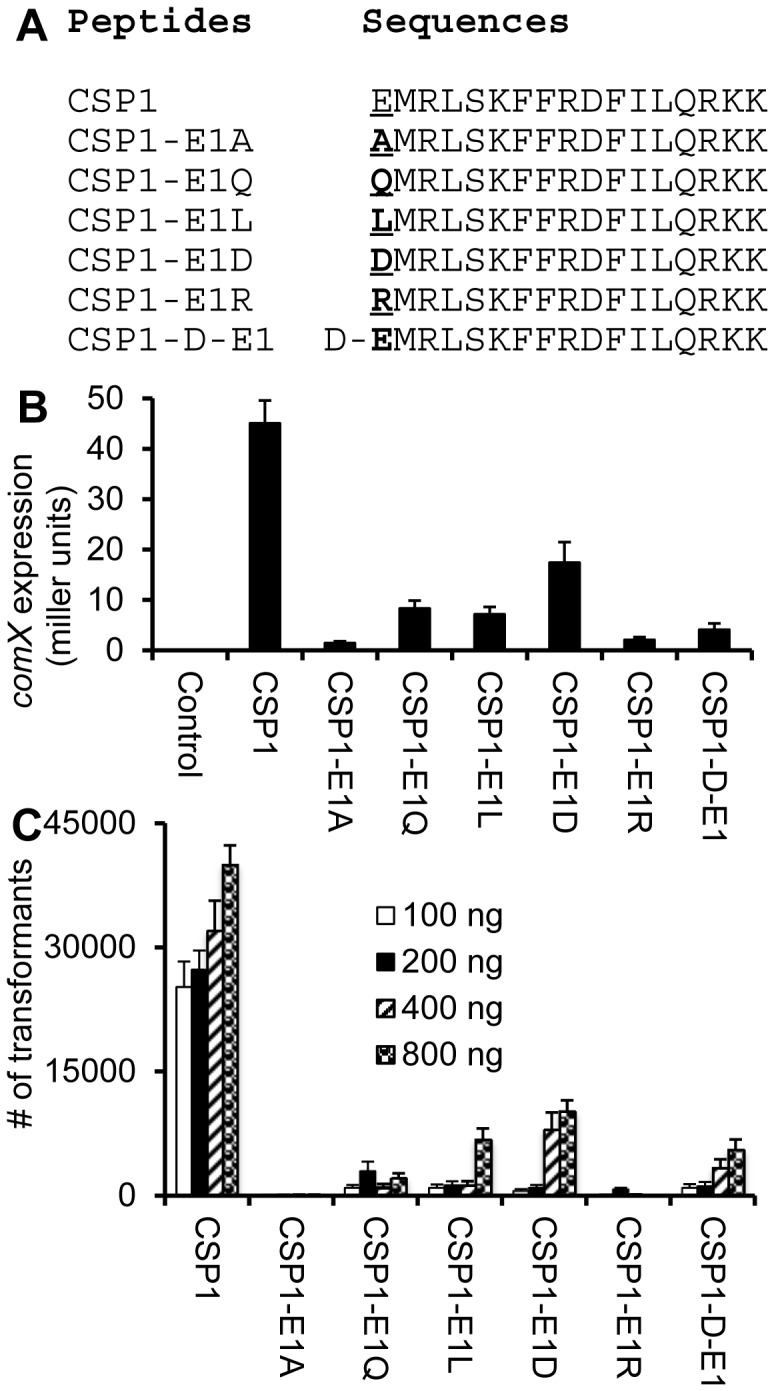
(A) Amino acid substitutions of the first glutamate residue in CSP1. (B) Induction of comX expression by CSP1 analogs. D39pcomX::lacZ were incubated with CSP1 or with each glutamate substituted analog at a final concentration of 100 ng/ml. The ability of each peptide to induce the competence regulon was measured by induction of the comX by ß-galactosidase assays. (C) Genetic transformation of D39 cells in the presence of indicated concentrations of CSP1 and its analogs using the rpsL gene. Transformants were selected on THB agar containing 100 µg/ml streptomycin. Experiments in B and C were performed in triplicates and repeated three times. The means ± SD of one typical experiment are shown.
Differential Amino Acid Substitutions of Glutamate at the First Position Generate New Inhibitors of CSP1
Next, we examined whether these CSP1 analogs with substitution at the glutamate residue could competitively inhibit the induction of comX and genetic transformation. All analogs inhibited comX induction and ComX-dependent genetic transformation to similar levels (Fig. 5A–B). The lack of distinction between the ability of CSP1-E1A and CSP1-E1R versus CSP1-E1D was surprising, probably caused by high amounts of analogs (100 ng/ml) that were used for these assays. To overcome the aforementioned difficulty, we examined CSP1 inhibition using lower concentrations of these analogs (Fig. 6). Analogs CSP1-E1R, CSP1-D-E1 and CSP1-E1Q had low ability to induce genetic transformation (Fig. 6A). Interestingly, at low concentrations, CSP1-E1Q and CSP1-E1D were found to be most effective at inhibiting CSP1-mediated transformation (Fig. 6B). Surprisingly, CSP1-E1R, which had the lowest ability to induce comX and genetic transformation (Figs. 6A), was least effective in inhibiting genetic transformation at low peptide concentrations (Fig. 6B). Conversely, CSP1-E1D, which retained one of the highest activities to induce comX expression and genetic transformation (Fig. 6A), was the most effective competitive inhibitor at low peptide concentrations (Fig. 6B). Finally, CSP1-E1Q, which was highly attenuated in its ability to induce comX expression and genetic transformation (Fig. 6A), was as effective as CSP1-E1D at competitively inhibiting CSP1 at low peptide concentrations (Fig. 6B).
Figure 5. Dose dependent inhibition of comX expression and genetic transformation by CSP1 analogs with substitution at glutamate residue in the first position.

(A) CSP1 analogs with substitution at the first glutamate acid competitively inhibit CSP1-mediated induction of comX. D39pcomX::lacZ cells were exposed to 100 ng/ml of CSP1 alone or simultaneously with increasing concentrations of CSP1 analogs. The activity of the comX gene promoter was measured by β-galactosidase activity. Experiments were performed in triplicates and repeated three times. The means ± SD of one typical experiment are shown. (B) CSP1 analogs with substitution at the first glutamate acid competitively inhibit the ability of CSP1 to induce genetic transformation. Genetic transformation was performed using the rpsL gene. Transformants were selected on THB agar containing 100 µg/ml streptomycin. Experiments were performed in triplicates and repeated three times. The means ± SD of one typical experiment are shown.
Figure 6. Competitive inhibition of CSP1 by low concentrations of analogs with substitutions of glutamate residue in the first position.
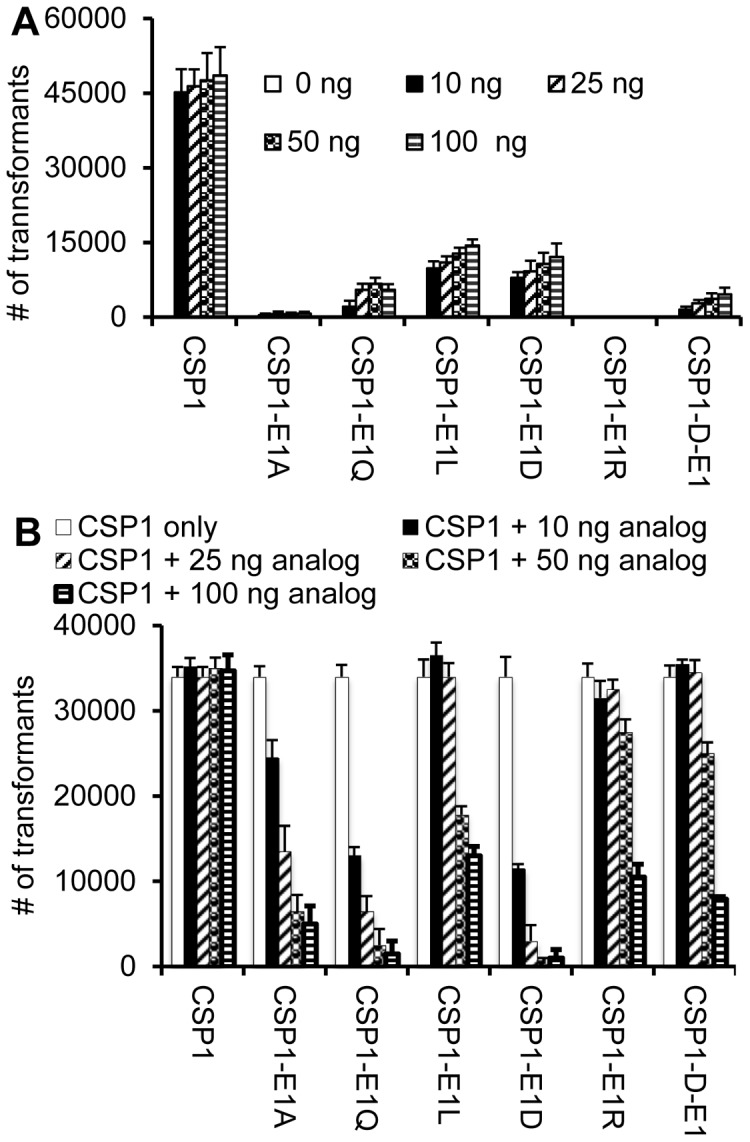
(A) Induction of genetic transformation by each analog using the genomic DNA containing the rpsL gene. Transformants were selected on THB agar containing 100 µg/ml streptomycin. Experiments were performed in triplicates and repeated three times. The means ± SD of one typical experiment are shown. (B) Competitive inhibition of genetic transformation by low concentrations of CSP1 analogs. Genetic transformation was performed using the genomic DNA containing the rpsL gene. Transformants were selected on THB agar containing 100 µg/ml streptomycin. Experiments were performed in triplicates and repeated three times. The means ± SD of one typical experiment are shown.
Discussion
The competence regulon regulates genetic transformation [11], [30] and virulence in S. pneumoniae [17]–[22]. In this study, we used saturated alanine scanning to identify synthetic analogs of CSP1 that could competitively inhibit genetic transformation of an antibiotic resistance gene. Substitutions of the 1st (CSP1-E1A), 3rd (CSP1-R3A), 7th (CSP1-F7A), 8th (CSP1-F8A) and 11th (CSP1-F11A) amino acid residues with alanine generate synthetic analogs with impaired ability to induce the competence regulon. However, only substitutions of the glutamate residue at the first position generate analogs that competitively inhibit CSP1. These results suggest that the glutamate residue is important for the ability of CSP1 to induce ComD, but is dispensable for binding to the histidine kinase receptor.
The chemical properties of individual amino acid residues, including charges, acidity, polarity and hydrophobicity may influence the activity of a peptide. Glutamate is negatively charged. The importance of negative charge is demonstrated by the analog CSP1-E1D, which has the glutamate in the first position substituted with negatively charged aspartate, preserves significant amounts of ability to induce competence regulon. This suggests that CSP1-E1D retains significant amount of binding to ComD1, and is thus, paradoxically, more efficient in competitively displacing the binding of CSP1 to ComD1. This argument is supported by the observation that CSP1-E1R, which the glutamate has been substituted with positively-charged arginine (CSP1-E1R), is least effective in inhibiting CSP1. Another highly effective inhibitory analog is CSP1-E1Q. Glutamine is neutrally charged, yet structurally most similar to glutamate. Structural similarity and lesser charge difference between glutamate-glutamine versus glutamate-arginine may have contributed to the different competitiveness between CSP1-E1Q versus CSP1-E1R. Because of its low ability to induce competence and genetic transformation, yet strong ability to competitively inhibit CSP1, we predict that CSP1-E1Q is the best inhibitor analog. Finally, the L-glutamate conformation is also important because enantiomeric substitution with D-glutamate severely reduces the activity of CSP1.
The use of inhibitory analogs of CSP1 may have an application in reducing horizontal transfer of antibiotic resistance genes. For example, competence in genetic transformation is induced by CSP1 and other peptide pheromones in streptococcal species by two major systems [13]. These systems differ in their methods of inducing the expression of ComX. Streptococcal species in Anginosus and Mitis groups use the ComCDE system to induce comX expression. Thus, CSP analogs that inhibit ComD to suppress competence may be applicable in these streptococcal species. The second system, ComRS, is utilized by streptococcal species in Pyogenic, Salvarius and Bovis groups [31]. In this system, the precursor of the competence peptide ComS is cleaved by an extracellular protease, and the processed peptide re-enters the cytoplasm through peptide permease to activate the cytoplasmic transcriptional regulator ComR to initiate the expression of comX. It will be interesting to explore if analogs of ComS could inhibit competence in the relevant streptococcal species.
In recent decades, incidence of S. pneumoniae resistance to β-lactams, macrolides, and other classes of antibiotics has escalated dramatically [2]–[8]. While conjugative elements appear to be more important for the spread of antibiotic resistance genes [4], [32]–[34], horizontal gene transfer of penicillin binding protein genes has occurred in clinical isolates of S. pneumoniae [10]. Importantly, it has been shown that under in vitro conditions, antibiotic stress stimulates horizontal gene transfer in bacteria, including S. pneumoniae [35]–[37]. For example, β-lactams induce the SOS response and horizontal transfer of virulence factors in Staphylococcus aureus [36]. Similarly, DNA damaging antibiotics trigger genetic exchange in Helicobacter pylori [35]. In this case, a coupling agent that inhibits horizontal gene transfer reduces the risk of generating antibiotic resistant pathogens during antibiotic treatment. In S. pneumoniae, antibiotic stress has been reported to induce the competence regulon and increase genetic transformation [37]. These authors also reported that induction of competence by antibiotic stress does not occur in a comA mutant that lacks the ABC transporter needed to export CSP, suggesting that this process is CSP-dependent. Collectively, these studies suggest that under certain clinical conditions, S. pneumoniae actively exploits the opportunity of antibiotic stress to acquire exogenous genes. Thus, antibiotic resistance mediated by the competence regulon may be under reported and worth more detailed investigations.
In conclusion, our data show that CSP analogs can effectively attenuate genetic transformation in S. pneumoniae. This strategy may be applicable to reduce the incidence of horizontal gene transfer and acquisition of antibiotic resistance genes. In addition, because competence systems in some streptococcal species, including S. mitis, are very similar to that of S. pneumoniae, there is a possibility that the former could be transformed by pneumococcal DNA [38], [39]. S. mitis is generally considered an avirulent species but has the potential to acquire virulence determinants from S. pneumoniae and transform itself into a pathogen. In this scenario, CSP1 analogs may be applicable to reduce the emergence of the new pathogen.
Materials and Methods
Bacterial Strains and Growth Conditions
S. pneumoniae strain D39 [40] was a generous gift from Dr. David Briles (University of Alabama-Birmingham). Strain D39pcomX::lacZ was generated by transforming D39 with the genomic DNA from CPM3 that harbored an insertion of the lacZ gene under the control of the comX promoter [27]. Aliquots of bacteria were stored at −80°C in Todd Hewitt broth (THB) containing 25% glycerol. For routine experiments, bacteria from frozen stocks were streaked onto THB agar containing 5% defibrinated horse blood and incubated for 12–24 hr at 37°C with 5% CO2. Fresh colonies were transferred to THB and cultured to desired density as measured by a spectrophotometer at OD 600 nm.
Synthetic CSPs
Both CSP1 and its analogs (≥95% purity) were synthesized by Elim Biopharm (Hayward, CA).
Activation Assay of comX
The ability of synthetic CSP-1 and its analogs to activate the expression of comX gene was compared in D39pcomX::lacZ cells grown in THB (pH 6.8) until OD 600 nm of 0.1, washed and resuspended in THB (pH 8.3). CSP1 and their analogs were added at indicated concentrations to the culture and incubated at 37°C for 30 min. ß-galactosidase activity in pneumococcal cells was measured according to previously published protocols [26], and expressed as Miller units.
S. pneumoniae Transformation Assay
Genetic transformation experiments were performed as we had previously described [18], [26]. Briefly, S. pneumoniae cells were grown to early log phase (OD 600 nm ∼ 0.1) in THB (pH 6.8), washed and resuspended in THB (pH 8.3) containing 30 µg/ml of D39 genomic DNA carrying a copy of the streptomycin resistance rpsL gene originated from strain CP1296 [41]. Transformation experiments were performed in the present CSP1 or its analogs. The transformation mix was incubated at 37°C with 5% CO2 for 1 hr. Transformants were selected on THB agar supplemented with 100 µg/ml streptomycin.
Acknowledgments
We thank Professor David Briles (University of Alabama at Birmingham College of Medicine) for his generous gift of S. pneumoniae strain D39; and Professor Donald Morrison (University of Illinois at Chicago) for his generous gift of genomic DNA samples containing the rpsL and pcomX-lacZ constructs, and his technical and biological insights on pneumococcal competence. We thank Ms. Jennifer Ida for critical reading and editing of the manuscript.
Funding Statement
This work was partially supported by the National Institutes of Health (HL090699) and a University of Illinois Research Board Arnold O. Beckman research award to G.W. Lau. This investigation was conducted in a facility constructed with the support from Research Facilities Improvement Program Grant Number C06 RR 16515-01 from the National Center for Research Resources, National Institutes of Health. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Kadioglu A, Weiser JN, Paton JC, Andrew PW (2008) The role of Streptococcus pneumoniae virulence factors in host respiratory colonization and disease. Nat Rev Microbiol 6: 288–301. [DOI] [PubMed] [Google Scholar]
- 2. Klugman KP, Lonks JR (2005) Hidden epidemic of macrolide-resistant pneumococci. Emerg Infect Dis 11: 802–807. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Jacobs MR (2008) Antimicrobial-resistant Streptococcus pneumoniae: trends and management. Expert Rev Anti Infect Ther 6: 619–635. [DOI] [PubMed] [Google Scholar]
- 4. Croucher NJ, Walker D, Romero P, Lennard N, Paterson GK, et al. (2009) Role of conjugative elements in the evolution of the multidrug-resistant pandemic clone Streptococcus pneumoniaeSpain23F ST81. J Bacteriol 191: 1480–1489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Coker TR, Chan LS, Newberry SJ, Limbos MA, Suttorp MJ, et al. (2010) Diagnosis, microbial epidemiology, and antibiotic treatment of acute otitis media in children: a systematic review. JAMA 304: 2161–2169. [DOI] [PubMed] [Google Scholar]
- 6. Linares J, Ardanuy C, Pallares R, Fenoll A (2010) Changes in antimicrobial resistance, serotypes and genotypes in Streptococcus pneumoniae over a 30-year period. Clin Microbiol Infect 16: 402–410. [DOI] [PubMed] [Google Scholar]
- 7. Lynch JP, 3rd, Zhanel GG (2010) Streptococcus pneumoniae: epidemiology and risk factors, evolution of antimicrobial resistance, and impact of vaccines. Curr Opin Pulm Med 16: 217–225. [DOI] [PubMed] [Google Scholar]
- 8. Jones RN, Jacobs MR, Sader HS (2010) Evolving trends in Streptococcus pneumoniae resistance: implications for therapy of community-acquired bacterial pneumonia. Int J Antimicrob Agents 36: 197–204. [DOI] [PubMed] [Google Scholar]
- 9. Dowson CG, Hutchison A, Brannigan JA, George RC, Hansman D, et al. (1989) Horizontal transfer of penicillin-binding protein genes in penicillin-resistant clinical isolates of Streptococcus pneumoniae. Proc Natl Acad Sci U S A 86: 8842–8846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Morrison DA, Lee MS (2000) Regulation of competence for genetic transformation in Streptococcus pneumoniae: a link between quorum sensing and DNA processing genes. Res Microbiol 151: 445–451. [DOI] [PubMed] [Google Scholar]
- 11. Claverys JP, Martin B, Polard P (2009) The genetic transformation machinery: composition, localization, and mechanism. FEMS Microbiol Rev 33: 643–656. [DOI] [PubMed] [Google Scholar]
- 12. Havarstein LS, Gaustad P, Nes IF, Morrison DA (1996) Identification of the streptococcal competence-pheromone receptor. Mol Microbiol 21: 863–869. [DOI] [PubMed] [Google Scholar]
- 13. Pestova EV, Havarstein LS, Morrison DA (1996) Regulation of competence for genetic transformation in Streptococcus pneumoniae by an auto-induced peptide pheromone and a two-component regulatory system. Mol Microbiol 21: 853–862. [DOI] [PubMed] [Google Scholar]
- 14. Havarstein LS (2010) Increasing competence in the genus Streptococcus. Mol Microbiol 78: 541–544. [DOI] [PubMed] [Google Scholar]
- 15. Martin B, Granadel C, Campo N, Henard V, Prudhomme M, et al. (2010) Expression and maintenance of ComD-ComE, the two-component signal-transduction system that controls competence of Streptococcus pneumoniae. Mol Microbiol 75: 1513–1528. [DOI] [PubMed] [Google Scholar]
- 16. Luo P, Li HY, Morrison DA (2003) ComX is a unique link between multiple quorum sensing outputs and competence in Streptococcus pneumoniae. Mol Microbiol 50: 623–633. [DOI] [PubMed] [Google Scholar]
- 17. Lau GW, Haataja S, Lonetto M, Kensit SE, Marra A, et al. (2001) A functional genomic analysis of type 3 Streptococcus pneumoniae virulence. Mol Microbiol 40: 555–571. [DOI] [PubMed] [Google Scholar]
- 18. Bartilson M, Marra A, Christine J, Asundi JS, Schneider WP, et al. (2001) Differential fluorescence induction reveals Streptococcus pneumoniae loci regulated by competence stimulatory peptide. Mol Microbiol 39: 126–135. [DOI] [PubMed] [Google Scholar]
- 19. Hava DL, Camilli A (2002) Large-scale identification of serotype 4 Streptococcus pneumoniae virulence factors. Mol Microbiol 45: 1389–1406. [PMC free article] [PubMed] [Google Scholar]
- 20. Guiral S, Mitchell TJ, Martin B, Claverys JP (2005) Competence-programmed predation of noncompetent cells in the human pathogen Streptococcus pneumoniae: genetic requirements. Proc Natl Acad Sci U S A 102: 8710–8715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Claverys JP, Martin B, Havarstein LS (2007) Competence-induced fratricide in streptococci. Mol Microbiol 64: 1423–1433. [DOI] [PubMed] [Google Scholar]
- 22. Kowalko JE, Sebert ME (2008) The Streptococcus pneumoniae competence regulatory system influences respiratory tract colonization. Infect Immun 76: 3131–3140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Pozzi G, Masala L, Iannelli F, Manganelli R, Havarstein LS, et al. (1996) Competence for genetic transformation in encapsulated strains of Streptococcus pneumoniae: two allelic variants of the peptide pheromone. J Bacteriol 178: 6087–6090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Whatmore AM, Barcus VA, Dowson CG (1999) Genetic diversity of the streptococcal competence (com) gene locus. J Bacteriol 181: 3144–3154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Iannelli F, Oggioni MR, Pozzi G (2005) Sensor domain of histidine kinase ComD confers competence pherotype specificity in Streptoccoccus pneumoniae. FEMS Microbiol Lett 252: 321–326. [DOI] [PubMed] [Google Scholar]
- 26. Johnsborg O, Kristiansen PE, Blomqvist T, Havarstein LS (2006) A hydrophobic patch in the competence-stimulating Peptide, a pneumococcal competence pheromone, is essential for specificity and biological activity. J Bacteriol 188: 1744–1749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Zhu L, Lau GW (2011) Inhibition of competence development, horizontal gene transfer and virulence in Streptococcus pneumoniae by a modified competence stimulating peptide. PLoS Pathog 7: e1002241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Coomaraswamy G (1996) Induction of genetic transformation in Streptococcus pneumoniae by a pheromone peptide and its synthetic analogs. Ph.D. thesis. University of Illinois, Chicago.
- 29. Sung CK, Morrison DA (2005) Two distinct functions of ComW in stabilization and activation of the alternative sigma factor ComX in Streptococcus pneumoniae. J Bacteriol 187: 3052–3061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Johnsborg O, Havarstein LS (2009) Regulation of natural genetic transformation and acquisition of transforming DNA in Streptococcus pneumoniae. FEMS Microbiol Rev 33: 627–642. [DOI] [PubMed] [Google Scholar]
- 31. Mashburn-Warren L, Morrison DA, Federle MJ (2010) A novel double-tryptophan peptide pheromone controls competence in Streptococcus spp. via an Rgg regulator. Mol Microbiol 78: 589–606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Pozzi G, Iannelli F, Oggioni MR, Santagati M, Stefani S (2004) Genetic elements carrying macrolide efflux genes in streptococci. Curr Drug Targets Infect Disord 4: 203–206. [DOI] [PubMed] [Google Scholar]
- 33. Shen X, Yang H, Yu S, Yao K, Wang Y, et al. (2008) Macrolide-resistance mechanisms in Streptococcus pneumoniae isolates from Chinese children in association with genes of tetM and integrase of conjugative transposons 1545. Microb Drug Resist 14: 155–161. [DOI] [PubMed] [Google Scholar]
- 34. Widdowson CA, Klugman KP (1998) The molecular mechanisms of tetracycline resistance in the pneumococcus. Microb Drug Resist 4: 79–84. [DOI] [PubMed] [Google Scholar]
- 35. Dorer MS, Fero J, Salama NR (2010) DNA damage triggers genetic exchange in Helicobacter pylori. PLoS Pathog 6: e1001026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Maiques E, Ubeda C, Campoy S, Salvador N, Lasa I, et al. (2006) beta-lactam antibiotics induce the SOS response and horizontal transfer of virulence factors in Staphylococcus aureus. J Bacteriol 188: 2726–2729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Prudhomme M, Attaiech L, Sanchez G, Martin B, Claverys JP (2006) Antibiotic stress induces genetic transformability in the human pathogen Streptococcus pneumoniae. Science 313: 89–92. [DOI] [PubMed] [Google Scholar]
- 38. Havarstein LS, Hakenbeck R, Gaustad P (1997) Natural competence in the genus Streptococcus: evidence that streptococci can change pherotype by interspecies recombinational exchanges. J Bacteriol 179: 6589–6594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Kilian M, Poulsen K, Blomqvist T, Havarstein LS, Bek-Thomsen M, et al. (2008) Evolution of Streptococcus pneumoniae and its close commensal relatives. PLoS One 3: e2683. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Avery OT, MacLeod CM, McCarty M (1979) Studies on the chemical nature of the substance inducing transformation of pneumococcal types. Inductions of transformation by a desoxyribonucleic acid fraction isolated from pneumococcus type III. J Exp Med 149: 297–326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Morrison DA, Baker MF (1979) Competence for genetic transformation in pneumococcus depends on synthesis of a small set of proteins. Nature 282: 215–217. [DOI] [PubMed] [Google Scholar]