

Abstract
The development of vaccines that prevent rabies has a long and distinguished history, with the earliest preceding modern understanding of viruses and the mechanisms of immune protection against disease. The correct application of inactivated tissue culture-derived vaccines is highly effective at preventing the development of rabies, and very few failures are recorded. Furthermore, oral and parenteral vaccination is possible for wildlife, companion animals and livestock, again using inactivated tissue culture-derived virus. However, rabies remains endemic in many regions of the world and causes thousands of human deaths annually. There also remain no means of prophylaxis for rabies once the virus enters the central nervous system (CNS). One reason for this is the poor immune response within the CNS to infection with rabies virus (RABV). New approaches to vaccination using modified rabies viruses that express components of the innate immune system are being applied to this problem. Preliminary reports suggest that direct inoculation of such viruses could trigger an effective anti-viral response and prevent a fatal outcome from RABV infection.
Keywords: central nervous system, rabies virus, reverse genetics, vaccination
Introduction
Rabies virus (RABV) is the type species of the genus Lyssavirus within the family Rhabdoviridae. The virus has a single-stranded ribonucleic acid (RNA) genome in the negative sense orientation that encodes five proteins in the following order: nucleoprotein (N), phosphoprotein (P), matrix protein (M), glycoprotein (G) and RNA polymerase (L) [1]. Infection with RABV causes in excess of 50 000 human deaths annually, the majority of which occur in Asia [2]. In addition to RABV, the lyssavirus genus contains a growing number of recognized and putative members (Table 1), many of which have been responsible for occasional human deaths [3]. RABV is a zoonotic virus and has a range of reservoir host species within the mammalian orders Carnivora and Chiroptera[4]. The most important of these reservoirs as a source of human disease is the domestic dog (Canis familiaris). In the Americas, a number of bat species are also responsible for transmission of rabies to humans, particularly the common vampire bat (Desmodus rotundus) in Latin America and a number of insectivorous bat species in North America [5]. This has emerged as a public health risk, as bites can occur without the victim realizing that an exposure has taken place, and many cases of bat-transmitted rabies have no recorded exposure to a bat prior to development of infection [6].
Table 1.
The lyssavirus genus
| Virus | Reservoir or known host | Causative agent of rabies in humans§ |
|---|---|---|
| Rabies virus (RABV) | Species of the orders Carnivora and Chiroptera* | Yes |
| Lagos bat virus (LBV) | Chiroptera† | No |
| Mokola virus (MOKV) | Not known | Yes |
| Duvenhage virus (DUVV) | Chiroptera† | Yes |
| European bat lyssavirus type 1 (EBLV-1) | Serotine bat (Eptesicus serotinus) | Yes |
| European bat lyssavirus type 2 (EBLV-2) | Daubenton's bat (Myotis daubentonii) | Yes |
| Australian bat lyssavirus (ABLV) | Chiroptera‡ | Yes |
| Aravan virus (ARAV) | Lesser mouse-eared bat (Myotis blythi) | No |
| Khujand virus (KHUV) | Whiskered bat (Myotis mystacinus) | No |
| Irkut virus (IRKV) | Greater tube-nosed bat (Murina leucogaster) | Yes |
| West Caucasian bat virus (WCBV) | Common bent-winged bat (Miniopterus schreibersii) | No |
| Shimoni bat virus (SHIBV) | Commerson's leaf-nosed bat (Hipposideros commersoni) | No |
| Bokeloh bat virus (BBLV) | Natterer's bat (Myotis nattereri) | No |
| Ikoma virus (IKOV) | Civet (Civettictis civetta) | No |
Bat species in the Americas only.
Bat species in Africa only.
Bat species in Australia only.
Rabies virus causes >50 000 human deaths annually; the other members of the genus Lyssavirus have caused occasional deaths (reviewed in [3]); no signifies no reported deaths.
Transmission of RABV occurs following a bite from an infected host resulting in the deposition of virus-laden saliva into a wound. RABV is highly neurotrophic and following a highly variable period, often lasting months, virus infects a peripheral nerve and ascends to the dorsal root ganglion [7]. Once within the spinal cord, the virus spreads rapidly to the brain, resulting in an overwhelming encephalitis that eventually kills the host. Examination of infected brains by histopathological methods reveals few gross changes with the exception, in many cases, of the presence of distinctive inclusion or Negri bodies. Immunolabelling reveals numerous infected neurones with accompanying gliosis and the development of perivascular cuffs around the parenchymal vasculature. This is found predominantly in the hindbrain [8]. Once signs of infection develop there is no effective treatment and, uniquely among infectious diseases, it has a case fatality rate of almost 100%. However, current vaccines are highly effective at protecting against this outcome provided that vaccination is given before or shortly after exposure to a biting incident. The development of virus-neutralizing antibodies is critical to preventing infection [9], and rabies vaccines are efficient at inducing an anti-rabies antibody response. However, the late development of neutralizing antibodies during RABV infection [10] is unexplained, and may be a critical factor responsible for the high fatality rate associated with the disease. Furthermore, there are no effective anti-viral treatments for rabies despite extensive investigations [11].
This review will provide an overview of the past, present and possible future of rabies vaccination, particularly considering the potential of vaccination to treat disease.
History of rabies vaccination
A comprehensive review on the development of rabies vaccines has been published recently [12]. What follows is a brief overview of key developments. Louis Pasteur developed the earliest effective vaccine against rabies that was first used to treat a human bite victim on 6 July 1885 [13]. The method involved inoculation with homogenates of RABV-infected rabbit spinal cord that had been desiccated progressively in sterile air. Initially, the recipient received a subcutaneous injection of homogenate that was fully inactivated. This was followed by injection of material derived from infections of spinal cord desiccated for shorter periods that contained progressively more virulent preparations of virus. Pasteur's approach proved highly effective, and the methodology spread widely. Two problems were associated with the approach. The first was the consistency of inactivation, which in some cases led to recipients possibly developing rabies from the vaccination, and secondly, the ability to produce sufficient vaccine from rabbits to meet the demand for treatment. These problems were resolved by inactivation of infected sheep or goat brain with chemical agents such as phenol [14,15]. These vaccines also proved successful but, like the original Pasteur vaccine, contained high levels of myelin that caused sensitization in some vaccine recipients and, in extreme cases, fatal encephalitis. Alternatives to this approach included inactivation of infected chick embryos [16] or inactivation of infected suckling mouse brain that has a lower level of myelin compared to the adult brain [17]. However, even these approaches were not entirely free of autoimmune reactions, and the World Health Organization (WHO) does not advocate the use of vaccines containing nervous tissue, although they are still used in a number of countries.
A new paradigm for rabies vaccines followed the development of cell culture for virus propagation. The first tissue culture vaccine was derived from virus grown in primary hamster kidney cells [18,19]. This was followed by growth of fixed RABV (see Box 1) in a human diploid cell line [21]. The lung-derived cell line WI-38 was used initially, but was switched subsequently to the MRC-5 cell line, which resulted in the development and licensing of a human diploid cell vaccine (HDCV) in the mid-1970s. An alternative to HDCVwas the use of purified chick embryo cells (PCEC) [22]. These vaccines are now used successfully worldwide.
Box 1. The concept of fixed virus
One of Louis Pasteur's achievements was to develop an animal model for the predictable passage of rabies virus (RABV). Pasteur solved this by infecting rabbits through introducing constant amounts of virus preparation onto the dura mater membrane after creating a hole in the skull (trepanation). After repeated passage by this method, in some instances more than 50 times, the incubation period from inoculation to the development of rabies became consistent at 7 days. The properties of the virus, which would now be termed a virus strain, were considered fixed. This has been applied to a large number of RABV strains that are now used commonly to develop rabies vaccines and in basic research on rabies pathogenesis. Examples include Pasteur virus (PV), Pitman Moore (PM), low egg passage (LEP) and Street Alabama Dufferin (SAD). The probable origins of these strains of RABV, many of which have been in existence for more than 50 years, were reviewed by Flamand and co-workers [20].
Administration of rabies vaccines
Uniquely among vaccines, those for rabies can be given both pre- and post-exposure to virus. Pre-exposure vaccination is appropriate for travellers to RABV-endemic regions, veterinarians and researchers working with the virus. Post-exposure vaccination is possible because the exposure event, usually a bite, is easily identifiable and the incubation period is of sufficient length for vaccination to induce a protective immune response. This is principally through the development of neutralizing antibodies. Post-exposure vaccination is usually accompanied by injection of anti-rabies immunoglobulin of either human (HRIG) or equine (ERIG) origin, and is referred to collectively as post-exposure prophylaxis (PEP). Whether PEP is given can be decided by the level of exposure (Table 2) which, despite the extreme consequences of developing disease, is a factor in resource-poor areas of the world.
Table 2.
World Health Organization recommendations for vaccination in response to contact with a potentially rabid animal
| Category | Contact | Action |
|---|---|---|
| I | Touch or normal animal husbandry such as feeding | No treatment required |
| II | Contact leads to minor scratches or abrasions that do not result in bleeding | Clean the wound with soap and water. Receive vaccination |
| Biting that does not lead to breaks in the skin | ||
| III | Bites or scratches that lead to skin damage | Clean wound with soap and water. Receive vaccination and anti-rabies immunoglobulin (human or equine origin) |
| Exposure of any sort to bats | ||
| Licks on broken skin/mucous membranes |
Pre-exposure vaccination consists of an intramuscular injection of 1 ml vaccine on days 0, 7, 21 and 28. Depending on the vaccine manufacturer, boosting is recommended at 3–5-year intervals. This has been borne out by recent cohort studies of UK bat workers who are required to be vaccinated against rabies prior to licensing to work with bats [23].
Post-exposure vaccination is given typically as an intramuscular injection on days 0, 3, 7, 14 and 30. HRIG is given on day 0, unless the recipient has received previous vaccination against rabies. One innovation has been the replacement of intramuscular inoculation with intradermal injection of vaccine [24]. This route of inoculation appears to require less vaccine to be effective, which reduces the cost of treatment. A disadvantage of this method is the increased difficulty in administering successful intradermal injections.
Cell culture-derived vaccines can be used for the parenteral vaccination of companion animals and livestock, and have also been used to develop oral vaccines for wildlife immunization [25]. The combination of high titres of attenuated strains of RABV with an oral bait attractive to wildlife vectors such as the red fox (Vulpes vulpes) have been highly effective at eliminating rabies from western Europe and remain in use throughout eastern Europe and Turkey [26,27].
Parenteral vaccination with tissue culture-derived vaccines has been administered since the 1970s, and has been used extensively in all continents of the world. They have low levels of side effects, can be produced at low cost and have found application in both human and veterinary medicine. The major disadvantage for PEP is the need for compliance with repeated injections to ensure that treatment is successful. In practice, this requires multiple trips to a health clinic which, if not followed, can lead to vaccine failure. The cost of a full course of vaccination, particularly in parts of Asia and Africa, also remains a problem regarding the delivery of appropriate, effective treatment to those who require it. These two factors, along with ignorance of both the disease and the consequences of a dog bite, explain the continuing persistence of human deaths due to rabies in many areas of the world. A vaccine that could achieve protection against rabies, but with fewer injections, would be of great benefit in the treatment of rabies.
Alternative development of rabies vaccines
Despite the undoubted success of current commercial vaccines against rabies there have been numerous attempts to develop alternatives, all taking advantage of the genetic manipulation revolution. Antibodies have been shown to be critical for protection against the spread of RABV [9]. The key target for antibodies is virus glycoprotein. Glycoprotein is the only surface-exposed protein on the virion particle, and a number of antigenic sites to which neutralizing monoclonal antibodies bind have been identified on this protein [28,29]. The ability to clone the RABV glycoprotein into bacterial plasmids and then express the protein in a range of systems has led to a number of alternative approaches with the potential for new vaccines against rabies. In each case the recombinant protein, expressed in a range of vectors, has been shown to be protective in mouse models of vaccination and virus challenge. The following are examples of this:
RABV glycoprotein expressed on the surface of the vaccinia virus [30].
RABV glycoprotein expressed on the surface of the canary pox virus [31].
RABV glycoprotein expressed by canine adenovirus [32].
A chimeric lyssavirus glycoprotein with segments derived from RABV and Mokola virus that provide immunization against more than one lyssavirus [33].
DNA vaccination with RABV glycoprotein cloned into a plasmid vector [34].
Despite the ability to rapidly induce high titres of RABV neutralizing antibodies, effective at preventing infection in small animal models, they have been unable to challenge existing vaccines, principally on cost and acceptance for human use.
Reverse genetics as a new paradigm in rabies vaccine development
A new avenue for research on RABV biology and rabies vaccine development was opened with the ability to manipulate the viral genome. An attenuated, fixed strain of RABV, SAD B19, a European derivative of an American SAD strain (see Box 1), was recovered from a plasmid-encoded genome by Conzelmann and Schnell [35]. This has paved the way for a range of developments in vaccination biology through manipulation of the RABV genome. Table 3 provides a visualization of these modified genomes and the outcomes from their application to pathogenesis and vaccine development studies.
Table 3.
Examples of recombinant rabies virus (RABV)
| Schematic | Comments | Reference |
|---|---|---|
 |
First demonstration of recovery of a recombinant RABV | [35] |
 |
Addition of a second glycoprotein coding sequenced increased expression, anti-glycoprotein antibody production and protection against subsequent infection with virulent RABV | [45] |
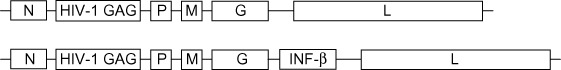 |
Introduction of the HIV GAG protein produced a potential vaccine candidate for HIV. Further introduction of the interferon β gene attenuated the virus but enhanced the induction of activated CD8+ T cells | [46] |
 |
Addition of CCL3 (MIP1α) reduced virulence, increased recruitment of dendritic cells to the inoculation site and increased anti-RABV antibody production | [47] |
 |
Intracerebral inoculation with a recombinant RABV expressing GM-CSF prevented rabies in mice infected with a virulent strain of RABV | [43] |
HIV: human immunodeficiency virus; MIP1α: macrophage inflammatory protein 1α; GM-CSF: granulocyte–macrophage colony-stimulating factor.
The persistent challenge in the field of RABV therapy is how to treat patients who have developed rabies beyond palliative care. A recent development and application of therapeutic coma [36] has met with a small number of successes, but also an increasing number of failures [10,37]. Experimental models suggest that the mammalian host produces a vigorous innate immune response to RABV infection in the brain [38–40]; however, there is strong evidence that this response is antagonized to some extent by the viral phosphoprotein [41,42]. This subversion may be the reason for the inability of the innate and adaptive immune response to control RABV replication in the central nervous system and leads ultimately to the death of the host. It is therefore encouraging that therapeutic treatment with modified rabies genomes appears to attenuate infection with a more virulent strain [43], and offers the future possibility that these may form the foundation of a future successful treatment to ameliorate the worst outcomes of RABV infection.
Conclusions
Rabies vaccines for humans are highly effective when given pre- or post-exposure to a bite from a potentially rabid animal. Recent alternatives have not (and are unlikely to in the near future) challenged current vaccines on the grounds of cost and acceptance. Indeed, it could be argued that there is no requirement for further development of vaccines for post-exposure treatment with current technologies, other than to ensure that the vaccine candidate is effective against the full spectrum of viruses within the genus Lyssavirus. However, the inability of current medicine to prevent patient death once RABV has entered the CNS remains a major challenge, particularly in the absence of effective anti-viral agents to RABV. Recent developments using reverse genetic approaches to modification of the RABV genome have generated potential tools for preventing rabies in murine models. Their use has been advocated for oral vaccination of dogs and wildlife [44]. These developments need to be explored further, but offer some hope for realizing the aim of preventing human deaths due to infection with RABV.
Acknowledgments
This work was funded by Department for Environment, Food and Rural Affairs (DEFRA) grant SV3500. This paper is published with the permission of the Controller of HMSO and the Queen's Printer for Scotland.
Disclosure
None.
References
- 1.Tordo N, Poch O, Ermine A, et al. Completion of the rabies virus genome sequence determination: highly conserved domains among the L (polymerase) proteins of unsegmented negative-strand RNA viruses. Virology. 1988;165:565–76. doi: 10.1016/0042-6822(88)90600-9. [DOI] [PubMed] [Google Scholar]
- 2.Sudarshan MK, Madhusudana SN, Mahendra BJ, et al. Assessing the burden of human rabies in India: results of a national multi-center epidemiological survey. Int J Infect Dis. 2007;11:29–35. doi: 10.1016/j.ijid.2005.10.007. [DOI] [PubMed] [Google Scholar]
- 3.Johnson N, Vos A, Freuling C, et al. Human rabies due to lyssavirus infection of bat origin. Vet Microbiol. 2010;142:151–9. doi: 10.1016/j.vetmic.2010.02.001. [DOI] [PubMed] [Google Scholar]
- 4.Badrane H, Tordo N. Host switching in lyssavirus history from the Chiroptera to the Carnivora orders. J Virol. 2001;75:8096–104. doi: 10.1128/JVI.75.17.8096-8104.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Messenger SL, Smith JS, Rupprecht CE. Emerging epidemiology of bat-associated cryptic cases of rabies in humans in the United States. Clin Infect Dis. 2002;35:738–47. doi: 10.1086/342387. [DOI] [PubMed] [Google Scholar]
- 6.De Serres G, Dallaire F, Côte M, Skowronski M. Bat rabies in the United States and Canada from 1950 through 2007: human cases with and without bat contact. Clin Infect Dis. 2008;46:1329–37. doi: 10.1086/586745. [DOI] [PubMed] [Google Scholar]
- 7.Schnell MJ, McGettigan JP, Wirblich C, Papaneri A. The cell biology of rabies virus: using stealth to reach the brain. Nat Rev Microbiol. 2010;8:51–61. doi: 10.1038/nrmicro2260. [DOI] [PubMed] [Google Scholar]
- 8.Hicks DJ, Nunez A, Healy DM, et al. Comparative pathological study of the murine brain after experimental infection with classical rabies virus and European bat lyssavirus. J Comp Pathol. 2009;140:113–26. doi: 10.1016/j.jcpa.2008.09.001. [DOI] [PubMed] [Google Scholar]
- 9.Hooper DC, Morimoto K, Bettle M, et al. Collaboration of antibody and inflammation in clearance of rabies virus from the central nervous system. J Virol. 1998;72:3711–9. doi: 10.1128/jvi.72.5.3711-3719.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Hunter M, Johnson N, Hedderwick S, et al. Immunovirological correlates in human rabies treated with therapeutic coma. J Med Virol. 2010;82:1255–65. doi: 10.1002/jmv.21785. [DOI] [PubMed] [Google Scholar]
- 11.Dacheux L, Delmas O, Bourhy H. Human rabies encephalitis prevention and treatment: progress since Pasteur's discovery. Infect Disord Drug Targets. 2011;11:251–99. doi: 10.2174/187152611795768079. [DOI] [PubMed] [Google Scholar]
- 12.Wu X, Smith TG, Rupprecht CE. From brain passage to cell adaptation: the road of human rabies vaccine development. Exp Rev Vaccines. 2011;10:1597–608. doi: 10.1586/erv.11.140. [DOI] [PubMed] [Google Scholar]
- 13.Bardenave G. Louis Pasteur (1822–1895) Microbes Infect. 2003;5:553–60. doi: 10.1016/s1286-4579(03)00075-3. [DOI] [PubMed] [Google Scholar]
- 14.Fermi C. Über die Immunisierung gegen Wutkrankheit. [Immunisation against rabies.] Z Hyg Infectionskrankh. 1908;58:233–76. [Google Scholar]
- 15.Semple D. The preparation of a safe and efficient antirabic vaccine. Sci Mem Med Sanit Dept India. 1911;44:1–31. [Google Scholar]
- 16.Koprowski H, Cox HR. Studies on chick embryo adapted rabies virus: I. Culture characteristics and pathogenicity. J Immunol. 1948;60:533–54. [PubMed] [Google Scholar]
- 17.Fuenzalida E, Palacios R, Borgono JM. Anti-rabies antibody responses in man to vaccine made from infected suckling-mouse brains. Bull World Health Organ. 1964;30:431–6. [PMC free article] [PubMed] [Google Scholar]
- 18.Kissling RE. Growth of rabies virus in non-nervous tissue. Proc Soc Exp Biol Med. 1958;98:223–5. doi: 10.3181/00379727-98-23997. [DOI] [PubMed] [Google Scholar]
- 19.Kissling RE, Reese DR. Anti-rabies vaccine of tissue culture origin. J Immunol. 1963;91:362–8. [PubMed] [Google Scholar]
- 20.Flamand A, Coulon P, Lafay F, Tuffereau C. Avirulent mutants of rabies virus and their use as live vaccine. Trends Microbiol. 1993;1:317–20. doi: 10.1016/0966-842x(93)90010-o. [DOI] [PubMed] [Google Scholar]
- 21.Wiktor TJ, Fernandes MV, Koprowski H. Cultivation of rabies virus in human diploid cell strain WI-38. J Immunol. 1964;93:353–66. [PubMed] [Google Scholar]
- 22.Kondo A. Growth characteristics of rabies virus in primary chicken embryo cells. Virology. 1965;27:199–204. doi: 10.1016/0042-6822(65)90160-1. [DOI] [PubMed] [Google Scholar]
- 23.Morris J, Crowcroft NS, Fooks AR, et al. Rabies antibody levels in bat handlers in the United Kingdom: immune response before and after purified chick embryo cell rabies booster vaccination. Hum Vaccin. 2007;3:165–70. doi: 10.4161/hv.3.5.4216. [DOI] [PubMed] [Google Scholar]
- 24.Verma R, Khanna P, Prinja S, Rajput M. Intra-dermal administration of rabies vaccines in developing countries: at an affordable cost. Hum Vaccin. 2011;7:792–4. doi: 10.4161/hv.7.7.15410. [DOI] [PubMed] [Google Scholar]
- 25.Gueu L, Schares S, Schinck C, et al. Genetic characterisation of attenuated SAD rabies virus strains used for oral vaccination of wildlife. Vaccine. 2008;26:3227–35. doi: 10.1016/j.vaccine.2008.04.007. [DOI] [PubMed] [Google Scholar]
- 26.Cliquet F, Aubert M. Elimination of terrestrial rabies in Western European countries. Dev Biol. 2004;119:185–204. [PubMed] [Google Scholar]
- 27.Vos A, Freuling C, Eskiizmirliler S, et al. Rabies in foxes, Aegean Region, Turkey. Emerg Infect Dis. 2009;15:1620–2. doi: 10.3201/eid1510.090203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Seif I, Coulon P, Rollin PE, Flamand A. Rabies virulence: effect on pathogenicity and sequence characterisation of rabies virus mutations affecting antigenic site III of the glycoprotein. J Virol. 1985;53:926–34. doi: 10.1128/jvi.53.3.926-934.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Prehaud C, Coulon P, Lafay F, et al. Antigenic site II of rabies virus glycoprotein: structure and role in viral virulence. J Virol. 1988;62:1–7. doi: 10.1128/jvi.62.1.1-7.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Wiktor TJ, Macfarlan RI, Reagan KJ, et al. Protection from rabies by a vaccinia virus recombinant containing the rabies virus glycoprotein gene. Proc Natl Acad Sci USA. 1984;81:7194–8. doi: 10.1073/pnas.81.22.7194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Cadoz M, Strady A, Meignier B, et al. Immunisation with canary pox virus expressing rabies glycoprotein. Lancet. 1992;339:1429–32. doi: 10.1016/0140-6736(92)92027-d. [DOI] [PubMed] [Google Scholar]
- 32.Yarosh OK, Wandeler AI, Graham FL, et al. Human adenovirus type 5 vectors expressing rabies glycoprotein. Vaccine. 1996;14:1257–564. doi: 10.1016/s0264-410x(96)00012-6. [DOI] [PubMed] [Google Scholar]
- 33.Jallet C, Jacob Y, Bahloul C, et al. Chimeric lyssavirus glycoproteins with increased immunological potential. J Virol. 1999;73:225–33. doi: 10.1128/jvi.73.1.225-233.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Lodmell DL, Ewalt LC. Post-exposure DNA vaccination protects mice against rabies virus. Vaccine. 2001;19:2468–73. doi: 10.1016/s0264-410x(00)00475-8. [DOI] [PubMed] [Google Scholar]
- 35.Conzelmann KK, Schnell M. Rescue of synthetic genome RNA analogs of rabies virus by plasmid encoded proteins. J Virol. 1994;68:713–9. doi: 10.1128/jvi.68.2.713-719.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Willoughby RE, Tieves KS, Hoffman GM, et al. Survival after treatment of rabies with induction of coma. N Engl J Med. 2005;352:2508–14. doi: 10.1056/NEJMoa050382. [DOI] [PubMed] [Google Scholar]
- 37.McDermid RC, Sacinger L, Lee B, et al. Human rabies encephalitis following bat exposure: failure of therapeutic coma. Can Med Assoc J. 2008;178:557–61. doi: 10.1503/cmaj.071326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Wang ZH, Sarmento L, Wang Y, et al. Attenuated rabies virus activities, while pathogenic rabies virus evades the host innate immune responses in the central nervous system. J Virol. 2005;79:12554–65. doi: 10.1128/JVI.79.19.12554-12565.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.McKimmie CS, Johnson N, Fooks AR, Fazakerley JK. Viruses selectively upregulate Toll-like receptors in the central nervous system. Biochem Biophys Res Commun. 2005;336:925–33. doi: 10.1016/j.bbrc.2005.08.209. [DOI] [PubMed] [Google Scholar]
- 40.Johnson N, Mansfield KL, Hicks D, et al. Inflammatory responses in the nervous system of mice infected with a street isolate of rabies virus. Dev Biol. 2008;131:65–72. [PubMed] [Google Scholar]
- 41.Vidy A, Chelbi-Alix M, Blondel D. Rabies virus P protein interacts with STAT 1 and inhibits interferon signal transduction. J Virol. 2005;79:14411–20. doi: 10.1128/JVI.79.22.14411-14420.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Brzozka K, Finke S, Conzelmann KK. Inhibition of interferon signalling by rabies virus phosphoprotein P: activation-dependent binding of STAT1 and STAT2. J Virol. 2006;80:2675–83. doi: 10.1128/JVI.80.6.2675-2683.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Wen Y, Wang H, Wu H, et al. Rabies virus expressing dendritic cell-activating molecules enhances the innate and adaptive immune response to vaccination. J Virol. 2011;85:1634–44. doi: 10.1128/JVI.01552-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Faber M, Dietzschold B, Li J. Immunogenicity and safety of recombinant rabies viruses used for oral vaccination of stray dogs and wildlife. Zoonoses Public Health. 2009;56:262–9. doi: 10.1111/j.1863-2378.2008.01215.x. [DOI] [PubMed] [Google Scholar]
- 45.Faber M, Pumanausakahul R, Hodawadekar SS, et al. Overexpression of the rabies virus glycoprotein results in enhancement of apoptosis and antiviral immune response. J Virol. 2002;76:3374–81. doi: 10.1128/JVI.76.7.3374-3381.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Faul EJ, Wanjalla CN, McGettigan JP, Schnell MJ. Interferon-β expressed by a rabies virus-based HIV-1 vaccine vector serves as a molecular adjuvant and decreases pathogenicity. Virology. 2008;382:226–38. doi: 10.1016/j.virol.2008.09.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Zhao L, Toriumi H, Wang H, et al. Expression of MIP-1alpha (CCL3) by a recombinant rabies virus enhances its immunogenicity by inducing innate immunity and recruiting dendritic cells and B cells. J Virol. 2010;84:9642–8. doi: 10.1128/JVI.00326-10. [DOI] [PMC free article] [PubMed] [Google Scholar]