

Abstract
Angiotensin II receptor blockers (ARBs) are widely used as hypertensive therapeutic agents. In addition, studies have provided evidence that ARBs have the potential to inhibit the growth of several types of cancer cells. It was reported that telmisartan (a type of ARB) has peroxisome proliferator-activated receptor (PPAR)-γ activation activity. We previously reported that the PPAR-γ ligand induces growth arrest in human urological cancer cells through apoptosis. In this study, we evaluated the effects of telmisartan and other ARBs on cell proliferation in renal cell carcinoma (RCC), bladder cancer (BC), prostate cancer (PC) and testicular cancer (TC) cell lines. The inhibitory effects of telmisartan and other ARBs (candesartan, valsartan, irbesartan and losartan) on the growth of the RCC, BC, PC and TC cell lines was investigated using an MTT assay. Flow cytometry and Hoechst staining were used to determine whether the ARBs induced apoptosis. Telmisartan caused marked growth inhibition in the urological cancer cells in a dose- and time-dependent manner. Urological cancer cells treated with 100 μM telmisartan underwent early apoptosis and DNA fragmentation. However, the other ARBs had no effect on cell proliferation in any of the urological cancer cell lines. Telmisartan may mediate potent anti-proliferative effects in urological cancer cells through PPAR-γ. Thus, telmisartan is a potent target for the prevention and treatment of human urological cancer.
Keywords: telmisartan, angiotensin II receptor blocker, urological cancer, apoptosis
Introduction
Angiogenetic factors play key roles in urological as well as other types of cancer. In recent years, the expression of angiogenic factors in solid human tumors has been widely reported (1). Growth factors secreted by tumor cells, such as fibroblast and transforming growth factors, increase neovascularization in vivo and in vitro (2). Studies have demonstrated that peroxisome proliferator-activated receptor (PPAR)-γ ligands inhibit the growth of cancer cells in vitro and in vivo (3). PPAR-γ, a nuclear hormone receptor, provides a strong link between lipid metabolism and the regulation of gene transcription (4). PPAR-γ acts in the adipose tissue and promotes lipogenesis under anabolic conditions. Recently, the receptor has also been implicated in inflammation and carcinogenesis. Significant evidence from many experimental systems suggests that PPAR-γ plays an important role in carcinogenesis.
Angiotensin II is known as a key biological peptide in the renin-angiotensin system, which regulates blood pressure and renal hemodynamics, and angiotensin II receptor blockers (ARBs) are widely used as anti-hypertensive drugs (5). It is well known that angiogenesis is essential for tumor progression and metastasis (6,7). Several studies have shown that angiotensin II induces neovascularization and that ARBs inhibit vascular endothelial growth factor (VEGF) production (8,9). Benson et al discovered a structural resemblance between telmisartan (a type of ARB) and a PPAR-γ ligand approved for the treatment of type II diabetes, and reported that telmisartan has PPAR-γ action (10).
Our previous research revealed the expression of PPARs in human urological cancers and investigated the administration of PPAR-γ ligands as an anti-cancer therapy (11–15). With this background, the present study aimed to evaluate the inhibitory effect of telmisartan on human renal cell carcinoma (RCC), bladder cancer (BC), prostate cancer (PC) and testicular cancer (TC) cell lines, and to determine whether telmisartan induces apoptosis in such cells.
Materials and methods
Reagents and materials
RPMI-1640 was purchased from Nissui Pharmaceutical Company (Tokyo, Japan). Fetal bovine serum (FBS) and penicillin-streptomycin mixture were from Biowhittaker (Walkersville, MD, USA). Trypsin/EDTA was from Gibco BRL (Rockville, MD, USA). The angiotensin II receptor blockers telmisartan, candesartan, valsartan and irbesartan were from Toronto Research Chemicals, Inc. (Ontario, Canada). One of the ARBs, losartan, was from Cayman Chemical (Ann Arbor, MI, USA).
Cell cultures
The human RCC cell line (Caki-1) was provided by Dr Shinichi Ikemoto (Department of Urology, Osaka City University School of Medicine, Osaka, Japan). The human BC cell line (transitional cell carcinoma T24), PC cell lines (LNCaP, PC3 and DU-145) and TC cell line (NEC-8) were obtained from the Health Science Research Resources Bank (HSRRB, Osaka, Japan). Cells were grown in a culture flask (Nunc, Roskilde, Denmark) in RPMI-1640 supplemented with 10% FBS, 100 U/ml penicillin and 100 μg/ml streptomycin in a humidified 5% CO2 atmosphere at 37°C. The media were changed every 3 days, and the cells were separated via trypsinization using trypsin/EDTA upon reaching subconfluence.
Cell-proliferative studies
Approximately 1.0×104 cells placed onto 8×8-mm-diameter multichamber slides (Nunc, Copenhagen, Denmark) were treated with telmisartan, and the other ARBs were dissolved in ethanol. The final concentration of ethanol was <0.05%. Cell viability was measured on day 1 by a microplate reader using a modified 3-[4,5-dimethylthiazol-2-thiazolyl]-2,5-diphenyltetrazolium bromide (MTT) assay (WST-1 assay; Dojindo, Kumamoto, Japan) and presented as the percentage of cells under the control culture conditions.
Flow cytometry
Annexin V and propidium iodide staining
The effects of telmisartan and the other ARBs (candesartan, valsartan, irbesartan and losartan) on the urological cancer cells were determined by dual staining with Annexin V-FITC and propidium iodide (PI) using the Annexin V-FITC Apoptosis Detection kit I (Biosiences Pharmingen). Annexin V-FITC and PI were added to the cellular suspension as per the manufacturer's instructions, and a sample fluorescence of 1.0×104 cells was analyzed by flow cytometry conducted with FACScan (Becton Dickinson, Heidelberg, Germany).
Annexin V-FITC-positive and PI-negative cells were identified as early apoptotic. Annexin V-FITC-positive and PI-positive cells were identified as late apoptotic or necrotic.
Identification of DNA fragmentation
An assay was performed using the TdT-mediated dUTP Nick End Labelling (TUNEL) method using the Apo-Direct™ kit (Becton Dickinson). Following the experiments, the urological cancer cells in suspension (1.0×106/ml) were fixed with 1% PBS, washed in PBS and suspended in 70% (v/v) ice-cold ethanol, then stored in ethanol at −20°C until use. Positive and negative controls and the sample were stained with FITC-dUTP by incubation in terminal deoxynucleotidyl transferase buffer as per the manufacturer's instructions, and sample fluorescence of 1.0×104 cells was analyzed by flow cytometry (Becton Dickinson). Results are expressed as the percentage (%) of TUNEL-positive cells.
Detection of apoptosis
DNA chromatin morphology was assessed using Hoechst staining. The urological cancer cells were incubated with 100 μM telmisartan and the other ARBs for 24 h. Cells were washed with RPMI-1640 and labeled with 8 mg/ml of Hoechst 33342 (Sigma-Aldrich Japan K.K., Tokyo, Japan) for 10 min; PI (Sigma-Aldrich Japan K.K.) was added (10 mg/ml final concentration), and the cells were examined by fluorescence microscopy.
Results
Telmisartan-induced growth inhibition in urological cancer cells determined by MTT
To investigate the effects of telmisartan and the other ARBs on RCC, BC, PC and TC cell proliferation, we analyzed cell viability in vitro using a modified MTT assay.
Telmisartan induced a reduction in cell viability with a half-maximal concentration of growth inhibition in the urological cancer cell lines (Table I) in the range of 25–100 μM (Table I). Furthermore, in cells counted on days 1, 2 and 3, marked inhibition of cell proliferation using 100 μM of telmisartan was clearly visible (Table II). Telmisartan arrested the growth of all the urological cancer cells. However, the other ARBs had no effect on cell proliferation in any of the urological cancer cell lines (Table I).
Table I.
Dose-dependent effects of angiotensin II receptor blockers (ARBs) on the viability of human urological cancer cell lines.
| ARB dose
|
||||
|---|---|---|---|---|
| 25 μM (%) | 50 μM (%) | 100 μM (%) | ||
| Telmisartan | ||||
| RCC cell line | Caki-1 | 53.2% | 36.3% | 19.1% |
| BC cell line | T24 | 44.8 | 21.7 | 12.4 |
| PC cell lines | LNCaP | 32.0 | 18.7 | 12.7 |
| PC3 | 56.9 | 49.2 | 24.3 | |
| DU-145 | 62.2 | 42.0 | 22.3 | |
| TC cell line | NEC-8 | 43.3 | 32.6 | 32.6 |
| Candesartan | ||||
| RCC cell line | Caki-1 | 92.7 | 90.6 | 102.3 |
| BC cell line | T24 | 99.5 | 91.9 | 75.1 |
| PC cell lines | LNCaP | 128.0 | 129.0 | 141.5 |
| PC3 | 117.5 | 110.0 | 107.6 | |
| DU-145 | 95.0 | 106.9 | 102.0 | |
| TC cell line | NEC-8 | 118.0 | 122.1 | 120.1 |
| Valsartan | ||||
| RCC cell line | Caki-1 | 125.2 | 110.4 | 101.8 |
| BC cell line | T24 | 105.4 | 101.9 | 89.7 |
| PC cell lines | LNCaP | 147.8 | 141.2 | 121.8 |
| PC3 | 109.5 | 98.1 | 123.1 | |
| DU-145 | 97.5 | 86.7 | 68.8 | |
| TC cell line | NEC-8 | 209.7 | 171.3 | 110.1 |
| Irbesartan | ||||
| RCC cell line | Caki-1 | 99.7 | 139.1 | 89.9 |
| BC cell line | T24 | 78.3 | 100.7 | 92.3 |
| PC cell lines | LNCaP | 111.7 | 112.9 | 95.8 |
| PC3 | 118.9 | 106.8 | 111.3 | |
| DU-145 | 88.4 | 109.1 | 82.2 | |
| TC cell line | NEC-8 | 117.0 | 112.4 | 152.7 |
| Losartan | ||||
| RCC cell line | Caki-1 | 83.4 | 100.2 | 105.4 |
| BC cell line | T24 | 98.4 | 124.5 | 133.0 |
| PC cell lines | LNCaP | 89.4 | 86.3 | 84.0 |
| PC3 | 102.6 | 106.4 | 112.5 | |
| DU-145 | 129.6 | 114.0 | 110.2 | |
| TC cell line | NEC-8 | 104.5 | 133.2 | 155.5 |
The dose-response analysis of viability in human urological cancer cells treated with ARBs (25–100 μM, 48 h) was measured using the MTT assay and expressed as the % of cells under control culture conditions. Telmisartan stopped the growth of all urological cancer cells.
Table II.
Effects of telmisartan on the cell growth of human urological cancer cell lines in a time-dependent manner.
| 0 h (%) | 24 h (%) | 48 h (%) | 72 h (%) | ||
|---|---|---|---|---|---|
| Control culture (cell number) | 1.0×105 | 20.0×105 | 130.0×105 | 300.0×105 | |
| Telmisartan | |||||
| RCC cell line | Caki-1 | 76.5 | 58.1 | 31.8 | |
| BC cell line | T24 | 34.3 | 12.7 | 18.3 | |
| PC cell lines | LNCaP | 64.3 | 23.7 | 25.5 | |
| PC3 | 60.9 | 40.0 | 39.4 | ||
| DU-145 | 60.9 | 11.1 | 29.2 | ||
| TC cell line | NEC-8 | 29.2 | 21.5 | 18.3 | |
Human urological cancer cell lines were treated with telmisartan (100 μM) for 3 days. Cell numbers were determined on days 1, 2 and 3, and expressed as the % of cells under control culture conditions. Telmisartan arrested the growth of all the urological cancer cell lines.
Telmisartan-induced apoptosis in urological cancer cells determined by flow cytometry
To evaluate whether or not the cell death induced by telmisartan and the other ARBs was achieved through apoptosis, flow cytometry was used (Fig. 1). Treatment with 100 μM telmisartan induced early apoptosis in almost all the urological cancer cell lines.
Figure 1.

Effects of telmisartan on early and late apoptosis as shown by flow cytometry. Treatment with 100 μM telmisartan induced early apoptosis in almost the total percentage of the urological cancer cell lines. The top left quadrants represent early apoptosis (Annexin V-FITC-positive and PI-negative cells). The top right quadrants represent late apoptosis and necrosis (Annexin V-FITC-positive and PI-positive cells). Typical flow cytometry analysis histograms in a representative experiment are shown (A, Caki-1; B, T24; C, LNCaP; D, PC3; E, DU-145; F, NEC-8).
At a concentration of 100 μM, telmisartan induced DNA fragmentation in all the urological cancer cell lines (Fig. 2A–F). In contrast, the other ARBs did not induce DNA fragmentation in the urological cancer cells (data not shown).
Figure 2.

Telmisartan induced DNA fragmentation in the urological cancer cell lines. Diagrams of FITC-Annexin V/PI flow cytometry in a representative experiment are shown. The top left quadrants represent early apoptosis (Annexin V-FITC-positive and PI-negative cells) and the top right quadrants represent late apoptosis and necrosis (Annexin V-FITC-positive and PI-positive cells). Telmisartan (100 μM) induced DNA fragmentation in all the urological cancer cells. Histograms of typical flow cytometry analysis in a representative experiment are shown (A, Caki-1; B, T24; C, LNCaP; D, PC3; E, DU-145; F, NEC-8).
Effect of telmisartan on the induction of apoptosis in urological cancer cells
To evaluate whether or not the cell death induced by telmisartan was due to apoptosis, we evaluated the chromatin morphology of the urological cancer cells using Hoechst 33342 staining. All the urological cancer cells treated with telmisartan showed significant chromatin and cytoplasmic condensation, cellular shrinkage and small membrane-bound (apoptotic) bodies, while the urological cancer cells treated with the other ARBs did not show any of the above characteristics. The former showed cellular changes that were typically common characteristics of apoptosis (Fig. 3A–F).
Figure 3.
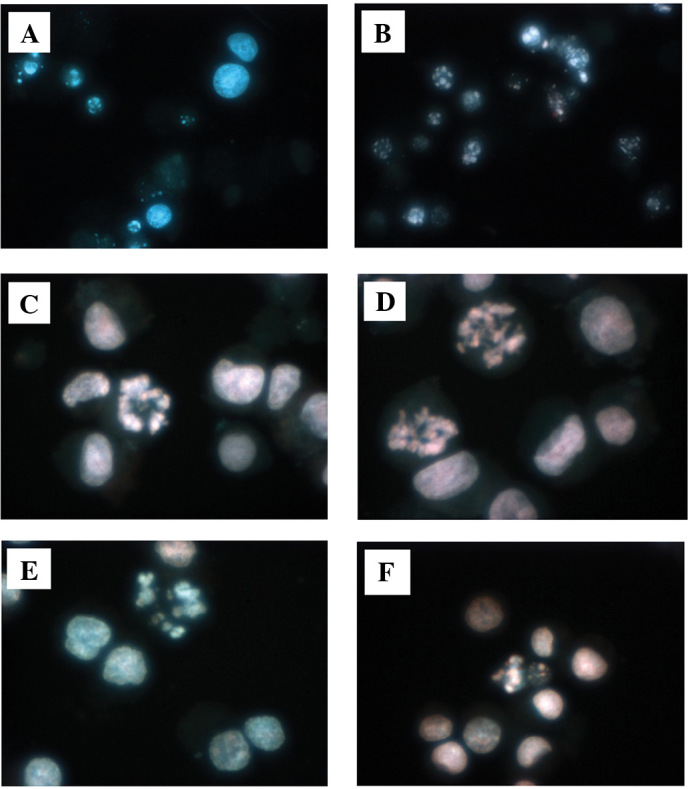
Effects of telmisartan on the induction of apoptosis in human urological cancer cell lines. Cells treated with termisartan showed significant chromatin and cytoplasmic condensation, cellular shrinkage and small membrane-bound (apoptotic) bodies. These cellular changes are typically common characteristics of apoptosis (A, Caki-1; B, T24; C, LNCaP; D, PC3; E, DU-145; F, NEC-8).
Discussion
ARBs have been synthesized and available for the treatment of hypertension since the 1990s (16,17). More recently, angiotensin II has been found to promote tumor growth and angiogenesis, and ARBs have been considered a significant anticancer and anti-angiogenesis therapeutic option (18).
Some types of tumor cells, such as melanoma, pancreatic cancer (19), RCC (20,21), breast cancer (22), BC (23) and PC (24), have been reported to express the angiotensin II receptor, and various studies have investigated the anti-tumor effects caused by the anti-angiogenesis of ARBs. Some researchers have demonstrated that candesartan (a type of ARB) inhibited the production of VEGF, one of the most potent and specific angiogenic factors, and decreased the growth of PC (24,28). Kosaka et al (24) found that a specific ARB suppressed VEGF production, resulting in reduced tumor angiogenesis and slower progression of PC in a tumor xenograft model. Concerning other tumor types, Kosugi et al (23) demonstrated that candesartan prevented the pulmonary metastasis of RCC and BC by inhibiting tumor angiogenesis through the suppression of VEGF in a xenograft model. Uemura et al (27) reported that, upon administering candesartan clinically to PC patients with hypertension, the level of prostate-specific antigen declined and the performance status of the patients improved. However, they also reported that candesartan had no effects on tumor growth in vitro, and did not detect apoptosis. Based on their in vitro and in vivo experiments, they suggested that the anti-tumor effect of ARB is not a result of direct toxicity or apoptotic induction, but of its anti-angiogenic effect.
The present study showed that candesartan and the other ARBs (except telmisartan) did not induce a reduction in cell viability and early apoptosis in urological cancer cells. Only telmisartan induced a reduction in cell viability with a half-maximal concentration of growth inhibition, and early apoptosis and DNA fragmentation in urological cancer cells.
Benson et al discovered a structural resemblance between telmisartan and pioglitazone, a PPAR-γ ligand approved for the treatment of type II diabetes. They found that telmisartan, not only blocks the angiotensin II receptor, but also activates PPAR-γ. Telmisartan functioned as a moderately potent selective PPAR-γ partial agonist, activating the receptor to 25–30% of the maximum level achieved by the full agonists pioglitazone and resiglitazone (10).
PPARs are members of the nuclear receptor superfamily of ligand-activated transcriptional factors such as steroids, thyroid hormones, vitamin D3 and retinoic acid. PPAR binds to the peroxisome proliferator response element as a heterodimer with the retinoic receptor in the regulation of PPAR target genes. PPARs are considered important immunomodulatory factors as well as fatty acid regulators. PPARs modulate these activities in different immune cell types, such as monocytes/macrophages, lymphocytes and endothelial cells (28).
PPAR-γ is expressed at a high level in adipose tissue and is a critical regulator of adipocyte differentiation. It is also expressed in the immune system, the spleen, monocyte bone-marrow precursors and helper T-cell clones, and in chondrocytes, synovial and bone tissues. Data have indicated that PPAR-γ ligands lead to the inhibition of phorbol ester-induced nitric oxide and macrophage-derived cytokines, such as tumor necrosis factor-α, interleukin-1β and interleukin-6, chemokines and adhesion molecules, in part by antagonizing the activities of transcriptional factors (29). It has been demonstrated that thiazolidinedione (a specific ligand for PPAR-γ, a new class of anti-diabetic medication) regulates the differentiation of cancer cells (30), and that nuclear-acting prostanoids including 15-d-PGJ2 are potent activators of the PPAR-γ receptor isoform (31,32). 15-d-PDJ2 induces apoptosis in macrophages, endothelial and choriocarcinoma cells (33–35), and thiazolidinedione induces fibroblast apoptosis (4).
We previously reported that PPAR-γ is strongly expressed in urological cancer tissues. The extent and intensity of PPAR-γ expression in urological cancer tissues were greater than in normal urological tissues. PPAR-γ ligands strongly induced early apoptosis in urological cancer cells as determined by flow cytometry and Hoechst staining (11–15). In this study, only telmisartan had a direct toxicity through apoptosis. Thus, telmisartan may mediate potent anti-proliferative effects against urological cancer cells through PPAR-γ. However, in our study, that dose was not clinically achievable. Further studies are needed to extend the use of telmisartan to clinical trials for the treatment for human urological cancer.
References
- 1.Weidner N, Folkman J, Pozza F, Bevilaqua P, Allred EN, Moore DH. Tumor angiogenesis: a new significant and independent prognostic indicator in early stage breast carcinoma. J Natl Cancer Inst. 1992;84:1875–1887. doi: 10.1093/jnci/84.24.1875. [DOI] [PubMed] [Google Scholar]
- 2.Lafyatis R, Thompson NL, Remmers EF, Flanders KC, Roche NS, Kim SJ. Transforming growth factor-beta production by synovial tissues from rheumatoid patients and streptococcal cell wall arthritic rats. Studies on secretion by synovial fibroblast-like cells and immunohistologic localization. J Immunol. 1989;143:1142–1148. [PubMed] [Google Scholar]
- 3.Kubota T, Koshizuka K, Williamson EA, et al. Ligand for Peroxisome proliferator-activated receptor-γ (troglitazone) has potent antitumor effect against human prostate cancer both in vitro and in vivo. Cancer Res. 1998;58:3344–3352. [PubMed] [Google Scholar]
- 4.Spiegelman BM. PPAR-gamma: adipogenic regulator and thiazolidinedione receptor. Diabetes. 1998;47:507–514. doi: 10.2337/diabetes.47.4.507. [DOI] [PubMed] [Google Scholar]
- 5.See S, Stirling AL. Candesartan cilexetil: an angiotensin II receptor blocker. Am J Health Syst Pharm. 2000;57:739–746. doi: 10.1093/ajhp/57.8.739. [DOI] [PubMed] [Google Scholar]
- 6.Folkman J. Tumor angiogenesis: therapeutic implications. N Engl J Med. 1971;285:1182–1186. doi: 10.1056/NEJM197111182852108. [DOI] [PubMed] [Google Scholar]
- 7.Folkman J. Angiogenesis in cancer, vascular, rheumatoid and other disease. Nat Med. 1995;1:27–31. doi: 10.1038/nm0195-27. [DOI] [PubMed] [Google Scholar]
- 8.Le Noble FA, Hekking JW, van Straaten HW, Slaaf DW, Struyker Boudier HA. Angiotensin II stimulates angiogenesis in the chorio-allantoic membrane of the chick embryo. Eur J Pharmacol. 1991;195:305–306. doi: 10.1016/0014-2999(91)90552-2. [DOI] [PubMed] [Google Scholar]
- 9.Le Noble FA, Schreurs NH, van Straaten HW, et al. Evidence for a novel angiotensin II receptor involved in angiogenesis in chick embryo chorioal-lantoic membrane. Am J Physiol. 1993;264:460–465. doi: 10.1152/ajpregu.1993.264.2.R460. [DOI] [PubMed] [Google Scholar]
- 10.Benson SC, Pershadsingh HA, Ho CI, et al. Identification of telmisartan as a unique angiotensin II receptor antagonist with selective PPARγ-modulating activity. Hypertension. 2004;43:993–1002. doi: 10.1161/01.HYP.0000123072.34629.57. [DOI] [PubMed] [Google Scholar]
- 11.Inoue K, Kawahito Y, Tsubouchi Y, et al. Expression of peroxisome proliferator-activated receptor gamma in renal cell carcinoma and growth inhibition by its agonists. Biochem Biophys Res Commun. 2001;287:727–732. doi: 10.1006/bbrc.2001.5640. [DOI] [PubMed] [Google Scholar]
- 12.Yoshimura R, Matsuyama M, Segawa Y, et al. Expression of peroxisome proliferator-activated receptors (PPARs) in human urinary bladder carcinoma and growth inhibition by its agonists. Int J Cancer. 2003;104:597–602. doi: 10.1002/ijc.10980. [DOI] [PubMed] [Google Scholar]
- 13.Segawa Y, Yoshimura R, Hase T, et al. Expression of peroxisome proliferator-activated receptor (PPAR) in human prostate cancer. Prostate. 2002;51:108–116. doi: 10.1002/pros.10058. [DOI] [PubMed] [Google Scholar]
- 14.Hase T, Yoshimura R, Mitsuhashi M, et al. Expression of peroxisome proliferator-activated receptors in human testicular cancer and growth inhibition by its agonists. Urology. 2002;60:542–547. doi: 10.1016/s0090-4295(02)01747-8. [DOI] [PubMed] [Google Scholar]
- 15.Yoshimura R, Matsuyama M, Hase T, et al. The effect of peroxisome proliferator-activated receptor-γ ligand on urological cancer cells. Int J Mol Med. 2003;12:861–865. [PubMed] [Google Scholar]
- 16.Burnier M. Angiotensin II type 1 receptor blockers. Circulation. 2001;103:904–912. doi: 10.1161/01.cir.103.6.904. [DOI] [PubMed] [Google Scholar]
- 17.Dina R, Jafari M. Angiotensin II-receptor antagonists. Am J Health-Syst Pharm. 2000;57:1231–1241. doi: 10.1093/ajhp/57.13.1231. [DOI] [PubMed] [Google Scholar]
- 18.Abali H, Güllü IH, Engin H, Haznedaroğlu IC, Erman M, Tekuzman G. Old antihypertensive as novel antineoplastics: angiotensin-I-converting enzyme inhibitors and angiotensin II type 1 receptor antagonists. Med Hypotheses. 2002;59:344–348. doi: 10.1016/s0306-9877(02)00185-8. [DOI] [PubMed] [Google Scholar]
- 19.Fujimoto Y, Sasaki T, Tsuchida A, Chayama K. Angiotensin II type 1 receptor expression in human pancreatic cancer and growth inhibition by angiotensin II type 1 receptor antagonist. FEBS Lett. 2001;495:197–200. doi: 10.1016/s0014-5793(01)02377-8. [DOI] [PubMed] [Google Scholar]
- 20.Miyajima A, Kosaka T, Asano T, et al. Angiotensin II type 1 antagonist prevents pulmonary metastasis of murine renal cancer by inhibiting tumor angiogenesis. Cancer Res. 2002;62:4176–4179. [PubMed] [Google Scholar]
- 21.Goldfarb DA, Diz DI, Tubbs RR, Ferrario CM, Novick AC. Angiotensin II receptor subtypes in the human renal cortex and renal cell carcinoma. J Urol. 1994;151:208–213. doi: 10.1016/s0022-5347(17)34918-2. [DOI] [PubMed] [Google Scholar]
- 22.Inwang ER, Puddefoot JR, Brown CL, et al. Angiotensin II type 1 receptor expression in human breast tissues. Br J Cancer. 1997;75:1279–1283. doi: 10.1038/bjc.1997.217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Kosugi M, Miyajima A, Kikuchi E, Horiguchi Y, Murai M. Angiotensin II type 1 receptor antagonist candesartan as an angiogenic inhibitor in a xenograft model of bladder cancer. Clin Cancer Res. 2006;12:2888–2893. doi: 10.1158/1078-0432.CCR-05-2213. [DOI] [PubMed] [Google Scholar]
- 24.Kosaka T, Miyajima A, Takayama E, et al. Angiotensin II type I receptor antagonist as an angiogenic inhibitor in prostate cancer. Prostate. 2007;67:41–49. doi: 10.1002/pros.20486. [DOI] [PubMed] [Google Scholar]
- 25.Egami K, Murohara T, Shimada T, et al. Role of host angiotensin II type 1 receptor in tumor angiogenesis and growth. J Clin Invest. 2003;112:67–75. doi: 10.1172/JCI16645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Koh WP, Yuan JM, van Den Berg D, Lee HP, Yu MC. Polymorphisms in angiotensin II type 1 receptor and angiotensin I converting enzyme genes and breast cancer risk among Chinese women in Singapore. Carcinogenesis. 2005;26:459–464. doi: 10.1093/carcin/bgh309. [DOI] [PubMed] [Google Scholar]
- 27.Uemura H, Hasumi H, Kawahara T, et al. Pilot study of angiotensin II receptor blocker in advanced hormone-refractory prostate cancer. Int J Clin Oncol. 2005;10:405–410. doi: 10.1007/s10147-005-0520-y. [DOI] [PubMed] [Google Scholar]
- 28.Kawahito Y, Kondo M, Tsubouchi Y, et al. 15-deoxy-delta (12,14)-PGJ(2) induces synoviocyte apoptosis and suppresses adjuvant-induced arthritis in rats. J Clin Invest. 2000;106:189–197. doi: 10.1172/JCI9652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Tsubouchi Y, Sano H, Kawahito Y, et al. Inhibition of human lung cancer cell growth by the peroxisome proliferator-activated receptor-γ agonists through induction of apoptosis. Biochem Biophys Res Commun. 2000;270:400–405. doi: 10.1006/bbrc.2000.2436. [DOI] [PubMed] [Google Scholar]
- 30.Dreyer C, Krey G, Keller H, Givel F, Helftenbein G, Wahli W. Control of the peroxisomal beta-oxidation pathway by a novel family of nuclear hormone receptors. Cell. 1992;68:879–887. doi: 10.1016/0092-8674(92)90031-7. [DOI] [PubMed] [Google Scholar]
- 31.Kliewer SA, Umesono K, Noonan DJ, Heyman RA, Evans RM. Convergence of 9-cis retinoic acid and peroxisome proliferator signalling pathways through heterodimer formation of their receptors. Nature. 1992;358:771–774. doi: 10.1038/358771a0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Kliewer SA, Forman BM, Blumberg B, et al. Differential expression and activation of a family of murine peroxisome proliferator-activated receptors. Proc Natl Acad Sci USA. 1994;91:7355–7359. doi: 10.1073/pnas.91.15.7355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Chinetti G, Griglio S, Antonucci M, et al. Activation of proliferator-activated receptors alpha and gamma induces apoptosis of human monocyte-derived macrophages. J Biol Chem. 1998;273:25573–25580. doi: 10.1074/jbc.273.40.25573. [DOI] [PubMed] [Google Scholar]
- 34.Altiok S, Xu M, Spiegelman BM. PPARgamma induces cell cycle withdrawal: inhibition of E2F/DP DNA-binding activity via down-regulation of PP2A. Genes Dev. 1997;11:1987–1998. doi: 10.1101/gad.11.15.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Keelan JA, Sato TA, Marvin KW, Lander J, Gilmour RS, Mitchell MD. 15-Deoxy-delta (12,14)-prostaglandin J(2), a ligand for peroxisome proliferator-activated receptor-gamma, induces apoptosis in JEG3 choriocarcinoma cells. Biochem Biophys Res Commun. 1999;262:579–585. doi: 10.1006/bbrc.1999.1257. [DOI] [PubMed] [Google Scholar]