

Abstract
Background
Homologous recombination, together with selection, laid the foundation for traditional plant breeding. The recombination process that takes place during meiotic cell division is crucial for the creation of novel variations of highly desired traits by breeders. Gaining control over this process is important for molecular breeding to achieve more precise, large-scale and quicker plant improvement. As conventional ubiquitous promoters are neither tissue-specific nor efficient in driving gene expression in meiocytes, promoters with high meiotic activities are potential candidates for manipulating the recombination process. So far, only a few meiotically-active promoters have been reported. Recently developed techniques to profile the transcriptome landscape of isolated meiocytes provided the means to discover promoters from genes that are actively expressed in meiosis.
Results
In a screen for meiotically-active promoters, we examined ten promoter sequences that are associated with novel meiotic candidate genes. Each promoter was tested by expressing a GFP reporter gene in Arabidopsis. Characterization of regulatory regions revealed that these meiotically-active promoters possessed conserved motifs and motif arrangement. Some of the promoters unite optimal properties which are invaluable for meiosis-directed studies such as delivering specific gene expression in early meiosis I and/or meiosis II. Furthermore, the examination of homologs of the corresponding genes within green plants points to a great potential of applying the information from Arabidopsis to other species, especially crop plants.
Conclusions
We identified ten novel meiotically-active promoters; which, along with their homologs, are prime candidates to specifically drive gene expression during meiosis in plants and can thus provide important tools for meiosis study and crop breeding.
Keywords: Meiosis, Homologous recombination, Promoter, GFP, cis-regulatory elements, Plant molecular breeding
Background
Meiosis is a key feature in the life cycle of flowering plants during which homologous chromosome pairing, synapsis and recombination are achieved [1-3]. Understanding the mechanisms of the meiotic process is crucial for not only the cell cycle regulation, but also plant breeding, because homologous recombination ensures genetic exchange between homologous chromosomes, generates the genetic variations, and maintains the inheritance of traits. Genome-wide gene expression analyses on isolated meiocytes revealed unique transcriptome-landscapes during male meiosis in the model systems of Arabidopsis and rice [4-7]. Over 1,000 protein coding genes demonstrated preferentially expression in male meiocytes, with a group of 55 genes that have mitochondrial genome origins, and 1,036 transposable element genes were up-regulated in male meiocytes [5]. The observation suggested that there is likely a specific transcription-regulatory mechanism during meiosis. As the first step toward understanding the molecular mechanism, we focus on characterizing the function and regulatory elements in selected candidate meiosis-gene promoters of this study. The objectives are to find common regulatory features in meiotically-active promoters and to explore the potential for applying the promoters in plant meiosis studies and crop breeding.
A prerequisite for meiotical engineering is the availability of effective meiotically-active promoters. However, the widely used CaMV 35 S promoter is not efficient in meiocytes. For example, AtCDC45 encodes a protein required for the normal fertility of the model plant Arabidopsis, and when an AtCDC45-RNAi construct driven by the 35 S promoter was transformed into wild type, only 20 of 59 transformants became sterile (34%), whereas a greater percentage of sterile plants (61%, 45 of 74 transformants) could be obtained by replacing the 35 S promoter with the meiosis-specific DMC1 promoter [8].
So far, only a limited number of meiotically-active promoters has been reported and investigated. The expression of the meiotic recombination gene AtDMC1 has been reported to be restricted to meiotic cells in anthers and carpels, and a β-glucuronidase (GUS) reporter fused to an AtDMC1 promoter revealed that the reporter gene activity initiated at the stages where meiosis takes place [9]. However, activity of the AtDMC1 promoter is not restricted to meiotic cells [9-11]. MS5 is a gene essential for male meiosis [12]; in situ hybridization showed that MS5 is localized specifically within anther cells undergoing meiosis [13].
In yeasts, rodents and human, the expression of genes in meiosis has been well studied [14-18]. The male meiocytes of Arabidopsis are of an extraordinary small size (1% of anther tissues) and are surrounded by somatic anther lobes, making the isolation and analysis of Arabidopsis meiocytes challenging. Recently, the application of effective meiocyte collection methods made it possible to investigate the meiotic transcriptome profile [5,6], thus allowing the bulk isolation and characterization of meiotically-active promoters. In this study, we experimentally verified the activity of twelve meiotically-active promoters out of fifteen candidate promoters, including ten new promoters.
Transcriptional regulation is critical for many developmental processes, making it important to analyze the transcriptional control to better understand the mechanisms that control spatial and temporal patterning in development [19]. The bulk isolation and characterization of meiotically-active promoters makes the study of important novel cis-regulatory motifs in these sequences feasible.
Comparative transcriptome analysis revealed similarity in meiocyte transcriptomes between organisms, for example, more than 500 single-copy genes are shared by meiotic cells of Arabidopsis[6], mouse (Mus musculus) [14] and fission yeast (Schizosaccharomyces pombe) [18], with a larger number of genes expressed in both mouse and fission yeast, or Arabidopsis and mouse. Therefore, analyzing meiotically-active promoters from Arabidopsis should provide not only clues of meiotic regulatory networks in Arabidopsis, but can also give hints for other species. These meiotically-active promoters or their close relatives could evolve into substantial tools for molecular breeding across species, especially in the plant kingdom where crop improvement is highly desired to cope with global climate changes and the increasing world population.
Results
Identification of meiotically-active promoters
A previous study performed in our labs compared the transcriptomes of whole anthers, isolated meiocytes and seedlings of Arabidopsis thaliana[5]. Among the transcripts that were found to have twofold or greater changes between both meiocytes versus seedlings and anther versus seedlings, we chose 15 candidates to further study their promoters (Table 1). These include two promoters of the previously reported meiosis-specific genes, AtDMC1[9,10] and MS5[12,13], which serve as positive controls, and were confirmed in their meiosis-preferential expression were also confirmed in our profiling study [5]. Since the 5' UTR (untranslated region) may have roles in the regulation of gene expression [20], all cloned promoters end one nucleotide ahead of the start codon of the annotated coding region. To generate a reporter system, putative promoters were amplified and cloned into the binary vector pCAMBIA1302, just upstream of the green fluorescent protein (GFP) coding sequence (Figure 1). A promoter which shows similar activity as the control promoters, marked by GFP fluorescence, is defined as a meiotically-active promoter. In addition, we checked for GFP expression in somatic cells of roots, stems and leaves of adult plants to exclude promoter activity during the major vegetative growth.
Table 1.
A List of gene IDs and associated primers for the 15 analyzed promoters
|
Promoter name |
Primer information |
|
|---|---|---|
| No. | Sequences | |
| (pATDMC1) * pAT3G22880 |
OMC1657 |
GCGTCGACTGGTAGAGTCATGTTACTTAAGGT |
| |
OMC1656 |
GGACTAGTCCATGGTCTCGCTCTAAGAGTCTCTAA |
| pAT3G19070* |
OCC555 |
ACGCGTCGACTCTTCAACATCAACCCGACC |
| OCC556 |
CATGCCATGGCTTGCAACTTAAGAAATTTGATTC |
|
| pAT2G28090* |
OCC563 |
ACGCGTCGACTGGAAACTTAATGCAAACGC |
| OCC564 |
CATGCCATGGTCGTCTCTAACTTCTTCTGC |
|
| (pMS5) * pAT4G20900 |
OCC553 |
ACGCGTCGACCTCGGCAAACGCCATAAC |
| | ||
| OCC554 |
CATGCCATGGTCTTTTTCGATTCTCTCTGTC |
|
| pAT1G26510* |
OCC557 |
ACGCGTCGACTTATTGCTCCCAACACTCG |
| OCC558 |
CATGCCATGGTCAATCGCTCTTGTTTCG |
|
| pAT2G32310 |
OCC561 |
ACGCGTCGACTACTTGGGTGCTTTCTTGTG |
| OCC562 |
CATGCCATGGTAACTTCTTTCCAAAGAATCTC |
|
| pAT3G49830* |
OCC565 |
ACGCGTCGACTAAGACTGATTTGCCAACAAGG |
| OCC566 |
CATGCCATGGTCGGTTGTTGAGTTCACC |
|
| pAT4G40020* |
OCC567 |
ACGCGTCGACGGGGTTTAGGTCTTTCCAT |
| OCC568 |
CATGCCATGGATGATAGAATGTTTTTTATTCAGC |
|
| pAT2G21640* |
OCC569 |
ACGCGTCGACTGAAAGGTTTCCCACTCC |
| OCC570 |
CATGCCATGGGAAAACAGAAAGAAATCTCATG |
|
| pAT3G07250* |
OCC571 |
AACTGCAGGCTTCGCAAATCCAACCT |
| OCC572 |
CATGCCATGGATAAAGATTCAACAAACATATAATGTC |
|
| pAT2G31141* |
OCC573 |
ACGCGTCGACCCATACAGAGTAAGCCAAACC |
| OCC574 |
CATGCCATGGCCCTCCGACTTTAGAATCC |
|
| pAT1G15320* |
OCC575 |
ACGCGTCGACCAACTCACCACCTCCCTCT |
| OCC576 |
CATGCCATGGTCGTGTTTCTTCTTCAGCACT |
|
| pAT1G64625* |
OCC577 |
ACGCGTCGACTTTCCTTGCTTGTGATCTTC |
| OCC552 |
CATGCCATGGTTAGGAATCCAAGCCGGG |
|
| pAT3G52770 |
OCC648 |
ACGCGTCGACTACAAAATGGTCCAAAACGG |
| OCC649 |
CATGCCATGGCTGCTTTCTTGCTACAAGTAAAAC |
|
| pAT1G24220 |
OCC625 |
ACGCGTCGACGAAGACATGAGATTTTGGGGTCA |
| OCC560 | CATGCCATGGCTAAACCCTCCAAG | |
* refers to meiotically-active promoter. For each primer pair, the former one is the sense primer, the later one is the antisense primer; The PstI (CTGCAG) or SalI (GTCGAC) sites in sense primers, the NcoI (CCATGG) sites in antisense primers are underlined.
Figure 1 .
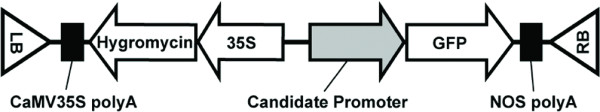
Schematic representation of the expression modules in pCAMBIA1302. CaMV35S polyA: Cauliflower mosaic virus 35 S poly-A terminator; 35 S: Cauliflower mosaic virus 35 S promoter, NOS polyA: Nopaline synthase poly-A terminator; GFP: Green fluorescent protein; Hygromycin represents the plant resistance selectable marker. LB: left border of the T-DNA, RB: right border.
Analysis of transgenic lines containing the GFP reporter driven by a candidate promoter showed that out of fifteen candidates, twelve expression modules had detectable GFP fluorescence within male meiocytes (Table 1, Figure 2 and Additional file 1, Figure S1). These include the expression modules containing the promoters of AtDMC1 and MS5 (Table 1), supporting the reliability of the GFP reporter system. In addition to these two already known promoters, ten additional meiotically-active promoters were newly identified in this study (Table 1, Figure 2 and Additional file 1, Figure S1). The particular expression pattern of the GFP modules were common to all positive lines, showing a single small and perinuclear spot per cell (Figure 2Additional file 1, Figure S1 and Additional file 2, Figure S2). In addition, we observed GFP signal in both nuclei and cytoplasm of pAtDMC1:AtDMC1:GFP meiocytes, which is similar to the CYCA1;2-GFP fusion [21]. In pAtDMC1:GFP meiocytes, however, the GFP signal was only detected in the cytoplasm and as a perinuclear green spot (data not shown), which is similar to all other tested promoter-GFP fusions in this study. No GFP signal was observed in somatic cells, such as anther walls ( Additional file 2, Figure S2). There are two visible types of meiocytes within an anther after dissection: I, cells that form columns (Figure 2a, b, e, f, I and j) contain meiocytes undergoing the prophase stages of meiosis-I; II, dissociated meiocytes (Figure 2c, d, g, h, k and l) which include meiosis-II cells. Most meiotically-active expression modules yield GFP signals in both cell types, thus spanning the whole duration of meiosis-I and II. However, module pAT1G15320:GFP only resulted in a signal in dissociated meiocytes ( Additional file 1, Figure S1), indicating expression restricted to later meiosis, whereas module pAT4G40020:GFP gives detectable signals only in cell columns ( Additional file 1, Figure S1), pointing to an early meiosis preferential expression pattern. For the ten examined promoters, no specific fluorescence signal was observed in other examined tissues, such as roots, stems and leaves of adult plants (data not shown), adding evidence to the meiotic specificity of these promoters, and the GFP signal was also not observed in meiocytes in the wild-type control (Figure 2a-d).
Figure 2 .

GFP signals in meiocytes driven by meiotically-active promoters. (a-d), wild type control meiocytes, a and c showing the normal weak background fluorescence; (e-h), pMS5:GFP meiocytes as a positive control, e and g showing GFP signals; (i-l) pAT3G19070:GFP meiocytes, i and k showing similar GFP signals as the positive control. (a, e, and i), early meiosis I cell clusters; (c, g and k), meiosis II meiocytes; images on their right are corresponding bright-field images, respectively. Scale bars, 10 μm.
In control tests, the levels of fluorescence driven by 35 S promoters were determined, including a multicolored set of organelle markers of endoplasmic reticulum, Golgi apparatus, tonoplast, peroxisomes, mitochondria and plastids [22]. The results showed that the 35 S promoter does not lead to the expression of a detectable level of fluorescence in meiotic cell columns, tetrads and pollen. As an example, Additional file 3, Figure S3 shows a signal from an endoplasmic reticulum marker [22] under the control of a 35 S promoter with dual enhancer elements (d35S) in somatic cells while being undetectable in meiotic cells.
Enriched regulatory elements in meiotically-active promoters
Gene expression is often regulated by the interaction of transcription factors and target cis-regulatory DNA elements in promoters. The identification of potential regulatory elements acting in meiotically-active promoters can be a useful tool for understanding regulatory networks [e.g. [20,23-26]]. We scanned enriched cis-acting regulatory DNA elements (CREs) in the promoters of our study to obtain clues about possible co-regulation of meiotically-active genes.
We first analyzed known plant regulatory elements in these promoter sequences by using the PLACE collection. We considered CREs within one kb upstream of these genes, since the effective regions of individual promoters are still unknown, and the normalized length also facilitated a better comparison between different sequences. The names of 17 broadly distributed CREs which appeared in all 12 promoters, and their occurrence in each promoter are shown in Figure 3a and Additional file 4, Table S1. These 17 CREs also rank in the 30 top most abundant CREs (with a total occurrence above 26, or an average occurrence above two per promoter) (Figure 3b, Additional file 4, Table S1).
Figure 3 .

Enrichedcis-acting regulatory DNA elements in meiotically-active promoters. (a), occurrence of 17 common CREs that distributed in all 12 promoters; (b), occurrence of top 30 most abundant CREs with a total occurrence above 26, or an average occurrence above two per promoter. See Additional file 4, Table S1 for description of each motif.
Several enriched PLACE motifs are universal or structural CREs that seem also common in meiotically-active promoters, such as TATABOX5 [27], POLASIG1 and POLASIG3 [28-30]. Interestingly, ROOTMOTIFTAPOX1 [31], NODCON2GM [32], RAV1AAT [33], OSE2ROOTNODULE [34,35] are all consensus CREs in root and nodule, pointing to a common property of these cells and meiocytes, likely due to their being either in the mitotic or the meiotic process. Many CREs are environment responsive motifs, for example, MYCCONSENSUSAT for cold [36-38], WRKY71OS for gibberellin and pathogenesis [39,40], ACGTATERD1 and IBOXCORE for light [41,42], MYBCORE for water stress [43], GT1GMSCAM4 for pathogen and salt [44] and WBOXNTERF3 for wounding [45]. Additionally, there is a high similarity to motifs in the promoters of rice sperm-cell-specific genes: the examined meiotically-active promoters share 9 out of 10 common motifs associated with rice sperm cell-specific genes, whereas one detected motif (ROOTMOTIFTAPOX1, ATATT) was only shown in sperm cell-specific genes [46].
To complement searches for possible motifs that have a statistically overrepresented frequency in the genome, we computationally analyzed the twelve promoters using Pscan [47]. As a result, six motifs were found enriched within the twelve promoters (p < 0.1, Figure 4 and Additional file 5, Table S2).
Figure 4 .

Sequence logos of overrepresented sequences in meiotically active promoters, found by Pscan. Letters abbreviating the nucleotides (A,C,G,T) in the images are sized relative to their occurrence. See Additional file 5, Table S2 for description of each motif.
Among these motifs (Figure 4 and Additional file 5, Table S2), MA0044.1 (p = 1.52751e-02) and MA0045.1 (p = 4.32294e-02) are binding sites for the chromatin-associated proteins HMG-1 and HMG-I/Y [48]. The high-mobility group proteins (HMG) are a group of chromosomal proteins that help with transcription, replication, recombination and DNA repair [49]. MA121.1 (p = 9.30809e-02) is a binding site of ARR10, whose multifunctional domain is responsible for both nuclear localization and DNA binding [50]; MA0096.1 (p = 9.96644e-02) is a binding site of two flower-specific bZIP proteins [51]. These motifs are likely basic elements that confer tissue- or developmental stage-specific activities to their promoters. Additionally, the motifs MA0128.1 (p = 9.03657e-03) and MA0129.1 (p = 4.67192e-02), which were implicated in abscisic acid (ABA)-mediated stress and light signaling [52], respectively, are found enriched in these promoters ( Additional file 5, Table S2), consistent with the notion that the meiotic process is sensitive to environmental factors and exogenous hormones including light and ABA [e.g. [52-55]].
Furthermore, we used MEME software to search for possible unknown CREs [56]. Three consensus motifs were found present in these promoter sequences ( Additional file 6, Figure S4). These motifs are characterized by enrichment of adenine (or thymine in the reverse complement strands) ( Additional file 6, Figure S4). Similar results were obtained with MClip tool [57] (data not shown). These adenine-rich motifs could be specific binding sites of transcriptional factors and enhancers [58]. Interestingly, the adenine-rich motifs were also found in promoter regions of 15 selected genes with a documented function in meiosis ( Additional file 7, Table S3 and Additional file 8, Figure S5).
Homologs of the examined meiotically-active genes
To know whether these meiotically-active genes have homologous sequences in other plant species, especially in crops, we investigated the “family history” of the 12 genes whose promoters were characterized in this study. As is shown in Figure 5, AT3G19070, AT3G49830 and AT2G3114 seem to be specific to Arabidopsis thaliana alone while AT3G07250 has at least a putative homolog in Arabidopsis lyrata. The other 8 genes are broadly distributed in different taxa, mostly found in flowering plants (Angiosperm), including many important crop species such as soybean (Glycine max), maize (Zea mays), rice (Oryza sativa) and Sorghum (Sorghum bicolor). Therefore, we assume that for these genes, there is a higher possibility that similar regulatory networks may be shared among different flowering plants.
Figure 5 .

Examination of putative homologs of meiotically-active genes in green plants. The upper panel displays the evolutionary clades represented by complete genome sequences in the Phytozome database. The evolutionary distance is shown on the left of the upper panel, with Viridiplantae representing all plants, and extending down into deeper evolutionary distances. For example, Arabidopsis is part of the Rosid clade and maize (Zea mays) is part of the Grass clade. The individual genes whose promoters were examined are shown in the lower panel, and the grey shading in cells indicates the breadth of their representation in plants. The number of genes similar to the gene whose promoter was characterized in that plant species is shown by the number in the box.
Discussion
The objectives of this study were to identify functional promoters that drive gene expression in meiosis and to find the common cis-regulatory elements that are present in all promoters. Results for meiosis-I, during which homologous recombination occurs, are of special interest to predict homologous recombination-related promoters in crop plants. We have tested 15 promoters that are associated with candidate meiosis genes that were discovered by a previous RNA-Seq experiment on isolated meiocytes [5], which include 13 promoters of functionally unknown genes and two reported meiotic gene promoters (pDMC1 and pMS5). Among the 13 candidate promoters that have no documented function in meiosis, ten have shown meiotic activity (≈77%) by driving the expression of GFP signal in meiocytes, thus revealing that our preliminary data has a high reliability for isolating meiotically-active promoters (Table 1). No GFP signal was observed in three transgenic lines of the candidate promoters (Table 1), although their respective gene transcripts were up-regulated in meiocytes in the RNA-Seq study. This may be attributed to distinctions in developmental age, time of harvest, sensitivity of the used method, the chosen promoter region or that these promoters function in a chromosome positional dependent manner [59,60]. Nevertheless, our work provided a significant number of promoters that can drive gene expression in meiosis. These promoters could evolve to be invaluable tools to drive meiotically-active expression in further fundamental meiosis studies as well as in applied molecular breeding.
Until now, most researchers use ubiquitous promoters such as the CaMV 35 S promoter to over-express genes in plants for functional analysis [61]. Those “ubiquitous” promoters, however, are inadequate for meiotic purposes, because they drive gene expression in meiosis at an insufficient level [62]. For example, fluorescent organelle markers [22] that are driven by a 35 S promoter demonstrated no signal in meiosis stages ( Additional file 3, Figure S3), although there is a strong GFP signal detected in somatic cells ( Additional file 3, Figure S3). In accordance with that, RNAi knock-down of a meiotic mutant is also achieved better by using the meiosis-specific DMC1 promoter than by using the 35 S promoter [61% vs 34%, [8]]. However, even the established and broadly used DMC1 promoter has its disadvantages in meiosis studies: Doutriaux et al. reported that AtDMC1 is expressed in mitotically active cells from a suspension culture and is even regulated during the mitotic cell cycle, linking it to the processes in proliferating cells [10]. Furthermore, the AtDMC1 promoter has been used for studies in young seedlings, yielding an efficient expression in a recombination reporter system [11], which is in accordance with the expression data for AtDMC1 obtained with ATH1 microarray chips: The eFP Browser tool displays that DMC1 is also highly expressed in vegetative rosette leaves and especially in the shoot apex and seedling roots [63]. Thus, there is no ultimately optimal meiosis-specific promoter in broad use yet.
The novel candidate meiotically-active promoters from this study should provide more powerful tools for a strict or specified meiotic expression. In our experimental setup, we first chose genes that are highly expressed in meiocytes [5]. We then relied not only on the positive expression that we got with our GFP reporter in meiotic cells but also looked at other tissues, e.g. roots, leaves and stems to validate its nonexistence there. Therefore we defined the promoters here as “meiotically-active” or “homologous recombination-related”, although we cannot completely exclude promoter activity in specific developmental stages or special conditions not covered or detectable by our setup. The decision of which promoter might be best depends on the special application and the preferences of the user, for example if a low or high expression is desired or if the expression should be restricted to a very specific time point in meiosis.
Interestingly, we observed diversified expression patterns in different cell types of meiocytes (cell columns and dissociated meiocytes) resulting from the examined promoters (Table 1). Transgenic lines harboring pAT1G15320:GFP only showed a specific fluorescence signal in dissociated meiocytes but not in meiocyte cell columns ( Additional file 1, Figure S1), which suggested a preferential activity in meiosis-II or after homologous recombination. In contrast, pAT4G40020:GFP plants showed detectable GFP signals only in early meiosis-I meiocytes, pointing to a homologous recombination-specific promoter. In addition to our results that pAT4G40020 drives gene expression at a high level during early meiosis, microarray data of developmental stages indicates that At4G40020 is further expressed only in microspores [eFP Browser, [63]]. There is also microarray data available for some of our other candidate genes, but not for all of them. Thus, we can confirm their meiosis-specific expression with our experimental setup but cannot completely rule out expression outside meiosis under special conditions or in specific developmental stages. Taken together, we have identified and validated 12 meiotically-active promoters and two of these promoters can be used to specifically address questions regarding roughly meiosis-I (pAT4G40020) or meiosis-II (pAT1G15320). For molecular engineering, expressing genes during prophase-I, the stage of recombination, will be of utmost interest.
Given the complexity and a relatively long duration of meiosis (for example, prophase I lasts 21.3 h) [64], the temporal specificity of different promoters might be even more confined to individual meiotic stages. In future work, it will be important to test this possibility and investigate the expression even closer to obtain stage-specific promoters which are powerful tools to meet different requirements.
The confirmation of the meiotic activity of the examined promoters also points to a meiotic function of the respective genes. In addition to the already characterized genes AtDMC1 and MS5[9,12,65,66], we have discovered an additional key gene with a role during meiosis by checking the T-DNA insertion mutants for the ten genes without documented function (unpublished data).
The gene transcription in eukaryotes is complex and is largely modulated by transcription factors that bind to regulatory elements within promoters. We scanned the identified promoter set for motifs with binding specificity for known transcription factors from the PLACE collection (Figure 3 and Additional file 4, Table S1) and used the software tool Pscan (Figure 4 and Additional file 5, Table S2). CREs that are common to the meiotically-active promoters from this study may reflect common binding sites for certain transcription factors that are required for meiotic activities (such as the binding sites of HMG-1, HMG-I/Y, ARR10 and bZIP910, Additional file 5, Table S2). It also provides a hint as to know how these promoters are shared by stimulus–response pathways (such as the binding sites of EMBP-1 and TGA1A, Additional file 5, Table S2). We also analyzed the promoters of 15 genes with a documented function in meiosis ( Additional file 7, Table S3) with Pscan. Although they are not all meiosis-specific under the criteria used in [5] Chen et al. (2010), the identified common elements include not only “basic element” such as HMG-1 binding sites, but also binding sites for proteins involved in gibberellin response and leaf development ( Additional file 9, Table S4). Therefore, it appears that the crosstalk between meiosis and environmental signals, especially hormone signals, are largely through their promoters. These identified CREs can also be further used to design the experimental verification of regulatory elements and the identification of transcriptional factors that regulate meiotically-active gene expression [46].
Since meiosis is a conserved process in all sexually reproducing eukaryotes, knowledge of gene function from one species could provide useful information transferable to other species. For example, studies in budding yeast (Saccharomyces cerevisiae) have revealed that a MER DNA helicase is required for the interference-sensitive pathway for crossover formation [67-71], and this finding led to the identification of a MER3 homolog, ROCK-N-ROLLERS (RCK) in Arabidopsis, supporting that as in budding yeast, both the interference-sensitive and insensitive pathways of recombination crossovers exist in plants [72,73]. Analysis of the “family history” of the meiotically-active genes from our study found a wide distribution of homologous sequences in many species in green plants (Viridiplantae), especially in flowering plants (Figure 5). This result suggests a great prospect of transferring the information obtained from Arabidopsis into other plants, including important crops such as soybean, maize, rice and Sorghum. Since low copies of putative homologous genes of AtDMC1, AT4G40020 and AT2G21640 seem to exist (Figure 5), exploring their correspondent promoter sequences in other species should be quite straightforward. For AT2G28090, MS5, AT1G26510, AT1G15320 and AT1G64625, many homologous genes were found in other plants and which might make it more difficult to elucidate “true” homologs and use their promoters; a more appropriate strategy in this case is to try to extend the usage of the promoter sequences from Arabidopsis directly to other plants.
In conclusion, we report here a bulk identification and experimental verification of meiotically-active promoters. The information provides not only invaluable clues about the meiotic regulatory system, but also a potential tool for the application in model and crop plants. In future work, it will be interesting and important to explore the relative activity levels of each promoter.
Conclusions
In conclusion, the ten isolated promoter sequences were confirmed to specifically drive gene expression in meiocytes. The findings can provide important tools for meiosis studies and crop breeding, especially two of these promoters are prime candidates for meiosis directed-studies that desire an expression focused on early meiosis-I and/or meiosis-II.
Methods
Plant material and growth conditions
Arabidopsis thaliana (L.) Heynh. Ecotype Columbia (Col-0) was used in this study. Seeds were sown on 50% Sun Gro Professional Growing Mix and 50% Sun Gro SPECIAL BLEND growing medium (Sun Gro Horticulture, USA) and imbibed at 4°C for 3 days in the dark before moving them to long-day conditions (16 h light/8 h dark) at 22°C, 40 to 60% RH and 63 mE`·s-1·m-2 light intensity. The ER marker line ER-gk was obtained from the Arabidopsis Biological Resource Center (ABRC) [74].
Examination of GFP fluorescence
Young inflorescences were dissected with syringe needles and the anthers were collected using 1xPBS buffer. The meiocytes were squashed out between a microscope slide and a cover glass. The GFP fluorescence was observed under an ERNST LEITZ WETZLAR 307–143.004 microscope (Wetzlar, Germany) and photographed with a SPOT Insight 4 Camera (Diagnostic Instruments, USA).
Cloning, vector construction and plant transformation
To clone the promoters, the genomic sequences upstream of the candidate genes’ start codons were amplified using specific primers as listed in Table 1, in which SalI or PstI restriction sites were introduced for sense primers and NcoI sites for antisense primers. In order to prevent overlap with neighbouring genes or to get the appropriate primer binding sites, the cloned regions ranged from 0.6 Kb to 1.8 Kb, covering the key cis-elements of most promoters [75,76]. The DMC1 promoter, which has a length of 3.0 Kb, is an exception, since it was cloned in our previous unpublished study. The promoters were cloned into the pCAMBIA1302 expression vector that contains the mgfp5 version of the Aequoria victoria GFP [77], substituting the CaMV 35 S promoter upstream of the GFP coding sequence. The sequences were confirmed and the plasmids were introduced into Agrobacterium tumefaciens C58. To test the constructs in planta, all plasmids were introduced to the Arabidopsis plants using a floral dip method [78]. Transgenic plants were first screened on medium containing 40 mg/l hygromycin and transferred to soil, and further validated by PCR with the sense primers used in the promoter cloning (Table 1) and an antisense primer near the 5’ end of GFP (GTT GCA TCA CCT TCA CCC TCT).
Analysis of cis-regulatory promoter elements
Known CREs were found by analysis with the PLACE database (http://www.dna.affrc.go.jp/PLACE/) [79-81]. The Pscan program was used to search for significantly overrepresented elements http://159.149.109.9/pscan/, [47]]. 1000 bp regions with respect to the annotated transcription start site of promoters were analyzed. The frequency matrices and visual logos of the sequences were obtained from the JASPAR CORE database. The p-values were computed by Pscan after z-test. An element was considered to be significantly overrepresented if the p-value was less than 0.1.
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
C.C. and E.F.R. conceived the project, designed the experiments and helped to draft the manuscript. J.L. carried out the molecular genetic studies, cis-element studies and drafted the manuscript. J.L. and C.C. performed the GFP reporter assay. A.D.F, I.E.L. and J.M. performed the homologs study. S.D. performed the endoplasmic reticulum marker assay and helped to draft the manuscript. T.L. helped in plant transformation and plant care. All authors read and approved the final manuscript.
Supplementary Material
Figure S1.GFP signals in meiocytes driven by meiotically-active promoters. (PDF 2536 kb).
Figure S2.The position of GFP spots and nuclear DNA/chromosomes in pMS5: GFP transformants. (PDF 636 kb).
Figure S3.GFP signals in both somatic and meiotic cells driven by a constitutive promoter d35S. (PDF 388 kb).
Table S1.PLACE motifs enriched in promoters of meiotically-active genes. (PDF 12 kb).
Table S2.Statistically overrepresented motifs in the promoters of meiotically-active genes. (PDF 8 kb).
Figure S4.Sequence logos of novel motifs in meiotically-active promoters found by MEME. (PDF 213 kb).
Table S3.A list of homologous recombination-related genes for comparative promoter analysis. (PDF 7 kb).
Figure S5.Sequence logos of novel motifs in promoters of homologous recombination-related genes found by MEME. (PDF 68 kb).
Table S4.Motifs enriched specifically in 15 known meiosis-related promoters. (PDF 7 kb)
Contributor Information
Junhua Li, Email: lijh0909@gmail.com.
Andrew D Farmer, Email: adf@ncgr.org.
Ingrid E Lindquist, Email: iel@ncgr.org.
Stefanie Dukowic-Schulze, Email: sdukowic@umn.edu.
Joann Mudge, Email: jm@ncgr.org.
Tao Li, Email: lixx0765@umn.edu.
Ernest F Retzel, Email: efr@ncgr.org.
Changbin Chen, Email: chenx481@umn.edu.
Acknowledgements
We are grateful for the research funds that are provided by the Biotechnology Research and Development Corporation (BRDC) to CC, EFR. CC and EFR are also supported by a NSF-PGRP grant (ISO1025881). The authors would like to thank two anonymous reviewers for their valuable comments and suggestions to improve the quality of the paper, and Ross Peterson, Duane McDowell, Doug Brinkman and Roger Meissner for plant care.
References
- Ma H. A molecular portrait of Arabidopsis meiosis. The Arabidopsis Book. 2006;4(1):1–39. doi: 10.1199/tab.0095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ronceret A, Sheehan M, Pawlowski W. Chromosome dynamics in meiosis. Verma DPS, Hong Z, Berlin/ Heidelberg: Springer-Verlag; 2007. pp. 103–124. (In Plant Cell Monogr (9), Cell Division Control in Plants). [Google Scholar]
- Yanowitz J. Meiosis: making a break for it. Curr Opin Cell Biol. 2010;22(6):744–751. doi: 10.1016/j.ceb.2010.08.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tang X, Zhang ZY, Zhang WJ, Zhao XM, Li X, Zhang D, Liu QQ, Tang WH. Global gene profiling of laser-captured pollen mother cells indicates molecular pathways and gene subfamilies involved in rice meiosis. Plant Physiol. 2010;154(4):1855–1870. doi: 10.1104/pp.110.161661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen C, Farmer AD, Langley RJ, Mudge J, Crow JA, May GD, Huntley J, Smith AG, Retzel EF. Meiosis-specific gene discovery in plants: RNA-Seq applied to isolated Arabidopsis male meiocytes. BMC Plant Biol. 2010;10:280. doi: 10.1186/1471-2229-10-280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang H, Lu P, Wang Y, Ma H. The transcriptome landscape of Arabidopsis male meiocytes from high-throughput sequencing: the complexity and evolution of the meiotic process. Plant J. 2011;65(4):503–516. doi: 10.1111/j.1365-313X.2010.04439.x. [DOI] [PubMed] [Google Scholar]
- Deveshwar P, Bovill WD, Sharma R, Able JA, Kapoor S. Analysis of anther transcriptomes to identify genes contributing to meiosis and male gametophyte development in rice. BMC Plant Biol. 2011;11:78. doi: 10.1186/1471-2229-11-78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stevens R, Grelon M, Vezon D, Oh J, Meyer P, Perennes C, Domenichini S, Bergounioux C. A CDC45 homolog in Arabidopsis is essential for meiosis, as shown by RNA interference-induced gene silencing. Plant Cell. 2004;16(1):99–113. doi: 10.1105/tpc.016865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klimyuk VI, Jones JD. AtDMC1, the Arabidopsis homologue of the yeast DMC1 gene: characterization, transposon-induced allelic variation and meiosis-associated expression. Plant J. 1997;11(1):1–14. doi: 10.1046/j.1365-313X.1997.11010001.x. [DOI] [PubMed] [Google Scholar]
- Doutriaux MP, Couteau F, Bergounioux C, White C. Isolation and characterisation of the RAD51 and DMC1 homologs from Arabidopsis thaliana. Mol Gen Genet. 1998;257(3):283–291. doi: 10.1007/s004380050649. [DOI] [PubMed] [Google Scholar]
- Orel N, Kyryk A, Puchta H. Different pathways of homologous recombination are used for the repair of double-strand breaks within tandemly arranged sequences in the plant genome. Plant J. 2003;35(5):604–612. doi: 10.1046/j.1365-313X.2003.01832.x. [DOI] [PubMed] [Google Scholar]
- Glover J, Grelon M, Craig S, Chaudhury A, Dennis E. Cloning and characterization of MS5 from Arabidopsis: a gene critical in male meiosis. Plant J. 1998;15(3):345–356. doi: 10.1046/j.1365-313X.1998.00216.x. [DOI] [PubMed] [Google Scholar]
- Sanders P, Bui A, Weterings K, McIntire K, Hsu Y, Lee P, Truong M, Beals T, Goldberg R. Anther developmental defects in Arabidopsis thaliana male-sterile mutants. Sexual Plant Reproduction. 1999;11(6):297–322. doi: 10.1007/s004970050158. [DOI] [Google Scholar]
- Chalmel F, Rolland AD, Niederhauser-Wiederkehr C, Chung SS, Demougin P, Gattiker A, Moore J, Patard JJ, Wolgemuth DJ, Jegou B. et al. The conserved transcriptome in human and rodent male gametogenesis. Proc Natl Acad Sci U S A. 2007;104(20):8346–8351. doi: 10.1073/pnas.0701883104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chu S, DeRisi J, Eisen M, Mulholland J, Botstein D, Brown PO, Herskowitz I. The transcriptional program of sporulation in budding yeast. Science. 1998;282(5389):699–705. doi: 10.1126/science.282.5389.699. [DOI] [PubMed] [Google Scholar]
- Juneau K, Palm C, Miranda M, Davis RW. High-density yeast-tiling array reveals previously undiscovered introns and extensive regulation of meiotic splicing. Proc Natl Acad Sci U S A. 2007;104(5):1522–1527. doi: 10.1073/pnas.0610354104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Primig M, Williams RM, Winzeler EA, Tevzadze GG, Conway AR, Hwang SY, Davis RW, Esposito RE. The core meiotic transcriptome in budding yeasts. Nat Genet. 2000;26(4):415–423. doi: 10.1038/82539. [DOI] [PubMed] [Google Scholar]
- Wilhelm BT, Marguerat S, Watt S, Schubert F, Wood V, Goodhead I, Penkett CJ, Rogers J, Bahler J. Dynamic repertoire of a eukaryotic transcriptome surveyed at single-nucleotide resolution. Nature. 2008;453(7199):1239–1243. doi: 10.1038/nature07002. [DOI] [PubMed] [Google Scholar]
- Watanabe K, Okada K. Two discrete cis elements control the Abaxial side-specific expression of the FILAMENTOUS FLOWER gene in Arabidopsis. Plant Cell. 2003;15(11):2592–2602. doi: 10.1105/tpc.015214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pickering BM, Willis AE. The implications of structured 5' untranslated regions on translation and disease. Semin Cell Dev Biol. 2005;16(1):39–47. doi: 10.1016/j.semcdb.2004.11.006. [DOI] [PubMed] [Google Scholar]
- Wang Y, Magnard JL, McCormick S, Yang M. Progression through meiosis I and meiosis II in Arabidopsis anthers is regulated by an A-type cyclin predominately expressed in prophase I. Plant Physiol. 2004;136(4):4127. doi: 10.1104/pp.104.051201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nelson BK, Cai X, Nebenfuhr A. A multicolored set of in vivo organelle markers for co-localization studies in Arabidopsis and other plants. Plant J. 2007;51(6):1126–1136. doi: 10.1111/j.1365-313X.2007.03212.x. [DOI] [PubMed] [Google Scholar]
- DeRisi JL, Iyer VR, Brown PO. Exploring the metabolic and genetic control of gene expression on a genomic scale. Science. 1997;278(5338):680–686. doi: 10.1126/science.278.5338.680. [DOI] [PubMed] [Google Scholar]
- Harmer SL, Hogenesch JB, Straume M, Chang HS, Han B, Zhu T, Wang X, Kreps JA, Kay SA. Orchestrated transcription of key pathways in Arabidopsis by the circadian clock. Science. 2000;290(5499):2110–2113. doi: 10.1126/science.290.5499.2110. [DOI] [PubMed] [Google Scholar]
- Nemhauser JL, Mockler TC, Chory J. Interdependency of brassinosteroid and auxin signaling in Arabidopsis. PLoS Biol. 2004;2(9):E258. doi: 10.1371/journal.pbio.0020258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tullai JW, Schaffer ME, Mullenbrock S, Kasif S, Cooper GM. Identification of transcription factor binding sites upstream of human genes regulated by the phosphatidylinositol 3-kinase and MEK/ERK signaling pathways. J Biol Chem. 2004;279(19):20167–20177. doi: 10.1074/jbc.M309260200. [DOI] [PubMed] [Google Scholar]
- Tjaden G, Edwards JW, Coruzzi GM. Cis elements and trans-acting factors affecting regulation of a nonphotosynthetic light-regulated gene for chloroplast glutamine synthetase. Plant Physiol. 1995;108(3):1109–1117. doi: 10.1104/pp.108.3.1109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Joshi CP. Putative polyadenylation signals in nuclear genes of higher plants: a compilation and analysis. Nucleic Acids Res. 1987;15(23):9627–9640. doi: 10.1093/nar/15.23.9627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Loke JC, Stahlberg EA, Strenski DG, Haas BJ, Wood PC, Li QQ. Compilation of mRNA polyadenylation signals in Arabidopsis revealed a new signal element and potential secondary structures. Plant Physiol. 2005;138(3):1457–1468. doi: 10.1104/pp.105.060541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O'Neill SD, Kumagai MH, Majumdar A, Huang N, Sutliff TD, Rodriguez RL. The alpha-amylase genes in Oryza sativa: characterization of cDNA clones and mRNA expression during seed germination. Mol Gen Genet. 1990;221(2):235–244. doi: 10.1007/BF00261726. [DOI] [PubMed] [Google Scholar]
- Elmayan T, Tepfer M. Evaluation in tobacco of the organ specificity and strength of the rolD promoter, domain A of the 35 S promoter and the 35 S2 promoter. Transgenic Res. 1995;4(6):388–396. doi: 10.1007/BF01973757. [DOI] [PubMed] [Google Scholar]
- Stougaard J, Jorgensen JE, Christensen T, Kuhle A, Marcker KA. Interdependence and nodule specificity of cis-acting regulatory elements in the soybean leghemoglobin lbc3 and N23 gene promoters. Mol Gen Genet. 1990;220(3):353–360. doi: 10.1007/BF00391738. [DOI] [PubMed] [Google Scholar]
- Kagaya Y, Ohmiya K, Hattori T. RAV1, a novel DNA-binding protein, binds to bipartite recognition sequence through two distinct DNA-binding domains uniquely found in higher plants. Nucleic Acids Res. 1999;27(2):470–478. doi: 10.1093/nar/27.2.470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fehlberg V, Vieweg MF, Dohmann EM, Hohnjec N, Puhler A, Perlick AM, Kuster H. The promoter of the leghaemoglobin gene VfLb29: functional analysis and identification of modules necessary for its activation in the infected cells of root nodules and in the arbuscule-containing cells of mycorrhizal roots. J Exp Bot. 2005;56(413):799–806. doi: 10.1093/jxb/eri074. [DOI] [PubMed] [Google Scholar]
- Vieweg MF, Fruhling M, Quandt HJ, Heim U, Baumlein H, Puhler A, Kuster H, Andreas MP. The promoter of the Vicia faba L. leghemoglobin gene VfLb29 is specifically activated in the infected cells of root nodules and in the arbuscule-containing cells of mycorrhizal roots from different legume and nonlegume plants. Mol Plant Microbe Interact. 2004;17(1):62–69. doi: 10.1094/MPMI.2004.17.1.62. [DOI] [PubMed] [Google Scholar]
- Abe H, Urao T, Ito T, Seki M, Shinozaki K, Yamaguchi-Shinozaki K. Arabidopsis AtMYC2 (bHLH) and AtMYB2 (MYB) function as transcriptional activators in abscisic acid signaling. Plant Cell. 2003;15(1):63–78. doi: 10.1105/tpc.006130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chinnusamy V, Ohta M, Kanrar S, Lee BH, Hong X, Agarwal M, Zhu JK. ICE1: a regulator of cold-induced transcriptome and freezing tolerance in Arabidopsis. Genes Dev. 2003;17(8):1043–1054. doi: 10.1101/gad.1077503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hartmann U, Sagasser M, Mehrtens F, Stracke R, Weisshaar B. Differential combinatorial interactions of cis-acting elements recognized by R2R3-MYB, BZIP, and BHLH factors control light-responsive and tissue-specific activation of phenylpropanoid biosynthesis genes. Plant Mol Biol. 2005;57(2):155–171. doi: 10.1007/s11103-004-6910-0. [DOI] [PubMed] [Google Scholar]
- Eulgem T, Rushton PJ, Schmelzer E, Hahlbrock K, Somssich IE. Early nuclear events in plant defence signalling: rapid gene activation by WRKY transcription factors. EMBO J. 1999;18(17):4689–4699. doi: 10.1093/emboj/18.17.4689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang ZL, Xie Z, Zou X, Casaretto J, Ho TH, Shen QJ. A rice WRKY gene encodes a transcriptional repressor of the gibberellin signaling pathway in aleurone cells. Plant Physiol. 2004;134(4):1500–1513. doi: 10.1104/pp.103.034967. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bovy A, Van den Berg C, De Vrieze G, Thompson WF, Weisbeek P, Smeekens S. Light-regulated expression of the Arabidopsis thaliana ferredoxin gene requires sequences upstream and downstream of the transcription initiation site. Plant Mol Biol. 1995;27(1):27–39. doi: 10.1007/BF00019176. [DOI] [PubMed] [Google Scholar]
- Simpson SD, Nakashima K, Narusaka Y, Seki M, Shinozaki K, Yamaguchi-Shinozaki K. Two different novel cis-acting elements of erd1, a clpA homologous Arabidopsis gene function in induction by dehydration stress and dark-induced senescence. Plant J. 2003;33(2):259–270. doi: 10.1046/j.1365-313X.2003.01624.x. [DOI] [PubMed] [Google Scholar]
- Urao T, Yamaguchi-Shinozaki K, Urao S, Shinozaki K. An Arabidopsis myb homolog is induced by dehydration stress and its gene product binds to the conserved MYB recognition sequence. Plant Cell. 1993;5(11):1529–1539. doi: 10.1105/tpc.5.11.1529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park HC, Kim ML, Kang YH, Jeon JM, Yoo JH, Kim MC, Park CY, Jeong JC, Moon BC, Lee JH. et al. Pathogen- and NaCl-induced expression of the SCaM-4 promoter is mediated in part by a GT-1 box that interacts with a GT-1-like transcription factor. Plant Physiol. 2004;135(4):2150–2161. doi: 10.1104/pp.104.041442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nishiuchi T, Shinshi H, Suzuki K. Rapid and transient activation of transcription of the ERF3 gene by wounding in tobacco leaves: possible involvement of NtWRKYs and autorepression. J Biol Chem. 2004;279(53):55355–55361. doi: 10.1074/jbc.M409674200. [DOI] [PubMed] [Google Scholar]
- Sharma N, Russell SD, Bhalla PL, Singh MB. Putative cis-regulatory elements in genes highly expressed in rice sperm cells. BMC Res Notes. 2011;4:319. doi: 10.1186/1756-0500-4-319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zambelli F, Pesole G, Pavesi G. Pscan: finding over-represented transcription factor binding site motifs in sequences from co-regulated or co-expressed genes. Nucleic Acids Res. 2009;37(Web Server issue):W247–W252. doi: 10.1093/nar/gkp464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Webster CI, Packman LC, Pwee KH, Gray JC. High mobility group proteins HMG-1 and HMG-I/Y bind to a positive regulatory region of the pea plastocyanin gene promoter. Plant J. 1997;11(4):703–715. doi: 10.1046/j.1365-313X.1997.11040703.x. [DOI] [PubMed] [Google Scholar]
- Rajeswari MR, Jain A. High Mobility Group chromosomal proteins, HMGA1 as potential tumor markers. Current Sci. 2002;82:838–844. [Google Scholar]
- Hosoda K, Imamura A, Katoh E, Hatta T, Tachiki M, Yamada H, Mizuno T, Yamazaki T. Molecular structure of the GARP family of plant Myb-related DNA binding motifs of the Arabidopsis response regulators. Plant Cell. 2002;14(9):2015–2029. doi: 10.1105/tpc.002733. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martinez-Garcia JF, Moyano E, Alcocer MJ, Martin C. Two bZIP proteins from Antirrhinum flowers preferentially bind a hybrid C-box/G-box motif and help to define a new sub-family of bZIP transcription factors. Plant J. 1998;13(4):489–505. doi: 10.1046/j.1365-313X.1998.00050.x. [DOI] [PubMed] [Google Scholar]
- Niu X, Renshaw-Gegg L, Miller L, Guiltinan MJ. Bipartite determinants of DNA-binding specificity of plant basic leucine zipper proteins. Plant Mol Biol. 1999;41(1):1–13. doi: 10.1023/A:1006206011502. [DOI] [PubMed] [Google Scholar]
- Jain M, Tyagi AK, Khurana JP. Constitutive expression of a meiotic recombination protein gene homolog, OsTOP6A1, from rice confers abiotic stress tolerance in transgenic Arabidopsis plants. Plant Cell Rep. 2008;27(4):767–778. doi: 10.1007/s00299-007-0491-8. [DOI] [PubMed] [Google Scholar]
- Lu BC. Genetic Recombination in Coprinus. IV. a Kinetic Study of the Temperature Effect on Recombination Frequency. Genetics. 1974;78(2):661–677. doi: 10.1093/genetics/78.2.661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu BC. The control of meiosis progression in the fungus Coprinus cinereus by light/dark cycles. Fungal Genet Biol. 2000;31(1):33–41. doi: 10.1006/fgbi.2000.1229. [DOI] [PubMed] [Google Scholar]
- Bailey TL, Boden M, Buske FA, Frith M, Grant CE, Clementi L, Ren J, Li WW, Noble WS. MEME SUITE: tools for motif discovery and searching. Nucleic Acids Res. 2009;37(Web Server issue):W202–W208. doi: 10.1093/nar/gkp335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frickey T, Weiller G. Mclip: motif detection based on cliques of gapped local profile-to-profile alignments. Bioinformatics. 2007;23(4):502–503. doi: 10.1093/bioinformatics/btl601. [DOI] [PubMed] [Google Scholar]
- Butler JE, Kadonaga JT. The RNA polymerase II core promoter: a key component in the regulation of gene expression. Genes Dev. 2002;16(20):2583–2592. doi: 10.1101/gad.1026202. [DOI] [PubMed] [Google Scholar]
- Alvarez M, Rhodes SJ, Bidwell JP. Context-dependent transcription: all politics is local. Gene. 2003;313:43–57. doi: 10.1016/s0378-1119(03)00627-9. [DOI] [PubMed] [Google Scholar]
- Serov O, Serova I. Genes and chromosomes: control of development. An Acad Bras Cienc. 2004;76(3):529–540. doi: 10.1590/S0001-37652004000300007. [DOI] [PubMed] [Google Scholar]
- Odell JT, Nagy F, Chua NH. Identification of DNA sequences required for activity of the cauliflower mosaic virus 35 S promoter. Nature. 1985;313(6005):810–812. doi: 10.1038/313810a0. [DOI] [PubMed] [Google Scholar]
- Fang RX, Nagy F, Sivasubramaniam S, Chua NH. Multiple cis regulatory elements for maximal expression of the cauliflower mosaic virus 35 S promoter in transgenic plants. Plant Cell. 1989;1(1):141–150. doi: 10.1105/tpc.1.1.141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Winter D, Vinegar B, Nahal H, Ammar R, Wilson GV, Provart NJ. An ‘Electronic Fluorescent Pictograph’ browser for exploring and analyzing large-scale biological data sets. PLoS One. 2007;2(8):e718. doi: 10.1371/journal.pone.0000718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Armstrong S, Franklin F, Jones G. A meiotic time-course for Arabidopsis thaliana. Sexual Plant Reproduction. 2003;16(3):141–149. doi: 10.1007/s00497-003-0186-4. [DOI] [Google Scholar]
- Couteau F, Belzile F, Horlow C, Grandjean O, Vezon D, Doutriaux MP. Random chromosome segregation without meiotic arrest in both male and female meiocytes of a dmc1 mutant of Arabidopsis. Plant Cell. 1999;11(9):1623–1634. doi: 10.1105/tpc.11.9.1623. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Muyt A, Pereira L, Vezon D, Chelysheva L, Gendrot G, Chambon A, Laine-Choinard S, Pelletier G, Mercier R, Nogue F. et al. A high throughput genetic screen identifies new early meiotic recombination functions in Arabidopsis thaliana. PLoS Genet. 2009;5(9):e1000654. doi: 10.1371/journal.pgen.1000654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mazina OM, Mazin AV, Nakagawa T, Kolodner RD, Kowalczykowski SC. Saccharomyces cerevisiae Mer3 helicase stimulates 3'-5' heteroduplex extension by Rad51; implications for crossover control in meiotic recombination. Cell. 2004;117(1):47–56. doi: 10.1016/S0092-8674(04)00294-6. [DOI] [PubMed] [Google Scholar]
- Nakagawa T, Flores-Rozas H, Kolodner RD. The MER3 helicase involved in meiotic crossing over is stimulated by single-stranded DNA-binding proteins and unwinds DNA in the 3' to 5' direction. J Biol Chem. 2001;276(34):31487–31493. doi: 10.1074/jbc.M104003200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakagawa T, Kolodner RD. The MER3 DNA helicase catalyzes the unwinding of holliday junctions. J Biol Chem. 2002;277(31):28019–28024. doi: 10.1074/jbc.M204165200. [DOI] [PubMed] [Google Scholar]
- Nakagawa T, Kolodner RD. Saccharomyces cerevisiae Mer3 is a DNA helicase involved in meiotic crossing over. Mol Cell Biol. 2002;22(10):3281–3291. doi: 10.1128/MCB.22.10.3281-3291.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakagawa T, Ogawa H. The Saccharomyces cerevisiae MER3 gene, encoding a novel helicase-like protein, is required for crossover control in meiosis. EMBO J. 1999;18(20):5714–5723. doi: 10.1093/emboj/18.20.5714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen C, Zhang W, Timofejeva L, Gerardin Y, Ma H. The Arabidopsis ROCK-N-ROLLERS gene encodes a homolog of the yeast ATP-dependent DNA helicase MER3 and is required for normal meiotic crossover formation. Plant J. 2005;43(3):321–334. doi: 10.1111/j.1365-313X.2005.02461.x. [DOI] [PubMed] [Google Scholar]
- Copenhaver GP, Housworth EA, Stahl FW. Crossover interference in Arabidopsis. Genetics. 2002;160(4):1631–1639. doi: 10.1093/genetics/160.4.1631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alonso JM, Stepanova AN, Leisse TJ, Kim CJ, Chen H, Shinn P, Stevenson DK, Zimmerman J, Barajas P, Cheuk R. et al. Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science. 2003;301(5633):653–657. doi: 10.1126/science.1086391. [DOI] [PubMed] [Google Scholar]
- Yamamoto YY, Ichida H, Abe T, Suzuki Y, Sugano S, Obokata J. Differentiation of core promoter architecture between plants and mammals revealed by LDSS analysis. Nucleic Acids Res. 2007;35(18):6219–6226. doi: 10.1093/nar/gkm685. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cooper SJ, Trinklein ND, Anton ED, Nguyen L, Myers RM. Comprehensive analysis of transcriptional promoter structure and function in 1 % of the human genome. Genome Res. 2006;16(1):1–10. doi: 10.1101/gr.4222606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Siemering KR, Golbik R, Sever R, Haseloff J. Mutations that suppress the thermosensitivity of green fluorescent protein. Curr Biol. 1996;6(12):1653–1663. doi: 10.1016/S0960-9822(02)70789-6. [DOI] [PubMed] [Google Scholar]
- Clough SJ, Bent AF. Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998;16(6):735–743. doi: 10.1046/j.1365-313x.1998.00343.x. [DOI] [PubMed] [Google Scholar]
- Higo K, Ugawa Y, Iwamoto M, Korenaga T. Plant cis-acting regulatory DNA elements (PLACE) database: 1999. Nucleic Acids Res. 1999;27(1):297–300. doi: 10.1093/nar/27.1.297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gubler F, Raventos D, Keys M, Watts R, Mundy J, Jacobsen JV. Target genes and regulatory domains of the GAMYB transcriptional activator in cereal aleurone. Plant J. 1999;17(1):1–9. doi: 10.1046/j.1365-313X.1999.00346.x. [DOI] [PubMed] [Google Scholar]
- Sessa G, Morelli G, Ruberti I. The Athb-1 and −2 HD-Zip domains homodimerize forming complexes of different DNA binding specificities. EMBO J. 1993;12(9):3507–3517. doi: 10.1002/j.1460-2075.1993.tb06025.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Figure S1.GFP signals in meiocytes driven by meiotically-active promoters. (PDF 2536 kb).
Figure S2.The position of GFP spots and nuclear DNA/chromosomes in pMS5: GFP transformants. (PDF 636 kb).
Figure S3.GFP signals in both somatic and meiotic cells driven by a constitutive promoter d35S. (PDF 388 kb).
Table S1.PLACE motifs enriched in promoters of meiotically-active genes. (PDF 12 kb).
Table S2.Statistically overrepresented motifs in the promoters of meiotically-active genes. (PDF 8 kb).
Figure S4.Sequence logos of novel motifs in meiotically-active promoters found by MEME. (PDF 213 kb).
Table S3.A list of homologous recombination-related genes for comparative promoter analysis. (PDF 7 kb).
Figure S5.Sequence logos of novel motifs in promoters of homologous recombination-related genes found by MEME. (PDF 68 kb).
Table S4.Motifs enriched specifically in 15 known meiosis-related promoters. (PDF 7 kb)