

Abstract
The disaccharide trehalose and trehalose-6-phosphate that are present in trace amounts are suggested to have a signaling function in plants. Recently, it was demonstrated that trehalose metabolism contributes to Arabidopsis thaliana defense against the green peach aphid (GPA; Myzus persicae Sülzer), an important insect pest of a large variety of plants. TPS11 (TREHALOSE PHOSPHATE SYNTHASE11)-dependent trehalose metabolism was shown to curtail GPA infestation by promoting starch accumulation and expression of the PAD4 (PHYTOALEXIN-DEFICIENT4) gene, which has important roles in regulating antibiosis and antixenosis against GPA. Here we show that trehalose metabolism is similarly activated in leaves of GPA-infested tomato (Solanum lycopersicum) plants and likely contributes to tomato defense against GPA. GPA-infested leaves of tomato accumulated trehalose, which was accompanied by the transient upregulation of SlTPS11, a homolog of the Arabidopsis TPS11. GPA-infestation was also accompanied by starch accumulation and the upregulation of SlPAD4, the tomato homolog of Arabidopsis PAD4. Furthermore, trehalose application induced SlPAD4 expression and starch accumulation, and curtailed GPA infestation, suggesting that like in Arabidopsis trehalose contributes to tomato defense against GPA.
Keywords: Myzus persicae, SlPAD4, SlTPS11, Solanum lycopersicum, plant defense, starch, trehalose
Sugars have important functions in carbon and energy metabolism of plants.1 In addition, sugars also have critical roles as signaling molecules in plant growth, development and response to environmental stress.1-3 The role of sucrose as a signaling molecule has been well documented for some time.1 In recent years, the α,α-1,1-linked glucose disaccharide trehalose and its metabolic intermediate trehalose-6- phosphate (T6P), which are present in trace amounts in most plants, were also shown to function as signaling molecules in plant growth, development and stress response.4,5 We recently demonstrated the involvement of trehalose metabolism in Arabidopsis thaliana defense against the green peach aphid (Myzus persicae Sülzer),6 which is an important pest of more than 50 families of plants.7 Trehalose levels and expression of the TREHALOSE PHOSPHATE SYNTHASE 11 (TPS11) gene, which encodes a trehalose synthesizing enzyme, transiently increased in GPA-infested Arabidopsis leaves.6 In addition to trehalose, sucrose levels also increased in GPA-infested Arabidopsis leaves.6 The upregulation of TPS11 promoted the flux of C into starch at the expense of sucrose accumulation. Starch accumulation in GPA-infested leaves was required for the full extent of basal resistance against GPA.6 TPS11 promoted the timely upregulation of PAD4 (PHYTOALEXIN-DEFICIENT4),6 which is an important modulator of antibiotic and antixenotic defenses against GPA. Here we present evidence that like in Arabidopsis, tomato leaves also respond to GPA-infestation with the activation of trehalose metabolism.
Trehalose Content Transiently Increases in GPA-infested Tomato Leaves
Tomato is a host for GPA. To determine if tomato, like Arabidopsis responds to GPA infestation with an increase in trehalose content, GPA was released on tomato variety Motelle plants and leaves from GPA-infested and un-infested plants harvested over a 24 h period. Trehalose content in these leaves was monitored as previously described.6 As shown in Figure 1A, although trehalose levels were undetectable in leaves of un-infested plants, trehalose content gradually increased in the GPA-infested leaves, reaching peak levels by 12 hpi and then declining at 24 hpi.
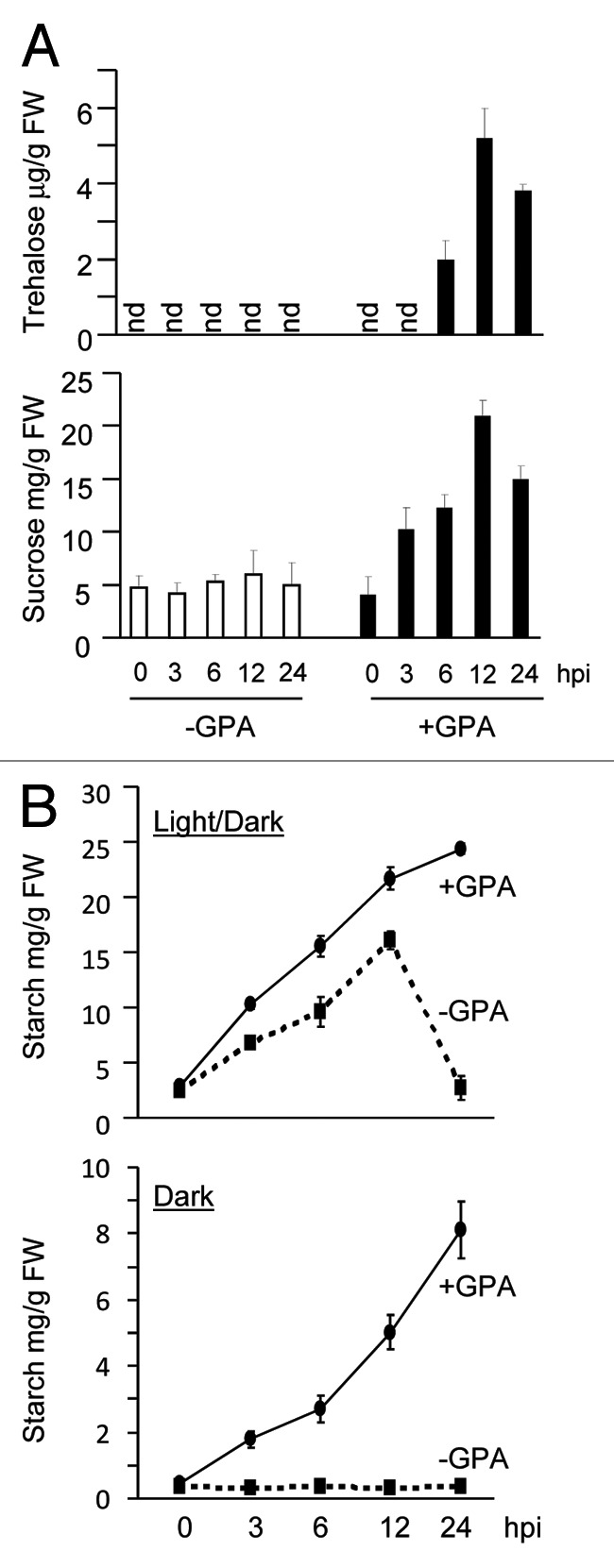
Figure 1. Trehalose, sucrose and starch content in GPA-infested tomato. (A) Trehalose and sucrose levels were measured at the indicated hours post infestation (hpi) in un-infested (-GPA) and GPA-infested (+GPA) leaves of the tomato variety Motelle (n = 3). (B) Starch content was measured in un-infested (-GPA) and GPA-infested (+GPA) tomato leaves of Motelle kept for 24 h in a 14/10 h light/dark cycle (Top panel) or for 24 h in complete darkness (Lower panel) (n = 3). For the light/dark cycle, insects were released on plants (0 h) immediately after start of the light period (8:00AM). In A and B, each plant received 25 adult aphids. Error bars represent SE. nd, non-detectable.
Sucrose and Starch Content Increase in GPA-infested Tomato Leaves
In Arabidopsis, GPA infestation results in an increase in sucrose and starch content.6 TPS11 promoted the accumulation of starch at the expense of sucrose in GPA-infested Arabidopsis leaves. No-choice assays with the starch-deficient Arabidopsis pgm1 mutant indicated that starch accumulation was required for controlling GPA infestation in Arabidopsis. Hence, it was suggested that starch accumulation is a mechanism that is utilized by the host plant to generate a secondary sink that counter’s the ability of the insect to manipulate host metabolism to increase sucrose content.6 We therefore examined if GPA-infestation also results in starch accumulation in tomato leaves. As shown in Figure 1B, a rapid increase in starch content was observed in the GPA-infested leaves of tomato. Starch accumulation paralleled sucrose accumulation in these GPA-infested leaves (Fig. 1A). Starch accumulation was also observed in experiments where the GPA-infested tomato plants were kept in darkness for the duration of the experiment (Fig. 1B). These results confirm that like Arabidopsis, tomato also responds to GPA-infestation by promoting sucrose and starch accumulation.
Trehalose Promotes Resistance Against GPA in Tomato Leaves
Trehalose applied to tomato leaves promoted starch accumulation (Fig. 2A) and curtailed GPA infestation (Fig. 2B), suggesting that similar to Arabidopsis trehalose metabolism may have a role in tomato defense against GPA. In Arabidopsis, expression of TPS11, which is required for trehalose accumulation, parallels the transient increase in trehalose in GPA-infested leaves.6 A survey of the nucleotide sequence databases revealed that the tomato genome contains a TPS11 homolog, which we have designated SlTPS11 (GenBank: AB368491). SlTPS11 expression was transiently induced in the GPA-infested leaves of tomato (Fig. 3A). The time-frame of SlTPS11 upregulation parallels the time-frame of trehalose accumulation in GPA-infested tomato leaves (Fig. 1A), suggesting that SlTPS11 expression is likely associated with the increase in trehalose in GPA-infested tomato leaves. By comparison to the transient upregulation of SlTPS11, expression of tomato SlPAD4 (GenBank: AI781175), which is homologous to PAD4 that is involved in Arabidopsis defense against GPA,8-10 gradually increased over the duration of the experiment (Fig. 3A). Similar to the Arabidopsis PAD4, SlPAD4 expression in tomato was upregulated in response to trehalose application (Fig. 3B). These results suggest that the expression of SlPAD4 in response to GPA infestation is similar to the expression of PAD4 in GPA-infested Arabidopsis. Furthermore, similar to our observations in Arabidopsis,6 GPA population was smaller on tomato plants that were pre-treated with trehalose as opposed to the water treated control plants (Fig. 2B).
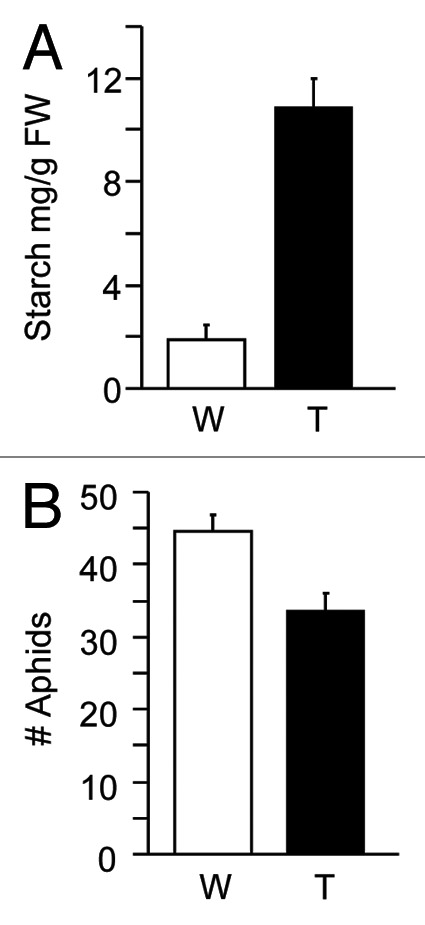
Figure 2. Trehalose application promotes tomato defense against GPA. (A) Starch content in trehalose-treated tomato leaves. Starch content was measured in leaves of tomato variety Motelle 12 h after infiltration of 10 µM Trehalose (T) and as control with water (W). (B) Trehalose application enhances resistance against GPA. No-choice assay: GPA numbers (adults + nymphs) on plants of the tomato variety Motelle infiltrated with 10 µM Trehalose (T) and as control with water (W), 12 h prior to release of 25 adult aphids per plant. Aphid numbers were taken at 2 dpi (n = 5). In A and B, error bars represent SE.
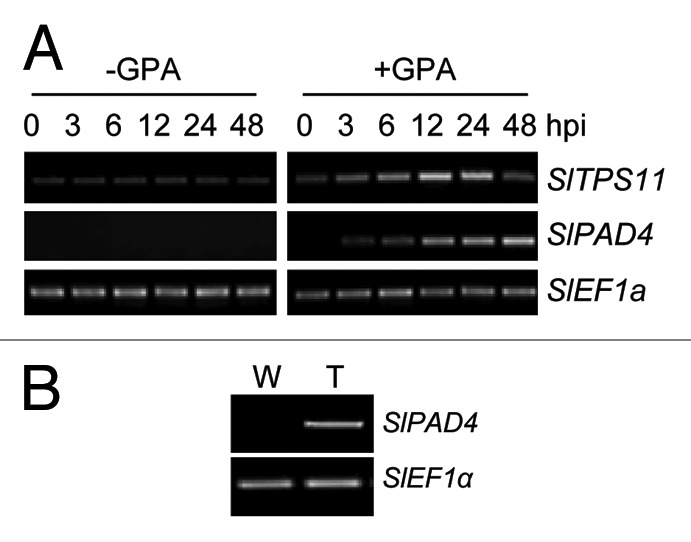
Figure 3. Expression of defense-related genes in tomato leaves. (A) RT-PCR analysis of tomato SlTPS11 (GenBank: AB368491) and SlPAD4 (GenBank: AI781175) gene expression in un-infested (-GPA) and GPA-infested (+GPA) leaves of tomato variety Motelle at the indicated hours post infestation (hpi). (B) RT-PCR analysis of tomato SlPAD4 expression in Motelle leaves 12 h post infiltration with a 10 µM solution of trehalose (T). Water (W) treated leaves provided negative controls. In A and B, expression of the SlEF1a gene (GenBank: X53043.1) provided the control for RT-PCR.
Taken together, the results presented here indicate similarities in the molecular and physiological alterations in tomato and Arabidopsis leaves in response to GPA-infestation. We suggest that similar to Arabidopsis, tomato engages trehalose metabolism to promote the activation of defenses that target GPA. This study also bolsters the idea that plants have evolved mechanisms that invoke their primary metabolic pathways to counter insect herbivory. Additional genetic studies are required to validate the contribution of trehalose and starch metabolism, and the SlTPS11 and SlPAD4 genes in tomato defense against GPA.
Note
GenBank IDs: SlPAD4 (AI781175); SlTPS11 (AB368491); SlEF1a (X53043.1)
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Footnotes
Previously published online: www.landesbioscience.com/journals/psb/article/20066
References
- 1.Rolland F, Baena-Gonzalez E, Sheen J. Sugar sensing and signaling in plants: conserved and novel mechanisms. Annu Rev Plant Biol. 2006;57:675–709. doi: 10.1146/annurev.arplant.57.032905.105441. [DOI] [PubMed] [Google Scholar]
- 2.Koch KE. Carbohydrate modulated gene expression in plants. Annu Rev Plant Physiol Plant Mol Biol. 1996;47:509–40. doi: 10.1146/annurev.arplant.47.1.509. [DOI] [PubMed] [Google Scholar]
- 3.Roitsch T. Source-sink regulation by sugar and stress. Curr Opin Plant Biol. 1999;2:198–206. doi: 10.1016/S1369-5266(99)80036-3. [DOI] [PubMed] [Google Scholar]
- 4.Paul MJ, Primavesi LF, Jhurreea D, Zhang Y. Trehalose metabolism and signaling. Annu Rev Plant Biol. 2008;59:417–41. doi: 10.1146/annurev.arplant.59.032607.092945. [DOI] [PubMed] [Google Scholar]
- 5.Fernandez O, Béthencourt L, Quero A, Sangwan RS, Clément C. Trehalose and plant stress responses: friend or foe? Trends Plant Sci. 2010;15:409–17. doi: 10.1016/j.tplants.2010.04.004. [DOI] [PubMed] [Google Scholar]
- 6.Singh V, Louis J, Ayre BG, Reese JC, Pegadaraju V, Shah J. TREHALOSE PHOSPHATE SYNTHASE11-dependent trehalose metabolism promotes Arabidopsis thaliana defense against the phloem-feeding insect Myzus persicae. Plant J. 2011;67:94–104. doi: 10.1111/j.1365-313X.2011.04583.x. [DOI] [PubMed] [Google Scholar]
- 7.Blackman RL, Eastop VF. Aphids on the World’s Crops: An Identification and Information Guide, 2nd edn, 2002; Chichester, UK: John Wiley. [Google Scholar]
- 8.Pegadaraju V, Knepper C, Reese J, Shah J. Premature leaf senescence modulated by the Arabidopsis PHYTOALEXIN DEFICIENT4 gene is associated with defense against the phloem-feeding green peach aphid. Plant Physiol. 2005;139:1927–34. doi: 10.1104/pp.105.070433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Pegadaraju V, Louis J, Singh V, Reese JC, Bautor J, Feys BJ, et al. Phloem-based resistance to green peach aphid is controlled by Arabidopsis PHYTOALEXIN DEFICIENT4 without its signaling partner ENHANCED DISEASE SUSCEPTIBILITY1. Plant J. 2007;52:332–41. doi: 10.1111/j.1365-313X.2007.03241.x. [DOI] [PubMed] [Google Scholar]
- 10.Louis J, Lorenc-Kukula K, Singh V, Reese J, Jander G, Shah J. Antibiosis against the green peach aphid requires the Arabidopsis thaliana MYZUS PERSICAE-INDUCED LIPASE1 gene. Plant J. 2010;64:800–11. doi: 10.1111/j.1365-313X.2010.04378.x. [DOI] [PubMed] [Google Scholar]