

Abstract
Background & objectives:
Acute tubular necrosis (ATN) caused by renal ischaemia, renal hypo-perfusion, or nephrotoxic substances is the most common form of acute kidney injury (AKI). There are a few treatment options for this life-threatening disease and the mortality rate exceeds 50 per cent. In critical cases of AKI the only option is renal transplantation. In the present study we evaluated whether bone marrow cells (BMCs) are involved in regeneration of kidney tubules following acute tubular necrosis in the mouse.
Methods:
Six to eight week old C57BL6/J and congenic enhanced green fluorescence protein (eGFP) mice were used. The relative contributions of eGFP-expressing BMCs were compared in two different approaches to kidney regeneration in the mercuric chloride (HgCl2)-induced mouse model of AKI: induced engraftment and forced engraftment. In vitro differentiation of lineage-depleted (Lin-) BMCs into renal epithelial cells was also studied.
Results:
In the forced engraftment approach, BMCs were found to play a role in the regeneration of tubules of renal cortex and outer medulla regions. About 70 per cent of donor-derived cells expressed megalin. In vitro culture revealed that Lin- BMCs differentiated into megalin, E-cadherin and cytokeratin-19 (CK-19) expressing renal epithelial cells.
Interpretation & conclusions:
The present results demonstrate that Lin- BMCs may contribute in the regeneration of renal tubular epithelium of HgCl2-induced AKI. This study may also suggest a potential role of BMCs in treating AKI.
Keywords: Acute kidney injury (AKI), bone marrow cells, megalin, plasticity, transplantation, tubular epithelial cells
Acute tubular necrosis (ATN) due to renal ischaemia, renal hypo-perfusion, or nephrotoxic substances is the most common form of acute kidney injury (AKI). There are only a few treatment options for this life-threatening disease with more than 50 per cent mortality rate1,2. The pathological manifestations of AKI are death of tubular cells and shedding of cellular components into the lumen of the tubules causing their blockage. The glomerular filtration rate is also reduced in AKI. Obstruction of the ureter may result in major destruction of the kidney medulla. Once the obstruction is removed, the tubules in the medulla can regenerate but without forming new nephrons3. It is believed that the cells eliciting such regeneration arise from dedifferentiation and migration of tubular cells in the damaged areas before redifferentiation4,5 and/or endogenous renal precursor cells6.
Several groups have examined the engraftment and retention of bone marrow (BM)-derived cells in the damaged kidney and their involvement in the overall regeneration process7–14. In various AKI models the engraftment potential of donor-derived cells in kidney has been reported to vary from low (less than 1%) to high (20%). A distinct fraction of these cells was incorporated into regenerating renal tubules. In contrast, two independent studies have proposed that BM-derived cells play a purely paracrine role in the functional recovery of injured nephrons and/or kidney15,16.
Majority of studies have been conducted in an ischaemia/reperfusion (I/R) model as the pathogenesis of this model is comparable to that of human AKI. However, different studies have reported highly variable results. In the present investigation, we have assessed the role of BMCs in the regeneration of kidney tubules in a mercuric chloride (HgCl2)-induced AKI mouse model. Kidney is the primary site for the uptake, accumulation, and toxicity of Hg ions. Following accidental ingestion these accumulate in the straight proximal tubules and cause acute toxic renal oncosis17. The mechanism of Hg toxicity is thought to involve oxidative stress, mitochondrial damage, and disruption of cadherin/catenin complexes, all of which lead to renal failure17,18. In this investigation, we have compared the contribution of eGFP expressing BMCs in induced-engraftment and forced-engraftment approaches for regeneration of tubular epithelium in mouse AKI model. We also report differentiation of lineage-depleted BMCs into renal tubular epithelial cells in culture.
Material & Methods
Animals: This study was conducted in the Stem Cell Biology Laboratory, National Institute of Immunology, New Delhi. Six to 8 wk old C57BL6/J and congenic enhanced green fluorescence protein (eGFP) expressing [CByJ.B6-Tg (UBC-GFP)30 Scha/J] mice were used in this investigation. Mice were obtained from the Jackson Laboratories (Bar Harbor, USA) and were maintained in the experimental animal facility of the Institute. Mice were kept in an isolator and were fed ad libitum with autoclaved acidified water and irradiated food. All experiments were conducted according to procedures approved by the Animal Ethics Committee of National Institute of Immunology.
Acute kidney injury model: AKI was induced by single subcutaneous injection of HgCl2 (7 mg/kg bw) in female mice19. Animals were sacrificed on days 1, 2, 3 and 5 post-injection; serum samples were collected for the estimation of creatinine and blood urea nitrogen (BUN) levels using respective kits (Transasia Bio-Medicals, India). Kidneys were dissected and fixed in 10 per cent buffered formalin. Sections (5 μm) of paraffin-embedded tissue were stained with haematoxylin and eosin (H&E) for histological examination.
Role of bone marrow cells in the regeneration of damaged tubules: Two different approaches were adopted in AKI model. In approach I, sub-lethally (700 cGy) irradiated mice were transplanted with 5 × 106 eGFP-expressing mononuclear BMCs through the tail vein. These mononuclear cells were isolated from eGFP-transgenic mouse bone marrow by flushing tibia and femurs with medium. Mice were maintained for 6 wk to obtain at least 50 per cent donor cell chimerism in peripheral blood (PB). AKI was induced in the chimeric mice by a single injection of HgCl2. This approach has been termed as induced-engraftment. In approach II, AKI was induced by a single injection of HgCl2. Forty eight hours after AKI, 5 × 106 eGFP-expressing mononuclear BMCs were transplanted through the tail vein. This approach has been termed forced-engraftment.
In vitro differentiation of Lin- cells: Lin- BMCs (5000/well) were cultured on 0.01 per cent collagen type I-coated 96-well plates in the presence of Iscove's modified Dulbecco's medium (IMDM) supplemented with 10 per cent damaged kidney serum (DKS), 10 ng/ml hepatocyte growth factor, 10 ng/ml epidermal growth factor, and 20 ng/ml insulin-like growth factor-I for 7 days. DKS was collected from the mouse 48 h after HgCl2 injection (7 mg/kg bw). Mice were bled through retro-orbital plexus till death. After clotting serum was collected, pooled and sterile filtered prior to storage at -70°C. Freshly-isolated Lin- BMCs were used to examine the expression of the haematopoietic marker (CD45). As a negative control, Lin- cells were cultured in medium supplemented with 10 per cent normal mouse serum. Fresh primary cells from the kidney cortex were used as a positive control. Prior to staining with antibodies cells were fixed with 4 per cent (w/v) paraformaldehyde for 30 min at 4°C and then permeabilized with 0.1 per cent Triton X-100 for 30 min at room temperature. The primary antibodies used were anti-megalin (a kind gift of Mr Dan Biemesderfer, Yale University, USA), anti-E-cadherin (Abcam plc., UK), anti-CK-19 (Santa Cruz Biotechnology Inc., USA), and mouse CD45/PE (BD Pharmingen, USA). Primary antibody staining was detected using goat anti-rabbit IgG Alexa Fluor 488 and donkey anti-goat IgG Alexa Fluor 594 (Molecular Probes Inc., USA), anti-rabbit IgG/PE (Jackson Immuno Research Lab., USA) and the nuclei were stained with 4’,6-diamidino-2-phenylindole (DAPI). As a control for immunofluorescence cells were stained with fluorochrome-conjugated secondary antibodies alone. Cells were imaged under an Olympus fluorescence microscope (Olympus, Japan) using LCPlanFl 20× objectives and a DP70 digital camera. Olysia Bioreport was the software (Olympus, Japan) used for image acquisition. Images were composed and edited in Photoshop 6.0 (Adobe).
Single cell preparation of kidney tissue: Kidney was dissected and the cortex region was carefully sliced-out and digested with 0.03 per cent collagenase IV at 37°C for 45 min. The larger tissue fragments were allowed to settle and single cell suspension was collected and centrifuged at 450 g for 5 min. The supernatant was discarded and the pellet was re-suspended in 1 ml of IMDM supplemented with 3 per cent FCS.
Isolation of Lin- cells: Lin- BMCs were fractionated by magnetic cell sorter using negative selection method (Miltenyi Biotech., Germany). The lineage (Lin) antibody cocktail used in this study was a mixture of antibodies against CD5, CD45R, CD11b, Gr1, Ter119, 7-4.
Immunohistochemistry: Kidneys were fixed in PLP buffer (2% paraformaldehyde, 75 mM L-Lysine, 10 mM sodium periodate, and 750 mM sucrose, pH 7.4) at 4°C for 24 h, and frozen in commercial optimal cutting temperature (OCT) medium (Leica Microsystems Nussloch GmbH, Germany). Serial sections of 5 μm thickness were cut and stored at -70°C. The cryosections were treated with 0.1 per cent Triton-X 100 for 30 min and stained with anti-megalin, mouse anti-GFP (Stressgen, USA), anti-F4/80 (Abcam, UK) and anti-vWF (Santa Cruz Biotechnology Inc., USA) antibodies. The washed sections were further treated with anti-rabbit IgG/PE, anti-goat IgG Alexa Flour 594, or anti-mouse Alexa Fluor 488, depending upon the source of the primary antibody. As a control for immunofluorescence sections were stained with fluorochrome-conjugated secondary antibodies. Cells were imaged under an Olympus fluorescence microscope using LCPlanFl 20x objectives and under a Zeiss LSM 510 META confocal laser scanning microscope using Acroplan 40X/0.65 Ph2 objective. LSM 510 software (Calr Zeiss Inc., USA) used for acquisition of images and processed by Zeiss LSM Image browser version 4.2.0.121.
Flow cytometry: An aliquot (0.5 × 106/200 μl) of single cells isolated from cortex of control and transplanted mice were permeabilized as above and stained by incubating with 50 μl of diluted primary antibodies (anti-megalin and anti-vWF) for 45 min at 4°C. The washed cells were further incubated with 50 μl of diluted secondary antibodies (anti-rabbit IgG/PE and anti-goat IgG/PE) for 30 min at 4°C. Finally, the washed cells were analyzed on a customized BD-LSR machine (BD Biosciences, USA).
Statistical analysis: Results of multiple experiments were reported as the means ± SEM. One-way ANOVA was employed to calculate the statistical significance.
Results
BMCs contribute to the repair of damaged renal tubules: A single injection of HgCl2 was found to induce AKI in mice. Serum creatinine and BUN in the mice were increased within 24 h of HgCl2 administration (Fig. 1A). These two serum parameters were further increased up to 48 h, reaching to highest levels at 2.6 ± 0.5 mg/dl and 183 ± 21.9 mg/dl, respectively (Fig. 1A). Pathological changes were predominantly observed in cortical and outer medullary regions of kidney within 24 h of HgCl2 administration (Fig. 1B). Changes included interstitial oedema, tubular dilation, and sloughing of individual epithelial cells. Sloughing of brush borders with tubular dilation was clearly evidenced in straight proximal tubules of outer medulla outer strip. Intraluminal granular casts were frequently observed (Fig. 1B).
Fig. 1.

(A): Effect of HgCl2 on plasma creatinine and blood urea nitrogen (BUN). Single dose of HgCl2 (7 mg/kg, bw) was administered in C57BL6/J mice. At each time interval, plasma creatinine and blood urea nitrogen level was estimated. The highest values were noticed on second day of the treatment. Number of mice (n) = 6 for each group, *P<0.01 compared to day 0 and day 1. (B) Effect of HgCl2 on histology of kidney. Normal saline and HgCl2 was administered in two groups of mice. At each time interval, mice were sacrificed and 5 μm tissue sections were stained with H&E. The pathological changes in the kidney tissue include interstitial oedema, tubular dilation, and sloughing of individual epithelial cells. Sloughing of brush borders with tubular dilation was clearly evidenced in straight proximal tubules of outer medulla outer strip. Intraluminal granular casts were frequently observed. Control section shows intact tubules and glomeruli. Arrows indicate granular casts. Magnification 200×.
To determine the transdifferentiation potential of the engrafted BMCs into renal tubular epithelial cells in AKI mice in the induced-engraftment approach serial kidney sections were co-stained with GFP and megalin antibodies. Megalin is expressed on the luminal surface of renal proximal tubules where it acts as an endocytic receptor for retrieving essential metabolites from the body fluid20. The representative photomicrographs of tissue sections of kidney (induced-engraftment approach) after 1 month of AKI are shown in Fig. 2 (panel A). Bone marrow-derived cells did not express megalin in all four mice of the group. Similar results were obtained in case of mice after 7 months of AKI. Most of the donor cells were engrafted in the interstitial space, as shown in Fig. 2 (panel C). In order to identify the type of cells engrafted in the interstitial space, the kidney sections were stained with von Willebrand factor (vWF) antibody. The representative images of the sections suggest that in the induced-engraftment, BM-derived cells were primarily involved in restoration of peritubular endothelial cells Fig.2E. In saline injected control mice no BMC was detected in the interstitial space (data not shown).
Fig. 2.
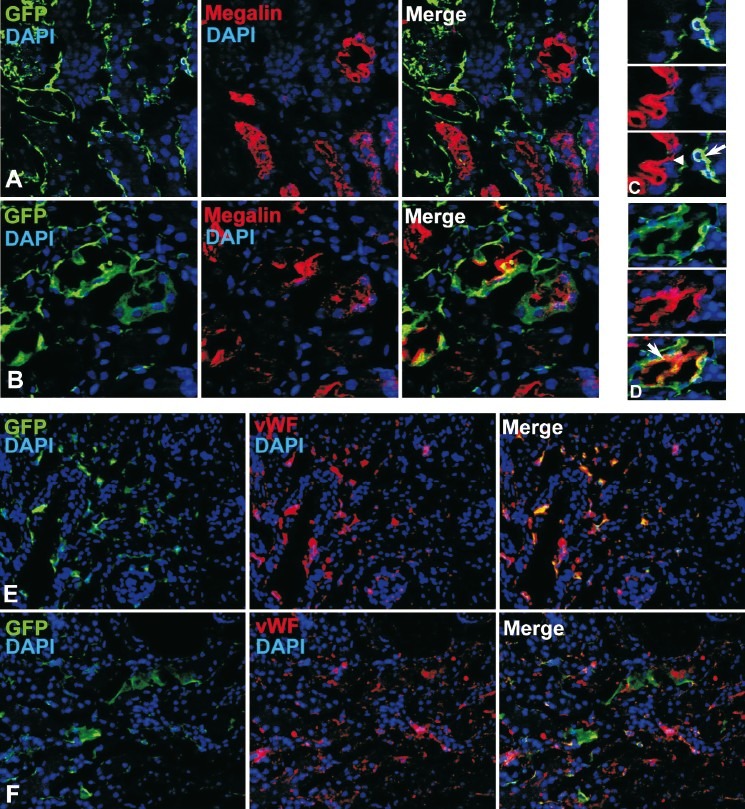
Expression of megalin and vWF in the donor-derived cells (induced-engraftment approach). eGFP chimeric mice received single injection of HgCl2. Serial kidney cryosections (5 μm) were stained with anti-GFP, anti-megalin, and anti-vWF antibodies. The representative photomicrographs after 1 month of HgCl2 treatment are shown. (A) eGFP-expressing cells are engrafted in the interstitial space, but do not express megalin. (C) Magnified view of a section distinctly shows that the donor cells (long white arrow) are not expressing megalin. The host tubular epithelial cells (short white arrow) are expressing megalin (red). (E) Co-expression of eGFP and vWF in non-tubular cells. Expression of megalin and vWF in the donor-derived cells (forced-engraftment approach). After 48 h of HgCl2 treatment, each mouse received tail vein injection of 5 × 106 eGFP-expressing BMCs. One month after transplantation mice were sacrificed and kidney tissues were analyzed. Serial kidney cryosections (5 μm) were stained with anti-GFP, anti-megalin, and anti-vWF antibodies. (B) eGFP-expressing cells are mostly engrafted on the tubules, many of them expressed megalin. (D) Magnified view of a section distinctly shows that donor cells (white long arrow) expressing megalin (red) on the apical surface of the tubular epithelial cells. (F) Photomicrographs show eGFP and vWF are not expressed by the same cells. Number of mice (n) = 4 for each group. Magnification: 400× (megalin); 200× (vWF).
The contribution of BMCs to the regeneration of tubules was examined after 1 month of transplantation in the forced-engraftment approach. Immunohisto-chemical analysis revealed that donor cells were present in the proximal tubules of the cortex and the outer medullary region of the kidney only in damaged-transplanted mice (Fig. 2, panel B and D). The engrafted cells did not stain with vWF antibody, indicated that in forced-engraftment approach BMCs were not involved in the restoration of peritubular endothelial cells (Fig. 2F). Kidney sections were also stained with F4/80 antibody to check the presence of macrophages in the engrafted cells. It was revealed that F4/80 reactive cells were absent, confirming that macrophages were not present. As a control of HgCl2 treatment, same numbers of BMCs were injected in saline treated mice, but no engraftment of eGFP-expressing cells was detected in the host tubules. Normal saline injection did not cause any injury to the interstitial tissue or kidney tubules, therefore eGFP-expressing BMCs were not engrafted. Donor cells were not detected in the organ like lungs. In this approach, a major fraction of the donor-derived cells expressed megalin, indicating that these were involved in regeneration of tubules. The donor-derived cells expressed megalin on the apical surface of the tubular epithelial cells (Fig. 2, panel D), as normally seen in the resident cells. Staining of megalin in the tissue sections was specific as it was limited to proximal tubules of cortex and outer medullary region. Again, the secondary antibody alone failed to stain any target cells further confirming that the staining was specific.
After confirming that BMCs transdifferentiate into megalin expressing cells only in the forced-engraftment approach, cell numbers were quantified by flow cytometry. Representative dot-plots of engrafted eGFP+ cells and their differentiated phenotypes expressing megalin and vWF are shown in Fig. 3A. Quantitative analysis revealed that about 2.2 ± 0.35 per cent (n=4) cells of the cortical and outer medullary regions of the kidney (n=4) were donor-derived. Interestingly, 1.6 ± 0.25 per cent and 0.3 ± 0.07 per cent of these cells were GFP+Meg+ and GFP+vWF+, respectively (Fig. 3B). These demonstrated that a significantly (P<0.01) larger fraction of BM-derived cells differentiated into tubular epithelial cells than endothelial cells.
Fig. 3.

Flow cytometric analysis of eGFP+, GFP+Megalin+, and GFP+vWF+ cells. Megalin (Meg) and vWF antibody stained cells from cortex and outer medullary regions (forced-engrafted approach) were analyzed by flowcytometer after 1 month of transplantation. (A) Representative dot-plots for eGFP+, GFP+Meg+, and GFP+vWF+ cells are shown. The analysis is based on ‘R1’ gated cells in SSC/FSC plot. (B) Percentage engraftment of eGFP+ cells and their differentiation into GFP+Meg+ and GFP+vWF+ cells determined from the dot-plots. n = 4. *P<0.01 with respect to GFP+vWF+ cells.
Lin- cells express megalin, E-cadherin and cytokeratin-19 in culture: Lineage negative (Lin-) BMCs were cultured on plates coated with collagen type I in the presence of damaged kidney serum (DKS) and growth factor-supplemented medium. The freshly harvested Lin- cells were typically round and some of these expressed CD45 (Fig. 4A). Lin- cells did not express megalin or cytokeratin-19 (CK-19) after 7 days of culture in normal mouse serum supplemented medium (Fig. 4C & 4D), but mesenchyma cells expressed vimentin (Fig. 4E). Staining with F4/80 antibody did not show any reactive cells, which showed that cultured cells are not macrophages. Thus, the expression of vimentin and the absence macrophage marker (F4/80) and tubular epithelial markers (megalin and CK-19) suggested that the adhered cells are BM-derived mesenchymal origin. After 7 days of culture in differentiation medium a small number of cells adhered to the plate (Fig. 4G). In these cells CD45 expression was negligible but megalin was robustly expressed (Fig. 4G). As a negative marker of differentiation, these cells were co-stained with megalin and vimentin. Interestingly, vimentin was completely downregulated in these cells with concomitant expression of megalin (Fig. 4, H-J), suggesting that mesenchymal phenotype has been lost. In addition to megalin, renal epithelial cells also expressed E-cadherin and CK-19. The adhered cells expressed both megalin and E-cadherin (Fig. 4, K-M), and megalin and CK-19 (Fig. 4, N-P), indicating that Lin- BMCs can differentiate into renal tubular epithelial cells. Further, to confirm that Lin- BMCs are the competent cells for differentiation and the specificity of megalin antibody, NIH3T3 cells were cultured in identical culture conditions. The results showed that megalin was not expressed in NIH3T3 cells. Again, it was confirmed that vimentin expressing fibroblasts like NIH3T3 was not the candidate cells for such differentiation. The primary renal tubular epithelial cells of the cortical region (positive control) were found to express both megalin and CK-19 (Fig. 4, Q-S). The culture medium supplemented with normal serum and growth factors did not support differentiation (data not shown), suggesting that DKS may contain other factors required for differentiation. These in vitro experiments suggest that depending on the culture environment, a fraction of Lin- BMCs can differentiate into megalin, CK-19 and E-cadherin expressing tubular epithelial cells.
Fig. 4.
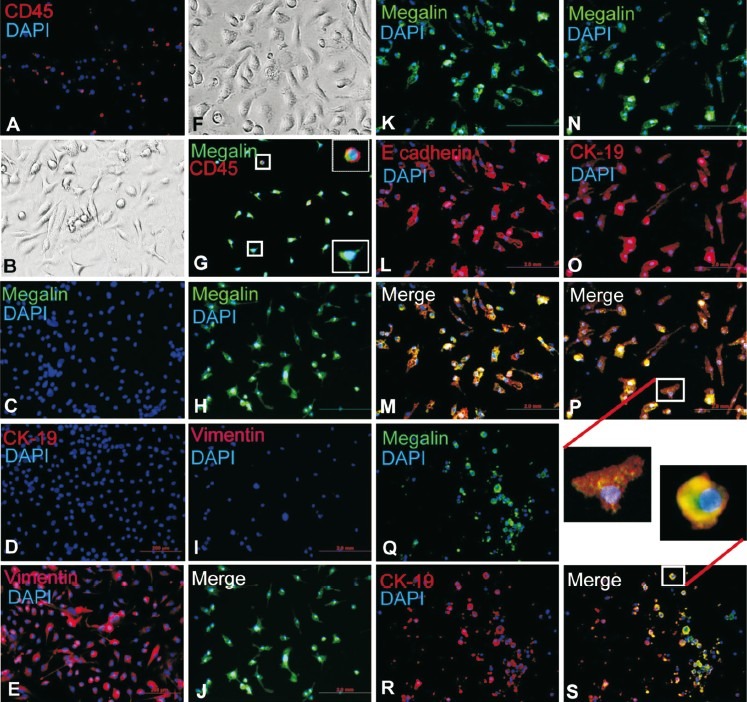
In vitro differentiation of Lin- BMCs into megalin, E-cadherin and CK-19 expressing epithelial cells. Lin- BMCs were cultured for 7 days. The cultured cells were stained with anti-CD45, anti-megalin, anti-CK19, anti-vimentin, and anti-E-cadherin antibodies. (A) Fresh Lin- BMCs stained with CD45/PE staining; (B-E): Lin- BMCs cultured in normal serum supplemented medium (negative control), (B) Adhered cells in bright field, (C) Megalin, (D) CK-19, (E) Vimentin; (F, G): Lin- BMCs were cultured in differentiating medium, (F) Adhered cells in bright field, (G) Co-stained for megalin and CD45 (enlarged images of cells in left two boxes are shown in the right); (H-J): Lin- BMCs were cultured in differentiating medium, (H) Megalin, (I) Vimentin, (J) Merge image of H & I; (K-M): Lin- BMCs were cultured in differentiating medium, K. Megalin, (L) E-cadherin, (M) Merge image of K & L; (N-P): Lin- BMCs were cultured in differentiating medium, (N) Megalin, (O) CK-19, (P) Merge image of N & O; (Q-R): Primary kidney cells (positive control), (Q) Megalin, (R) CK-19, (S) Merge of Q & R. Not all cells show the expression of megalin and CK-19 due to crude preparation of the primary cells. Inset show magnified images of cells (differentiated tubular epithelial and primary renal epithelial) co-expressing megalin and CK-19. Nuclei were stained blue with DAPI. Number of experiment = 3. Magnification: 200×.
Discussion
Our findings showed that BMCs contributed to the regeneration of the renal tubular epithelium in HgCl2-induced AKI mice. It was previously reported that the mechanisms of tubular damage and recovery in I/R and HgCl2-induced injury are not identical21. However, in this study it was observed that HgCl2-induced tubular damage in the cortical and outer medullary regions was comparable to that seen in an I/R model12. To study the contribution of BMCs to tubular regeneration, two different approaches were followed for engraftment of cells at the site of injury: (i) indirectly, through mobilization, and (ii) directly, via tail vein injection.
The administration of HgCl2 caused significant pathological changes in the proximal tubules of the cortex region. As a consequence of this injury, mobilization of Lin-Sca-1+ cells was observed in the peripheral circulation and cell engraftment into the cortex of the damaged kidney. Injury-induced mobilization of Lin-Sca-1+ cells in peripheral blood suggested that these cells might play a role in the recovery of necrotic tissues. However, we have not noticed the involvement of BM-derived (mobilized) cells in tubular regeneration. The majority of the engrafted cells differentiated into endothelial cells of the peritubular region.
In the forced-engraftment approach donor-derived megalin-expressing cells were seen in the proximal tubules of the cortex within 15 days of transplantation (data not shown). Flow cytometric analysis of renal cells from cortical region after 1 month of transplantation indicated that a major fraction of the engrafted cells differentiated into megalin expressing tubular epithelial cells. Earlier studies in I/R model showed that BM-derived cells insignificantly contributed in the restoration of epithelial integrity of the tubules, whereas majority of these were differentiated as interstitial cells14,15,22. The difference in the outcome of present and earlier studies may be attributed to the extent of kidney damage, and the type and number of cell transplanted.
One study using an I/R model23 and two studies using the HgCl2 model19,24 demonstrated that BM-derived cells could make a minor but distinct contribution to the regeneration process following ATN. It has been hypothesized that the natural flux of BMCs to kidney is stimulated in case of ATN for assisting regeneration and repair19. We observed increase in homing of BMCs in damaged kidney for the purpose other than regeneration of renal parenchyma. In a similar AKI model27 it has been concluded that haematopoietic lineage-committed BMCs, but not the mesenchymal stem cells (MSCs), involve in the repair process after tubular injury. In our study, crude BMCs were used, so based on in vivo experiments it could not be concluded whether haematopoietic or mesenchymal lineage was involved for tubular regeneration. However, our in vitro experiment suggested that BM-derived mesenchymal cells were the candidate for differentiation of tubular epithelial cells. Exogenous MSCs have been reported to be involved during the development of kidney25, as well as in tubular regeneration after the damage26. The present comparative study suggested that in the forced engraftment approach, BMCs were found to contribute for the regeneration of tubules in non-irradiated mice. This study contradicted an earlier report showing that exogenous BMCs apparently contribute little to the repair of the HgCl2-damaged kidney in non-irradiated mice27. The exact reason for such difference in outcome between these studies is not clear, but irradiation cannot be the issue in this model. Mercuric chloride specifically depletes proximal renal tubular cells, even mouse is not irradiated. The dead cells create space for the engraftment of competent donor cells. We have already shown that in non-irradiated mice, BMCs injected through tail vein can engraft and differentiate into hepatocytes in CCl4/acetaminophen-induced damaged liver28. Overall, the difference of results in the present study with earlier in the similar model may be attributed to the extent of kidney damage, time and the type(s) of cell transplantation.
The relative contribution of exogenous BMCs for tubular regeneration may be much lower than the intrarenal cells. Many growth factors (e.g. hepatocyte growth factor, epithelial growth factor, insulin-like growth factor-I, fibroblast growth factor-I) are known to be involved in the tubular regeneration following AKI20,29. These growth factors are thought to be synthesized by the mesenchyma of the damaged kidney. We suggest that, under the influence of these growth factors, competent cells present in the graft can differentiate into tubular epithelial cells. Indeed, in vitro experiments suggested that a fraction of Lin- cells differentiated into tubular epithelial-like cells in medium supplemented with DKS and growth factors. Since Lin- BMCs did not express megalin and CK-19 in normal culture conditions (in absence of DKS), it is concluded that the specific differentiation process is dependent on one or more unknown factors present in DKS. The BM-derived tubular epithelial cells may not be morphologically similar to the primary epithelial cell in culture. This may be due to short duration of the culture.
The results of our induced-engraftment approach suggested that a few BM-derived cells differentiated into tubular epithelial cells. On the other hand, the majority of these engrafted in the interstitial space differentiated into endothelial cells. In the forced-engraftment approach almost two-thirds of the engrafted cells differentiated into megalin-expressing tubular epithelial cells. The exact reason for the difference in outcomes between these two approaches is not know, but we envisage two explanations. First, a small number of cells that had differentiation potential were engrafted in the damaged kidney by induced-engraftment approach, whereas in the forced-engraftment approach many competent cells were engrafted. Second, in the forced-engraftment approach, the donor cells were transplanted 48 h after AKI. The regeneration of non-tubular cells is expected to be initiated by host-derived BMCs and/or intrarenal cells prior to the transplantation. Thus, a few donor-derived non-tubular cells were detected in the forced-engraftment approach.
Our results do not rule out the possibility of fusion between renal tubular epithelial cells and BMCs. Bone marrow cells have a tendency to fuse with polyploid cells such as hepatocytes and cardiomyocytes30. An earlier study reported a low frequency (<4%) of fusion in BM-derived tubular epithelial cells19. The results of in vitro experiments suggested in favour of direct differentiation of Lin- BMCs into megalin, E-cadherin and CK-19 expressing epithelial cells. Overall, this study demonstrated that BMCs were involved in the regeneration of HgCl2-induced damage tubular epithelial cells. The relative contribution of BMCs to tubular regeneration was dependent on the availability of competent cells at the site of injury, the extent of damage, and the time when BMCs were transplanted following kidney damage.
Acknowledgment
This research was funded from the core grant of the National Institute of Immunology, New Delhi.
References
- 1.Edelstein CL, Ling H, Wangsiripaisan A, Schrier RW. Emerging therapies for acute renal failure. Am J Kidney Dis. 1997;30:S89–95. doi: 10.1016/s0272-6386(97)90548-5. [DOI] [PubMed] [Google Scholar]
- 2.Kelly KJ, Molitoris BA. Acute renal failure in the new millennium: Time to consider combination therapy. Semin Nephrol. 2000;20:4–19. [PubMed] [Google Scholar]
- 3.Cochrane AL, Kett MM, Samuel CS, Campanale NV, Anderson WP, Hume DA, et al. Renal structural and functional repair in a mouse model of reversal of ureteral obstruction. J Am Soc Nephrol. 2005;16:3623–30. doi: 10.1681/ASN.2004090771. [DOI] [PubMed] [Google Scholar]
- 4.Witzgall R, Brown D, Schwarz C, Bonventre JV. Localization of proliferating cell nuclear antigen, vimentin, c-Fos, and clusterin in the postischemic kidney. Evidence for a heterogenous genetic response among nephron segments, and a large pool of mitotically active and dedifferentiated cells. J Clin Invest. 1994;93:2175–88. doi: 10.1172/JCI117214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Bonventre JV. Dedifferentiation and proliferation of surviving epithelial cells in acute renal failure. J Am Soc Nephrol. 2003;14:S55–S61. doi: 10.1097/01.asn.0000067652.51441.21. [DOI] [PubMed] [Google Scholar]
- 6.Ricardo SD, Deane JA. Adult stem cells in renal injury and repair. Nephrology (Carlton) 2005;10:276–82. doi: 10.1111/j.1440-1797.2005.00373.x. [DOI] [PubMed] [Google Scholar]
- 7.Poulsom R, Forbes SJ, Hodivala-Dilke K, Ryan E, Wyles S, Navaratnarasah S, et al. Bone marrow contributes to renal parenchymal turnover and regeneration. J Pathol. 2001;195:229–35. doi: 10.1002/path.976. [DOI] [PubMed] [Google Scholar]
- 8.Grimm PC, Nickerson P, Jeffery J, Savani RC, Gough J, McKenna RM, et al. Neointimal and tubulointerstitial infiltration by recipient mesenchymal cells in chronic renal-allograft rejection. N Engl J Med. 2001;345:93–7. doi: 10.1056/NEJM200107123450203. [DOI] [PubMed] [Google Scholar]
- 9.Ito T, Suzuki A, Imai E, Okabe M, Hori M. Bone marrow is a reservoir of repopulating mesangial cells during glomerular remodeling. J Am Soc Nephrol. 2001;12:2625–35. doi: 10.1681/ASN.V12122625. [DOI] [PubMed] [Google Scholar]
- 10.Lagaaij EL, Cramer-Knijnenburg GF, Van Kemenade FJ, Van Es L, Bruijn J, Van Krieken J. Endothelial cell chimerism after renal transplantation and vascular rejection. Lancet. 2001;357:33–7. doi: 10.1016/S0140-6736(00)03569-8. [DOI] [PubMed] [Google Scholar]
- 11.Gupta S, Verfaillie C, Chmielewski D, Kim Y, Roisenberg ME. A role for extra-renal cells in the regeneration following acute renal failure. Kidney Int. 2000;62:1285–90. doi: 10.1111/j.1523-1755.2002.kid569.x. [DOI] [PubMed] [Google Scholar]
- 12.Kale S, Karihaloo A, Clark PR, Kashgarian M, Krause DS, Cantley LG. Bone marrow stem cells contribute to repair of the ischemically injured renal tubule. J Clin Invest. 2003;112:42–9. doi: 10.1172/JCI17856. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Fang TC, Alison MR, Cook HT, Jeffery R, Wright NA, Poulsom R. Proliferation of bone marrow-derived cells contributes to regeneration after folic acid-induced acute tubular injury. J Am Soc Nephrol. 2005;16:1723–32. doi: 10.1681/ASN.2004121089. [DOI] [PubMed] [Google Scholar]
- 14.Lin F, Moran A, Igarashi P. Intra renal cells, not bone marrow-derived cells, are the major source for regeneration in postischemic kidney. J Clin Invest. 2005;115:1756–64. doi: 10.1172/JCI23015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Duffield JS, Park KM, Hsiao LL, Kelly VR, Scadden DT, Ichimura T, et al. Restoration of tubular epithelial cells during repair of the postischemic kidney occurs independently of bone marrow-derived stem cells. J Clin Invest. 2005;115:1743–55. doi: 10.1172/JCI22593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Humphreys BD, Bonventre JV. Mesenchymal stem cells in acute kidney injury. Annu Rev Med. 2008;59:311–25. doi: 10.1146/annurev.med.59.061506.154239. [DOI] [PubMed] [Google Scholar]
- 17.Harriman JF, Schnellmann RG. Mechanisms of renal cell death. In: Tarloff JB, Lash LH, editors. Toxicology of the kidney. Boca Raton Florida: CRC Press; 2004. pp. 245–98. [Google Scholar]
- 18.Jiang J, Dean D, Burghardt RC, Parrish AR. Disruption of cadherin/catenin expression, localization, and interactions during HgCl 2 -induced nephrotoxicity. Toxicol Sci. 2004;80:170–82. doi: 10.1093/toxsci/kfh143. [DOI] [PubMed] [Google Scholar]
- 19.Yen T-H, Alison MR, Cook HT, Jeffery R, Otto WR, Wright NA, et al. The cellular origin and proliferative status of regenerating renal parenchyma after mercuric chloride damage and erythropoietin treatment. Cell Prolif. 2007;40:143–56. doi: 10.1111/j.1365-2184.2007.00423.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Biemesderfer D, Nagy T, DeGray B, Aronson PS. Specific association of megalin and the Na1/H1 exchanger isoform NHE3 in the proximal tubule. J Biol Chem. 1999;274:17518–24. doi: 10.1074/jbc.274.25.17518. [DOI] [PubMed] [Google Scholar]
- 21.Boffa JJ, Just A, Coffman TM, Arendshorst WJ. Thromboxane receptor mediates renal vasoconstriction and contributes to acute renal failure in endotoxemic mice. J Am Soc Nephrol. 2004;15:2358–65. doi: 10.1097/01.ASN.0000136300.72480.86. [DOI] [PubMed] [Google Scholar]
- 22.Dekel B, Shezen E, Even-Toc-Friedman S, Katchman H, Margalit R, Nagler A, et al. Transplantation of human hematopoietic stem cells into ischemic and growing kidneys suggests a role in vasculogenesis but not tubulogenesis. Stem Cells. 2006;24:1185–93. doi: 10.1634/stemcells.2005-0265. [DOI] [PubMed] [Google Scholar]
- 23.Li L, Truong P, Igarashi P, Lin F. Renal and bone marrow cells fuse after renal ischemic injury. J Am Soc Nephrol. 2007;18:3067–77. doi: 10.1681/ASN.2007030284. [DOI] [PubMed] [Google Scholar]
- 24.Fang T-C, Otto WR, Rao J, Jeffery R, Hunt T, Alison MR, et al. Haematopoietic lineage-committed bone marrow cells, but not cloned cultured mesenchymal stem cells, contribute to regeneration of renal tubular epithelium after HgCl2 -induced acute tubular injury. Cell Prolif. 2008;41:575–91. doi: 10.1111/j.1365-2184.2008.00545.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Yokoo T, Ohashi T, Shen JS, Sakurai K, Miyazaki Y, Utsunomiya Y, et al. Human mesenchymal stem cells in rodent wholeembryo culture are reprogrammed to contribute to kidney tissues. Proc Natl Acad Sci USA. 2005;102:3296–300. doi: 10.1073/pnas.0406878102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Morigi M, Imberti B, Zoja C, Corna D, Tomasoni S, Abbate M, et al. Mesenchymal stem cells are renotropic, helping to repair the kidney and improve function in acute renal failure. J Am Soc Nephrol. 2004;15:1794–804. doi: 10.1097/01.asn.0000128974.07460.34. [DOI] [PubMed] [Google Scholar]
- 27.Fang T-C, Otto WR, Jeffery R, Hunt T, Alison MR, Cook HT, et al. Exogenous bone marrow cells do not rescue non-irradiated mice from acute renal tubular damage caused by HgCl2, despite establishment of chimaerism and cell proliferation in bone marrow and spleen. Cell Prolif. 2008;41:592–606. doi: 10.1111/j.1365-2184.2008.00546.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Yadav N, Sumod K, Kumar S, Jain M, Halder A, Saxena R, et al. The therapeutic effect of bone marrow-derived liver cells in the phenotypic correction of murine hemophilia A. Blood. 2009;114:4552–61. doi: 10.1182/blood-2009-02-202788. [DOI] [PubMed] [Google Scholar]
- 29.Wang Z, Chen K, Wang S-W, Moeckel G, Harris RC. Importance of functional EGF receptors in recovery from acute nephrotoxic injury. J Am Soc Nephrol. 2003;14:3147–54. doi: 10.1097/01.asn.0000098681.56240.1a. [DOI] [PubMed] [Google Scholar]
- 30.Alvarez-Dolado1 M, Pardal R, Garcia-Verdugo JM, Fike JR, Lee H, Pfeffer K, et al. Fusion of bone-marrow-derived cells with Purkinje neurons, cardiomyocytes and hepatocytes. Nature. 2003;425:968–73. doi: 10.1038/nature02069. [DOI] [PubMed] [Google Scholar]