

Abstract
The unique mutualism between corals and their photosynthetic zooxanthellae (Symbiodinium spp.) is the driving force behind functional assemblages of coral reefs. However, the respective roles of hosts and Symbiodinium in this endosymbiotic association, particularly in response to environmental challenges (e.g., high sea surface temperatures), remain unsettled. One of the key obstacles is to produce and maintain aposymbiotic coral hosts for experimental purposes. In this study, a simple and gentle protocol to generate aposymbiotic coral hosts (Isopora palifera and Stylophora pistillata) was developed using repeated incubation in menthol/artificial seawater (ASW) medium under light and in ASW in darkness, which depleted more than 99% of Symbiodinium from the host within 4∼8 days. As indicated by the respiration rate, energy metabolism (by malate dehydrogenase activity), and nitrogen metabolism (by glutamate dehydrogenase activity and profiles of free amino acids), the physiological and biochemical performances of the menthol-induced aposymbiotic corals were comparable to their symbiotic counterparts without nutrient supplementation (e.g., for Stylophora) or with a nutrient supplement containing glycerol, vitamins, and a host mimic of free amino acid mixture (e.g., for Isopora). Differences in biochemical responses to menthol-induced bleaching between Stylophora and Isopora were attributed to the former digesting Symbiodinium rather than expelling the algae live as found in the latter species. Our studies showed that menthol could successfully bleach corals and provided aposymbiotic corals for further exploration of coral-alga symbioses.
Introduction
The unique mutualism between corals and their photosynthetic zooxanthellae (Symbiodinium spp.) underpins ecological success of corals in shallow and oligotrophic seawater. However, this association is highly vulnerable to rising seawater temperatures. A rise of only 1∼2°C above the summer average under moderate to high irradiance will likely be enough to disrupt the symbiotic relationships by causing the symbionts to be expelled from the host, precipitating so-called ‘coral bleaching’ [1], [2]. Coral bleaching events are known to further cause a breakdown [1]–[4] or phase shift [5]–[7] in coral reefs. These situations are predicted to worsen with time if the increase in seawater surface temperatures cannot be slowed [8], [9].
In order to understand if corals can survive the coming stressful environments, the mechanisms underlying coral bleaching have been intensively studied (reviewed in Weis [10]). It is widely accepted that reactive oxygen species (ROS) generated by Symbiodinium photoinhibition and/or mitochondrial dysfunction in the host can cause breakdown of the symbiotic association [10]–[12]. However, the comparative susceptibility of coral hosts and Symbiodinium to thermal stresses is not completely understood. In studies of symbionts, cultured and freshly isolated Symbiodinium (FIS) was widely used to explore the symbiont physiology. Different physiological performances, such as the photosynthesis capability under thermal stress, of FIS or cultured Symbiodinium were also revealed at the clade or subclade levels [13]–[16]. In contrast, studies on physiological responses of aposymbiotic coral hosts are limi'ted due to a lack of suitable protocols.
Several methods were used to deplete Symbiodinium from cnidarian hosts, including cold shock (e.g., 4°C) [17]–[19], a high seawater temperature (e.g., 33°C) [20], and 3-(3,4-dichlorophenyl)-1,1-dimethylurea (DCMU) treatment [21], but few of them generated healthy aposymbiotic coral hosts which could be used for further studies. Aposymbiotic corals induced by high seawater temperatures either take a long time and need antibiotics treatment [20] or result in high coral mortality [22]. High-temperature treatment might also implant a heat experience in corals which might influence the performance of bleached corals in thermal-tolerance studies. On the other hand, bleaching corals with DCMU requires high light intensities (e.g., 70% of ambient insolation) and large volumes of seawater (ca. 1000 L) to maintain the animals, which prevents laboratories without ample seawater supplies and outdoor facilities from conducting coral-bleaching experiments. Consequently, physiological and biochemical studies on aposymbiotic hosts in Symbiodinium-cnidarian symbioses are mostly confined to sea anemones [19], [23]–[26] and aposymbiotic larvae from limited coral species [27], [28]. Nevertheless there are still gaps in applying the knowledge obtained from sea anemones to corals when the coral skeleton, calcification, and surface and endoskeletal microbes should be taken into account [29]. Therefore, a general method needs to be developed to prepare as many species of aposymbiotic corals from adult individuals to conduct comparative analyses among coral species.
Menthol is a cyclic terpene alcohol which is usually used to anesthetize cnidarians in marine biological studies [30]. This compound was occasionally found to bleach Symbiodinium-associated corals and sea anemones during anesthetization (unpublished data). Despite menthol's toxicity to corals being unclear, menthol was found to be less toxic to an aquatic invertebrate (Daphnia magna) for which the 24-h 50% lethal concentration (LC50) is 37.7∼71.0 mg L−1 [31]. In this study, therefore, extant corals from two major lineages, respectively of robust and complex clades, were used to explore a workable procedure to prepare aposymbiotic corals from adult colonies. Furthermore, the physiological and biochemical performances of the aposymbiotic coral hosts were examined, and their comparability to their symbiotic counterparts was evaluated. Feeding an artificial diet was also used to examine the effect of exogenous nutrients on maintaining physiological and biochemical performances of aposymbiotic coral compared to their symbiotic counterparts.
Materials and Methods
Experimental organisms
Isopora palifera (robust clade) and Stylophora pistillata (complex clade) were respectively collected from 3 and 7 m in depth within Kenting National Park, Taiwan (21°55′54″N, 120°44′45″E) between October 2010 and November 2011. Coral colonies were transferred to the laboratory within 3 h in an aerated plastic box, and maintained in an aquarium (90×45×45 cm) equipped with illumination [12: 12-h light-dark regime and ca. 50 µmol photons m−2s−1 photosynthetically active radiation (PAR)], temperature control (25°C), filtration (EHEIM, Germany), and a protein skimmer. Corals were acclimatized to laboratory conditions for 1 week before conducting the experiments.
Bleaching coral in menthol-artificial seawater (ASW) and feeding trial
Menthol-induced coral bleaching was examined by incubating an Isopora fragment (ca. 5×5 cm in size) in a crystallizing dish (125×65 mm, Corning, Kaiserslautern, Germany) containing 300 ml menthol supplemented with ASW (Instant Ocean, Aquarium Systems, Sarrebourg Cedex, France) with aeration and under standard illumination as described above at 25°C. The menthol/ASW medium was prepared by diluting a 20% (w/v) menthol stock (in ethanol) with ASW and was used to bleach Isopora at concentrations of 0.19, 0.38, and 0.58 mM. Released Symbiodinium was collected by centrifuging the medium at 860×g for 3 min. The bleaching test was stopped when the coral tissue began to shrink, and the remaining Symbiodinium alga in Isopora was collected by air-blasting and centrifugation as described in a previous paper [32]. Numbers of Symbiodinium cells collected were counted with a Neubauer improved hemocytometer (Marienfeld Superior, Lauda-Königshofen, Germany) to determine the coral bleaching rate. Two nutrient cocktails (A and B) were used to feed bleached Isopora to test if nutrient supplementation was necessary to maintain the physiological and biochemical performance comparable to their symbiotic counterparts. The common supplements for nutrient A and B were 200 µg ml−1 cobalamin, 4 µg ml−1 biotin, and 10% glycerol. The amino acid supplement (see details in Table 1) to nutrient A was 10.5 mM of a free amino acid (FAA) mixture, an FAA pool mimic of that in the Isopora tissue, and that to nutrient B was a 10.5 mM so-called ‘essential’ FAA mixture. The 10% glycerol supplement was used to provide the coral host with organic carbon and also to increase the supplement viscosity, such that the nutrient cocktail would remain on the coral surface for awhile.
Table 1. Mole percentages of amino acids in the nutrient supplements for maintaining aposymbiotic Isopora palifera.
| Amino acid | Free amino acid supplement (mol %) | |
| Nutrient A | Nutrient B | |
| Aspartate | 4.2 | - |
| Glutamate | 13.5 | - |
| Asparagine | 2.0 | - |
| Serine | 2.9 | - |
| Histidine | 6.0 | 10.0 |
| Glutamine | 4.5 | - |
| Glycine | 1.9 | - |
| Threonine | 10.9 | 10.0 |
| Arginine | 5.7 | - |
| Taurine | 1.4 | - |
| Alanine | 4.2 | - |
| Tyrosine | 10.3 | 10.0 |
| γ-Aminobutyric acid | 2.2 | - |
| Tryptophane | 0.9 | 10.0 |
| Methionine | 1.5 | 10.0 |
| Valine | 5.0 | 10.0 |
| Phenylalanine | 2.7 | 10.0 |
| Isoleucine | 3.4 | 10.0 |
| Leucine | 3.9 | 10.0 |
| Lysine | 12.8 | 10.0 |
| Total | 100.0 | 100.0 |
Determination of physiological and biochemical indices
The respiration rate was used to represent the general physiological performance of symbiotic and aposymbiotic coral hosts, which was determined in a custom-made respiration chamber (400 ml) which was connected with a BOD probe (YSI 5905, Yellow Springs, OH, USA) and a dissolved oxygen (DO) meter (YSI 52). Oxygen consumption by the coral host in the respiration chamber was continuously determined by connecting the meters to a personal computer for 15 min in darkness. The respiration rate of the coral host per se in symbiosis was determined by subtracting the dark respiration rate of an equivalent amount of Symbiodinium in the whole symbiotic consortium from the total oxygen consumption by the symbiotic coral. The dark respiration rate of Symbiodinium was determined with freshly isolated algae in a Hansatech Oxygraph System (Hansatech Instrument, Norfolk, UK).
Biochemical indices of the coral host were determined with the apparent activities of malate dehydrogenase (MDH) and glutamate dehydrogenase (GDH), and the FAA profile. MDH, one of the key enzymes in energy anabolism [33], was used to represent the energy-synthesizing capacity of the coral host. GDH and the composition of FAAs are two key factors which are usually used to reveal the nitrogen status of Symbiodinium-associated corals and sea anemone hosts [25], [34]. To prepare the host homogenate, a Stylophora branch of about 10 cm or an area of 25 cm2 of Isopora was stripped of tissues with 4°C seawater buffer [25] carried by air blasting. The resulting tissue slurry stored on ice was homogenized in a syringe and then centrifuged at 21,500×g for 10 min (4°C) to remove cell debris and Symbiodinium. The enzyme extract was immediately stored at −80°C and analyzed within 3 days. MDH activity was determined by adding 100 µl of host homogenate to 1 ml of the reaction mixture containing 80 mM imidazole-HCl buffer (pH 7.0), 100 mM KCl, 0.3 mM oxaloacetate, and 0.15 mM NADH at room temperature. The activity of GDH was determined in the aminating direction, in which 100 µl of host homogenate was incubated with 1 ml of reaction mixture containing 50 mM HEPES buffer at pH 7.4, 0.2 mM NADPH, and 10 mM α-ketoglutarate at room temperature. The enzyme activity was measured by the decrease in absorbance at 340 nm, and was expressed as nmol NAD(P)+ formed per milligram tissue protein per minute. Co-incubation of the host extract and reaction mixture without substrate (oxaloacetate for MDH and α-ketoglutarate for GDH) was used as a blank to confirm the specificity of the enzyme reaction, which reduced the activity by over 95%. Prior to determining FAAs in the host homogenate, samples were precipitated with 70% ethanol. FAAs in the resulting ethanol extract were quantified by a reverse-phase high-performance liquid chromatographic (HPLC) system using a pre-column derivatization method modified from Jones et al. [35]. For the HPLC analysis, the amino acids were derivatized with o-phthaldialdehyde and separated with a solvent delivery system (Hitachi L-2130, Tokyo, Japan), using a C18-ultrasphere column and fluorescence detector (Hitachi L-2485). The reference amino acid mixture was AA-S-18 (Sigma) supplemented with asparagine, glutamine, γ-aminobutyric acid (GABA), trypophan, and taurine. It was noted that cysteine and proline could not be detected by this technique. The protein content in the host homogenate was quantified by a protein assay kit of Bio-Rad Chemical (Hercules, CA, USA), following the manufacturer's instruction for the microassay with 0∼12 µg bovine serum albumin as the standard.
Statistical analysis
Data in this study are presented as the mean±S.E. from numbers of different colonies. Comparisons of the host respiration rates, MDH and GDH activities, and FAA contents in the host homogenate between symbiotic and aposymbiotic coral were made using a one-way analysis of variance (ANOVA) followed by Fisher's least significance difference (LSD) test for multiple comparisons at a significance level of 0.05. For the similarity analysis of the FAA composition between symbiotic and aposymbiotic coral hosts, the mole % data of FAAs were arc cosine-transformed to meet the normality and homogeneity of variance assumptions. The similarity of the FAA compositions was compared using multidimensional scaling (MDS) ordination [36], [37]. An analysis of similarity (ANOSIM) was used to determine whether FAA profiles from different treatments separated by MDS ordination significantly differed [38]. The analyses were carried out using the computer package, PRIMER 6 [38].
Results
When I. palifera was incubated in menthol-supplemented ASW, the Symbiodinium density in the coral declined with an increase in the incubation time, and rates of algal depletion were dependent on the concentration of menthol used (Fig. 1A). In order to examine the dose response of menthol, the decline in Symbiodinium density with time was converted to an equation using a curve-fitting model provided by SigmaPlot. The equation that best fit as determined by r
2 values was: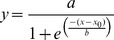 . Parameters of the equations for 0.19, 0.38, and 0.58 mM menthol treatment were: a = 101.22, b = −7.37, and x0 = 42.24 (r
2 = 0.979); a = 102.08, b = −7.08, and x0 = 29.65 (r
2 = 0.875); and a = 104.43, b = −3.79, and x0 = 12.71 (r
2 = 0.934), respectively. Using this equation, the times for 50% coral bleaching at different menthol concentrations were estimated and regressed on the menthol concentration used. As shown in Fig. 1B, the time for 50% coral bleaching was significantly correlated with the menthol concentration used (p<0.0001), and the correlation was fit to the linear regression equation: y = 59.11–78.76x (r
2 = 0.983). Although 0.58 mM menthol could bleach Isopora comparatively rapidly, continuous incubation at that concentration for 24 h always caused high (>80%) mortality. In order to obtain a rapid and gentle bleaching procedure, the duration of menthol treatment was reduced to 8 h following by 16 h of resting in an aquarium without menthol, and the mortality rate was significantly reduced in this way. With the protocol described in Fig. 2, 4 repeats of the above treatment/resting cycle could expel almost all Symbiodinium from Isopora and Stylophora (see as Fig. 3) within 4∼8 days after being maintained in an aquarium without menthol, which resulted in respective 0% and <10% mortalities in aposymbiotic Stylophora and Isopora preparations. It was also found that Isopora and Stylophora released Symbiodinium in different modes during menthol treatment. Symbiodinium released by menthol-treated Isopora was in a cloudy suspension and retained some PSII activity (Fv/Fm = 0.3∼0.5), but that from menthol-treated Stylophora aggregated into black granules which displayed no detectable PSII activity. When coral was bleached, a nutrient cocktail was fed from day 5 for aposymbiotic Isopora, but aposymbiotic Stylophora was not fed due to its physiological and biochemical performances being comparable to its symbiotic counterpart (see below). As shown in Fig. 3, the aposymbiotic and symbiotic Isopora and Stylophora displayed comparably healthy shapes to each other.
. Parameters of the equations for 0.19, 0.38, and 0.58 mM menthol treatment were: a = 101.22, b = −7.37, and x0 = 42.24 (r
2 = 0.979); a = 102.08, b = −7.08, and x0 = 29.65 (r
2 = 0.875); and a = 104.43, b = −3.79, and x0 = 12.71 (r
2 = 0.934), respectively. Using this equation, the times for 50% coral bleaching at different menthol concentrations were estimated and regressed on the menthol concentration used. As shown in Fig. 1B, the time for 50% coral bleaching was significantly correlated with the menthol concentration used (p<0.0001), and the correlation was fit to the linear regression equation: y = 59.11–78.76x (r
2 = 0.983). Although 0.58 mM menthol could bleach Isopora comparatively rapidly, continuous incubation at that concentration for 24 h always caused high (>80%) mortality. In order to obtain a rapid and gentle bleaching procedure, the duration of menthol treatment was reduced to 8 h following by 16 h of resting in an aquarium without menthol, and the mortality rate was significantly reduced in this way. With the protocol described in Fig. 2, 4 repeats of the above treatment/resting cycle could expel almost all Symbiodinium from Isopora and Stylophora (see as Fig. 3) within 4∼8 days after being maintained in an aquarium without menthol, which resulted in respective 0% and <10% mortalities in aposymbiotic Stylophora and Isopora preparations. It was also found that Isopora and Stylophora released Symbiodinium in different modes during menthol treatment. Symbiodinium released by menthol-treated Isopora was in a cloudy suspension and retained some PSII activity (Fv/Fm = 0.3∼0.5), but that from menthol-treated Stylophora aggregated into black granules which displayed no detectable PSII activity. When coral was bleached, a nutrient cocktail was fed from day 5 for aposymbiotic Isopora, but aposymbiotic Stylophora was not fed due to its physiological and biochemical performances being comparable to its symbiotic counterpart (see below). As shown in Fig. 3, the aposymbiotic and symbiotic Isopora and Stylophora displayed comparably healthy shapes to each other.
Figure 1. Bleaching Isopora palifera with menthol.

(A) Decrease in the Symbiodinium density in treated coral with different menthol concentrations; (B) correlation between the menthol concentration and the time for 50% coral bleaching.
Figure 2. Flow diagram of the preparation of aposymbiotic Isopora palifera and Stylophora pistillata.

Figure 4. Respiration rates of symbiotic and menthol-bleached Isopora palifera and Stylophora pistillata.

Aposymbiotic I. palifera with feeding was treated by supplementing animals with a nutrient cocktail containing glycerol, vitamins, and a host mimic free amino acid mixture as described in the [text/Table 1??]. Data represent the mean±S.E.
Figure 3. Polyps of symbiotic and aposymbiotic corals under microscopic examination.

(A) and (C) are Stylophora pistillata; (B) and (D) are Isopora palifera.
The extents of physiological and biochemical comparability between symbiotic and aposymbiotic corals were further examined. In this study, the term, aposymbiotic host, represents freshly bleached corals which were examined at 6∼10 days after menthol treatment. When comparing respiration rates, as shown in Fig. 4, those of the aposymbiotic hosts were 12.5±1.1 nmol min−1cm−2 (n = 5) for Isopora and 9.0±1.2 nmol min−1cm−2 (n = 5) for Stylophora. These data did not significantly differ from their symbiotic counterparts [10.3±0.5 nmol min−1cm−2 (n = 7) for Isopora, F1,11 = 3.996, p>0.05; and 9.0±1.1 nmol min−1cm−2 (n = 9) for Stylophora, F1,12 = 0.000, p>0.05]. Feeding aposymbiotic Isopora and Stylophora with the nutrient cocktail did not produce significant differences between the symbiotic and aposymbiotic corals (data not shown).
Biochemical indices (MDH, GDH, and the FAA pool) in the host homogenate were further examined. As shown in Table 2, GDH activity, total FAAs, and “essential” FAAs in Isopora were significantly reduced by 50.0% , 44.7%, and 43.7%, respectively, after bleaching (p<0.05). However, depletion of Symbiodinium produced no difference in MDH activities between the symbiotic and aposymbiotic Isopora (p>0.05). “Essential” FAAs noted here followed the definition applied to the sea anemone Aiptasia pulchella [19]. Levels of GDH and FAAs (total and essential) in aposymbiotic Isopora could be reverted to comparable levels of the symbiotic counterpart by feeding with nutrient A. However, feeding with nutrient B (containing a mixture of essential FAAs) was less effective than nutrient A in reverting GDH and FAA levels back to those of the symbiotic counterpart. Total FAA content in nutrient B supplemented-aposymbiotic Isopora reverted to about 73% of that in the symbiotic counterpart, but the value was still significantly lower (p<0.01) than that from the symbiotic counterpart. On the contrary, nutrient B supplementation reverted levels of GDH and essential FAAs in aposymbiotic Isopora to levels comparable to the symbiotic counterpart (p>0.05). Inconsistent with Isopora, levels of GDH activity, and total and essential FAAs displayed no significant (p>0.05) differences between symbiotic and aposymbiotic Stylophora (Table 3). However, as shown in Table 3, depletion of Symbiodinium from Stylophora still caused a significant increase in host MDH activity by 49.5% (p<0.005). Samples from aposymbiotic Stylophora with feeding were not further examined because data in Table 3 suggested that feeding might not be necessary.
Table 2. Contents of free amino acids and activities of malate dehydrogenase (MDH) and glutamate dehydrogenase (GDH) in tissue homogenates of symbiotic and bleached Isopora palifera with or without nutrient supplementation.
| Treatment | MDH | GDH | Free amino acids | |
| Total | Essential | |||
| (nmole NAD(P)+ min−1mg−1) | (pmole mg−1) | |||
| Symbiotic control | 77±18a (14) | 40±6a (13) | 385±43a (9) | 103±15a (9) |
| Apo-symbiotic host | 86±6a (9) | 20±4b (9) | 213±21b (11) | 58±8b (11) |
| Apo-symbiotic host fed nutrient A | 109±26a (5) | 24±4ab (5) | 372±29a (5) | 94±11a (5) |
| Apo-symbiotic host fed nutrient B | 42±9a (11) | 41±5a (8) | 281±34b (11) | 80±8ab (11) |
| F3,35 = 2.331 P>0.05 | F3,31 = 3.292 P<0.05 | F3,32 = 5.864 P<0.01 | F3,32 = 3.264 P<0.05 | |
Essential amino acids followed the definition applied to the sea anemone Aiptasia pulchella [19]. Enzyme activities were determined as the amount of NAD(P)H (in nmol) converted to NAD(P) by 1 mg of protein in 1 min. Nutrient compositions of A and B are described in “Materials and Methods” and Table 1. Numbers in parentheses represent the number of colony replicates, and means followed by the same letter do not significantly differ at p = 0.05 (Fisher's least significance difference test). Data are the mean±S.E.
Table 3. Contents of free amino acids and activities of malate dehydrogenase (MDH) and glutamate dehydrogenase (GDH) in tissue homogenates of symbiotic and bleached Stylophora pistillata.
| Treatment | MDH | GDH | Free amino acids | |
| Total | Essential | |||
| (nmole NAD(P)+ min−1mg−1) | (pmole mg−1) | |||
| Symbiotic control | 46±11 (12) | 21±1 (12) | 250±20 (12) | 75±12 (12) |
| Apo-symbiotic host | 91±6 (18) | 21±2 (20) | 271±11 (21) | 95±8 (21) |
| F1,28 = 12.948 p<0.001 | F1,30 = 0.026 p>0.05 | F1,31 = 0.004 p>0.05 | F1,31 = 2.231 p>0.05 | |
Essential amino acids followed the definition applied to the sea anemone Aiptasia pulchella [19]. Enzyme activities were determined as the amount of NAD(P)H (in nmol) converted to NAD(P) by 1 mg of protein in 1 min. Numbers in parentheses represent the number of colony replicates, and data are the mean±S.E.
A comparative analysis of FAAs indicated that dominant FAAs in the host homogenate of Isopora were glutamate, glycine, threonine, arginine, and lysine, which comprised >50 mol% of the total detected FAAs. When Symbiodinium was depleted from the host, threonine was the dominant FAA in the aposymbiotic host (25.6±4.7 mole%) and nutrient B-supplemented aposymbiotic host (20.4±3.0 mole%). The dominant FAA in the host homogenate of nutrient A-supplemented aposymbiotic Isopora was arginine (21.3±1.9 mole%) instead of the amino acids mentioned above. However, the dominant FAA in the host homogenate of Stylophora was aspartate for both the symbiotic (29.5±1.4 mole%) and aposymbiotic hosts (28.7±1.0 mole%). The variance analysis of the FAA composition further indicated that FAA profiles in the host homogenate significantly differed between symbiotic and aposymbiotic Isopora (with or without nutrient supplementation) (ANOSIM test, global R = 0.315, p<0.01, Fig. 5A), but no differences were found between symbiotic and aposymbiotic Stylophora (ANOSIM test, global R = 0.076, p>0.05, Fig. 5B). Pair-wise comparisons between different treatments in Isopora indicated a significant separation of FAA profiles in symbiotic coral from those in aposymbiotic coral (R = 0.205, p<0.01) and aposymbiotic coral supplemented with nutrient B (R = 0.466, p<0.001). However, the FAA profile in symbiotic Isopora could not be separated from that in the aposymbiotic counterpart supplemented with nutrient A (R = 0.122, p>0.05).
Figure 5. Multi-dimensional scaling (MDS) ordination of arc cosine-transformed free amino acid concentrations (mole%) in tissue extracts from symbiotic, bleached, and bleached coral with nutrient supplementation.

(A) Isopora palifera (stress = 0.11); (B) Stylophora pistillata. (stress = 0.14). Data labels represent the treatment for coral (1, symbiotic control; 2, bleached coral; 3, bleached coral with nutrient A supplement; 4, bleached coral with nutrient B supplement).
Discussion
In this study, we applied menthol to develop a simple and gentle protocol to prepare aposymbiotic corals which retained comparable physiological and biochemical performances to their symbiotic counterparts by incubation in seawater only (for Stylophora) or with additional feeding of a nutrient cocktail containing glycerol, vitamins, and a host mimic FAA mixture (for Isopora). Bleaching coral by menthol, as indicated in Fig. 1B, occurred in a significant dose-dependent manner. However, because continuous incubation always caused high mortality, a repeated 8: 16-h menthol (treatment): ASW (resting) treatment cycle was essential for the success of the protocol (Fig. 2).
Menthol is a compound known to act on a variety of different membrane receptors, including the transient receptor potential (TRP)M8, TRPA1, and other ionotrophic receptors [39]. The binding of menthol to TRPM8 results in an increase in intracellular Ca2+ concentrations and causes a cold sensation in vertebrates [40]–[43]. Menthol was also found to cause antinociceptive and local anesthetic effects in neuronal and skeletal muscles via blocking voltage-operated sodium channels [44]. Menthol is also known to cause many adverse effects to plants, including photoinhibition [45]. In Symbiodinium-associated corals, the mechanism underpinning menthol-induced coral bleaching is not clear. However, based on two different Symbiodinium-releasing modes (ejecting the alga in a cloudy suspension by Isopora and releasing digested alga by Stylophora), the bleaching mechanism might be attributed to Ca2+-triggered exocytosis as described by Pang and Südhof [46] and/or photoinhibition in Symbiodinium. We have no information about Ca2+ movements in the coral host during menthol treatment, but a preliminary study indicated that menthol might inhibit Symbiodinium photosynthesis II activity in the millimolar range (4-h IC50 of 0.72∼1.96 mM) which was at a similar level that caused coral bleaching (unpublished data). Further studies are needed to clarify the mechanism of menthol-induced coral bleaching.
When depleting Symbiodinium from a cnidarian host, a cessation in the supply of photosynthate released from the algal symbiont would greatly upset the host physiology and metabolism. Although respiration rates of some corals (Montastraea annularis, Agarwia lamarcki, Porites compressa, and Montipora capitata) decreased when Symbiodinium algae were depleted [47], [48], those of freshly bleached Isopora and Stylophora did not significantly differ from the symbiotic counterparts (Fig. 4). No differences in respiration rates between symbiotic and aposymbiotic corals were found in the temperate coral Astrangia danae, which was interpreted as holozoic feeding in the aposymbiotic coral possibly compensating for the energy loss from the deprivation of photosynthate release by Symbiodinium [49]. Because no food sources are available in ASW, energy sources for the bleached Stylophora and Isopora to balance the loss from lack of photosynthate release by Symbiodinium might be derived from consuming previous reserves or digestion of impaired Symbiodinium.
Depletion of symbiotic algae would also result in significant changes in nitrogen metabolism of the host [25], [34], [50]. For example, GDH, a key enzyme for assimilating (or releasing) ammonium into (or from) amino acids, increases in alga-depleted corals and sea anemones [34]. FAAs, especially the so-called essential amino acids, in the host homogenates were also found to have decreased by nearly half after depletion of symbiotic algae [25], [50]. In this study, the responses of coral nitrogen metabolism to algal depletion differed between Stylophora and Isopora. Algal depletion caused significant decreases in Isopora GDH activity and FAA contents but not in Stylophora. However, supplementation of the aposymbiotic Isopora with nutrients containing glycerol, a host mimic FAA mixture, and vitamins reverted the nitrogen metabolic indices back to a level and composition comparable to the symbiotic counterpart (Table 2, Fig. 5A). This result is similar to previous findings in Aiptasia [25], [50]. Therefore, aposymbiotic coral generated by expelling Symbiodinium alive during bleaching would need to be fed a proper nutrient supplement before being subjected to physiological studies. With the nutrient A supplement, we successfully maintained Isopora for the test of reinfection with heterogenic Symbiodinium (unpublished data).
In summary, comparisons of physiological performances and gene expression profiles between different species of coral hosts per se will be available by preparing freshly bleached aposymbiotic coral with the menthol protocol combined with nutrient supplementation if necessary. This technique will also potentially benefit the search for a generalist coral to re-establish symbiosis with different heterogenic Symbiodinium, which will make the contributions of different Symbiodinium subclades to coral symbiosis more straightforward.
Acknowledgments
The authors would like to thank members of the Coral Reef Evolutionary, Ecology and Genetics (CREEG) Group, Biodiversity Research Center, Academia Sinica (BRCAS) for field support. This is CREEG-BRCAS contribution no. 83.
Funding Statement
Academic Sinica Thematic grant (2008–2010) to JTW and CAC, and National Science Council (NSC-96-2628-B-001-004-MY3) to CAC. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Fitt WK, Brown BE, Warner ME, Dunne RP (2001) Coral bleaching: interpretation of thermal tolerance limits and thermal thresholds in tropical corals. Coral Reefs 20: 51–65. [Google Scholar]
- 2. Lesser MP, Farrell JH (2004) Exposure to solar radiation increases damage to both host tissues and algal symbionts of corals during thermal stress. Coral Reefs 23: 367–377. [Google Scholar]
- 3. Brown BE (1997) Coral bleaching: causes and consequences. Coral Reefs 16: S129–S138. [Google Scholar]
- 4. Hoegh-Guldberg O (1999) Climate change, coral bleaching and the future of world's coral reefs. Marine and Freshwater Research 50: 839–866. [Google Scholar]
- 5. Hughes TP (1994) Catastrophes, phase-shifts and large-scale degradation of Caribbean reef. Science 265: 1547–1551. [DOI] [PubMed] [Google Scholar]
- 6. Chen CA, Dai C-F (2004) Local phase shift from Acropora-dominant to Condylactis-dominant community in the Tiao-Shi Reef, Kenting National Park, southern Taiwan. Coral Reefs 23: 508–508. [Google Scholar]
- 7. Hughes TP, Rodrigues MJ, Bellwood DR, Ceccarelli D, Hoegh-Guldberg O, et al. (2007) Phase shifts, herbivory, and the resilience of coral reefs to climate change. Current Biology 17: 360–365. [DOI] [PubMed] [Google Scholar]
- 8. Hoegh-Guldberg O, Mumby PJ, Hooten AJ, Steneck RS, Greenfield P, et al. (2007) Coral Reefs Under Rapid Climate Change and Ocean Acidification. Science 318: 1737–1742. [DOI] [PubMed] [Google Scholar]
- 9. Carpenter KE, Abrar M, Aeby G, Aronson RB, Banks S, et al. (2008) One-Third of Reef-Building Corals Face Elevated Extinction Risk from Climate Change and Local Impacts. Science 321: 560–563. [DOI] [PubMed] [Google Scholar]
- 10. Weis VM (2008) Cellular mechanisms of cnidarian bleaching: stress causes the collapse of symbiosis. Journal of Experimental Biology 211: 3059–3066. [DOI] [PubMed] [Google Scholar]
- 11. Downs CA, Fauth JE, Halas JC, Dustan P, Bemiss J, et al. (2002) Oxidative stress and seasonal coral bleaching. Free Radical Biology and Medicine 33: 533–543. [DOI] [PubMed] [Google Scholar]
- 12. Smith DJ, Suggett DJ, Baker NR (2005) Is Photoinhibition of zooxanthellae photosynthesis the primary cause of thermal bleaching in corals? Global Change Biology 11: 1–11. [Google Scholar]
- 13. Bhagooli R, Hidaka M (2003) Comparison of stress susceptibility of in hospite and isolated zooxanthellae among five coral species. Journal of Experimental Marine Biology and Ecology 291: 181–197. [Google Scholar]
- 14. Tchernov D, Gorbunov MY, Vargas Cd, Yadav SN, Milligan AJ, et al. (2004) Membrane lipids of symbiotic algae are diagnostic of sensitivity to thermal bleaching in corals. Proceedings of the National Academy of Sciences of the United States of America 101: 13531–13535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Robison JD, Warner ME (2006) Differential impacts of photoacclimation and thermal stress on the photobiology of four different phylotypes of Symbiodinium (Pyrrhophyta). Journal of Phycology 42: 568–579. [Google Scholar]
- 16. Wang J-T, Meng P-J, Chen Y-Y, Chen CA (2012) Determination of the thermal tolerance of Symbiodinium using the activation energy for inhibiting photosystem II activity. Zoological Studies 51: 137–142. [Google Scholar]
- 17. Steen RG, Muscatine L (1987) Low temperature evokes rapid exocytosis of symbiotic algae by a sea anemone. Biological Bulletin 172: 246–263. [Google Scholar]
- 18. Wang J-T, Douglas AE (1997) Nutrients, signals, and photosynthate release by symbiotic algae: the impact of taurine on the dinoflagellate alga Symbiodinium from the sea anemone Aiptasia pulchella . Plant Physiology 114: 631–636. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Wang J-T, Douglas AE (1999) Essential amino acid synthesis and nitrogen recycling in an alga–invertebrate symbiosis. Marine Biology 135: 219–222. [Google Scholar]
- 20. Coffroth MA, Poland DM, Petrou EL, Brazeau DA, Holmberg JC (2010) Environmental Symbiont Acquisition May Not Be the Solution to Warming Seas for Reef-Building Corals. PLoS ONE 5: e13258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Jones RJ (2004) Testing the ‘photoinhibition’ model of coral bleaching using chemical inhibitors. Marine Ecology Progress Series 284: 133–145. [Google Scholar]
- 22. Berkelmans R, van Oppen MJ (2006) The role of zooxanthellae in the thermal tolerance of corals: a ‘nugget of hope’ for coral reefs in an era of climate change. Proceedings of The Royal Society B 273: 2305–2312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Fitt WK, Pardy RL (1981) Effects of starvation, and light and dark on the energy metabolism of symbiotic and aposymbiotic sea anemones, Anthopleura elegantissima . Marine Biology 61: 199–205. [Google Scholar]
- 24. Weis VM, Levine RP (1996) Differential protein profiles reflect the different lifestyles of symbiotic and aposymbiotic Anthopleura elegantissima, a sea anemone from temperate waters. Journal of Experimental Biology 199: 883–892. [DOI] [PubMed] [Google Scholar]
- 25. Wang J-T, Douglas AE (1998) Nitrogen recycling or nitrogen conservation in an alga-invertebrate symbiosis? Journal of Experimental Biology 201: 2445–2453. [DOI] [PubMed] [Google Scholar]
- 26. Kuo J, Liang Z-C, Lin C-H (2010) Suppression subtractive hybridization identifies genes correlated to symbiotic and aposymbiotic sea anemone associated with dinoflagellate. Journal of Experimental Marine Biology and Ecology 388: 11–19. [Google Scholar]
- 27. Mieog JC, Olsen JL, Berkelmans R, Bleuler-Martinez SA, Willis BL, et al. (2009) The roles and interactions of symbiont, host and environment in defining coral fitness. PLoS ONE 4: e6364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Rodriguez-Lanetty M, Harii S, Hoegh-Guldberg OVE (2009) Early molecular responses of coral larvae to hyperthermal stress. Molecular Ecology 18: 5101–5114. [DOI] [PubMed] [Google Scholar]
- 29. Weis VM, Davy SK, Hoegh-Guldberg O, Rodriguez-Lanetty M, Pringle JR (2008) Cell biology in model systems as the key to understanding corals. Trends in Ecology & Evolution 23: 369–376. [DOI] [PubMed] [Google Scholar]
- 30. Moore SJ (1989) Narcotizing sea anemones. Journal of the Marine Biological Association of the United Kingdom 69: 803–811. [Google Scholar]
- 31.OECD SIDS (2003) Menthols. CAS No: 2216-51-5, 15356-60-2, 89-78-1, 1490-04-6, United Nations Environment Programme Publications. p. 5.
- 32. Wang J-T, Meng P-J, Sampayo E, Tang S-L, Chen C (2011) Photosystem II breakdown induced by reactive oxygen species in freshly-isolated Symbiodinium from Montipora (Scleractinia; Acroporidae). Marine Ecology Progress Series 422: 51–62. [Google Scholar]
- 33. MacDonald MJ (1982) Evidence for the malate aspartate shuttle in pancreatic islets. Archives of Biochemistry and Biophysics 213: 643–649. [DOI] [PubMed] [Google Scholar]
- 34. Rahav O, Dubinsky Z, Achituv Y, Falkowski PG (1989) Ammonium metabolism in the zooxanthellate Coral, Stylophora pistillata . Proceedings of the Royal Society of London Series B 236: 325–337. [Google Scholar]
- 35. Jones BN, Pääbo S, Stein S (1981) Amino acid analysis and enzymatic sequence determination of peptides by an improved o-phthaldialdehyde precolumn labeling procedure. Journal Liquid Chromatography 4: 565–586. [Google Scholar]
- 36. González-Teuber M, Heil M (2009) The role of extrafloral nectar amino acids for the preferences of facultative and obligate ant mutualists. Journal of Chemical Ecology 35: 459–468. [DOI] [PubMed] [Google Scholar]
- 37.Kruskal JB, Wish M (1978) Multidimensional scaling.
- 38.Clarke KR, Warwick RM (1994) Changes in marine communities: an approach to statistical analysis and interpretation.
- 39. Hans M, Wilhelm M, Swandulla D (2012) Menthol suppresses nicotinic acetylcholine receptor functioning in sensory neurons via allosteric modulation. Chemical Senses 37: 463–469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Okazawa M, Terauchi T, Shiraki T, Matsumura K, Kobayashi S (2000) l-Menthol-induced [Ca2+]i increase and impulses in cultured sensory neurons. Neuroreport 11: 2151–2155. [DOI] [PubMed] [Google Scholar]
- 41. McKemy DD, Neuhausser WM, Julius D (2002) Identification of a cold receptor reveals a general role for TRP channels in thermosensation. Nature 416: 52–58. [DOI] [PubMed] [Google Scholar]
- 42. Peier AM, Moqrich A, Hergarden AC, Reeve AJ, Andersson DA, et al. (2002) A TRP channel that senses cold stimuli and menthol. Cell 108: 705–715. [DOI] [PubMed] [Google Scholar]
- 43. Reid G, Babes A, Pluteanu F (2002) A cold- and menthol-activated current in rat dorsal root ganglion neurones: properties and role in cold transduction. The Journal of Physiology 545: 595–614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Haeseler G, Maue D, Grosskreutz J, Bufler J, Nentwig B, et al. (2002) Voltage-dependent block of neuronal and skeletal muscle sodium channels by thymol and menthol. European Journal of Anaesthesiology 19: 571–579. [DOI] [PubMed] [Google Scholar]
- 45. Brown JT, Hegarty PK, Charlwood BV (1987) The Toxcity of monoterpenes to plant cell cultures. Plant Science 48: 195–201. [Google Scholar]
- 46. Pang ZP, Südhof TC (2010) Cell biology of Ca2+-triggered exocytosis. Current Opinion in Cell Biology 22: 496–505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Rodrigues LJ, Grottoli AaG (2007) Energy reserves and metabolism as indicators of coral recovery from bleaching. Limnology Oceanography 52: 1874–1882. [Google Scholar]
- 48. Porter JW, Fitt WK, Spero HJ, Rogers CS, White MW (1989) Bleaching in reef corals: Physiological and stable isotopic responses. Proceedings of the National Academy of Sciences 86: 9342–9346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Szmant-Froelich A, Pilson ME (1984) Effects of feeding frequency and symbiosis with zooxanthellae on nitrogen metabolism and respiration of the coral Astrangia danae . Marine Biology (Berlin) 81: 153–162. [Google Scholar]
- 50. Rees TAV (1986) The Green Hydra Symbiosis and Ammonium I. The Role of the Host in Ammonium Assimilation and its Possible Regulatory Significance. Proceedings of the Royal Society of London Series B Biological Sciences 229: 299–314. [Google Scholar]