

Graphical abstract

Keywords: Endothelial nitric oxide synthase (eNOS), Ca2+, Benzofuran derivative, AMPK, CaMKKβ
Abstract
Endothelial nitric oxide synthase (eNOS) mediates important vaso-protective and immunomodulatory effects. Aim of this study was to examine whether lignan derivatives isolated from the roots of the anti-inflammatory medicinal plant Krameria lappacea influence eNOS activity and endothelial nitric oxide (NO) release. The study was performed using cultured human umbilical vein endothelial cells (HUVECs) and HUVEC-derived EA.hy926 cells. Among the eleven isolated compounds only 2-(2,4-dihydroxyphenyl)-5-(E)-propenylbenzofuran (DPPB) was able to increase eNOS enzyme activity.
DPPB (1–10 μM) treatment for 24 h induced a significant and dose-dependent increase in eNOS activity as determined by the [14C]l-arginine/[14C]l-citrulline conversion assay. Immunoblotting studies further revealed a time-dependent DPPB-induced increase in eNOS-Ser1177 and decrease in eNOS-Thr495 phosphorylation, as well as increased AMPK phosphorylation at Thr172, whereas Akt phosphorylation at Ser473 was not affected. Si-RNA-mediated knockdown of AMPK and inhibition of CaMKKβ by STO 609, as well as intracellular Ca2+ chelation by Bapta AM abolished the stimulating effect of DPPB on eNOS-Ser1177 and AMPK-Thr172 phosphorylation. Furthermore, we could show that DPPB increases intracellular Ca2+ concentrations assessed with the fluorescent dye Fluo-3-AM. DPPB enhances eNOS activity and endothelial NO release by raising intracellular Ca2+ levels and increases signaling through a CaMKKβ–AMPK dependent pathway.
1. Introduction
The endothelium is responsible for the maintenance of vascular homeostasis thereby regulating vascular tone, thrombosis, and smooth muscle cell proliferation and migration [1,2]. In a diseased state this balance is disturbed and the endothelium is in a dysfunctional state. Endothelial dysfunction is a key event in the development of atherosclerosis and can therefore be regarded as an early marker for cardiovascular disease [3]. Hallmark of a dysfunctional endothelium is an impaired action of the enzyme endothelial nitric oxide synthase (eNOS) [4]. The reaction catalyzed by eNOS leads to the conversion of molecular oxygen and l-arginine to l-citrulline and nitric oxide (NO). NO is a major vasoprotective mediator released by the endothelium, with multiple effects including next to vasodilatation also inhibition of platelet aggregation and leukocyte adhesion as well as control of smooth muscle cell proliferation. This overall leads to an anti-inflammatory and immunomodulatory action of NO [5–8].
Besides different cofactors (e.g. NADPH, FMN, FAD, BH4), adaptor and regulatory proteins, eNOS requires Ca2+ for sustained action. When the intracellular Ca2+ concentration ([Ca2+]i) reaches a certain level, eNOS binds to the regulatory protein Ca2+/calmodulin (CaM), thereby leading to an increased rate of electron transfer from NADPH bound at the reductase domain of the enzyme to the heme center at the N-terminal oxygenase domain, activating the enzyme [9]. Extracellular ligands of G-protein coupled receptors, such as acetylcholine, bradykinin, adenosine, thrombin or histamine, rapidly increase [Ca2+]i and activate eNOS via this mechanism [10–12]. ENOS activation, however, can also be induced by stimuli that act independently of a [Ca2+]i increase like shear stress induced by blood flow [13]. Regulation of eNOS further involves modulation of eNOS protein expression and post-translational modification of the protein such as phosphorylation [14]. Among the most important kinases directly activating eNOS by phosphorylation are Akt and AMP-activated protein kinase (AMPK) [15].
There is an increasing body of evidence that some natural products contained in dietary sources or in specific medicinal plants may positively influence endothelial NO production and thus promote endothelial function [14]. The identification of such compounds and better understanding of their molecular mechanism of action provides valuable input for both the prevention and the treatment of cardiovascular disease. Plant secondary metabolites from the benzofuran class are known to exhibit a broad range of bioactivities, including anti-inflammatory [16,17], cardio- and vaso-protective [18,19], cytostatic [20], and antioxidant actions [21,22]. However, an influence on eNOS activity was not described so far.
The roots of Krameria lappacea (Dombey) Burdet et Simpson (syn. K. triandra Ruiz et Pavon), which are a rich source for such compounds, have been traditionally used in South America for the treatment of different inflammation-related complaints, and have been introduced in the European medicine since the 18th century [23–25]. We have isolated eleven lignans (nine benzofuran lignans and two epoxy lignans) including the benzofuran derivative 2-(2,4-dihydroxyphenyl)-5-(E)-propenylbenzofuran (DPPB) from the dried roots of K. lappacea. So far only one report shows the identification of DPPB as the major active compound of a standardized K. triandra root extract exhibiting a potent cytoprotective effect on different cell lines exposed to stress [23]. We examine here a potential positive effect of the isolated lignan derivatives on eNOS activity in endothelial cells. Furthermore, we investigate the molecular mode of action of the most active compound DPPB and characterize the upstream signaling pathways that lead to increased eNOS activity.
2. Methods
2.1. Chemicals and cell culture reagents
Dulbecco's modified Eagle's medium (DMEM) without phenol red containing 4.5 g/L glucose, endothelial growth medium EBM™, EGM™ SingleQuots, glutamine, amphotericin B, benzylpenicillin and streptomycin were purchased from Lonza (Verviers, Belgium), HAT supplement (100 μM hypoxanthine, 0.4 μM aminopterin and 16 μM thymidine) from Biochrom (Berlin, Germany), and trypsin from Cambrex (Verviers, Belgium). Fetal bovine serum (FBS) was obtained from Gibco via Invitrogen (Paisley, UK). A23187 and 4,5-diaminofluorescein (DAF-2) were bought from Alexis Biochemicals (Lausen, Switzerland) and [14C]l-arginine (346 mCi/mmol) from New England Nuclear (Bosten, MA, USA). Bapta AM was purchased from Tocris (Bristol, UK) and Fluo-3-AM was bought from Invitrogen (Paisley, UK). Antibodies were obtained from the following companies: eNOS from BD (Becton, Dickinson and Company; Franklin Lakes, NJ, USA), phospho-eNOS-Ser1177, phospho-eNOS-Thr495, phospho-AMPK-Thr172, AMPK, phospho-Akt-Ser473, Akt, and horseradish peroxidase-conjugated goat anti-rabbit secondary antibody from Cell Signaling (Bosten, MA, USA), anti-tubulin from Santa Cruz (CA, USA), anti-actin from MP Biomedicals (Solon, OH, USA), and horseradish peroxidase-conjugated goat anti-mouse secondary antibody from Upstate (Millipore, Vienna, Austria). All other chemicals were bought from Sigma–Aldrich (Vienna, Austria). TLC plates were bought from Machery-Nagel (Markus Bruckner Analysentechnik, Linz, Austria).
2.2. Origin of tested compounds
Compounds 1–11 were isolated and identified in a previous study [26] as: 5-(3-hydroxypropyl)-2-(2-methoxy-4-hydroxyphenyl)benzofuran (1), (−)-larreatricin (2), meso-3,30-didemethoxynectandrin B (3), (2S,3S)-2,3-dihydro-3-hydroxymethyl-2-(4-hydroxyphenyl)-5-(E)-propenylbenzofuran (4), 2-(2-hydroxy-4-methoxyphenyl)-5-(3-hydroxypropyl)benzofuran (5), 2-(2,4-dihydroxyphenyl)-5-(E)-propenylbenzofuran (DPPB; 6), (+)-conocarpan (7), 2-(4-hydroxyphenyl)-5-(E)-propenylbenzofuran (8), rataniaphenol III (9), rataniaphenol I (10), and rataniaphenol II (11). The purity of all isolated compounds was ≥96% (determined by HPLC). The used plant material (Ratanhiae radix, dried ground roots of Krameria lappacea (Dombey) Burdet et Simpson (syn. K. triandra Ruiz et Pavon; Krameriaceae); 500 g; KL 6269) was purchased from Mag. pharm. Kottas-Heldenberg & Sohn (Vienna, Austria) and complied with the European Pharmacopoeia. A voucher specimen (KL 6269) is deposited at the Institute of Pharmacy/Pharmacognosy, University of Innsbruck (Austria).
2.3. Cell culture
The human endothelial cell line EA.hy926 (kindly provided by Dr. C.-J.S. Edgell, University of North Carolina, Chapel Hill, NC, USA) [28] was grown in DMEM without phenol red supplemented with 2 mM glutamine, 100 U/mL benzylpenicillin, 100 μg/mL streptomycin, HAT supplement, and 10% FBS until passage 26. For experiments, cells were seeded in six-well plates at a density of 5 × 105 cells/well and treated with test compounds at confluence, after approximately 72 h. HUVECs were obtained from Lonza and cultivated in EBM™ growth medium supplemented with 10% FBS, EGM™ SingleQuots, 100 U/mL benzylpenicillin, 100 μg/mL streptomycin, and 1% amphotericin until passage five. For experiments cells were seeded in gelatine-coated six-well plates at a density of 1 × 105 cells/well. DPPB and other tested compounds of K. lappacea, STO 609, compound C, and Bapta AM were dissolved in dimethyl sulfoxide (DMSO) and stored at −80 °C. Final DMSO concentrations did not exceed 0.1%. Control cells were always treated with an equal volume of solvent.
2.4. [14C]l-arginine/[14C]l-citrulline conversion assay
The enzymatic reaction catalyzed by eNOS converts the amino acid arginine into citrulline and NO. [14C]l-citrulline production can thus serve as a surrogate marker of NO production. The assay was performed as previously described [29]. Briefly, endothelial cells were equilibrated in HEPES buffer (HEPES 10 mM, NaCl 145 mM, KCl 5 mM, MgSO4 2 mM, α-d(+)-Glucose 10 mM, CaCl2 × 2H2O 1.5 mM, pH 7.4) for 10 min at 37 °C. Then 0.32 μM [14C]l-arginine (346 mCi/mmol) and 1 μM of the calcium ionophore A23187 were added. The reaction was stopped by lysing cells followed by extraction with ethanol and ethanol/water. The extracts were dried under vacuum (SPD 1010 SpeedVac, Thermo Savant, Thermo Scientific, Langenselbold, Germany) and resolved in water/methanol (1:1). After separation of [14C]l-arginine from [14C]l-citrulline by thin layer chromatography (Polygram SIL N-HR, Machery-Nagel, Austria) in the solvent system water:chloroform:methanol:ammonium hydroxide 25% (2:1:9:4, v/v/v/v) [14C]l-citrulline was quantified by autoradiography in a phosphoimager (BAS-1800II, Fujifilm, Düsseldorf, Germany). AIDA software (raytest, Langenzersdorf, Austria) was used for densitometric analysis.
2.5. Quantification of NO release by 4,5-diaminofluorescein-2 (DAF-2)
Quantification of NO released from endothelial cells was performed using 4,5-diaminofluorescein-2 (DAF-2), a NO-sensitive fluorescent probe. EA.hy926 cells were seeded in 96-well plates at a density of 2.5 × 104 cells/well and were treated with test compounds at confluence after approximately 72 h. Cells were washed two times with PBS+ (137 mM NaCl, 2.68 mM KCl, 8.1 mM Na2HPO4, 1.47 mM KH2PO4, 0.5 mM MgCl2 × 6H2O, 0.68 mM CaCl2 × 2H2O) containing 100 μM arginine and equilibrated 10 min in this buffer. Then A23187 was added to a final concentration of 1 μM and DAF-2 to a final concentration of 0.1 μM and the cells were incubated for 1 h at 37 °C. Addition of l-NAME to a final concentration of 200 μM allowed correction for non-NO-specific fluorescence. The supernatant was transferred to a black 96-well plate and fluorescence was measured in a plate reader (Genios Pro, Tecan, Grödig, Austria) with an excitation wavelength of 485 nm and an emission wavelength of 520 nm. Fluorescence values were normalized to viable cells as determined by resazurin conversion method [30]. For this, the cells were incubated with 0.1 mg/mL resazurin in PBS for 30 min before measuring the fluorescence in a plate reader (Genios Pro, Tecan, Grödig, Austria) at an excitation wavelength of 535 nm and an emission wavelength of 590 nm.
2.6. Gel electrophoresis and immunoblot analysis
Preparation of cell extracts, SDS-PAGE, immunoblot analysis, and densitometric evaluations were performed as described previously [31]. For detection of multiple proteins with similar molecular weights in one sample, two or more identical membranes were processed in parallel.
2.7. siRNA mediated knockdown of AMPKα
HUVECs were seeded in six-well plates at a density of 0.3 × 106 cells/well and transfected one day later with 33 pmol AMPKα siRNA (Santa Cruz, CA, USA) and scrambled control (Invitrogen, Paisley, UK), respectively, using the OptiMEM/Oligofectamine system (Invitrogen, Paisley, UK). 72 h after transfection the cells were used for experiments. Successful knockdown of the target proteins was confirmed by Western blot analysis.
2.8. Determination of intracellular Ca2+ concentration
The intracellular calcium concentration ([Ca2+]i) was measured using the intracellular calcium indicator Fluo-3-AM. EA.hy926 cells were seeded at a density of 0.5 × 106 cells/well in a 6-well plate and treated with test compounds at confluence, approximately after 72 h. Cells were loaded with the fluorescent dye Fluo-3-AM (2 μM) for 45 min at 37 °C. Then [Ca2+]i was measured in a flow cytometer.
2.9. Statistical methods
Statistical analysis was done using GraphPad Prism software version 4.03 (GraphPad Software Inc., La Jolla, CA, USA). To determine statistical significance Student's t test or one- or two-way analysis of variance (ANOVA) were performed. P values < 0.05 were considered significant (*). Figures with bar graphs represent means ± SEM of at least three independent experiments.
3. Results
3.1. DPPB is the only compound isolated from K. lappacea that stimulates endothelial NO release and increases eNOS enzyme activity
Eleven lignan derivatives (Fig. 1) were isolated from the dried roots of K. lappacea [26] and investigated for their ability to alter NO release from endothelial cells. Treatment of endothelial EA.hy926 cells [28] with compounds 1, 5, and 9 showed a significantly decreased release in endothelial NO whereas only compound 6 increased it (Fig. 2A). The other compounds had no significant effect. We also determined possible toxic effects of the isolated compound on endothelial EA.hy926 cells by a LDH release and resazurin conversion assay. None of the compounds were toxic (data not shown). Since eNOS activity is the major determinant of the endothelial NO production, we further determined eNOS enzyme activity by quantifying the conversion of [14C]l-arginine to [14C]l-citrulline. We tested compounds 6, 7, 8 and 11 in this assay, as they were the most promising compounds regarding the results from the endothelial NO release measurement, although only compound 6 showed a statistically significant effect (Fig. 2B). Interestingly only compound 6 (DPPB, 2-(2,4-dihydroxyphenyl)-5-(E)-propenylbenzofuran) was able to significantly increase eNOS enzyme activity. Therefore DPPB was further investigated. Treatment of endothelial EA.hy926 cells with 10 μM DPPB for 24 h resulted in a more than 2-fold increase in NO availability in comparison to the solvent control (Fig. 2C). Mevastatin, which is known to potently upregulate eNOS expression and thereby NO release, was used as a positive control [32,33]. Additionally, DPPB treatment resulted in a dose-dependent increase of eNOS activity in EA.hy926 cells, reaching 1.2-fold activation at a concentration of 10 μM (Fig. 2D). As a positive control for the [14C]l-arginine/[14C]l-citrulline conversion assay we used ascorbic acid, which is known to stabilize the eNOS cofactor tetrahydrobiopterin and therefore increases eNOS activity [34,35]. To verify the activity of DPPB also in primary HUVECs, cells were treated with 10 μM DPPB and eNOS enzyme activity was measured using the [14C]l-arginine/[14C]l-citrulline conversion assay, resulting in a very similar activity of DPPB (Fig. 2E).
Fig. 1.

Chemical structures of investigated lignan derivatives of K. lappacea: 5-(3-hydroxypropyl)-2-(2-methoxy-4-hydroxyphenyl)benzofuran (1), (−)-larreatricin (2), meso-3,30-didemethoxynectandrin B (3), (2S,3S)-2,3-dihydro-3-hydroxymethyl-2-(4-hydroxyphenyl)-5-(E)-propenylbenzofuran (4), 2-(2-hydroxy-4-methoxyphenyl)-5-(3-hydroxypropyl)benzofuran (5), 2-(2,4-dihydroxyphenyl)-5-(E)-propenylbenzofuran (DPPB; 6), (+)-conocarpan (7), 2-(4-hydroxyphenyl)-5-(E)-propenylbenzofuran (8), rataniaphenol III (9), rataniaphenol I (10), and rataniaphenol II (11).
Fig. 2.

DPPB increases eNOS activity and endothelial NO release. (A) EA.hy926 cells were treated with 10 μM of the indicated compound or solvent vehicle for 24 h. Endothelial NO release of eleven lignan derivatives was measured by incubation with the NO-sensitive fluorescent probe diaminofluorescein-2. Fluorescence values were normalized to the number of viable cells as determined by resazurin staining (*p < 0.05; **p < 0.01) (mean ± SEM, n = 3). (B) EA.hy926 cells were treated with 10 μM of the indicated compound or solvent vehicle for 24 h. Then a [14C]l-arginine/[14C]l-citrulline conversion assay was performed as described in Section 2. [14C]l-citrulline production was normalized to the untreated control (**p < 0.01) (mean ± SEM, n = 4). (C) EA.hy926 cells were incubated with 10 μM DPPB for 24 h. Mevastatin (Mev, 1 μM) was used as positive control. NO release was quantified as in (A) (*p < 0.01) (mean ± SEM, n = 3). (D) EA.hy926 cells were treated with the indicated concentration of DPPB for 24 h. Then a [14C]l-arginine/[14C]l-citrulline conversion assay was performed as in (B). Ascorbic acid (Asc, 100 μM) served as a positive control. [14C]l-citrulline production was normalized to the untreated control (**p < 0.01; ***p < 0.001; n.s., not significant) (mean ± SEM, n = 3). (E) HUVECs were incubated with 10 μM of DPPB for 24 h, and eNOS activity was determined as in (B) (**p < 0.01) (mean ± SEM, n = 3).
3.2. DPPB changes the eNOS phosphorylation pattern
Modulation of eNOS activity is linked to changes in the phosphorylation state of the enzyme [15]. Enhanced phosphorylation at eNOS-Ser1177, and concomitantly decreased phosphorylation at eNOS-Thr495, is known to stimulate eNOS activity [15]. Treatment of EA.hy926 cells and HUVECs with DPPB led to an increase in eNOS-Ser1177 phosphorylation and a decrease in eNOS-Thr495 phosphorylation in a time-dependent manner (Fig. 3A and B). DPPB had its maximal effect after 24 h of incubation. Therefore we decided to continue with this condition. On the contrary, total eNOS protein level (Fig. 3A and B) remained unchanged, overall suggesting a direct stimulatory effect of DPPB on eNOS enzyme activity. Additionally we checked for changes in neuronal and inducible NOS expression upon treatment with DPPB, but no differences were detectable (data not shown).
Fig. 3.

Changes in phosphorylation status of different proteins upon DPPB treatment. (A), (C) EA.hy926 cells or (B), (D) HUVECs were treated with 10 μM of DPPB or solvent vehicle for the indicated time points or for 24 h in figure (B) and (D) and subjected to Western blot and subsequent densitometric analysis for the detection and quantification of (phospho-) eNOS levels, (phospho-) Akt levels or (phospho-) AMPK levels, respectively. One representative blot is shown. Band intensities are normalized to tubulin or actin and expressed as fold activation in comparison to the untreated control (***p < 0.001; **p < 0.01; *p < 0.05; n.s., not significant) (mean ± SEM, n = 3).
3.3. DPPB does not affect Akt but increases the phosphorylation of AMPK-Thr172
Among the most prominent upstream determinants of eNOS activity, known to directly catalyze the phosphorylation of eNOS-Ser1177 and thereby stimulate endothelial NO production, are the kinases Akt and AMPK [36,37]. Akt activation was estimated by analyzing the phosphorylation of the Akt-Ser473 residue [37]. The activation of AMPK was monitored by determining the phosphorylation of the AMPK-Thr172 residue [38]. Treatment with DPPB did neither alter Akt phosphorylation in EA.hy926 cells nor in HUVECs (Fig. 3C and D), indicating no changes in Akt activity. However, upon DPPB treatment the phosphorylation of AMPK at Thr172 was increased in both cell types (Fig. 3C and D), indicating an increased AMPK activity.
3.4. Increased eNOS-Ser1177 phosphorylation upon DPPB treatment is dependent on AMPK and CaMKKβ
To check if the AMPK activity is indeed required for the DPPB-induced eNOS-Ser1177 activation we utilized compound C, an inhibitor of AMPK. Application of compound C (10 μM) blocked DPPB-mediated phosphorylation at AMPK-Thr172 as well as at eNOS-Ser1177 in EA.hy926 cells (Fig. 4A), underlining the causal link between the AMPK activation and the observed increase in eNOS phosphorylation. This was further confirmed by downregulation of AMPK levels by transfection with a specific siRNA. When HUVECs were transfected with this siRNA, treatment with DPPB failed to elicit enhanced eNOS-Ser1177 phosphorylation (Fig. 4B). Interestingly treatment of endothelial cells with compound C alone led to a basally elevated eNOS phosphorylation at Ser1177 and AMPK phosphorylation at Thr172. In addition, AMPK siRNA transfection also led to an elevated level of eNOS phosphorylation at Ser1177. This is probably due to off-target effects or compensatory mechanisms triggered by these treatments.
Fig. 4.

eNOS phosphorylation is enhanced after DPPB administration in an AMPK- and CaMKKβ- dependent manner. (A) EA.hy926 cells were pretreated with 10 μM compound C and incubated with 10 μM DPPB for 24 h as indicated. Western blot and subsequent densitometric analyses were performed to detect and quantify (phospho-) AMPK and (phospho-) eNOS protein levels. One representative blot is shown. Band intensities are normalized to tubulin and expressed as fold difference to the vehicle treated control (**p < 0.01; n.s., not significant) (mean ± SEM, n = 3). (B) HUVEC were transfected with AMPKα siRNA or scrambled control prior to treatment with 10 μM DPPB for 1 h as indicated. Western blot and subsequent densitometric analyses were performed to detect and quantify (phospho-) eNOS and AMPK protein. One representative blot is shown. Band intensities are normalized to actin and expressed as fold untreated control (*p < 0.05; n.s., not significant) (mean ± SEM, n = 3). (C) EA.hy926 cells were pretreated with 10 μM compound C and incubated with 10 μM DPPB for 24 h as indicated. Then a [14C]l-arginine/[14C]l-citrulline conversion assay was performed as described in Section 2. [14C]l-citrulline production was normalized to the untreated control (***p < 0.001; n.s., not significant) (mean ± SEM, n = 3). (D) EA.hy926 cells were pretreated with 10 μM STO 609 and incubated with 10 μM DPPB for 24 h as indicated. Western blot and subsequent densitometric analyses were performed to detect and quantify (phospho-) AMPK and (phospho-) eNOS protein levels. One representative blot is shown. Band intensities are normalized to tubulin and expressed as fold untreated control (**p < 0.01; n.s., not significant) (mean ± SEM, n = 3).
To strengthen the connection between AMPK and eNOS activity upon DPPB treatment we further assessed the effect of DPPB on eNOS enzyme activity by measuring conversion of [14C]l-arginine to [14C]l-citrulline in the presence of compound C. Incubation of EA.hy926 cells with 10 μM compound C completely blocked the effect of DPPB on eNOS enzyme activity (Fig. 4C). This result confirms a contribution of AMPK on the DPPB-induced increase in eNOS activity.
To further study how AMPK is activated upon DPPB treatment, we investigated a potential involvement of CaMKKβ, an upstream kinase of AMPK [38,39]. Upon incubation of EA.hy926 cells with 10 μM STO 609, an inhibitor of CaMKKβ, DPPB failed to increase AMPK-Thr172 and eNOS-Ser1177 phosphorylation, suggesting an important role of CaMKKβ for the DPPB-induced activation of eNOS (Fig. 4D).
3.5. DPPB enhances intracellular Ca2+ levels
CaMKKβ and eNOS are both enzymes that are regulated by [Ca2+]i [12,40]. Therefore we were interested whether the increase in eNOS activity and NO production upon treatment with DPPB is due to an increase in [Ca2+]i. For determination of [Ca2+]i we used the fluorescent probe Fluo-3-AM. Incubation of EA.hy926 cells with different concentrations of DPPB showed a dose-dependent increase in [Ca2+]i (Fig. 5A).
Fig. 5.
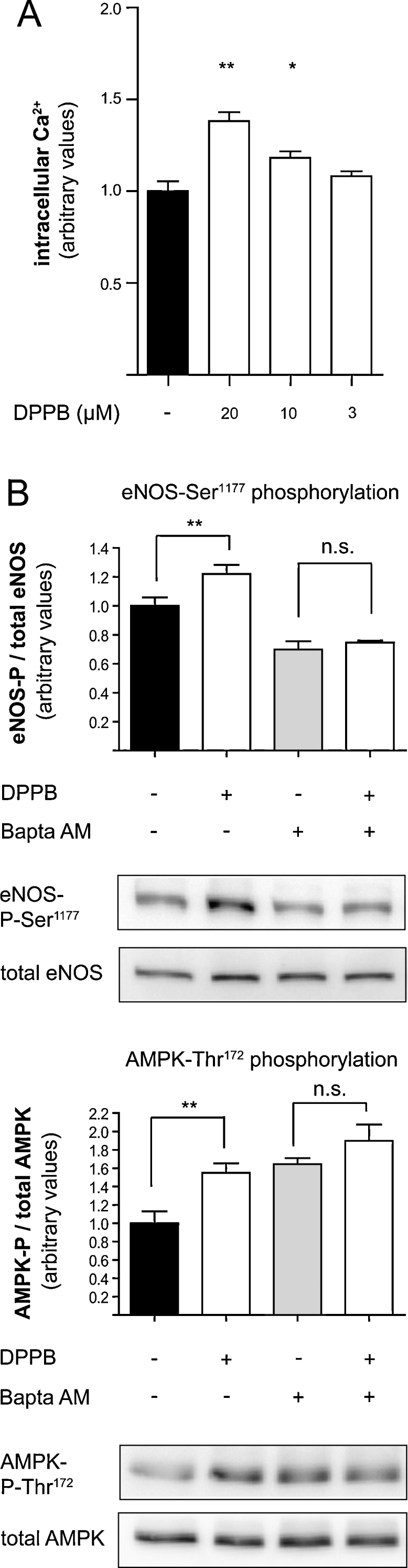
The effect of DPPB is due to increase of intracellular Ca2+ levels. (A) EA.hy926 cells were incubated with the indicated concentrations of DPPB for 24 h. Intracellular Ca2+ was quantified in a flow cytometer after incubation with the fluorescent dye Fluo-3-AM (**p < 0.01; *p < 0.05) (mean ± SEM, n = 3). (B) EA.hy926 cells were treated with 10 μM DPPB for 24 h and with 30 μM Bapta AM for 2 h as indicated. Western blot and subsequent densitometric analyses were performed to detect and quantify (phospho-) eNOS and (phospho-) AMPK protein levels. One representative blot is shown. Band intensities are normalize to actin and expressed as fold untreated control (**p < 0.01, n.s., not significant) (mean ± SEM, n = 6).
3.6. Chelation of intracellular Ca2+ abolishes the positive effect of DPPB on eNOS-Ser1177 and AMPK-Thr172 phosphorylation
The data obtained so far suggested that an increase in [Ca2+]i may be the reason for the increased eNOS activity upon DPPB treatment. To obtain proof for this assumption we tested whether incubation of EA.hy926 cells with the Ca2+ chelator Bapta AM (30 μM) could abolish the activating effects induced by DPPB. Chelation of intracellular Ca2+ indeed abrogated the stimulating effect of DPPB on eNOS-Ser1177 and AMPK-Thr172 phosphorylation (Fig. 5B), similar to the effects previously observed with compound C and STO 609 (Fig. 4A and B). Again, treatment of the cells with Bapta AM led to a basal increase in AMPK phosphorylation at Thr172, which, however, could not be further increased upon treatment with DPPB. All these results regarding basal activation of different proteins after treatment with pharmacological inhibitors strengthens our belief that there is a delicate balance between the upstream kinases of eNOS-Ser1177 (Akt, CaMKKβ, AMPK) and that interference with one of these kinases is likely to trigger compensatory actions by the others.
All data therefore point to a strong causal link between the increase in [Ca2+]i and the observed DPPB-induced increase of eNOS activity.
4. Discussion
In this study we show that the benzofuran derivative DPPB enhances eNOS activity leading to an increased NO availability in cultured human endothelial cells. The observed effect is associated with an increase in [Ca2+]i leading to the activation of eNOS via a signal transduction pathway involving Ca2+/CaM, CaMKKβ, and AMPK (Fig. 6).
Fig. 6.

Scheme of the proposed mechanism by which DPPB increases eNOS activity. Administration of DPPB results in an increase in intracellular Ca2+ and enhanced Ca2+/CaM signaling thereby activating CaMKKβ, AMPK and finally eNOS, ultimately resulting in enhanced NO production. The inhibitors used in the study (Bapta AM, STO 609, AMPKα siRNA and Compound C) and their place of action is also indicated.
Secondary metabolites from the benzofuran class are known to exhibit a broad range of bioactivities [16–18,20,21], however, an influence on eNOS activity was not described so far. Interestingly, out of several benzofuran derivatives isolated from K. lappacea [26], DPPB is the only compound which promotes eNOS activity in human endothelial cells.
DPPB increases endothelial NO release by a mechanism involving increased [Ca2+]i. eNOS activation via an increase of [Ca2+]i is a well-established mechanism since eNOS can be directly activated by protein–protein interaction with the intracellular Ca2+ receptor CaM enabling the electron transfer from NADPH at the reductase domain of the enzyme to the heme center at the oxygenase domain [12]. In addition, CaM is a known mediator of Ca2+ signaling directly binding and regulating a number of different target proteins, thereby affecting many different cellular functions [41]. A well-established target of CaM is CaMKKβ, which further translates the intracellular Ca2+ increase into phosphorylation of downstream target proteins, including AMPK phosphorylation at the threonine residue 172 [38,39]. Although the AMPK pathway is mainly considered as an important regulator of metabolism, it is also crucial for the maintenance of endothelial function and redox balance [42]. AMPK exerts anti-atherosclerotic effects by influencing various signaling cascades resulting in improved NO bioavailability, attenuated free radical generation and activation of angiogenic factors [39]. Furthermore, activated AMPK is able to reduce the activation of NF-κB in endothelial cells [43]. In addition to its beneficial effect on endothelial function, AMPK inhibits the proliferation of vascular smooth muscle cells thus supporting anti-atherosclerotic mechanisms [44]. Activated AMPK enhances eNOS enzyme activity by phosphorylation at the eNOS-Ser1177 residue [15].
Upon DPPB treatment we observed a strong increase of AMPK-Thr172 phosphorylation concomitant with increased eNOS-Ser1177 phosphorylation (Fig. 3) suggesting that eNOS is not only activated via a direct protein–protein interaction with Ca2+/CaM, but also via an additional regulatory loop involving Ca2+/CaM-dependent CaMKKβ activation, AMPK activation, and finally increased eNOS-Ser1177 phosphorylation (Fig. 6). The causal connection of these regulatory steps was indeed confirmed by the application of CaMKKβ and AMPK inhibitors (STO 609 and compound C, respectively) as well as by knockdown of AMPK (Fig. 4). These approaches were able to block the DPPB-induced increase of eNOS-Ser1177 phosphorylation, an accepted posttranslational modification activating eNOS [15]. Accompanying the increase in eNOS-Ser1177 phosphorylation, we observed a decreased phosphorylation of eNOS-Thr495 after treatment with DPPB (Fig. 3). The eNOS-Thr495 residue is located in the CaM-binding domain of the enzyme and decreased phosphorylation of Thr495 is known to correlate with a stronger CaM-eNOS interaction due to interference of the phosphorylated residue with CaM binding to the eNOS CaM-binding domain [45,46].
It is difficult to attribute specific effects to AMPK activation as there are several reports showing crosstalk between the Akt and the AMPK pathway [39]. However, our data showed no influence of DPPB on Akt, also a major regulator of eNOS-Ser1177 phosphorylation. Similar results have been shown for the regulation of eNOS by some agonists of G protein-coupled receptors such as bradykinin or histamine, which are independent on signaling from the PI3K/Akt pathway, but are known to utilize an increase in [Ca2+]i for their signal transduction in endothelial cells [46,47].
Taken together our results reveal that the benzofuran derivative DPPB, isolated from the medicinal plant K. lappacea, increases eNOS activity and NO availability in cultured endothelial cells. This action is mediated by a mechanism involving elevation of the intracellular Ca2+ levels and increased signaling via a pathway involving Ca2+/CaM, CaMKKβ, and AMPK. Furthermore, our study outlines DPPB as an interesting natural product that might have significant contribution in mediating the bioactivity of K. lappacea. Clearly further studies are necessary to better estimate whether DPPB has a good potential to be used as a pharmaceutical or health-promoting food supplement additive.
Conflict of interest
None.
Funding
This work was supported by grants from the Austrian Science Fund (FWF) [NFN S10703-B03 and S10703-B13 and S10704-B03 and S10704-B13].
Acknowledgements
We gratefully acknowledge the excellent technical assistance of Daniel Schachner, Judith Benedics, and Hortenzia Beres. We would also like to thank Dr. Hartmut Kleinert (Mainz, Germany) for the human iNOS control cell lysates and the respective iNOS antibody.
Contributor Information
Angela Ladurner, Email: angela.ladurner@univie.ac.at.
Atanas G. Atanasov, Email: atanas.atanasov@univie.ac.at.
Elke H. Heiss, Email: elke.heiss@univie.ac.at.
Lisa Baumgartner, Email: l.baumgartner@gmx.net.
Stefan Schwaiger, Email: stefan.schwaiger@uibk.ac.at.
Judith M. Rollinger, Email: judith.rollinger@uibk.ac.at.
Hermann Stuppner, Email: hermann.stuppner@uibk.ac.at.
Verena M. Dirsch, Email: verena.dirsch@univie.ac.at.
References
- 1.Luscher T.F., Barton M. Biology of the endothelium. Clin Cardiol. 1997;20:II-3–II-10. [PubMed] [Google Scholar]
- 2.Kinlay S., Libby P., Ganz P. Endothelial function and coronary artery disease. Curr Opin Lipidol. 2001;12:383–389. doi: 10.1097/00041433-200108000-00003. [DOI] [PubMed] [Google Scholar]
- 3.Chatterjee A., Black S.M., Catravas J.D. Endothelial nitric oxide (NO) and its pathophysiologic regulation. Vascul Pharmacol. 2008;49:134–140. doi: 10.1016/j.vph.2008.06.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Davignon J., Ganz P. Role of endothelial dysfunction in atherosclerosis. Circulation. 2004;109:III27–III32. doi: 10.1161/01.CIR.0000131515.03336.f8. [DOI] [PubMed] [Google Scholar]
- 5.Forstermann U., Munzel T. Endothelial nitric oxide synthase in vascular disease: from marvel to menace. Circulation. 2006;113:1708–1714. doi: 10.1161/CIRCULATIONAHA.105.602532. [DOI] [PubMed] [Google Scholar]
- 6.Loscalzo J. Nitric oxide insufficiency, platelet activation, and arterial thrombosis. Circ Res. 2001;88:756–762. doi: 10.1161/hh0801.089861. [DOI] [PubMed] [Google Scholar]
- 7.De Caterina R., Libby P., Peng H.B., Thannickal V.J., Rajavashisth T.B., Gimbrone M.A., Jr. Nitric oxide decreases cytokine-induced endothelial activation Nitric oxide selectively reduces endothelial expression of adhesion molecules and proinflammatory cytokines. J Clin Invest. 1995;96:60–68. doi: 10.1172/JCI118074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Rizzo N.O., Maloney E., Pham M., Luttrell I., Wessells H., Tateya S. Reduced NO-cGMP signaling contributes to vascular inflammation and insulin resistance induced by high-fat feeding. Arterioscler Thromb Vasc Biol. 2010;30:758–765. doi: 10.1161/ATVBAHA.109.199893. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Forstermann U. Nitric oxide and oxidative stress in vascular disease. Pflugers Arch. 2010;459:923–939. doi: 10.1007/s00424-010-0808-2. [DOI] [PubMed] [Google Scholar]
- 10.Dudzinski D.M., Michel T. Life history of eNOS: partners and pathways. Cardiovasc Res. 2007;75:247–260. doi: 10.1016/j.cardiores.2007.03.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Li L., Bressler B., Prameya R., Dorovini-Zis K., Van Breemen C. Agonist-stimulated calcium entry in primary cultures of human cerebral microvascular endothelial cells. Microvasc Res. 1999;57:211–226. doi: 10.1006/mvre.1998.2131. [DOI] [PubMed] [Google Scholar]
- 12.Michel T., Vanhoutte P.M. Cellular signaling and NO production. Pflugers Arch. 2010;459:807–816. doi: 10.1007/s00424-009-0765-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Fleming I., Busse R. Molecular mechanisms involved in the regulation of the endothelial nitric oxide synthase. Am J Physiol Regul Integr Comp Physiol. 2003;284:R1–R12. doi: 10.1152/ajpregu.00323.2002. [DOI] [PubMed] [Google Scholar]
- 14.Schmitt C.A., Dirsch V.M. Modulation of endothelial nitric oxide by plant-derived products. Nitric Oxide. 2009;21:77–91. doi: 10.1016/j.niox.2009.05.006. [DOI] [PubMed] [Google Scholar]
- 15.Mount P.F., Kemp B.E., Power D.A. Regulation of endothelial and myocardial NO synthesis by multi-site eNOS phosphorylation. J Mol Cell Cardiol. 2007;42:271–279. doi: 10.1016/j.yjmcc.2006.05.023. [DOI] [PubMed] [Google Scholar]
- 16.Halici Z., Dengiz G.O., Odabasoglu F., Suleyman H., Cadirci E., Halici M. Amiodarone has anti-inflammatory and anti-oxidative properties: an experimental study in rats with carrageenan-induced paw edema. Eur J Pharmacol. 2007;566:215–221. doi: 10.1016/j.ejphar.2007.03.046. [DOI] [PubMed] [Google Scholar]
- 17.Hu Z.F., Chen L.L., Qi J., Wang Y.H., Zhang H., Yu B.Y. Two new benzofuran derivatives with anti-inflammatory activity from Liriope spicata var. prolifera. Fitoterapia. 2011;82:190–192. doi: 10.1016/j.fitote.2010.09.002. [DOI] [PubMed] [Google Scholar]
- 18.Hur K.Y., Seo H.J., Kang E.S., Kim S.H., Song S., Kim E.H. Therapeutic effect of magnesium lithospermate B on neointimal formation after balloon-induced vascular injury. Eur J Pharmacol. 2008;586:226–233. doi: 10.1016/j.ejphar.2008.02.072. [DOI] [PubMed] [Google Scholar]
- 19.Fung K.P., Zeng L.H., Wu J., Wong H.N., Lee C.M., Hon P.M. Demonstration of the myocardial salvage effect of lithospermic acid B isolated from the aqueous extract of Salvia miltiorrhiza. Life Sci. 1993;52:PL239–PL244. doi: 10.1016/0024-3205(93)90471-e. [DOI] [PubMed] [Google Scholar]
- 20.Lee S.K., Cui B., Mehta R.R., Kinghorn A.D., Pezzuto J.M. Cytostatic mechanism and antitumor potential of novel 1H-cyclopenta[b]benzofuran lignans isolated from Aglaia elliptica. Chem Biol Interact. 1998;115:215–228. doi: 10.1016/s0009-2797(98)00073-8. [DOI] [PubMed] [Google Scholar]
- 21.Kapche G.D., Fozing C.D., Donfack J.H., Fotso G.W., Amadou D., Tchana A.N. Prenylated arylbenzofuran derivatives from Morus mesozygia with antioxidant activity. Phytochemistry. 2009;70:216–221. doi: 10.1016/j.phytochem.2008.12.014. [DOI] [PubMed] [Google Scholar]
- 22.Mei R.Q., Wang Y.H., Du G.H., Liu G.M., Zhang L., Cheng Y.X. Antioxidant Lignans from the Fruits of Broussonetia papyrifera. J Nat Prod. 2009;72:621–625. doi: 10.1021/np800488p. [DOI] [PubMed] [Google Scholar]
- 23.Carini M., Aldini G., Orioli M., Antioxidant Facino R.M. photoprotective activity of a lipophilic extract containing neolignans from Krameria triandra roots. Planta Med. 2002;68:193–197. doi: 10.1055/s-2002-23167. [DOI] [PubMed] [Google Scholar]
- 24.Simpson B. The past and present uses of rhatany (Krameria, Krameriaceae) Econ Bot. 1991;45:397–409. [Google Scholar]
- 25.Simpson B.B. Krameriaceae Flora Neotropica. 1989:1–108. [Google Scholar]
- 26.Baumgartner L., Sosa S., Atanasov A.G., Bodensieck A., Fakhrudin N., Bauer J. Lignan derivatives from Krameria lappacea roots inhibit acute inflammation in vivo and pro-inflammatory mediators in vitro. J Nat Prod. 2011;74:1779–1786. doi: 10.1021/np200343t. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Edgell C.J., McDonald C.C., Graham J.B. Permanent cell line expressing human factor VIII-related antigen established by hybridization. Proc Natl Acad Sci USA. 1983;80:3734–3737. doi: 10.1073/pnas.80.12.3734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Schmitt C.A., Handler N., Heiss E.H., Erker T., Dirsch V.M. No evidence for modulation of endothelial nitric oxide synthase by the olive oil polyphenol hydroxytyrosol in human endothelial cells. Atherosclerosis. 2007;195:e58–e64. doi: 10.1016/j.atherosclerosis.2007.02.024. [DOI] [PubMed] [Google Scholar]
- 30.O’Brien J., Wilson I., Orton T., Pognan F. Investigation of the Alamar Blue (resazurin) fluorescent dye for the assessment of mammalian cell cytotoxicity. Eur J Biochem. 2000;267:5421–5426. doi: 10.1046/j.1432-1327.2000.01606.x. [DOI] [PubMed] [Google Scholar]
- 31.Schmitt C.A., Heiss E.H., Aristei Y., Severin T., Dirsch V.M. Norfuraneol dephosphorylates eNOS at threonine 495 and enhances eNOS activity in human endothelial cells. Cardiovasc Res. 2009;81:750–757. doi: 10.1093/cvr/cvn326. [DOI] [PubMed] [Google Scholar]
- 32.Schalkwijk C.G., van Dam B., Stehouwer C.D., van Hinsbergh V.W. Mevastatin increases eNO synthase expression and inhibits lipid peroxidation in human endothelial cells. Clin Hemorheol Microcirc. 2007;37:179–184. [PubMed] [Google Scholar]
- 33.Sen-Banerjee S., Mir S., Lin Z., Hamik A., Atkins G.B., Das H. Kruppel-like factor 2 as a novel mediator of statin effects in endothelial cells. Circulation. 2005;112:720–726. doi: 10.1161/CIRCULATIONAHA.104.525774. [DOI] [PubMed] [Google Scholar]
- 34.Heller R., Unbehaun A., Schellenberg B., Mayer B., Werner-Felmayer G., Werner E.R. l-ascorbic acid potentiates endothelial nitric oxide synthesis via a chemical stabilization of tetrahydrobiopterin. J Biol Chem. 2001;276:40–47. doi: 10.1074/jbc.M004392200. [DOI] [PubMed] [Google Scholar]
- 35.Huang A., Vita J.A., Venema R.C., Keaney J.F., Jr. Ascorbic acid enhances endothelial nitric-oxide synthase activity by increasing intracellular tetrahydrobiopterin. J Biol Chem. 2000;275:17399–17406. doi: 10.1074/jbc.M002248200. [DOI] [PubMed] [Google Scholar]
- 36.Chen Z.P., Mitchelhill K.I., Michell B.J., Stapleton D., Rodriguez-Crespo I., Witters L.A. AMP-activated protein kinase phosphorylation of endothelial NO synthase. FEBS Lett. 1999;443:285–289. doi: 10.1016/s0014-5793(98)01705-0. [DOI] [PubMed] [Google Scholar]
- 37.Michell B.J., Griffiths J.E., Mitchelhill K.I., Rodriguez-Crespo I., Tiganis T., Bozinovski S. The Akt kinase signals directly to endothelial nitric oxide synthase. Curr Biol. 1999;9:845–848. doi: 10.1016/s0960-9822(99)80371-6. [DOI] [PubMed] [Google Scholar]
- 38.Woods A., Dickerson K., Heath R., Hong S.P., Momcilovic M., Johnstone S.R. Ca2+/calmodulin-dependent protein kinase kinase-beta acts upstream of AMP-activated protein kinase in mammalian cells. Cell Metab. 2005;2:21–33. doi: 10.1016/j.cmet.2005.06.005. [DOI] [PubMed] [Google Scholar]
- 39.Fisslthaler B., Fleming I. Activation and signaling by the AMP-activated protein kinase in endothelial cells. Circ Res. 2009;105:114–127. doi: 10.1161/CIRCRESAHA.109.201590. [DOI] [PubMed] [Google Scholar]
- 40.Swulius M.T., Waxham M.N. Ca(2+)/calmodulin-dependent protein kinases. Cell Mol Life Sci. 2008;65:2637–2657. doi: 10.1007/s00018-008-8086-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Chin D., Means A.R. Calmodulin: a prototypical calcium sensor. Trends Cell Biol. 2000;10:322–328. doi: 10.1016/s0962-8924(00)01800-6. [DOI] [PubMed] [Google Scholar]
- 42.Xu Q., Si L.Y. Protective effects of AMP-activated protein kinase in the cardiovascular system. J Cell Mol Med. 2010;14:2604–2613. doi: 10.1111/j.1582-4934.2010.01179.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Devaraj S., Torok N., Dasu M.R., Samols D., Jialal I. Adiponectin decreases C-reactive protein synthesis and secretion from endothelial cells: evidence for an adipose tissue-vascular loop. Arterioscler Thromb Vasc Biol. 2008;28:1368–1374. doi: 10.1161/ATVBAHA.108.163303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Igata M., Motoshima H., Tsuruzoe K., Kojima K., Matsumura T., Kondo T. Adenosine monophosphate-activated protein kinase suppresses vascular smooth muscle cell proliferation through the inhibition of cell cycle progression. Circ Res. 2005;97:837–844. doi: 10.1161/01.RES.0000185823.73556.06. [DOI] [PubMed] [Google Scholar]
- 45.Fleming I., Fisslthaler B., Dimmeler S., Kemp B.E., Busse R. Phosphorylation of Thr(495) regulates Ca(2+)/calmodulin-dependent endothelial nitric oxide synthase activity. Circ Res. 2001;88:E68–E75. doi: 10.1161/hh1101.092677. [DOI] [PubMed] [Google Scholar]
- 46.Fleming I. Molecular mechanisms underlying the activation of eNOS. Pflugers Arch. 2010;459:793–806. doi: 10.1007/s00424-009-0767-7. [DOI] [PubMed] [Google Scholar]
- 47.Tiruppathi C., Minshall R.D., Paria B.C., Vogel S.M., Malik A.B. Role of Ca2+ signaling in the regulation of endothelial permeability. Vascul Pharmacol. 2002;39:173–185. doi: 10.1016/s1537-1891(03)00007-7. [DOI] [PubMed] [Google Scholar]