

Abstract
Entomoparasitic nematodes (EPNs) are being commercialized as a biocontrol measure for crop insect pests, as they provide advantages over common chemical insecticides. Mass production of these nematodes in liquid media has become a major challenge for commercialization. Producers are not willing to share the trade secrets of mass production and by doing so, have made culturing EPNs extremely difficult to advance existing technologies. Theoretically, mass production in liquid media is an ideal culturing method as it increases cost efficiency and nematode quantity. This paper will review current culturing methodologies and suggest basic culturing parameters for mass production. This review is focused on Heterorhabditis bacteriophora; however, this information can be useful for other nematode species.
Keywords: Beneficial nematodes, Photorhabdus luminescens, Heterorhabditis bacteriophora, Mass production
Introduction
The use of beneficial or entomoparasitic nematodes (EPNs) as a biocontrol measure for insect pests is increasing because they provide several advantages: (i) more effective than chemical agents; (ii) persist within soil; (iii) sustainability; and (iv) safe for user and environment [1–5]. The problem of insect resistance to chemicals has created a genuine interest in EPNs and therefore their market viability is increasing. Currently, these “products” are produced by a few companies and the increasing use of integrated pest management is creating pressures for producers to keep up with customer demands. It is the aim of this review to highlight the methodologies utilized in the mass production of Heterorhabditis bacteriophora.
Nematode Products as Biocontrol Agents
EPN producers/retailers offer multiple products depending on application whereas nematodes can be purchased in pack quantities ranging from 5 to 500 million. The producer/distributor ships nematodes cold-packed and recommends that they be stored at 4 °C if they are not used immediately. From our experience, H. bacteriophora can remain viable for one month at this temperature (unpublished data). Producers may package EPNs on a variety of solid substrates (clay, vermiculite, alginate gels, diatomaceous earth) [5].
Nematodes are roundworms (Fig. 1) that exhibit different lifestyles that include free-living, predaceous or parasitic. Biodiversity of these animals is extensive; however, EPNs strictly belong to the families Steinernematidae and Heterorhabditidae. Many species of nematodes have been identified from these families; however, only few of them are commercially available [6]. Steinernematidae and Heterorhabditidae nematodes are of interest as they are the only nematodes that can parasitize a broad range of insects; however, they will only persist as insect hosts are available. EPNs have a high degree of safety for plants, livestock and humans making them an attractive environmental choice [7, 8]. The likelihood of the nematode becoming a human pathogen is extremely remote because sudden genetic mutations of either the nematode or bacterium would disrupt symbiosis and neither organism would flourish. Furthermore, no life-threatening risks have been associated with the use of beneficial nematodes.
Fig. 1.
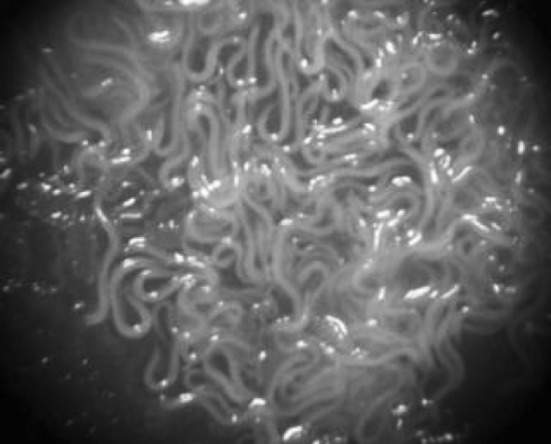
Nematodes of H. bacteriophora growing on a lawn of Photorhabdus luminescens
Research has investigated the utilization of liquid cultures of the bacterial symbionts in order to circumvent the problems with nematode rearing; however, this approach may present safety and/or environmental problems when applied directly to crops [9]. In another approach, the use of cloned toxin genes from bacterial symbionts as an alternative to the Bt toxin was discussed. Other approaches included toxin preparations as potential oral insecticides; however, there is concern for non-target organisms [10, 11].
One of the main issues of mass producing EPNs is yield. Batch requirements are problematic for Heterorhabditidae nematodes; however, they can be reared on solid media, but large-scale production is not cost efficient [12–14]. A more efficient approach is to utilize bioreactors containing liquid media [15, 16]. Bioreactors are expensive; however, scale-up is much easier and economical (Fig. 2).
Fig. 2.

Culturing H. bacteriophora in liquid media: (Left) Fermentor prior to Photorhabdus inoculation. (Right) After 21 days of rearing nematodes
The Symbiotic Relationship
Heterorhabditis bacteriophora nematodes only exist symbiotically with its bacterial symbiont Photorhabdus luminescens. P. luminescens benefits from this relationship by utilizing H. bacteriophora as a vector and ultimately bioconverts the insect into nutrients that can be utilized by both partners. P. luminescens also creates an environment for nematode reproduction within the insect [17–19]. Nematodes can enter the insect larvae through many of the insect’s orifices including cuticle penetration. Upon entry, the bacteria are regurgitated into the insect hemocoel where they proliferate causing insect mortality within 2 days [20, 21].
Photorhabdus Luminescens
Photorhabdus luminescens is a Gram-negative, bioluminescent, pigment producing, phase varying entomopathogen (Fig. 3). This bacterium is not free-living and can only be isolated from its nematode symbiont or infected insects. When viewed under electron microscopy, P. luminescens is 5 microns in length and 1 micron in width (Fig. 4) suggesting that H. bacteriophora can feed upon them (Fig. 5).
Fig. 3.
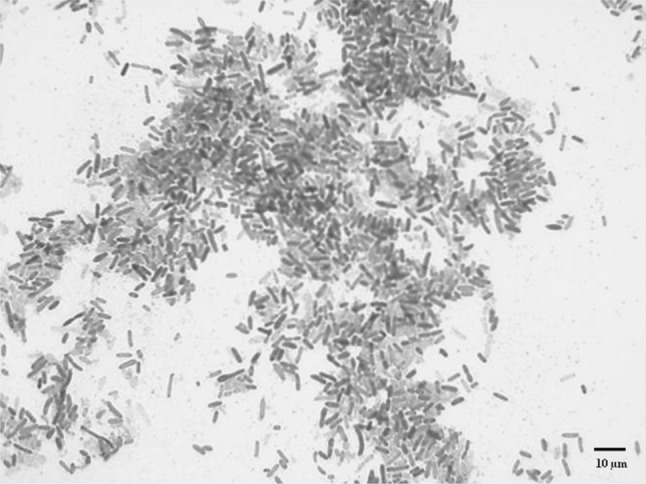
Gram stain of P. luminescens (1,000×)
Fig. 4.

Scanning electron micrograph of P. luminescens
Fig. 5.

Scanning electron micrograph of nematode buccal region
P. luminescens is considered to be a model to study symbiosis, host-pathogen interactions and phase variation. Research has suggested that a single infective juvenile nematode may carry ten cells of P. luminescens, but only three cells are necessary to cause mortality [22]. Other research shows that P. luminescens produces several biologics that are responsible for bioconversion, protection, and nematode transmission [23–27]. Recent studies indicate that insect colonization is due to protective mechanisms against insect macrophages [28]. The makes caterpillars’ floppy toxin that is produced is essential for insect mortality as it causes the insect to lose body turgor [29]. P. luminescens contains a lux operon that is similar to that of the genus Vibrio; however, the lux operon is not associated with quorum sensing [30]. Photorhabdus is genetically related to Xenorhabdus where they form a unique group of entomopathogenic bacteria; however, luminosity and pigmentation is used to differentiate between the genera [31].
Two related species of P. luminescens have been identified: P. temperata and P. asymbiotica. P. luminescens and P. temperata are known to be associated with H. bacteriophora and other species; however, P. asymbiotica is associated with a new species H. gerrardi [32, 33]. Research has shown that the ability of P. luminescens to support a culture of Heterorhabditis spp. is strain and species specific; however, nematode reproduction is influenced by the phase variant [34–36]. Strauch and Ehlers [37] suggested that the bacteria provide a “food signal” that modulates nematode development; however, this signal is yet to be identified [38].
Photorhabdus luminescens undergoes phase variation, meaning that it can shift to a more stable state. This process produces primary and secondary forms (phase I and phase II) which differ morphologically and physiologically [39–41] such as the association of coccoid bodies with phase I cells (Fig. 6). P. luminescens adapts by shifting between these two distinct phenotypes [42]. Each phenotype is continuously generated, but the phenotype with the best fitness increases [43–45].
Fig. 6.

SEM micrograph depicting phase I cells and the associated coccoid bodies (arrow)
Optimum temperature and pH for culturing Photorhabdus is 28 ±2 °C and 7.30 ± 0.01, respectively [46]. There are different media formulations for culturing Photorhabdus. Nutrient broth (5 g digested gelatin, 3 g beef extract) supplemented with 2.5 % trehalose (NBT) adjusted to pH 7.3 works well for routine culturing. Trehalose, a multifunctional sugar, is utilized because of its abundance within the insect and has been suggested to stabilize characteristics such as luminosity [47, 48]. Upon sequencing, Duchaud et al. [26] reported that the genome of P. luminescens does encode a trehalose operon consisting of a repressor protein (treR), a trehalose permease (treB) and a trehalose 6-phosphate hydrolase (treC). Presently, the metabolism of trehalose and trehalose 6-phosphate in both phase variants has yet to be described. Inman and Holmes [47] demonstrated that P. luminescens cultured in 1.5 % trehalose did not form acidic end products when compared to 1.5 % glucose cultures. This suggests that trehalose, at best, is slowly metabolized. Biological phase characteristics (Table 1) are used to differentiate phase I and II cells. Identifying and culturing phase I cells is critical as phase II-shifted bacteria can impact nematode reproduction and yield; therefore phase II cells should be avoided [49].
Table 1.
Phase characteristics of Photorhabdus luminescens [56]
| Characteristic | Phase I | Phase II |
|---|---|---|
| Relative luminosity | +++ | + |
| Red pigmentation | Pos | Neg |
| Yellow pigmentation | Pos | Pos |
| Protease activity | Pos | Neg |
| Lipase activity | ||
| Tween 80 | Pos | Neg |
| Tween 40 | Pos | Pos |
| Antimicrobial production | Pos | Neg |
| Inclusion bodies | Pos | Neg |
| Colony description on: | ||
| NBTA | Maroon-elevated colonies with feathered edges. Media assumes a blue tint | Transparent-elevated colonies that assumes color of media |
| Nutrient agar | Red-elevated colonies. Red pigment diffuses | Yellow-elevated colonies. Yellow pigment does not diffuse |
+++ Highly luminescent; + minimal luminosity, Pos positive; Neg negative
Heterorhabditis bacteriophora
In 1975, H. bacteriophora was described as a new species, genus, and family in the order Rhabditida and was found that infective juveniles were similar to the dauer stage of Caenorhabditis elegans. Also, this “new” species (Fig. 7) is able to transmit a bacterium into the insect [50]. Since then, eleven species of Heterorhabditis have been isolated, described, and studied [51, 52]. The life cycle of H. bacteriophora is an intriguing process (Fig. 8) that consists of five growth stages (Fig. 9). The infective stage is the only stage that is free-living. Han and Ehlers [49] reported that H. bacteriophora develops slowly when phase II bacteria are used because they are not retained by IJs and ultimately do not support growth and reproduction [53].
Fig. 7.
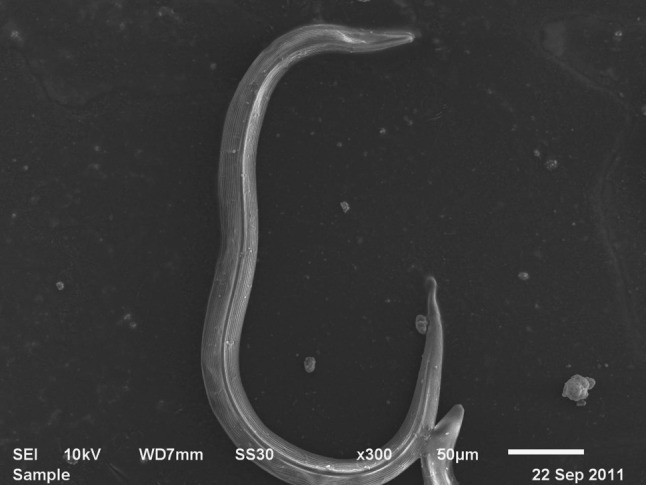
Scanning electron micrograph of H. bacteriophora
Fig. 8.

Life cycle of H. bacteriophora. Used with permission [49]
Fig. 9.
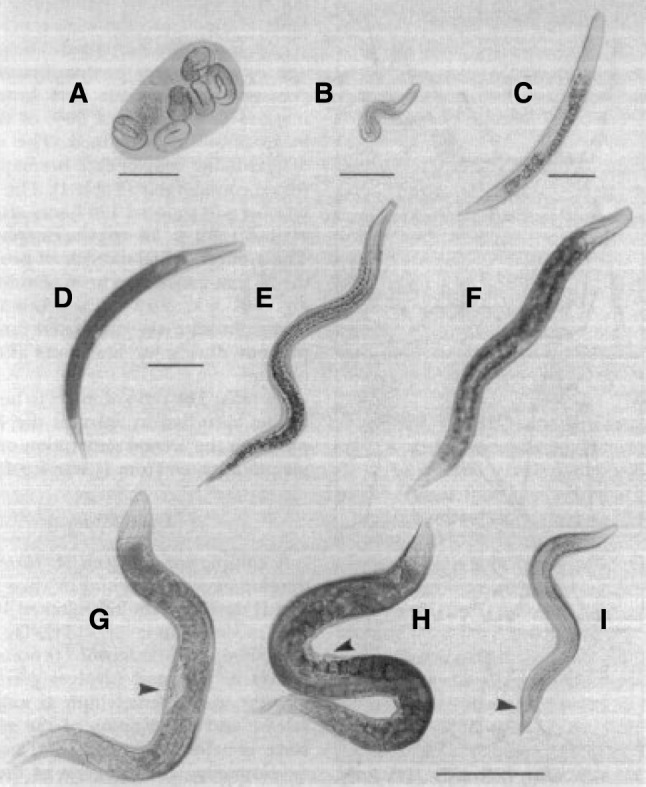
Developmental stages of H. bacteriophora. A Eggs; B Stage 1; C Stage 2; D Feeding stage 3; E Infective stage 3; F Stage 4; G Hermaphrodite (arrow undeveloped vulva); H Mature female (arrow protruding vulva); I Adult male nematode (arrow specula). Scales: A = 12 μm; B = 25 μm; C = 10 μm; D–F = 50 μm; and G–I = 100 μm. Figure used with permission [53]
Mass Production in Liquid Media
There are several ways that cultures of H. bacteriophora can be grown: shake flasks, stirred bioreactors [54], airlift bioreactors [55] and in vivo production. Nematode inoculum densities can vary between 300–4,000 nematodes per milliliter. This section focuses on the use of bench-scale stirred reactors and the parameters used: (i) bacterial culture; (ii) media composition; (iii) oxygen availability/agitation; and (iv) temperature/pH.
Bacterial Culture
Photorhabdus luminescens is isolated from the nematode to ensure that the bacterial strain is specific. The simplest way to isolate the bacteria is to inoculate an insect with IJs. After 2 days, red pigmented larvae are placed into a clear microcentrifuge tube and luminosity measured [56]. These cadavers are aseptically dissected (Fig. 10) and the infected hemolymph is streaked onto nutrient agar containing trehalose (NAT).
Fig. 10.
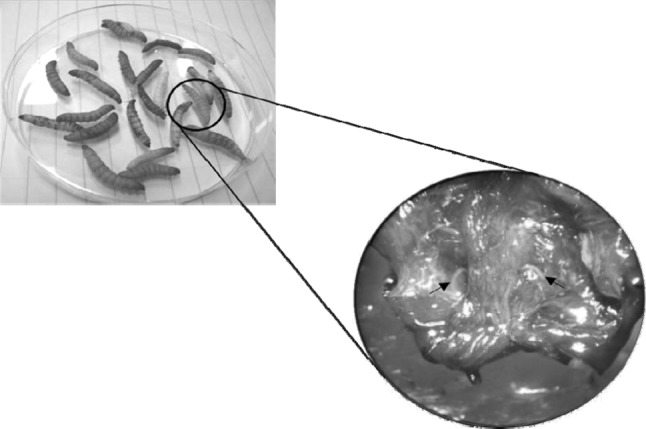
Dissection of a G. mellonellared coloration is due to pigment production. Adult nematodes were seen (arrows)
After three days, presumptive phase I colonies on NAT media are cultured onto NBTA plates for phase confirmation (Fig. 11). Luminosity is measured by suspending a portion of a colony into a milliliter of water in a clear micro-tube. Colonies exhibiting high luminosity and red pigmentation are predominately phase I cells and can be utilized to prepare NBT seed cultures. Seed cultures are then scaled-up for reactor inoculation. Since the nematode culture will be growing for a few weeks in the bioreactor, sterility is critical. Time and effort spent maintaining purity will prevent many processing problems. Since P. luminescens is naturally resistant to ampicillin, the addition of ampicillin reduces potential contamination [57]. Performing Gram stains, measuring luminosity and monitoring pH are convenient ways to monitor culture health.
Fig. 11.
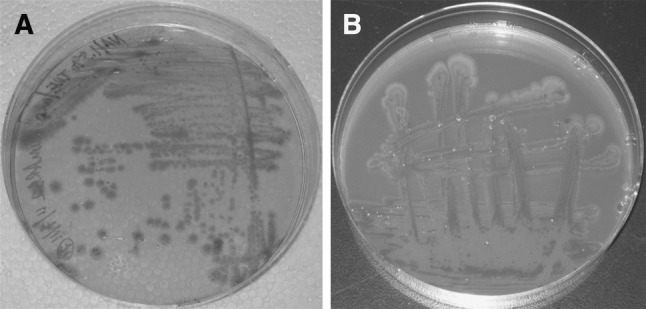
P. luminescens on a NAT and b NBTA plates
Trehalose (Fig. 12), a multifunctional sugar, is abundant in many insects and fungi. Trehalose is also found in C. elegans; however, the role of trehalose is not yet understood [58]. Trehalose serves many biological purposes as it is known to: (i) stabilize cellular constituents during environmental stresses such as high osmolarity and (ii) play a role in bacterial carbohydrate metabolism [59–63].
Fig. 12.

Structure of trehalose
Culture Media
There has been a wide-range of media employed for co-culturing P. luminescens and H. bacteriophora (Table 2), however, NBT is a good formulation for the bacteria. Cultures of P. luminescens grown in NBT will produce red pigmentation and strong bioluminescence. We also found that pigmentation varies with media composition (Figs. 13, 14) and pH [47]. The phase I inoculation concentration should be ~2.5 % of the growth media volume and tested for luminosity and contamination prior to fermentor upscale. After inoculation, P. luminescens is grown for 24 h at 28 °C, with an air flow rate of 1 vvm and agitation of 100 rpm [64].
Table 2.
Liquid media formulations for mass production
| Medium | Scale | Features | Reference |
|---|---|---|---|
| Complex liquid media | Bench to pilot | Chicken egg | [69] |
| Swine kidney/bovine fat | 0.5 L | Polyether/urethane sponges | [35] |
| Lipid culture media | 50 mL | Increasing lipids | [66] |
| Chicken offal | 10 L | Paddle-stirred | [54] |
| Nutrient roux | 0.25 L | Polyurethane foam; soy flour | [12] |
| Modified wouts’ 1 | 20 L | Air lift | [69] |
| Modified wouts’ 2 | 20 and 500 L | Milk powder | [69] |
| Liquid culture medium | 5 L | Increased aeration rates | [37] |
| Liquid nematode media | 10 L | Variable agitation rates; egg yolks | [16] |
| Nematode growth media | 0.5–2 L | Cholesterol, liver extract, | [12] |
| Liquid chicken extract | 2 and 10 L, predicted 30 L | Culture stabilizer; biological buffer | (unpublished data) |
Fig. 13.
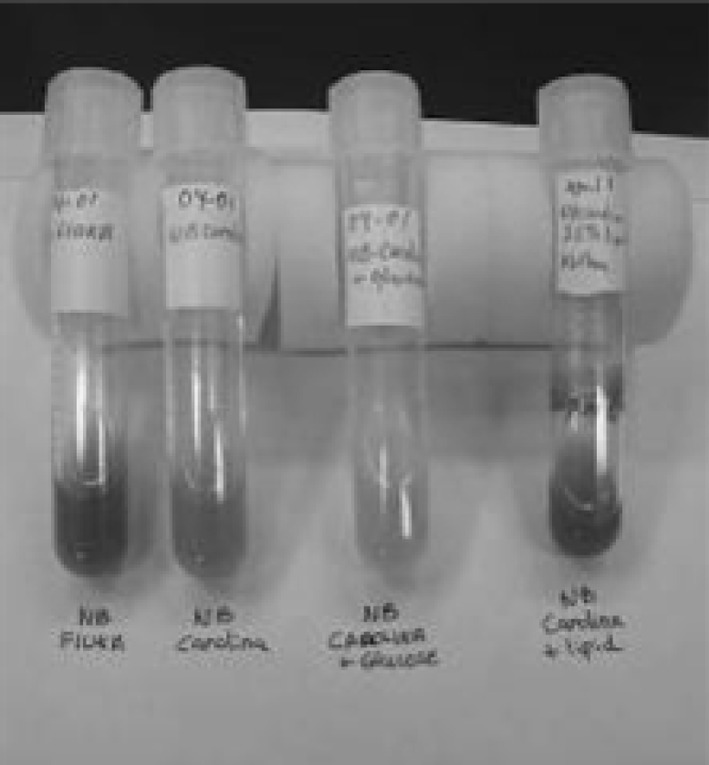
Media pigmentation left to right: Fluka NB gray; Carolina NB orange; Carolina NB + glucose yellow; Carolina NB + lipid brick red. (Color figure online)
Fig. 14.

An upscaled culture of P. luminescens
The medium for the co-culture is much more complex than bacterial media alone [65]. Published formulations contain varying concentrations of the following: (i) peptone; (ii) yeast extract; (iii) beef extract; (iv) lipid; (v) carbohydrate; (vi) cholesterol; and (vii) salts. These media are rich in proteins and lipids because P. luminescens secretes lipases and proteases to degrade these molecules into nutrition for both the nematode and itself. The proteins and lipids found within the media are required due to the longevity of the production process [66]. It is important that growth conditions must satisfy both H. bacteriophora and P. luminescens because as IJs develop, they will require P. luminescens for nutrition. P. luminescens is the diet for all nematode stages except the IJs, which do not feed, but require P. luminescens for the food/development signal [20]. If the bacteria are maintained in the phase I state, the nematodes will develop and produce progeny resulting in maximum yields.
Oxygen Availability and Agitation
Oxygen is poorly soluble in water; therefore, oxygen transfer to the media is critical. Gas to liquid transfer is influenced by the viscosity and volume of the medium, stirrer design/speed and flow rates [67]. The fragile nature of adult nematodes requires low impeller velocities to prevent disruption; therefore, a compromise between agitation and oxygen flow must be reached. Oxygenation is crucial because bacterial demands will compete with those of the nematodes; therefore, an oxygen flow of 1 volume of gas to 1 volume of media per min (vvm) is recommended to prevent the build-up of CO2 [67]. When culturing EPNs, the dissolved oxygen should be high, approximately 40 %, for juveniles and approaching 60 % for adults and the agitation should not be greater than 60 rpm [37].
Temperature and pH
Nematodes and bacteria grow best at 28 °C ± 2 °C [46]. Other temperatures may not produce satisfactory results due to the phase sensitivity of the bacteria (unpublished data). We recommend that the media pH be buffered around 7.30 because it has been observed that the pH increases to 8.0 after 48 h when a buffer is not used [47].
Discussion
Nematode reproduction in vivo is stimulated by the environment inside the insect. Culturing in vitro requires media conditioning with the bacterial symbiont. Others have shown that best production occurs when nematodes are inoculated during late logarithmic growth of phase I cells [46]. After 7 days in culture, juvenile populations should begin to recover (unpublished data). This first generation feeds upon P. luminescens and develop into hermaphrodites. The self-fertilized eggs of the hermaphroditic nematode will hatch internally to give rise to offspring that will feed upon and develop within the maternal nematode (termed endotokia matricida). It is through this process that 80 % of the hermaphroditic nematodes will bear live young after 12 days post-inoculation [68]. These nematodes will emerge from the hermaphroditic nematode in the J3 stage and the life cycle continues. It is clear that the hermaphrodite transmits the symbiotic bacteria required by the infective juveniles. Furthermore, eggs that are produced by sexual reproduction are laid outside of the adults and will hatch within the media; however, in liquid culture adults may or may not be able to reproduce sexually based upon culturing parameters mainly agitation [38]. Development of infective juveniles (IJs) is induced by nutrient depletion of the maternal nematode, insect or media.
Mass producing beneficial nematodes in liquid culture is a challenging task; however, understanding both the nematode and its bacterial symbiont in liquid culture increases the chance of success. For starters, understanding the symbiotic relationship provides information about how they will respond when cultured together. Another daunting aspect of mass production technologies is the role of phase variation in the bacterial symbiont. The phase I variant is crucial to the life cycle of the nematode because the bacteria have the ability to (1) produce toxins and enzymes that kill and bioconvert the insect into nutritional components for both symbionts, (2) produce and excrete antimicrobials to ward off contaminating microbes, (3) produce bioluminescence; however, its function is yet to be determined, (4) secrete “food signals” for nematode recovery, and (5) serve as the main food source of the nematodes. All of the above mentioned processes must occur to create the optimal environment for nematode reproduction.
To mass produce beneficial nematodes in liquid culture, a liquid media formulation must be created that can be utilized by the variant bacteria to produce the necessary reproductive environment for the nematodes. To date, most liquid media consists of high concentrations of proteins and lipids. The addition of a biological buffer and a culture stabilizer may benefit the culturing process. The pH of the media being utilized should be tolerable by both symbionts (pH 7.0–7.3). Another process parameter, dissolved oxygen (DO) is also important. The nematode oxygen demand is dependent on the current nematode growth stage. It is recommended that the DO should be 40 % when the prevalence of juveniles are seen and up to 60 % when adults are in abundance. Agitation, another important parameter in fermentation, should be avoided due to its negative impact on gravid adult nematodes; however, if agitation is desired the speed should not exceed 60 rpm. Published yields of IJs vary considerably with average counts of around 20,000 to 40,000 per mL. Based upon our current media formulation and process parameters we can achieve IJ counts of 50,000 per mL (unpublished data) which exceeds the data range obtained by other researchers. Therefore, consistency in nematode yields continues to be a present-day challenge.
Acknowledgments
Financial support is provided in part by the National Institute of Child Health and Development (5G11HD052381-05); the North Carolina Biotechnology Center (2010-IDG-1008); and Farm Bureau of Robeson County, North Carolina USA.
References
- 1.Bathon H. Impact of entomopathogenic nematodes on non-target hosts. Biocontrol Sci Technol. 1996;6:421–434. doi: 10.1080/09583159631398. [DOI] [Google Scholar]
- 2.Bedding RA, Akhurst RJ, Kaya HK. Future prospects for entomogenous and entomopathogenic nematodes. In: Bedding RA, Akhurst RJ, Kaya HK, editors. Nematodes and the biological control of insect pests. Melbourne: CSIRO; 1993. pp. 157–170. [Google Scholar]
- 3.Ehlers RU. Entomopathogenic nematodes in the European biocontrol market. Commun Agric Appl Biol Sci. 2003;68(4a):3–16. [PubMed] [Google Scholar]
- 4.Ehlers RU, Hokkanen HM. Insect biocontrol with non-endemic entomopathogenic nematodes (Steinernema spp. and Heterorhabditis spp.): conclusions and recommendations of a combined OECD and COST workshop on scientific and regulatory policy issues. Biocontrol Sci Technol. 1996;6:295–302. doi: 10.1080/09583159631280. [DOI] [Google Scholar]
- 5.Gaugler R, Han R. Production Technology. In: Gaugler R, editor. Entomopathogenic nematology. Cambridge: CAB International; 2002. pp. 289–310. [Google Scholar]
- 6.Shapiro-Ilan DI, Gaugler R. Production technology for entomopathogenic nematodes and their bacterial symbionts. J Ind Microbiol Biotechnol. 2002;28:137–146. doi: 10.1038/sj.jim.7000230. [DOI] [PubMed] [Google Scholar]
- 7.Williams RN, Fickle DS, Grewal PS, Dutcher J. Field efficacy against the grape root borer Vitacea polistiformis (Lepidoptera: Sesiidae) and persistence of Heterorhabditis zealandica and H. bacteriophora (Nematoda: Heterorhabditidae) in vineyards. Biol Control. 2010;53:86–91. doi: 10.1016/j.biocontrol.2009.11.004. [DOI] [Google Scholar]
- 8.Boemare NE, Laumond C, Mauleon H. The entomopathogenic nematode–bacterium complex: biology, life cycle and vertebrate safety. Biocontrol Sci Technol. 1996;6:333–345. doi: 10.1080/09583159631316. [DOI] [Google Scholar]
- 9.Mahar AN, Jan ND, Mahar AQ, Mahar GM, Hullio MH, Lanjar AG. Efficacy of entomopathogenic bacterium Photorhabdus luminescens and its metabolites against diamondback moth Plutella xylostella larvae on chinese cabbage and artificial diet. Pak J Nematol. 2008;26(1):69–82. [Google Scholar]
- 10.Bowen D, Rocheleau TA, Blackburn M, Andreev O, Golubeva E, Bhartia R, Ffrench-Constant RH. Insecticidal toxins from the bacterium Photorhabdus luminescens. Science. 1998;280:2129–2132. doi: 10.1126/science.280.5372.2129. [DOI] [PubMed] [Google Scholar]
- 11.Mohan S, Sabir N. Biosafety concerns on the use of Photorhabdus luminescens as biopesticide: experimental evidence of mortality in egg parasitoid Trichogramma spp. Curr Sci. 2005;89(7):1268–1272. [Google Scholar]
- 12.Wouts WM. Mass production of the entomopathogenic nematode Heterorhabditid heliothidis (Nematoda: Heterorhabditidae) on artificial media. J Nematol. 1981;13(4):467–469. [PMC free article] [PubMed] [Google Scholar]
- 13.Torre M. Challenges for mass production of nematodes in submerged culture. Biotechnol Adv. 2003;21:407–416. doi: 10.1016/S0734-9750(03)00057-0. [DOI] [PubMed] [Google Scholar]
- 14.Tabassum KA, Shahina F. In vitro mass rearing of different species of entomopathogenic nematodes in monoxenic solid culture. Pak J Nematol. 2004;22(2):167–175. [Google Scholar]
- 15.Friedman MJ. Commercial production and development. In: Gaugler R, editor. Entomopathogenic nematodes in biological control. Boca Raton: CRC Press; 1990. pp. 152–172. [Google Scholar]
- 16.Friedman MJ, Langston SE, Pollitt S (1991) Mass production in liquid culture of insect-killing nematodes. United States Patent, 5,023,183. Accessed on 3 Oct 2011
- 17.Gouge DH, Snyder JL. Temporal association of entomopathogenic nematodes (Rhabditida: Steinernematidae and Heterorhabditidae) and bacteria. J Invertebr Pathol. 2006;91:147–157. doi: 10.1016/j.jip.2005.12.003. [DOI] [PubMed] [Google Scholar]
- 18.Li XY, Cowles RS, Cowles EA, Gaugler R, Cox-Foster DL. Relationship between the successful infection by entomopathogenic nematodes and the host immune response. Int J Parasitol. 2007;37:365–374. doi: 10.1016/j.ijpara.2006.08.009. [DOI] [PubMed] [Google Scholar]
- 19.Waterfield NR, Ciche T, Clarke D. Photorhabdus and a host of hosts. Annu Rev Microbiol. 2009;63:557–574. doi: 10.1146/annurev.micro.091208.073507. [DOI] [PubMed] [Google Scholar]
- 20.Nguyen KB, Smart GC., Jr Morphology of the life stages of three Heterorhabditis spp. from the infective juvenile to the hermaphrodite. Proc Soil Crop Sci Soc Fla. 1998;57:101–107. [Google Scholar]
- 21.Kahel-Raifer H, Glazer I. Environmental factors affecting sexual differentiation in the entomopathogenic nematode Heterorhabditis bacteriophora. J Exp Zool. 2000;287:158–166. doi: 10.1002/1097-010X(20000701)287:2<158::AID-JEZ6>3.0.CO;2-4. [DOI] [PubMed] [Google Scholar]
- 22.Poinar GO, Thomas GM, Hess R. Characteristics of the specific bacterium associated with Heterorhabditis bacteriophora. Nematol. 1977;23:97–102. doi: 10.1163/187529277X00273. [DOI] [Google Scholar]
- 23.Forst S, Clarke D. Bacteria-nematode symbiosis. In: Gaugler R, editor. Entomopathogenic nematology. Wallingford: CAB International; 2002. pp. 57–77. [Google Scholar]
- 24.Munch A, Stingl L, Jung K, Heermann R. Photorhabdus luminescens genes induced upon insect infection. BMC Genomics. 2008;9:229. doi: 10.1186/1471-2164-9-229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Akhurst RJ. Antibiotic activity of Xenorhabdus spp. bacteria symbiotically associated with insect pathogen nematodes of the families Heterorhabditidae and Steinernematidae. J Gen Microbiol. 1983;128:3061–3066. doi: 10.1099/00221287-128-12-3061. [DOI] [PubMed] [Google Scholar]
- 26.Duchaud E, Rusniok C, Frangeul L, et al. The genome sequence of the entomopathogenic bacterium Photorhabdus luminescens. Nat Biotechnol. 2003;21:1307–1313. doi: 10.1038/nbt886. [DOI] [PubMed] [Google Scholar]
- 27.Somvanshi VS, Kaufmann-Daszczuk B, Kim K, Mallon S, Ciche TA. Photorhabdus phase variants express a novel fimbrial locus, mad, essential for symbiosis. Mol Microbiol. 2010;77(4):1021–1038. doi: 10.1111/j.1365-2958.2010.07270.x. [DOI] [PubMed] [Google Scholar]
- 28.Brugirard-Ricaud K, Duchaud E, Givaudan A, Girard PA, Kunst F, Boemare N, Brehelin M, Zumbihl R. Site-specific antiphagocytic function of Photorhabdus luminescens type III secretion system during insect colonization. Cell Microbiol. 2005;7(3):363–371. doi: 10.1111/j.1462-5822.2004.00466.x. [DOI] [PubMed] [Google Scholar]
- 29.Daborn PJ, Waterfield N, Silva CP, Au CPY, Sharma S, Ffrench-Constant RH. A single Photorhabdus gene, makes caterpillars floppy (mcf), allows Escherichia coli to persist within and kill insects. Proc Natl Acad Sci USA. 2002;99(16):10742–10747. doi: 10.1073/pnas.102068099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Ruby EG, Urbanowski M, Campbell J. Complete gene sequence of Vibrio Fischeri: a symbiotic bacterium with pathogenic congeners. Proc Natl Acad Sci USA. 2005;102(8):3004–3009. doi: 10.1073/pnas.0409900102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Brenner DJ, Farmer JL., III . Family I. Enterobacteriaceae. In: Garrity GM, Brenner DJ, Krieg NR, Staley TJ, editors. Bergey’s manual of systematic bacteriology, volume 2, the proteobacteria, Part B: the gammaproteobacteria. 2. New York: Spinger; 2005. pp. 587–607. [Google Scholar]
- 32.Gerritsen LJM, Smits PH. The influence of Photorhabdus luminescens strains and form variants on the reproduction and bacterial retention of Heterorhabditis megidis. Fundam Appl Nematol. 1997;20(4):317–322. [Google Scholar]
- 33.Plichta KL, Joyce SA, Clarke D, Waterfield N, Stock SP. Heterorhabditis gerrardi n. sp. (Nematoda: Heterorhabditidae): the hidden host of Photorhabdus asymbiotica (Enterobacteriaceae: γ-Proteobacteria) J Helminthol. 2009;83:309–320. doi: 10.1017/S0022149X09222942. [DOI] [PubMed] [Google Scholar]
- 34.Han R, Wouts WM, Li L. Development of Heterorhabditis spp. strains as characteristics of possible Xenorhabdus luminescens subspecies. Rev Nematol. 1990;13(4):411–415. [Google Scholar]
- 35.Bedding RA. Low cost in vitro mass production of Neoaplectana and Heterorhabditis species (Nematoda) for field control of insect pests. Nematologica. 1981;27:109–114. doi: 10.1163/187529281X00115. [DOI] [Google Scholar]
- 36.Ehlers RU, Stoessel S, Wyass U. The influence of phase variants of Xenorhabdus spp. and Escherichia coli (Enterobacteriaceae) on the propagation of entomopathogenic nematodes of the genera Steinernema and Heterorhabditis. Rev Nematol. 1990;13(4):417–424. [Google Scholar]
- 37.Strauch O, Ehlers RU. Influence of the aeration rate on the yields of the biocontrol nematode Heterorhabditis megidis in monoxenic liquid cultures. Appl Microbiol Biotechnol. 2000;54:9–13. doi: 10.1007/s002530000352. [DOI] [PubMed] [Google Scholar]
- 38.Strauch O, Ehlers RU. Food signal production of Photorhabdus luminescens inducing the recovery of entomopathogenic nematodes Heterorhabditis spp. in liquid culture. Appl Microbiol Biotechnol. 1998;50:369–374. doi: 10.1007/s002530051306. [DOI] [Google Scholar]
- 39.Boemare NE, Akhurst RJ. Biochemical and physiological characterization of colony form variants in Xenorhabdus spp. (Enterobacteriaceae) J Gen Microbiol. 1988;134:751–761. doi: 10.1099/00221287-134-7-1835. [DOI] [PubMed] [Google Scholar]
- 40.Forst S, Nealson K. Molecular biology of the symbiotic-pathogenic bacteria Xenorhabdus spp. and Photorhabdus spp. Microbiol Rev. 1996;60:21–43. doi: 10.1128/mr.60.1.21-43.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Lango L, Clarke DJ. A metabolic switch is involved in lifestyle decisions in Photorhabdus luminescens. Mol Microbiol. 2010;77(6):1394–1405. doi: 10.1111/j.1365-2958.2010.07300.x. [DOI] [PubMed] [Google Scholar]
- 42.Rosner BM, Ensign JC, Schink B. Anaerobic metabolism of primary and secondary forms of Photorhabdus luminescens. FEMS Microbiol Lett. 1996;140:227–232. doi: 10.1111/j.1574-6968.1996.tb08341.x. [DOI] [Google Scholar]
- 43.Salaün L, Snyder LAS, Saunders NJ. Adaption by phase variation in pathogenic bacteria. Adv Appl Microbiol. 2003;52:263–301. doi: 10.1016/S0065-2164(03)01011-6. [DOI] [PubMed] [Google Scholar]
- 44.Clarke DJ. Photorhabdus: a model for the analysis of pathogenicity and mutualism. Cell Microbiol. 2008;10(11):2159–2167. doi: 10.1111/j.1462-5822.2008.01209.x. [DOI] [PubMed] [Google Scholar]
- 45.Crawford JM, Kontnik R, Clardy J. Regulating alternative lifestyles in entomopathogenic bacteria. Curr Biol. 2010;20(1):69–74. doi: 10.1016/j.cub.2009.10.059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Wouts WM. Steinernema (Neoaplectana) and Heterorhabdititis species. In: Nickle WR, editor. Manual of agricultural nematology. New York: Marcel Dekker; 1991. pp. 855–897. [Google Scholar]
- 47.Inman FL, III, Holmes LD. The effects of trehalose on the bioluminescence and pigmentation of the phase I variant of Photorhabdus luminescens. J Life Sci. 2012;5:454–465. [Google Scholar]
- 48.Wyatt GR, Kalf GF. The chemistry of insect hemolymph. Pt II. Trehalose and other carbohydrates. J Gen Physiol. 1957;40(6):833–847. doi: 10.1085/jgp.40.6.833. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Han R, Ehlers RU. Effect of Photorhabdus luminescens phase variants on the in vivo and in vitro development and reproduction of Heterorhabditis bacteriophora and Steinernema carpocapsae. FEMS Microbiol Ecol. 2001;35(3):239–247. doi: 10.1111/j.1574-6941.2001.tb00809.x. [DOI] [PubMed] [Google Scholar]
- 50.Poinar GO. Description and biology of a new insect parasitic rhabditoid, Heterorhabditis bacteriophora n. gen., n. sp. (Rhabditida; Heterorhabditidae n. fam.) Nematologica. 1975;21:463–470. doi: 10.1163/187529275X00239. [DOI] [Google Scholar]
- 51.Adams BJ, Fodor A, Koppenhofer HS, Stackebrandt E, Stock SP, Klein MG. Biodiversity and systematics of nematode–bacterium entomopathogens. Biol Control. 2006;37:32–49. doi: 10.1016/j.biocontrol.2005.11.008. [DOI] [Google Scholar]
- 52.Nguyen KB, Hunt DJ. Heterorhabditidae: species descriptions. In: Nguyen KB, Hunt DJ, editors. Entomopathogenic nematodes: systematics, phylogeny and bacterial symbiosis. Leiden: Koninklijke NV; 2007. pp. 611–692. [Google Scholar]
- 53.Zioni S, Glazer I, Segal D. Life cycle and reproductive potential of the nematode Heterorhabditis bacteriophora strain HP88. J Nematol. 1992;24(3):352–358. [PMC free article] [PubMed] [Google Scholar]
- 54.Ehlers RU, Lunau S, Krasomil-Osterfeld K, Osterfeld KH. Liquid culture of the entomopathogenic nematode–bacterium-complex Heterorhabditis megidis/Photorhabdus luminescens. Biocontrol. 1998;43:77–86. doi: 10.1023/A:1009965922794. [DOI] [Google Scholar]
- 55.Stanbury PF, Whitaker A, Hall SJ. Principles of fermentation technology. 2. Butterworth, Heinemann: Oxford Press; 2003. [Google Scholar]
- 56.Bleakley B, Nealson KH. Characterization of primary and secondary forms of Xenorhabdus luminescens strain Hm. FEMS Microbiol Ecol. 1988;53:241–250. [Google Scholar]
- 57.Dunphy GB. Physicochemical properties and surface components of Photorhabdus luminescens influencing bacteria interaction with non-self response systems of nonimmune Galleria mellonella larvae. J Invertebr Pathol. 1995;65:25–34. doi: 10.1006/jipa.1995.1004. [DOI] [Google Scholar]
- 58.Pellerone FI, Archer SK, Behm CA, Grant WN, Lacey MJ, Somerville AC. Trehalose metabolism genes in Caenorhabditis elegans and filarial nematodes. Int J Parasitol. 2003;33:1195–1206. doi: 10.1016/S0020-7519(03)00173-5. [DOI] [PubMed] [Google Scholar]
- 59.Chandra GK, Chater F, Bornemann S. Unexpected and widespread connections between bacterial glycogen and trehalose metabolism. Microbiol. 2011;157:1565–1572. doi: 10.1099/mic.0.044263-0. [DOI] [PubMed] [Google Scholar]
- 60.Jain NK, Roy I. Effect of trehalose on protein structure. Protein Sci. 2008;18:24–36. doi: 10.1002/pro.3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Iturriaga G, Suarez R, Nova-Franco B. Trehalose metabolism: from osmoprotection to signaling. Int J Mol Sci. 2009;10:3793–3810. doi: 10.3390/ijms10093793. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Ohtake S, Wang YJ. Trehalose: current use and future applications. J Pharm Sci. 2010;100(6):2020–2053. doi: 10.1002/jps.22458. [DOI] [PubMed] [Google Scholar]
- 63.Krasomil-Osterfeld KC. Influence of osmolarity on phase shift in Photorhabdus luminescens. Appl Environ Microbiol. 1995;61(10):3748–3749. doi: 10.1128/aem.61.10.3748-3749.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Moreau E, Inman FL, III, Singh S, Walters H, Holmes LD. Remote control of fed-batch fermentation systems. J Chem Chem Eng. 2011;5:897–902. [Google Scholar]
- 65.Singh S, Moreau E, Inman III FL, Holmes LD (2011). Characterization of Photorhabdus luminescens growth for the rearing of the beneficial nematode Heterorhabditis bacteriophora. Indian J Microbiol. doi:10.1007/s12088-011-0238-7 [DOI] [PMC free article] [PubMed]
- 66.Yoo SK, Brwon I, Gaugler R. Liquid media development for Heterorhabditis bacteriophora: lipid source and concentration. Appl Microbiol Biotechnol. 2000;54:759–763. doi: 10.1007/s002530000478. [DOI] [PubMed] [Google Scholar]
- 67.Garcia-Ochoa F, Gomez E. Bioreactor scale-up and oxygen transfer rate in microbial processes: an overview. Biotechnol Adv. 2009;27:153–176. doi: 10.1016/j.biotechadv.2008.10.006. [DOI] [PubMed] [Google Scholar]
- 68.Baliadi Y, Yosiga T, Kondo E. Development of endotokia matricida and emergence of originating infective juveniles of Steinernematid and Heterorhabditid nematodes. Jpn J Nematol. 2001;31(1/2):26–35. [Google Scholar]
- 69.Surrey MR, Davies RJ (1996) Pilot-scale liquid culture and harvesting of an entomopathogenic nematode, Heterorhabditis bacteriophora. J Invertebr Pathol 67:92–99