

Abstract
In the bacterial ISC system for iron–sulfur cluster assembly, IscU acts as a primary scaffold protein, and the molecular co-chaperones HscA and HscB specifically interact with IscU to facilitate ATP-driven cluster transfer. In this work, cluster transfer from Azotobacter vinelandii [Fe2S2]2+ cluster-bound IscU to apo-Grx5, a general purpose monothiol glutaredoxin in A. vinelandii, was monitored by circular dichroism spectroscopy, in the absence and in the presence of HscA/HscB/Mg-ATP. The results indicate a 700-fold enhancement in the rate of [Fe2S2]2+ cluster transfer in the presence of the co-chaperones and Mg-ATP, yielding a second-order rate constant of 20 000 M–1 min–1 at 23 °C. Thus, HscA and HscB are required for efficient ATP-dependent [Fe2S2]2+ cluster transfer from IscU to Grx5. The results support a role for monothiol Grx’s in storing and transporting [Fe2S2]2+ clusters assembled on IscU and illustrate the limitations of interpreting in vitro cluster transfer studies involving [Fe2S2]-IscU in the absence of the dedicated HscA/HscB co-chaperone system.
The ubiquitous and essential IscU protein serves as the primary scaffold for cysteine desulfurase-mediated iron–sulfur cluster assembly in the ISC machinery for cluster biogenesis that is used by many bacteria and in eukaryotic mitochondria.1−3 Under reconstitution conditions, the initial product is a stable form containing one [Fe2S2]2+ cluster per homodimer, [Fe2S2]-IscU, which is slowly converted under strictly anaerobic conditions into a form containing one [Fe4S4]2+ cluster per homodimer, [Fe4S4]-IscU, via reductive coupling of two [Fe2S2]2+ clusters at the subunit interface.4,5In vivo studies have shown that IscU-type proteins have critical roles in the maturation of both [Fe2S2] and [Fe4S4] cluster-containing proteins under aerobic and anaerobic conditions.6 Taken together with recent structural evidence that the initial product of IscS/IscU cluster biosynthesis is an IscU monomer with a solvent-exposed [Fe2S2] cluster,3 there is little doubt that the stable [Fe2S2]-IscU dimer is physiologically relevant for initiating cellular [Fe2S2] cluster trafficking. Moreover, the bacterial isc operon also contains an essential and dedicated molecular co-chaperone system, HscA/HscB in bacteria (Ssq1/Jac1 in yeast mitochondria), which serves to specifically enhance the rate of [Fe2S2]2+ cluster transfer from the [Fe2S2]-IscU dimer to apo acceptor proteins such as Isc ferredoxin (Fdx) in an ATP-dependent reaction.7,8 However, it is currently unclear if IscU transfers clusters directly to acceptor proteins or via proposed intermediate cluster carrier proteins such as the A-type,9,10 Nfu-type,11,12 or monothiol glutaredoxins (Grx’s).13,14
In vivo studies in Saccharomyces cerevisiae have demonstrated an important role for Grx5 in Fe–S cluster biogenesis.15 Yeast Grx5 is a member of a ubiquitous and well-defined class of monothiol Grx’s with CGFS active sites that exhibit low glutathione-dependent thiol–disulfide oxidoreductase activity.14 Rather, 55Fe-radiolabeled immunoprecipitation studies have indicated a role in facilitating transfer of Fe–S clusters assembled on Isu1, a yeast homologue of IscU.16 Moreover, spectroscopic and structural studies have shown that monothiol Grx’s can bind subunit-bridging [Fe2S2]2+ clusters, ligated by the active-site cysteines of each monomer and two glutathiones, that can be rapidly transferred to physiologically relevant acceptor proteins.13,17,18 However, there is currently no direct evidence for [Fe2S2]2+ cluster transfer from IscU to apo monothiol Grx’s. In this work, we present direct spectroscopic evidence for rapid, ATP-driven, [Fe2S2]2+ cluster transfer from [Fe2S2]-IscU to apo-Grx5 in the presence of HscA and HscB using recombinant proteins from Azotobacter vinelandii. The results demonstrate the critical role that HscA and HscB play in facilitating cluster transfer from [Fe2S2]-IscU to monothiol Grx’s and suggest an important role for monothiol Grx’s in the trafficking of [Fe2S2]2+ clusters assembled on IscU. The experimental methods for expressing, purifying, and assaying the proteins used in this work and the protocols used for cluster transfer reactions are described in the Supporting Information.
The marked differences in the CD spectra of the [Fe2S2]2+ centers in A. vinelandii IscU and Grx5, red and blue spectra, respectively, in Figure 1, make this the method of choice for monitoring cluster transfer between these two proteins. No cluster transfer was observed from [Fe2S2]-Grx5 to apo-IscU over a period of 3 h using a 2-fold excess of [Fe2S2] clusters per apo-IscU dimer, in the presence of physiologically relevant levels of glutathione (3 mM). However, the reverse reaction involving cluster transfer from [Fe2S2]-IscU to apo-Grx5 does occur, albeit very slowly, in the presence of 3 mM glutathione. This is shown in Figure 1, which indicates ∼30% cluster transfer over a period of 3 h, using IscU containing 0.9 [Fe2S2]2+ cluster per homodimer and a 1.7-fold excess of dimeric Grx5 per IscU [Fe2S2] cluster. In contrast, in the presence of HscA, HscB, Mg-ATP, and KCl (required for optimal ATPase activity of HscA), the CD spectrum of [Fe2S2]-IscU is perturbed by binding to HscA and HscB, as previously observed,7 and the rate of cluster transfer from [Fe2S2]-IscU to apo-Grx5 is dramatically enhanced, going to completion within 6 min of initiating the reaction by the addition of Mg-ATP, see Figure 2.
Figure 1.
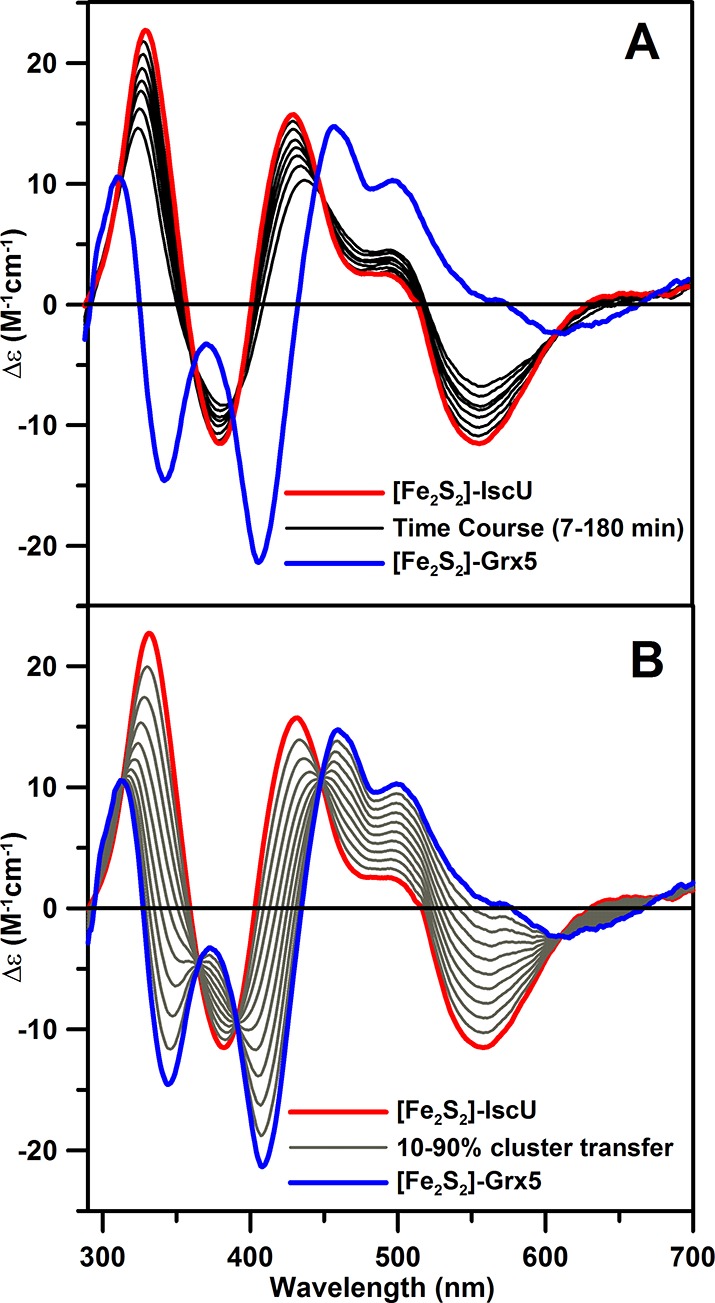
Time course of cluster transfer from A. vinelandii [Fe2S2]-IscU (45 μM in [Fe2S2]2+ clusters) to apo-Grx5 (150 μM in monomer) monitored by UV–visible CD spectroscopy at 23 °C. (A) CD spectra recorded at 0, 7, 20, 40, 60, 80, 120, and 180 min after adding [Fe2S2]-IscU to apo-Grx5 in 100 mM Tris-HCl buffer, pH 7.8, with 3 mM glutathione. (B) Simulated CD spectra corresponding to quantitative [Fe2S2]2+ cluster transfer from [Fe2S2]-IscU to apo-Grx5 in 10% increments. Δε values are based on the [Fe2S2]2+ cluster concentration, and the path length was 1 cm.
Figure 2.
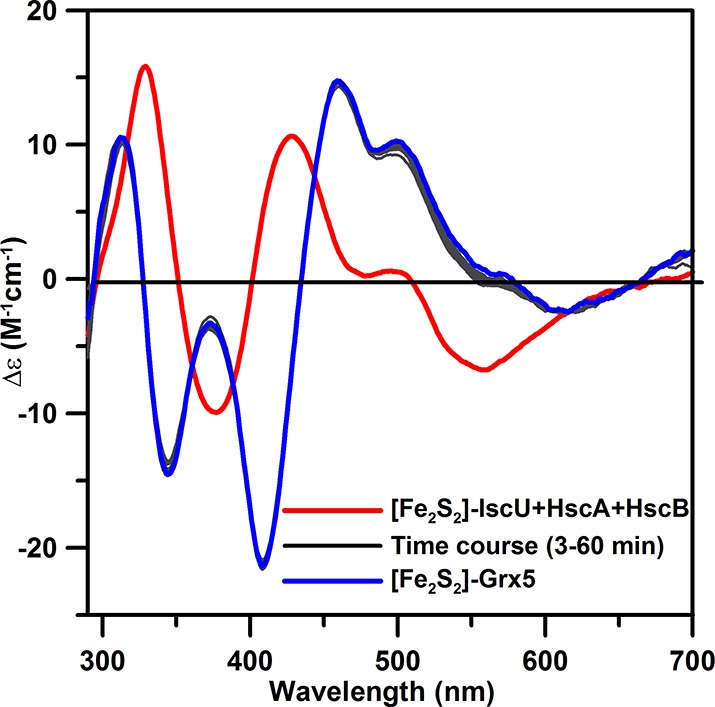
Time course of cluster transfer from A. vinelandii [Fe2S2]-IscU to apo-Grx in the presence of 0.10 mM A. vinelandii HscA and HscB, 40 mM MgCl2, 2 mM ATP, and 150 mM KCl monitored by UV–visible CD spectroscopy at room temperature. CD spectra were recorded at 3, 6, 10, 14, 18, 22, 26, 30, 40, 50, and 60 min after the addition of Mg-ATP to the reaction mixture. All other conditions are the same as described in Figure 1.
Rate constants for [Fe2S2]-IscU to apo-Grx5 cluster transfer in the absence and in the presence of the co-chaperones, Mg-ATP and KCl, were quantitatively assessed by fitting CD intensities as a function of time to second-order kinetics based on the initial concentrations of donor and acceptor, see Figure 3. The rate constant increases ∼700-fold, from 30 to 20 000 M–1 min–1, on addition of the co-chaperones, Mg-ATP and KCl. This is much greater than the ∼20-fold increase (from 36 to 800 M–1 min–1) in the rate of cluster transfer from [Fe2S2]-IscU to apo-IscFdx that occurs on addition of the same co-chaperones.7 This suggests that [Fe2S2]-IscU is unlikely to be the immediate [Fe2S2] cluster donor for maturation of IscFdx, which functions as an essential electron donor for ISC-mediated cluster assembly in A. vinelandii.19 Rather, as discussed below, IscFdx may receive [Fe2S2] clusters directly from Grx5.
Figure 3.

Comparison of the kinetics of cluster transfer from A. vinelandii [Fe2S2]-IscU to apo-Grx5 in the presence and in the absence of HscA/HscB/ATP. All conditions are the same as described in Figures 1 and 2. The data in the presence of HscA/HscB/ATP (■) were obtained by continuously monitoring the CD intensity at 460 nm after initiation of the reaction with Mg-ATP, and the solid line is a best-fit simulation to second-order kinetics with a rate constant of 20 000 M–1 min–1. The data in the absence of HscA/HscB/ATP (•) were obtained by monitoring the difference in the CD intensity at 457 and 408 nm, and the solid line is a best-fit simulation to second-order kinetics with a rate constant of 30 M–1 min–1.
The ability of Grx5 to rapidly and quantitatively accept [Fe2S2]2+ clusters from [Fe2S2]-IscU in the presence of the co-chaperones, in an ATP-dependent reaction, supports a physiological role for monothiol Grx’s in the trafficking of [Fe2S2]2+ clusters that are assembled on IscU. Monothiol Grx’s therefore have the capacity to store and/or deliver [Fe2S2]2+ clusters assembled on U-type scaffold proteins. Interestingly, slow and reversible [Fe2S2] cluster exchange via direct protein interaction between human Isu and Grx2, in the absence of the human Fe–S cluster biogenesis co-chaperone system (HSPA9 and HSC20)20,21 and Mg-ATP, has recently been reported by monitoring loss or gain in disulfide oxidoreductase activity and isothermal titration calorimetry.22 Although human Grx2 (CSYC active site) is a dithiol Grx and has not been implicated in Fe–S cluster biogenesis, it has been shown to exist in a mononuclear apo form with high disulfide oxidoreductase activity and a [Fe2S2]2+ cluster-bridged dimer, ligated by the first active-site cysteine of each Grx2 monomer and two glutathiones, which lacks disulfide oxidoreductase activity.23 In light of the stability of the cluster-bound form with respect to cluster transfer and sensitivity to oxidative stress, the [Fe2S2]2+ cluster on human Grx2 has been proposed to function as a sensor that responds to oxidative stress by activating the disulfide oxidoreductase activity via cluster degradation.24
A [Fe2S2]2+ cluster storage function for monothiol Grx’s may be required under Fe-replete conditions, and the extent of cluster loading may be an important sensor of the cellular Fe–S cluster status. This latter hypothesis is supported by the accumulating evidence that the Fe regulon in yeast is controlled by the extent of [Fe2S2] cluster-loading of the cytosolic Grx3 and Grx4 monothiol glutaredoxins.25−27 In S. cerevisiae, the sensing mechanism involves interaction of the [Fe2S2]2+ cluster-bound form of the Grx3 or Grx4 homodimer with a BolA-type protein, termed Fra2, to form a less labile [Fe2S2]2+ cluster-bound Grx3/4-Fra2 heterodimer that prevents accumulation of the Aft transcription factor in the nucleus, where it functions in activating Fe uptake systems. A related Fe or Fe–S cluster regulatory function may also occur in bacteria since a stable [Fe2S2]2+ cluster-bound Grx4/BolA heterodimer has been reported in Escherichia coli, where Grx4 is the sole monothiol Grx.28 An homologous BolA protein is also present in A. vinelandii. Alternatively, either the [Fe2S2]2+ cluster-bound monothiol Grx homodimer or the Grx-BolA heterodimer may serve to regulate Fe–S cluster biogenesis in bacteria by acting as the [Fe2S2]2+ cluster donor for IscR, which acts as a transcriptional repressor of the entire isc operon in its [Fe2S2]2+ cluster-bound form.29
In addition to a potential [Fe2S2]2+ cluster storage or sensing role for monothiol Grx’s, the available evidence for rapid cluster transfer to physiologically relevant acceptor proteins suggests a role as a delivery system for clusters assembled on primary scaffold proteins. This was first demonstrated in plant chloroplasts, in which [Fe2S2]2+ cluster-bound monothiol GrxS14 was found to rapidly and quantitatively transfer its [Fe2S2] cluster to apo plant Fdx with a second-order rate contant of 20 000 M–1 min–1 at 23 °C.11 Subsequently, cluster transfer from [Fe2S2]-Grx4 to apo-IscFdx in E. coli was demonstrated, although the rate constant was not determined.28 Based on the CD studies shown in Figure 4, intact and quantitative cluster transfer from [Fe2S2]-Grx5 to apo-IscFdx also occurs in A. vinelandii, with a second-order rate constant of 2100 M–1 min–1 at 23 °C. This rate constant is 2–3 times larger than that reported for co-chaperone-assisted [Fe2S2] cluster transfer from IscU (800 M–1 min–1 at 23 °C),7 indicating that Grx5 is a viable intermediate carrier protein for delivering [Fe2S2] clusters assembled on IscU to apo-IscFdx.
Figure 4.
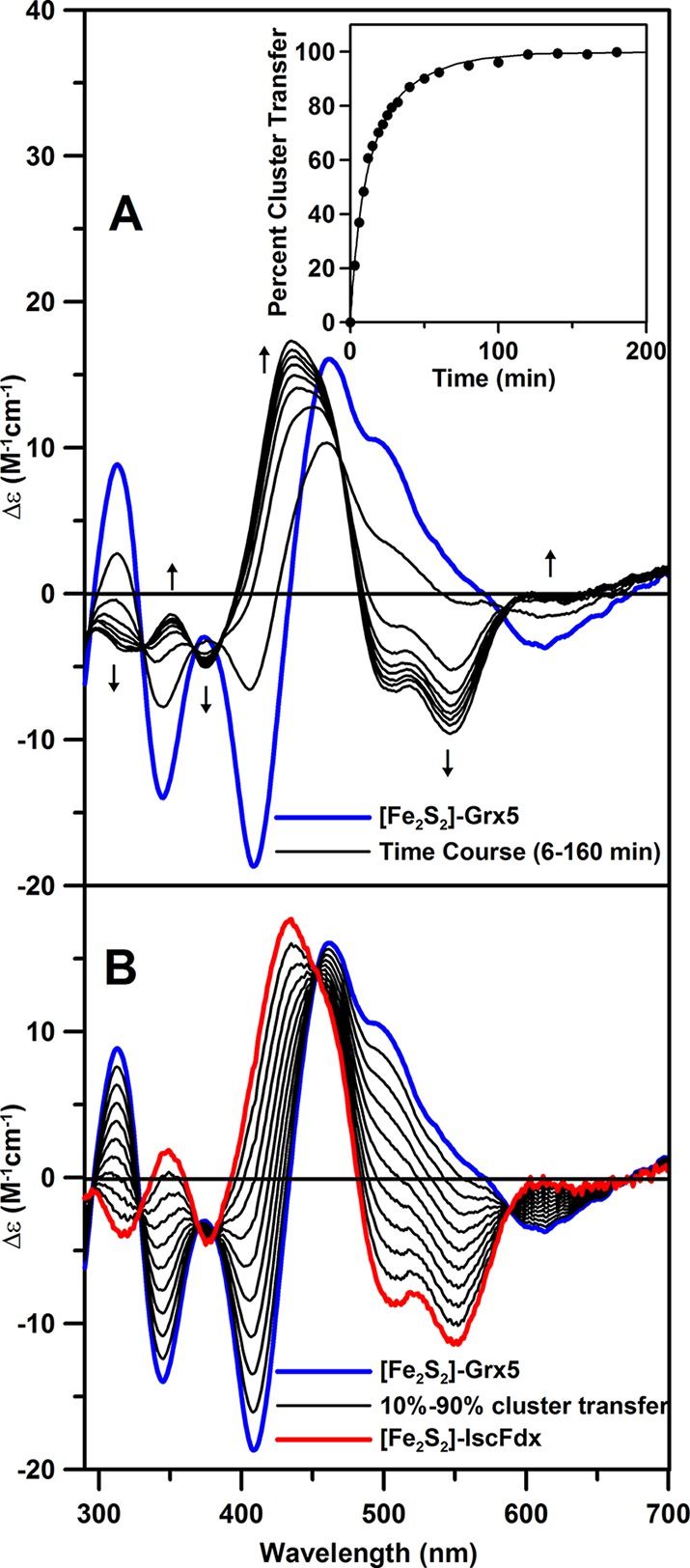
Time course of cluster transfer from A. vinelandii [Fe2S2]-Grx5 (32 μM in [Fe2S2]2+ clusters) to apo-IscFdx (48 μM) monitored by UV–visible CD spectroscopy at 23 °C . (A) CD spectra recorded at 6, 20, 40, 60, 80, 100, 120, and 160 min after adding [Fe2S2]-Grx5 to apo-IscFdx in 100 mM Tris-HCl buffer, pH 7.8, with 2 mM dithiothreitol. The arrows indicate the direction of intensity change with increasing time at selected wavelengths. Inset shows kinetic data for the cluster transfer measured at 434 nm, and the solid line is a best-fit simulation to second-order kinetics with a rate constant of 2100 M–1 min–1. (B) Simulated CD spectra corresponding to quantitative [Fe2S2]2+ cluster transfer from [Fe2S2]-Grx5 to apo-IscFdx in 10% increments. Δε values are based on the [Fe2S2]2+ cluster concentration, and the path length was 1 cm.
Much work needs to be done to identify specific cluster acceptor proteins for [Fe2S2]2+ cluster-bound forms of monothiol Grx’s. These could be other proposed carrier proteins such as A-type and Nfu-type proteins and/or specific apo Fe–S proteins and enzymes. Identifying the specificity of [Fe2S2] cluster-bound forms of monothiol Grx’s with respect to acceptor proteins is under active investigation in our laboratory. In addition, since IscU functions as a catalyst for Fe–S cluster assembly,30 it is clearly important to develop a robust in vitro catalytic system that includes the co-chaperones for investigating cluster assembly on target proteins using Fe(II) and cysteine as the primary substrates.
Acknowledgments
We thank Dr. Dennis Dean and co-workers for providing plasmids for the recombinant expression of A. vinelandii IscU, HscA, HscB, Grx5, and IscFdx. This work was supported by a grant from the NIH (GM62542 to M.K.J.)
Supporting Information Available
Experimental methods for expressing, purifying, and assaying the proteins used in this work and protocols used for cluster transfer reactions. This material is available free of charge via the Internet at http://pubs.acs.org.
Author Contributions
† P.S. and B.Z. contributed equally to this work.
The authors declare no competing financial interest.
Funding Statement
National Institutes of Health, United States
Supplementary Material
References
- Johnson D. C.; Dean D. R.; Smith A. D.; Johnson M. K. Annu. Rev. Biochem. 2005, 74, 247. [DOI] [PubMed] [Google Scholar]
- Lill R. Nature 2009, 460, 831. [DOI] [PubMed] [Google Scholar]
- Marinoni E. N.; de Oliveira J. S.; Nicolet Y.; Raulfs E. C.; Amara P.; Dean D. R.; Fontecilla-Camps J. C. Angew. Chem., Int. Ed. 2012, 51, 5439. [DOI] [PubMed] [Google Scholar]
- Agar J. N.; Krebs B.; Frazzon J.; Huynh B. H.; Dean D. R.; Johnson M. K. Biochemistry 2000, 39, 7856. [DOI] [PubMed] [Google Scholar]
- Chandramouli K.; Unciuleac M.-C.; Naik S.; Dean D. R.; Huynh B. H.; Johnson M. K. Biochemistry 2007, 46, 6804. [DOI] [PubMed] [Google Scholar]
- Mühlenhoff U.; Richter N.; Pines O.; Pierik A. J.; Lill R. J. Biol. Chem. 2011, 286, 41205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chandramouli K.; Johnson M. K. Biochemistry 2006, 45, 11087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vickery L. E.; Cupp-Vickery J. R. Crit. Rev. Biochem. Mol. Biol. 2007, 42, 95. [DOI] [PubMed] [Google Scholar]
- Ollagnier-de-Choudens S.; Sanakis Y.; Fontecave M. J. Biol. Inorg. Chem. 2004, 9, 828. [DOI] [PubMed] [Google Scholar]
- Vinella D.; Brochier-Armanet C.; Loiseau L.; Talla E.; Barras F. PLoS Genet. 2009, 5, e1000497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bandyopadhyay S.; Naik S.; O’Carroll I. P.; Huynh B. H.; Dean D. R.; Johnson M. K.; Dos Santos P. C. J. Biol. Chem. 2008, 283, 14092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Angelini S.; Gerez C.; Ollagnier-de-Choudens S.; Sanakis Y.; Fontecave M.; Barras F.; Py B. J. Biol. Chem. 2008, 289, 14084. [DOI] [PubMed] [Google Scholar]
- Bandyopadhyay S.; Gama F.; Molina-Navarro M. M.; Gualberto J. M.; Claxton R.; Naik S. G.; Huynh B. H.; Herrero E.; Jacquot J.-P.; Johnson M. K.; Rouhier N. EMBO J. 2008, 27, 1122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rouhier N.; Couturier J.; Johnson M. K.; Jacquot J. P. Trends Biochem. Sci. 2010, 35, 43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodríguez-Manzaneque M. T.; Tamarit J.; Bellí G.; Ros J.; Herrero E. Mol. Biol. Cell 2002, 13, 1109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mühlenhoff U.; Gerber J.; Richhardt N.; Lill R. EMBO J. 2003, 22, 4815. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Picciocchi A.; Saguez C.; Boussac A.; Cassier-Chauvat C.; Chauvat F. Biochemistry 2007, 46, 15018. [DOI] [PubMed] [Google Scholar]
- Iwema T.; Picciocchi A.; Traore D. A.; Ferrer J. L.; Chauvat F.; Jacquamet L. Biochemistry 2009, 48, 6041. [DOI] [PubMed] [Google Scholar]
- Johnson D. C.; Unciuleac M.-C.; Dean D. R. J. Bacteriol. 2006, 188, 7551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Uhrigshardt H.; Singh A.; Kovtunovych G.; Ghosh M.; Rouault T. A. Hum. Mol. Genet. 2010, 3816. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rouault T. A.; Tong W. H. Cell 2008, 24, 398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qi W.; Cowan J. A. Chem. Commun. 2011, 47, 4889. [DOI] [PubMed] [Google Scholar]
- Johansson C.; Kavanagh K. L.; Gileadi O.; Oppermann U. J. Biol. Chem. 2007, 282, 3077. [DOI] [PubMed] [Google Scholar]
- Lillig C. H.; Berndt C.; Vergnolle O.; Lonn M. E.; Hudermann C.; Bill E.; Holmgren A. Proc. Natl. Acad. Sci. U.S.A. 2005, 102, 8168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ojeda L.; Keller G.; Muhlenhoff U.; Rutherford J. C.; Lill R.; Winge D. R. J. Biol. Chem. 2006, 281, 17661. [DOI] [PubMed] [Google Scholar]
- Li H.; Mapolelo D. T.; Dingra N. N.; Naik S. G.; Lees N. S.; Hoffman B. M.; Riggs-Gelasco P. J.; Huynh B. H.; Johnson M. K.; Outten C. E. Biochemistry 2009, 48, 9569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li H.; Mapolelo D. T.; Dingra N. N.; Keller G.; Riggs-Gelasco P. J.; Winge D. R.; Johnson M. K.; Outten C. E. J. Biol. Chem. 2011, 286, 867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yeung N.; Gold B.; Liu N. L.; Prathapam R.; Sterling H. J.; Williams E. R.; Butland G. Biochemistry 2011, 50, 8957. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwartz C. J.; Giel J. L.; Patschkowski T.; Luther C.; Ruzicka F. J.; Beinert H.; Kiley P. J. Proc. Natl. Acad. Sci. U.S.A. 2011, 98, 14895. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonomi F.; Iametti S.; Ta D. T.; Vickery L. E. J. Biol. Chem. 2005, 280, 29513. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.