

Abstract
Capillary electrophoresis with laser induced fluorescence detection (CE-LIF) was employed for rapid sialic acid speciation, facilitating the quantitative determination of N-glycolylneuraminic acid (Neu5Gc) and N-acetylneuraminic acid (Neu5Ac) on glycoproteins. Derivatization of the sialic acids with 2-aminoacridone (2-AMAC), using classical reductive amination in a non-aqueous solvent, led to the spontaneous decarboxylation of the sialic acid residues as determined by CE-LIF and offline mass spectrometric analysis. Modification of both the labeling conditions to drive the decarboxylation reaction to completion and the CE-LIF parameters to separate the neutral species by complexation with a neutral coated capillary and borate reversed polarity, led to a robust platform for the rapid, sensitive and quantitative speciation of sialic acids. The method can readily be used for quality control of recombinant biopharmaceuticals.
Keywords: Sialic acid, N-acetylneuraminic acid, N-glycolylneuraminic acid, capillary electrophoresis, monosaccharide, 2-aminoacridone, decarboxylation
Introduction
Sialic acids are nine carbon monosaccharides that typically terminate the antennary chains of both N- and O-glycans via enzymatic addition by a variety of sialyltransferases1–3. N-acetylneuraminic acid (Neu5Ac) and its hydroxylated form, N-glycolylneuraminic acid (Neu5Gc), are the two major sialic acids found in mammals4. Neu5Gc is not expressed in humans due to the evolutionary loss of the gene encoding the enzyme that converts Neu5Ac into Neu5Gc (CMP-Neu5Ac hydroxylase)5. However, studies have demonstrated the metabolic incorporation of Neu5Gc into human glycoproteins by both normal and cancerous tissues due to the dietary availability of Neu5Gc6. Furthermore, it has been shown that humans possess circulating antibodies against Neu5Gc7–9. As a consequence, glycans attached to protein therapeutics expressed in cell lines capable of Neu5Gc incorporation into the attached oligosaccharides have an associated immunogenic potential10. Therefore, identification and quantitation of Neu5Gc in therapeutic glycoproteins is of high importance11.
Methods for the speciation of sialic acids include the analysis of liberated sialic acids by capillary electrophoresis (CE) and gas (GC) or liquid chromatography (LC) after derivatization with a chromophore or fluorophore for sensitive detection12–15. Mass spectrometry14 has also been employed due to the sixteen Dalton mass difference between the Neu5Ac and Neu5Gc forms11, 16. For separation methods with optical detection, derivatization of the sialic acids is routinely accomplished with a neutral dye using reductive amination, for example, 2-aminacridone (2-AMAC) in DMSO with sodium cyanoborohydride or 1,2-diamino-4,5-dimethoxybenzene (DMB) in mercaptoethanol and sodium dithionite17. Herein, we examined the conditions of sialic acid labeling with 2-AMAC to improve the yield of the reaction, thus increasing the overall reliability and sensitivity of the CE-LIF method. Interestingly, during derivatization using classical reductive amination with 2-AMAC and sodium cyanoborohydride in DMSO/acetic acid, the sialic acids were observed to spontaneously decarboxylate. The underlying process governing the derivatization was characterized using CE and offline MS analysis. A modified labeling method was combined with an optimized CE separation leading to the rapid, quantitative determination of sialic acid species present on glycoproteins. The simplicity of the approach allows for straightforward transfer of the method into a biopharmaceutical quality control analytical environment.
Experimental
Chemicals and reagents
Neu5Ac and Neu5Gc sialic acid standards, 2-aminoacridone (2-AMAC), anhydrous DMSO, sodium cyanoborohydride (1 M solution in THF), acetic acid and alpha-1-acid glycoprotein from both bovine and human plasma were all from Sigma Aldrich (St. Louis, MO). Water and acetonitrile were Thermo Fisher LC-MS Optima Grade (Fair Lawn, NJ).
Sialic acid derivatization
During initial experiments, Neu5Ac and Neu5Gc were derivatized using 2 μL of 0.1 M 2-AMAC prepared in 85:15 anhydrous DMSO acetic acid, followed by the addition of 2 μL of 1 M sodium cyanoborohydride in THF and incubation at 55 °C for 60 minutes. Later, after optimization studies, sialic acid derivatization was performed under the same conditions except that 2-AMAC was prepared in 15% v/v acetic acid.
Sialic acid release
Sialic acids were released from glycoproteins either by acid hydrolysis (2M acetic acid at 80°C for two hours) or by treating the oligosaccharides with sialidase A from Arthrobacter ureafaciens (Prozyme, Hayward, CA). Glycans were reconstituted in 50 mM ammonium acetate pH 5.5, and sialidase was added to 10% of the final volume; the reaction mixture was incubated at 37°C overnight. Released sialic acids were collected by filtration through prewashed 10 kDa molecular weight cut off filters. The filtrate was reduced to dryness and then derivatized, as previously described.
Capillary electrophoresis
Capillary electrophoresis was conducted on a PA 800 Plus Pharmaceutical Analysis System equipped with laser-induced fluorescence detection (488 nm excitation/520 nm emission) (Beckman Coulter Inc., Brea, CA). The specific capillaries and background electrolytes are described in the figure legends.
Mass spectrometry
MS analysis was performed in the flow injection mode using 70:30:0.1 methanol, water, formic acid as mobile phase at 50 μL/min flow rate, delivered by a Waters 2695 Alliance HPLC (Milford, MA). The injection volume was 10 μL. Mass spectrometric measurements were conducted using a Waters LCT ESI-Tof instrument operated in the positive ion mode. The ESI voltage was 3.2 kV, and the cone and extraction voltages were 30 V and 5 V, respectively. The lens voltage was adjusted to 180 V, and the desolvation temperature was 100°C. The source temperature was 120°C, and the flow rates of cone gas and desolvation gas were 50 L/hr and 450 L/hr, respectively.
Results and Discussion
Speciation of sialic acids present on therapeutic glycoproteins is a necessary requirement to identify the sialic acid forms and to quantitate the amount of non-human sialic acids, such as N-glycolylneuraminic acid. Herein, we describe the optimization of the reductive amination conditions necessary for labeling of released sialic acids with 2-AMAC, leading to a rapid and simple CE-LIF method for quantitative sialic acid speciation.
Sialic acids labeled with 2-AMAC using classical reductive amination spontaneously decarboxylate
Neu5Ac and Neu5Gc standards were initially derivatized using 0.1 M 2-AMAC solution containing 85% v/v anhydrous DMSO/15% v/v acetic acid and 1 M sodium cyanoborohydride (NaBH3CN). CE-LIF separation was performed using a 20 μm ID bare fused silica capillary with normal polarity, as previously described18. As shown in Fig. 1(A), five major peaks were observed, one of which corresponded to excess unreacted 2-AMAC, which migrated with the electroosmotic flow (EOF). The minor peaks in Fig. 1(A) arise from low level contaminants in the 2-AMAC dye as revealed in the analysis of a reagent blank, data not shown. The presence of 4 peaks, rather than 2, suggests potential analyte degradation or modification of the sialic acids during the derivatization reaction. Such changes could clearly affect quantitative results. In addition, the experimental reproducibility was found to significantly decline from injection to injection (Supplementary Figure S-1). Moreover, the bare fused silica capillary required washing and extensive equilibration between analyses to maintain the desired EOF19. The method was clearly not satisfactory.
Fig. 1.

Separation of Neu5Ac and Neu5Gc. 10 μg of Neu5Ac and Neu5Gc were labeled in 0.1 M 2-AMAC solution in 15% v/v acetic acid in DMSO. (A) Capillary: bare fused silica, total length: 60 cm, effective length: 50 cm, 20 μm ID, applied field: 500 V/cm, normal polarity. BGE: 200 mM boric acid buffer, pH 10. (B) Capillary: neutral coated, total length: 60 cm, effective length: 50 cm, 30 μm ID, applied field: 500 V/cm, reversed polarity. BGE: same as (A). (C) Separation after heat treatment of the sample. Separation conditions are the same as (B). (D) Injection of the heat treated sample. Separation conditions are the same as (B) and (C) except for the BGE: 20 mM ammonium acetate buffer, pH 10. All separations were carried out at 25°C on a Beckman PA800 Plus Instrument.
To overcome the column re-equilibration issue, the sialic acids were analyzed using a neutral coated capillary (no EOF) in the reversed polarity mode using same BGE as in Fig. 1(A). In the absence of EOF, peaks 1 and 2, Fig. 1(B) migrated later than peaks 3 and 4, suggesting that peaks 1 and 2 were neutral species and peaks 3 and 4 were the negatively charged sialic acids. The neutrality of peaks 1 and 2 could result from the loss of the charged carboxylic group of the sialic acids. To investigate this possibility, the sialic acids were heated at 80°C for 60 minutes to promote decarboxylation of the sialic acid residue followed by analysis with reversed polarity CE-LIF. Fig. 1(C) shows the absence of peaks 3 and 4 after the treatment at 80°C, with increased area of peaks 1 and 2. To demonstrate that peaks 1 and 2 are neutral species, boric acid was next removed from the BGE. As expected, peaks 1 and 2 were no longer observed, Fig. 1(D), as the borate was not present to form the charged complex and the neutral species would not migrate in the electric field.
MS investigation of sialic acid decarboxylation during 2-AMAC derivatization
Offline mass spectrometry using an electrospray ionization time of flight mass spectrometer (ESI-Tof MS) was next employed to confirm the identity of peaks 1 and 2 and to examine various conditions that could affect the decomposition process. Fig. 2(A) shows the mass spectrum of standard Neu5Ac without derivatization. Both the [M+H]+ molecular ion and a sodiated adduct at m/z 332 and 354, respectively, are observed. When the labeling reaction was carried out with the standard protocol under non-aqueous conditions (0.1 M 2-AMAC in 85% DMSO/15% of glacial acetic acid (v/v), overnight labeling at 37°C and 1 M NaBH3CN), the protonated 2-AMAC-Neu5Ac conjugate and its sodiated adduct appeared at m/z 504.36 and 526, respectively, Fig. 2(B), corresponding to the mass shift associated with the addition of the 2-AMAC fluorophore. However, this conjugate underwent decomposition during the labeling reaction, as evidenced by the peak at m/z 460, Fig. 2(B), 44 mass units lower than the ion corresponding to [2-AMAC Neu5Ac+H]+. This ion is due to the neutral loss of carbon dioxide from the sialic acid residue via decarboxylation during the derivatization reaction. The result supports the interpretation of analyte behavior shown in Fig. 1, wherein using the combination of CE migration under differential conditions accompanied by offline MS analysis the presence of the decarboxylated sialic derivatives can be confirmed.
Fig. 2.

Mass spectra and electropherograms of Neu5Ac under different labeling conditions: (A) Neu5Ac solution (10 μg/100 μL) without labeling; (B) Labeling of 10 μg of Neu5Ac in 5 μL of 0.1 M 2-AMAC- in 15% acetic acid/85% DMSO and 5 μL 1 M NaBH3CN in DMSO at 37°C overnight; (C) Labeling of 10 μg Neu5Ac in 5 μL of 0.05 M aqueous 2-AMAC in 15% acetic acid at 37°C and 5 μL 1 M NaBH3CN in DMSO at 37°C overnight; (D) 10 μg Neu5Ac in 50 μL of 2 M acetic acid solution, pre-heated at 80°C for 2 hours, then dried and labeled according to the procedure described in (B); (E) Neu5Ac solution pretreated at 80°C for 2 hours in 2 M acetic acid; (F) 10 μg of Neu5Ac labeled according to the procedure described in (C), followed by quenching of the reaction by the addition of 50 μL of water, and the addition of APTS labeled maltopentaose as internal standard 8; (G) 10 μg of Neu5Ac labeled according to the procedure described in (C), and processed as described in (F). Electropherogram (I) was acquired before heating, whereas electropherogram (II) was acquired after incubation at 80°C for 1 hour. Peak 1: unknown, peak 2: 2-AMAC-Neu5Ac (intact), peak 3: 2-AMAC-Neu5Ac (decomposed).
Importantly, Fig. 2(B) shows that labeling of sialic acids using classical reductive amination by sodium cyanoborohydride in a DMSO/acetic acid solvent system is incomplete, as indicated by the ions at m/z 332 and 354, corresponding to the native material. The incomplete reaction occurs even when the dye is used in a 3-fold excess (data not shown). In contrast, when the sialic acids were labeled with a 50 mM solution of 2-AMAC dissolved in 15% v/v aqueous acetic acid, no unlabeled sialic acid derivative was detected, Fig. 2(C), indicating that the reaction was complete. To further investigate the labeling mechanism, the reaction was next repeated under aqueous and anhydrous conditions. Fig. 2(C) clearly shows that in the presence of water, the decomposition of AMAC-sialic acid conjugate was complete as only the decarboxylated conjugate (m/z 460) and its corresponding sodiated adduct (m/z 482) were observed. Fig. 2(B–D) also reveals that the decarboxylated sialic acid underwent further loss of water, yielding an ion at m/z 442. Conducting the labeling reaction in 50 mM 2-AMAC prepared in 15% v/v aqueous acetic acid solution and 1 M NaBH3CN resulted in complete decarboxylation and quantitative labeling of sialic acid as the 2-AMAC-Neu5Ac neutral conjugate was generated, peak 3 Fig. 2(F). Neither the Neu5Ac conjugate in the electropherogram nor unreacted Neu5Ac in the mass spectrum, Fig. 2(C) were observed. In comparison, Neu5Ac labeling in 15% v/v acetic acid in DMSO and 1 M NaBH3CN resulted in only 15% of peak 3 and the intact 2-AMAC-Neu5Ac conjugate, peak 2 Fig. 2(G), electropherogram I. Heating this mixture at 80°C for 1 hour increased the yield of peak 3 to 36%. The disappearance of peak 2 demonstrates the increased lability of 2-AMAC-Neu5Ac at higher temperature. A similar yield of 30% of neutral 2-AMAC-Neu5Gc in 15% v/v acetic acid in DMSO was observed, suggesting identical behavior of both sialic acids under the same derivatization conditions, Fig. 2(G), electropherogram II and Supplementary Figure S-2.
Fig. 2(D and E) show that pretreatment of sialic acids, i.e. exposure to elevated temperatures commonly used for thermally assisted desialylation with no subsequent derivatization, does not alter their structure. These data suggest that the native ring conformation of sialic acids is very stable. However, labeling with 2-AMAC promotes ring opening, resulting in conjugate lability and subsequent decarboxylation. It is interesting to note that this decomposition cannot be avoided even if the reaction is carried out under anhydrous conditions due to the presence of residual trace quantities of water in the non-aqueous solvent, as determined by KF titration (likely due to trace absorption of atmospheric moisture). Therefore, to facilitate quantitative speciation of sialic acids released from therapeutic glycoproteins, methods for analyte stabilization are required. In this case, the use of aqueous acetic acid solution to promote complete labeling and decarboxylation of the sialic acid-dye conjugate is recommended. With this approach, quantitation of sialic acids can easily be achieved through analysis of the decomposition products using CE in reversed polarity.
Rapid CE-LIF speciation of 2-AMAC labeled sialic acids
Once sialic acids were labeled and quantitatively converted to their neutral forms, the CE-LIF separation conditions were fine-tuned by optimizing the boric acid concentration (in the BGE), the electric field and the temperature. Sialic acid speciation analysis using a BGE containing 200 mM boric acid proved optimal. Complete baseline resolution between Neu5Gc and Neu5Ac was achieved at 25°C with 500 V/cm electric field, Supplementary Figure S-3 (A). The electric field strength was further increased to 600 V/cm by shortening the length of the capillary, resulting in faster and more efficient separation, Supplementary Figure S-3 (B). Separation of 2-AMAC derivatized sialic acids was also significantly influenced by the column temperature due to the strong complexation between boric acid and the labeled conjugates20. For CE-LIF separations performed at 25°C, Fig. 3, the peak corresponding to Neu5Gc showed fronting. Increasing the temperature from 25°C to 45°C resulted in a significant improvement of the symmetry of Neu5Gc peak, (As Neu5Gc: 0.63 at 25°C, 0.88 at 45°C). At elevated temperatures, complexation with borate present in the BGE leads to more efficient analyte peaks allowing impurities to be separated from the Neu5Gc peak, Supplementary Figure S-4.
Fig. 3.
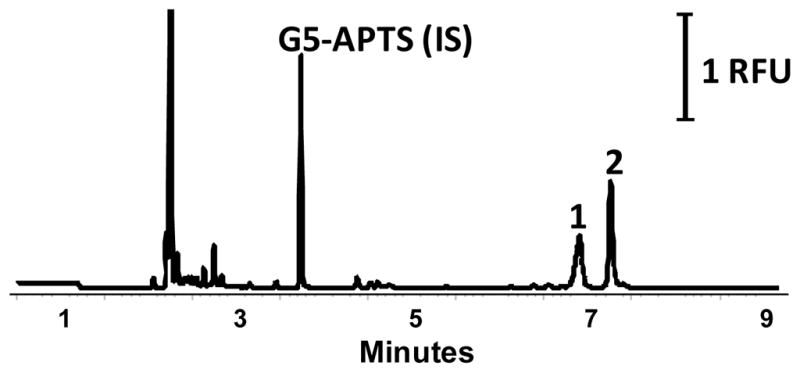
(A) Optimized CE-LIF separation of the decarboxylated 2-AMAC sialic acid conjugates; separation conditions as outlined in the text, E = 600 V/cm, and BGE of 200 mM boric acid pH 10, T = 45°C,
The developed method was sensitive and linear over the range of 1–150 ng/μL, the correlation co-efficient (R2) being greater than 0.995 for both Neu5Ac and Neu5Gc. To illustrate the application of the analysis, 50 μg of human and bovine α-1-acid-glycoprotein were desialylated using sialidase, derivatized under optimized conditions and analyzed by CE-LIF. Reproducibility for five replicate injections for both the released bovine and human sialic acids pools was less than 2% RSD, Table 1. The evaluation of quantitative accuracy is complicated by the lack of available reference materials and the heterogeneity of commercially available glycoproteins. The present approach is suitable not only for the reproducible detection and quantitation of sialic acids, such as Neu5Ac and Neu5Gc, but also O-acetylated sialic acid variants.
Table 1.
Analytical reproducibility for sialic acids released from α-1-acid glycoprotein from bovine and human plasma.
| μg Neu5Gc/1mg bovine A1G | μg Neu5Ac/1mg bovine A1G | μg Neu5Ac/1mg human A1G | |
|---|---|---|---|
| Release | |||
| 1 | 29.72 | 34.17 | 74.77 |
| 2 | 28.31 | 31.31 | 74.93 |
| 3 | 25.13 | 29.91 | 76.55 |
| 4 | 26.89 | 29.37 | 73.78 |
| 5 | 27.49 | 30.50 | 75.66 |
| Average | 27.51 | 31.05 | 75.14 |
| STD | 1.70 | 1.89 | 1.04 |
Conclusions
A novel CE-LIF separation method was developed to speciate and quantify sialic acids present on glycoproteins. The results demonstrate that modification of the labeling conditions can result in the formation of neutral dye-sugar conjugates that can be rapidly separated using reversed polarity CE-LIF in a coated capillary in the absence of EOF. Mass spectrometric analysis revealed that the labeling reaction in the presence of water was complete, whereas in non aqueous solutions, the reaction was incomplete due to the neutral loss of carbon dioxide, thus limiting the quantitation of sialic acid speciation. The presented method offers advantage over HPLC based profiling methods using DMB as the fluorophore as the separation mechanism in our CE method is based on the formation of borate-sialic acid complexes with minimal interference from the 2-AMAC fluorophore. A problem with HPLC profiling of DMB labeled sialic acids is the presence of co-eluting DMB impurity peaks leading to complicated peak annotation and structure assignment.. The described method can accurately and reproducibly detect and quantify sialic acids in glycoproteins, and its simplicity has potential for quality control of biotechnology products.
Supplementary Material
Acknowledgments
This research was supported by NIH GM 15847. The authors thank Beckman Coulter for their generous gifts of products to support this study. Sandor Spisak and Matthew Trahan are kindly acknowledged for their technical assistance. Contribution No.1018 from the Barnett Institute.
References
- 1.Angata T, Varki A. Chemical diversity in the sialic acids and related alpha-keto acids: an evolutionary perspective. Chem Rev. 2002;102(2):439–69. doi: 10.1021/cr000407m. [DOI] [PubMed] [Google Scholar]
- 2.Kelm S, Schauer R. Sialic acids in molecular and cellular interactions. Int Rev Cytol. 1997;175:137–240. doi: 10.1016/S0074-7696(08)62127-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Traving C, Schauer R. Structure, function and metabolism of sialic acids. Cell Mol Life Sci. 1998;54(12):1330–49. doi: 10.1007/s000180050258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Varki A. Colloquium paper: uniquely human evolution of sialic acid genetics and biology. Proc Natl Acad Sci U S A. 2010;107(Suppl 2):8939–46. doi: 10.1073/pnas.0914634107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Nguyen DH, Tangvoranuntakul P, Varki A. Effects of natural human antibodies against a nonhuman sialic acid that metabolically incorporates into activated and malignant immune cells. J Immunol. 2005;175(1):228–36. doi: 10.4049/jimmunol.175.1.228. [DOI] [PubMed] [Google Scholar]
- 6.Tangvoranuntakul P, Gagneux P, Diaz S, Bardor M, Varki N, Varki A, Muchmore E. Human uptake and incorporation of an immunogenic nonhuman dietary sialic acid. Proc Natl Acad Sci U S A. 2003;100(21):12045–50. doi: 10.1073/pnas.2131556100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Lu Q, Padler-Karavani V, Yu H, Chen X, Wu SL, Varki A, Hancock WS. LC-MS analysis of polyclonal human anti-Neu5Gc xeno-autoantibodies immunoglobulin G Subclass and partial sequence using multistep intravenous immunoglobulin affinity purification and multienzymatic digestion. Anal Chem. 2012;84(6):2761–8. doi: 10.1021/ac2030893. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Padler-Karavani V, Hurtado-Ziola N, Pu M, Yu H, Huang S, Muthana S, Chokhawala HA, Cao H, Secrest P, Friedmann-Morvinski D, Singer O, Ghaderi D, Verma IM, Liu YT, Messer K, Chen X, Varki A, Schwab R. Human xeno-autoantibodies against a non-human sialic acid serve as novel serum biomarkers and immunotherapeutics in cancer. Cancer Res. 2011;71(9):3352–63. doi: 10.1158/0008-5472.CAN-10-4102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Padler-Karavani V, Yu H, Cao H, Chokhawala H, Karp F, Varki N, Chen X, Varki A. Diversity in specificity, abundance, and composition of anti-Neu5Gc antibodies in normal humans: potential implications for disease. Glycobiology. 2008;18(10):818–30. doi: 10.1093/glycob/cwn072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Ghaderi D, Taylor RE, Padler-Karavani V, Diaz S, Varki A. Implications of the presence of N-glycolylneuraminic acid in recombinant therapeutic glycoproteins. Nat Biotechnol. 2010;28(8):863–7. doi: 10.1038/nbt.1651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Maeda E, Kita S, Kinoshita M, Urakami K, Hayakawa T, Kakehi K. Analysis of nonhuman N-glycans as the minor constituents in recombinant monoclonal antibody pharmaceuticals. Anal Chem. 2012;84(5):2373–9. doi: 10.1021/ac300234a. [DOI] [PubMed] [Google Scholar]
- 12.Hara S, Yamaguchi M, Takemori Y, Nakamura M, Ohkura Y. Highly sensitive determination of N-acetyl- and N-glycolylneuraminic acids in human serum and urine and rat serum by reversed-phase liquid chromatography with fluorescence detection. J Chromatogr. 1986;377:111–9. doi: 10.1016/s0378-4347(00)80766-5. [DOI] [PubMed] [Google Scholar]
- 13.Morimoto N, Nakano M, Kinoshita M, Kawabata A, Morita M, Oda Y, Kuroda R, Kakehi K. Specific distribution of sialic acids in animal tissues as examined by LC-ESI-MS after derivatization with 1,2-diamino-4,5-methylenedioxybenzene. Anal Chem. 2001;73(22):5422–8. doi: 10.1021/ac0104328. [DOI] [PubMed] [Google Scholar]
- 14.Shahrokh Z, Royle L, Saldova R, Bones J, Abrahams JL, Artemenko NV, Flatman S, Davies M, Baycroft A, Sehgal S, Heartlein MW, Harvey DJ, Rudd PM. Erythropoietin produced in a human cell line (Dynepo) has significant differences in glycosylation compared with erythropoietins produced in CHO cell lines. Mol Pharm. 2011;8(1):286–96. doi: 10.1021/mp100353a. [DOI] [PubMed] [Google Scholar]
- 15.Tzanakakis GN, Syrokou A, Kanakis I, Karamanos NK. Determination and distribution of N-acetyl- and N-glycolylneuraminic acids in culture media and cell-associated glycoconjugates from human malignant mesothelioma and adenocarcinoma cells. Biomed Chromatogr. 2006;20(5):434–9. doi: 10.1002/bmc.573. [DOI] [PubMed] [Google Scholar]
- 16.Stehling P, Gohlke M, Fitzner R, Reutter W. Rapid analysis of O-acetylated neuraminic acids by matrix assisted laser desorption/ionization time-of-flight mass spectrometry. Glycoconj J. 1998;15(4):339–44. doi: 10.1023/a:1006965600322. [DOI] [PubMed] [Google Scholar]
- 17.Guttman A. Analysis of monosaccharide composition by capillary electrophoresis. J Chromatogr A. 1997;763(1–2):271–7. doi: 10.1016/s0021-9673(96)00750-9. [DOI] [PubMed] [Google Scholar]
- 18.Grill E, Huber C, Oefner P, Vorndran A, Bonn G. Capillary zone electrophoresis of p-aminobenzoic acid derivatives of aldoses, ketoses and uronic acids. Electrophoresis. 1993;14(10):1004–10. doi: 10.1002/elps.11501401160. [DOI] [PubMed] [Google Scholar]
- 19.Oefner PJ, Chiesa C. Capillary electrophoresis of carbohydrates. Glycobiology. 1994;4(4):397–412. doi: 10.1093/glycob/4.4.397. [DOI] [PubMed] [Google Scholar]
- 20.Hoffstetter-Kuhn SPA, Gassmann E, Widmer HM. Influence of borate complexation on the electrophoretic behavior of carbohydrates in capillary electrophoresis. Anal Chem. 1991;63(15):1541–1547. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.