

Abstract
Purpose
Prior studies from our laboratory have demonstrated the efficacy of a combined treatment of low doses of dietary agents curcumin and phenylethylisothiocyanate in effectively suppressing prostate cancer in vitro in human prostate cancer PC3 cells as well as in vivo in immunodeficient mice implanted with PC3 cells. Hence, this study was undertaken to examine the potential chemopreventive properties of the two agents against transgenic adenocarcinoma of the mouse prostate.
Materials and Methods
The efficacy of AIN-76A diet supplemented with 2% curcumin or 0.05% PEITC or a combination of 1% curcumin and 0.025% PEITC for periods of 10 and 16 weeks was tested against adenocarcinoma of the mouse prostate. Immunohistochemistry and Western blot analysis were used to examine the expression of proliferation and apoptotic biomarkers. All statistical tests were two-sided.
Results
Supplementing AIN-76A diet with dietary phytochemicals curcumin or PEITC either alone or in combination, significantly decreased incidence of prostate tumor formation (P=0.0064). Immunohistochemistry revealed a significant inhibition of high-grade PIN (P=0.0006, 0.000069, 0.00029 for a treatment period of 10 weeks and P=0.02582, 0.022179, 0.0317 for a treatment period of 16 weeks) along with decreased proliferation and increased apoptotic index in the curcumin, PEITC or curcumin and PEITC treated animals, respectively. Furthermore, Western blot analysis revealed that downregulation of the Akt signaling pathway may in part play a role in decreasing cell proliferation ultimately retarding prostate tumor formation.
Conclusion
Our data lucidly evidence the chemopreventive merits of dietary phytochemicals curcumin and PEITC in suppressing prostate adenocarcinoma.
Keywords: cancer chemoprevention, curcumin, PEITC, prostate cancer, TRAMP
INTRODUCTION
Prostate cancer (CaP) is one of the most frequently diagnosed form of cancer and the second leading cause of cancer related death among men in the USA. CaP when truly confined only to the prostate can be cured by radiation therapy or androgen ablation. Such treatment causes apoptosis in androgen-dependent cells and substantially reduces tumor growth in ~80% of patients. However, in all too many cases, androgen-independent tumor cells, that are refractory to hormone therapy, ultimately develop and eventually result in a relapse. Currently, the number of patients suffering from this invasive disease is on the rise and given the fact that when clinically significant this disorder is associated with a very high mortality rate, prevention may prove to be the best approach to combat it (1). The use of recently developed, genetically manipulated animals provides a good strategy to study CaP chemoprevention. The transgenic adenocarcinoma of the mouse prostate (TRAMP) is one such genetically manipulated model that recapitulates many salient aspects of the progressive forms of human prostatic cancer. In this model, expression of the SV40 early genes (T and t antigens, Tag) are driven by the prostate-specific promoter probasin that leads to cell transformation within the prostate. The TRAMP transgenic mice develop high-grade prostatic intraepithelial neoplasia (PIN) within 12 weeks of birth and ultimately develop metastases by 30 weeks, primarily to the lungs, liver and lymph nodes. PIN is considered the precursor to invasive carcinoma since it is most often than not associated with malignant form of the disease and the primary architectural and cytological features of PIN resemble that of invasive carcinoma. Hence the ability to retard PIN formation is considered as an important merit in agents that are tested against prostate carcinoma (2-4).
Epidemiological studies have continually supported the contention that naturally occurring dietary agents demonstrate chemopreventive properties and can suppress several malignancies including that of the prostate. Antineoplastic effect of cruciferous vegetables is mainly attributed to isothiocyanates which occur as thioglucoside conjugates (glucosinolates) in a variety of edible plants including broccoli, watercress, cabbage etc. Organic isothiocyanates are generated due to hydrolysis of corresponding glucosinolates through catalytic activity of enzyme myrosinase which is released on damage of plant cells during processing (cutting or chewing) of cruciferous vegetables. Phenylethylisoathiocyanate (PEITC) is one such naturally occurring isothiocyanate compound that has attracted a great deal of attention due to its remarkable cancer chemopreventive properties. Data from literature clearly demonstrates that PEITC can effectively suppress proliferation of human prostate cancer cells by inducing apoptosis and cell cycle arrest (5-9). Curcumin (diferuloyl-methane), the yellow pigment found in spice turmeric extracted from the rhizome of the plant Curcuma longa has been shown to possess strong anti-oxidant and anti-inflammatory effects (10). Due to these properties it has been very widely investigated for its potential chemopreventive activity. Curcumin has been shown to inhibit the induction of cancers of the skin, forestomach, duodenum and colon in models of chemical carcinogenesis in mice and rats (11,12). It has also shown therapeutic efficacy against human prostate cancer xenografts in nude mice (13,14).
Recent studies from our laboratory have established that combined treatment of low doses of PEITC and curcumin can suppress human prostate cancer cell growth in vitro as well as in immunodeficient (Nu/Nu) mice bearing xenografts of androgen-independent human prostate cancer cells (PC-3) (15,16). The following research study was undertaken to extend the promising chemopreventive effects of a combined treatment of PEITC and curcumin to the TRAMP mouse model. Our results clearly demonstrate that a combination of PEITC and curcumin can effectively inhibit the development and progression of CaP in TRAMP mice.
MATERIALS AND METHODS
Animals
Female hemizygous C57BL/TGN TRAMP mice, line PB Tag 8247NG, and male C57BL/6 mice were purchased from The Jackson Laboratory (Bar Harbor, ME). The animals were bred on same genetic background and maintained in the Animal Care Facility of Rutgers University. Housing and care of the animals was in accordance with the guidelines established by the University’s Animal Research Committee consistent with the NIH Guidelines for the Care and Use of Laboratory Animals. Transgenic males for these studies were routinely obtained as [TRAMP × C57BL/6] F1 or as [TRAMP × C57BL/6] F2 offspring. Identity of transgenic mice was established by the PCR-based DNA screening. Throughout the experiment the animals were housed in cages with wood chip bedding in a temperature-controlled room (68–72°F) with a 12 h light dark cycle, at a relative humidity of 45% to 55%. The animals were fed with irradiated AIN-76A diet (Research Diets Inc, New Brunswick, NJ).
Diets and Study Design
Curcumin (66–70% purity) and phenylethylisothiocyanate—PEITC (>99% purity) were obtained from Sigma chemicals Inc (St. Louis, MO). AIN-76A diets containing either 2% curcumin or 0.05% PEITC or 1% curcumin and 0.025% PEITC were prepared by Research Diets Inc, New Brunswick, NJ. Based on our previous experiments a similar dose of curcumin, PEITC or a combination of the two agents in xenograft model as well as Apc min mice model, no toxicity was observed. These doses were well-tolerated. The prepared diets were stored at −20°C. Fresh diets were weighed each day and added to the cages. Two studies were initiated using 8-week old TRAMP males. One was a short term study involving treatment for 10 weeks (i.e. till the mice were 18 weeks old) while the second was a long term study involving treatment for 16 weeks (i.e. till the mice were 24 weeks old). In addition, four TRAMP males of age 8 weeks and 12 weeks were used as untreated controls. A summary of study design is as shown in Fig. 1A.
Fig. 1.
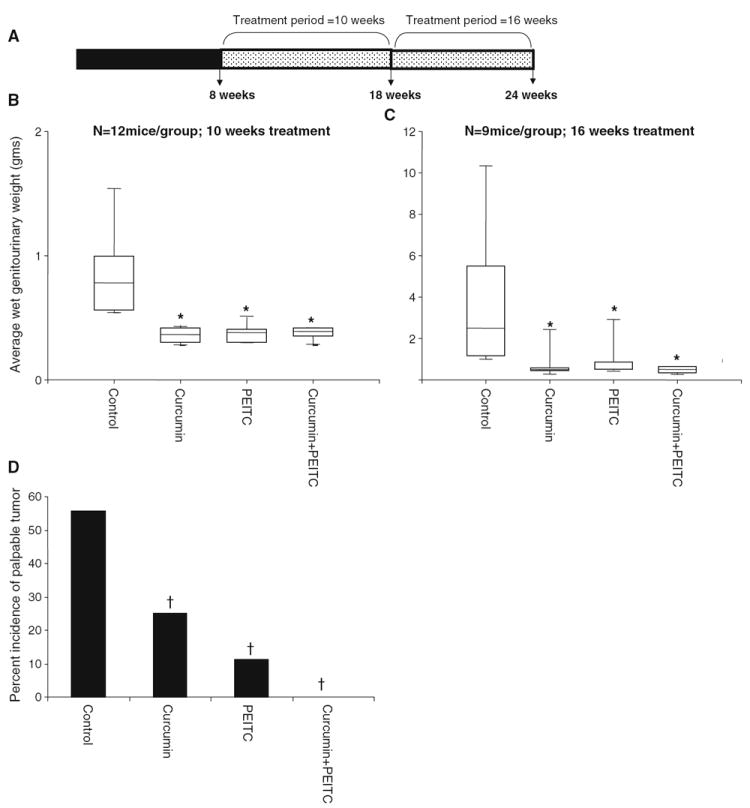
The effects of curcumin and/or PEITC supplemented diet on the genitourinary weight. A Time line. Eight week old TRAMP males were put on an AIN-76A diet supplemented with either (2%) curcumin or (0.05%) PEITC alone or a combination of the two chemopreventive agents (1% and 0.025%). The diets were maintained for two treatment periods of 10 and 16 weeks till the age of the mice were 18 and 24 weeks, respectively at which point they were sacrificed and the genitourinary apparatus consisting of the prostate, seminal vesicles and bladder, the lungs, livers and periaortic renal lymph nodes were harvested. B, C Effects of curcumin and/or PEITC on the genitourinary weights of animals treated for periods of 10 and 16 weeks. The upper boundary of the box represents the 75th percentile while the lower boundary of the box represents the 25th percentile of the data distribution, the horizontal line within each box represents the median value and the error bars represent the 95% confidence intervals. Asterisk Significantly different from the control. D Incidence of palpable tumor in the 24 week old control versus treated animals. Dagger Significantly different based on Chi’s square test.
All the animals were put on AIN-76A diet 1 week prior to the study. The control animals received AIN-76A diet throughout the experiment while the treatment groups received the corresponding chemopreventive agent supplemented AIN-76A diet. The animals were weighed weekly and monitored on a regular basis for their general health. At each time point the mice were killed by cervical dislocation and the genitourinary apparatus (GUT) consisting of the seminal vesicles, prostate and the bladder were isolated for further analyses.
Histopathology
The dorso-lateral prostate was excised and fixed overnight in 10% formalin and then transferred to 70% ethanol. Sections (4 μM) were cut from paraffin embedded tissue and mounted on slides. The sections were stained with Haematoxylin and Eosin to observe any neoplastic changes. Sections were blindly evaluated by histopathologist Dr. Xingpei Hao to classify PIN lesions. Lesions were classified as PIN I, PIN II, PIN III and PIN IV as described by Park and co-workers (17). Briefly prostatic tissue displaying columnar epithelial cells with surrounding stroma consisting of two to three layers of smooth muscle with loose connective tissue was classified as normal. Tissue displaying fairly small foci with one or two layers of atypical cells with or without papillary structures and none of them being secondary branched was classified as PIN I. In PIN I the cells appear slightly taller than normal cells, though the fibromuscular stromal sheath appears intact. PIN II displays larger foci with multiple layers of atypical cells that do not, however, fill up the lumen space. The underlying fibromuscular stromal sheath appears intact, though the epithelial cells display papillary, cribiform or tufting patterns. The atypical cells appear the same as in PIN I though there is increased nuclear pleomorphism and hyperchromasia. PIN III displays foci with multiple layers of atypical cells that fill up roughly two thirds of the lumen. The epithelial cells exhibit either papillary with multiple branches or cribiform or tufting patterns. The atypical cells have inverted nuclear to cytoplasmic ratio. They appear poorly oriented with increasingly severe nuclear pleomorphism and hyperchromasia. Mitotic figures are present. PIN IV displays foci with multiple layers of atypical cells that fill up the lumen of the ducts. The ducts appear distorted with irregular or absent fibromuscular sheath. The epithelial cells form solid, cribiform or tufting growth patterns. The nuclear to cytoplasmic ratio is inverted. The atypical cells display increasingly severe nuclear pleopmorphism and hyperchromasia. Mitotic figures are present.
For the purpose of ease, PIN I and PIN II have been grouped as low grade PIN while PIN III and PIN IV have been grouped as high grade PIN.
Apoptosis and Proliferation
The apoptotic cells were detected using an ApopTag In Situ Apoptosis Detection Kit (Chemicon). The assay was performed according to the manufacturer’s manual. After deparaffinization, the tissues sections were incubated in proteinase K for 15 min in room temperature. The sections were then incubated with terminal deoxynucleotidyl transferase (TdT) enzyme at 37°C for 1 h, washed in three changes of PBS and incubated with anti-digoxigenin conjugate in a humidified chamber at room temperature for 30 min. The color was developed by incubating the sections with peroxidase substrate and then counterstained with hematoxylin for 30 s. For detection of proliferative cells, PCNA antibody (1:50; Dakocytomaton, CA) was used. The assay was performed following the manufacturer’s protocols. A positive control slide of rat mammary glands provided by the manufacturer was used as positive control for the in situ apoptosis detection assay. For the PCNA staining, mouse intestinal crypt cells were used as a positive control. The apoptotic or proliferative index was calculated as number of positive cells/total cells counted×200.
Immunoblot Analysis
The dorso-lateral prostate tissues removed from both treated and control groups were pooled and homogenized with RIPA buffer (50 mM NaCl, 0.5% Triton-X 100, 50 mM Tris–HCl, pH 7.4, 25 mM NaF, 20 mM EGTA, 1 mM DTT, 1 mM Na3VO4, protease inhibitor cocktail tablet (Roche, Manheim, Germany) at a concentration of 10 μg/ml and placed on ice for 40 min, followed by centrifugation at 14,800×g for 15 min. The protein concentrations were measured by Bicinchonic Acid (BCA) solution (Pierce, Rockford, IL). Protein (20 μg) was loaded onto Biorad pre-cast gels (4–12%) and after electrophoresis, transferred onto polyvinylidene difluoride membrane. The membrane was blocked with 5% BSA in 0.1% Tween-20 in PBS (TBST) for 1 h followed by incubation with primary antibody in 3% BSA overnight at 4°C. After three 15-min washes with TBST, the membrane was incubated with horseradish peroxidase conjugated secondary antibody in 3% BSA for 1 h at room temperature followed by washing with TBST. The proteins were then visualized using Super-signal West Femto (Pierce, Rockford, IL).
Statistics
The results for tumor incidence and poorly differentiated carcinoma between 18 and 24 week old control males were analyzed by Student’s test. Statistical significance of differences in all other measurements was determined by one-way analysis of variance followed by Tukey’s procedure for all pairwise multiple comparisons, excluding the control group. All statistical tests were two-sided and P<0.05 was considered statistically significant.
RESULTS
Effects of Curcumin and/or PEITC Supplemented Diet and General Health Observations
The overall health of all the mice was observed on a daily basis and was found to be good throughout the study period. No significant change in the body weights of the animals was found throughout the experimental period.
Effect of 10 weeks of Curcumin and/or PEITC Supplemented Diet on Prostate Tumorigenesis
The mice were put on respective diets for a period of 10 weeks and sacrificed when they were 18 weeks old. The effect of such treatment for a period of 10 weeks on the wet weight of the GUT apparatus is shown in Fig. 1B. Curcumin and/or PEITC supplemented diet significantly decreased the GUT weight with P values of 0.000322, 0.000295, 0.000357 respectively. Such treatment resulted in decreased hyperplasia in the GUT apparatus, especially in the seminal vesicles as evidenced by their reduced size. Hematoxylin and Eosin staining of the dorso-lateral prostates of these animals revealed that the curcumin or PEITC or a combination of curcumin and PEITC treated animals demonstrated the presence of significantly higher proportion of normal prostatic tissue as compared to the control animals that demonstrated almost no normal prostatic tissue (P=0.0069, 0.0019, 0.0019, respectively). In addition the treated animals demonstrated low levels of high grade PIN as compared to the control animals (P=0.007, 0.00533 and 0.00883, respectively). Although PEITC treated animals demonstrated higher levels of normal tissue, the degree of inhibition of high-grade PIN was the same as the curcumin treated animals. A combination diet, surprisingly, did not inhibit high grade PIN as much as individual agents curcumin or PEITC did, although this difference was not found to be statistically significant. Results are as depicted in Figure 2B. Thus, it may be inferred that a combination of low doses of potential chemopreventive agents curcumin and PEITC for a treatment period of 10 weeks, lacked any synergistic effect.
Fig. 2.
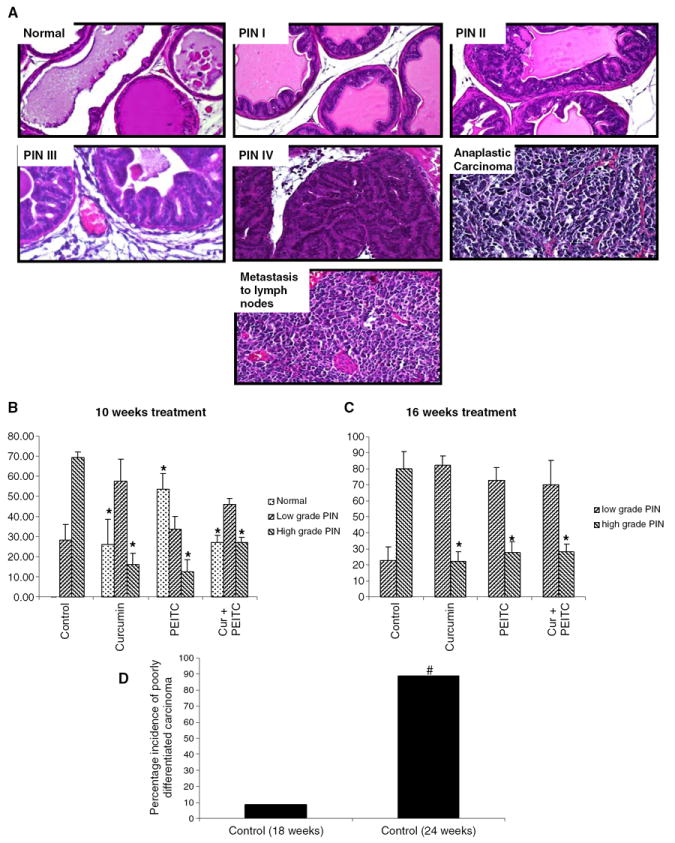
Histological evaluation of prostatic intra-epithelial lesions and adenocarcinoma and their inhibition by curcumin and/or PEITC supplemented diets. A Representative sections depicting normal tissue, PIN I, PIN II, PIN III, PIN IV, anaplastic carcinoma and metastasis to lymph nodes. B Levels of low and high grade PIN in animals treated for a period of 10 weeks. Asterisk Significantly different from the control. C Levels of low and high grade PIN in animals treated for a period of 16 weeks. Asterisk Significantly different from the control. D Bar graph representing number of 18 and 24 week old control animals that demonstrated poorly differentiated carcinoma, histologically. Pound sign Significantly different based on Student’s t test. Data represented is the mean value and the error bars represent ±SE.
Effects of 16 weeks of Curcumin and/or PEITC Supplemented Diet on Prostate Tumorigenesis
The mice were put on the respective diets for a period of 16 weeks and sacrificed when they were 24 weeks old. The effect of such treatment on the wet weight of the GUT is summarized in Fig. 1C. There was a statistically significant reduction in GUT weight followed by curcumin and/or PEITC supplemented diet (P=0.025, 0.04 and 0.01345, respectively). A significant reduction in the incidence of palpable tumor was observed in the treated animals (P= 0.0064). Six out of nine control animals fed AIN-76A diet developed palpable tumors. In contrast, only two curcumin fed and one PEITC fed animal demonstrated the presence of palpable tumors. None in the curcumin and PEITC fed groups of TRAMP males developed palpable tumors. Results are as depicted in Fig. 1D. These findings were further confirmed by histological observations that showed that eight out of the nine control animals developed poorly differentiated carcinoma with one of them demonstrating necrosis. In contrast only one 18 week old control animal developed poorly differentiated carcinoma (P=0.00022). This confirms previous findings that as age of the TRAMP mouse advances, the incidence of carcinoma increases. Results are as depicted in Fig. 2D. Among the treated groups, one out of nine combination diet fed animal demonstrated poorly differentiated carcinoma although there was no evidence of any palpable tumor. Of most importance is the observation that the invasiveness of CaP was suppressed in the treated groups as compared to the control. This is evidenced by effective inhibition of high grade PIN in the curcumin, PEITC and combination diet fed animals (P=0.0023, 0.014, 0.0036 respectively). Results are as depicted in Figure 2C. Thus it can be inferred that a combination of low doses of chemopreventive agents curcumin and PEITC can effectively suppress prostate tumor growth and this effect maybe moderately synergistic when treated for longer periods.
Effect of Curcumin and/or PEITC Supplemented Diet on Proliferation Marker (PCNA) and Apoptosis
PCNA serves as an auxiliary protein for DNA polymerase δ-driven DNA synthesis and is known to be cell-cycle regulated (18). Treatment with either curcumin or PEITC or the two in combination for periods of 10 weeks resulted in significantly low levels of PCNA (P=0.001818, 0.0005657, 0.01288, respectively) as observed by immunohistochemical analysis. A similar statistically significant decrease in PCNA levels was observed in TRAMP males fed curcumin and/or PEITC diet for a period of 16 weeks (P=0.00240, 0.00075, 0.002509, respectively). Results are as shown in Fig. 3A and B.
Fig. 3.
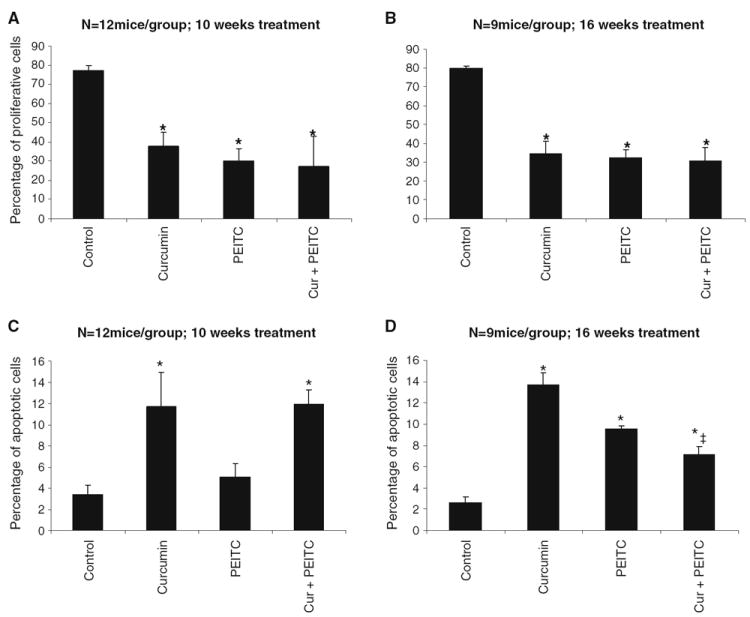
Immunohistochemical analysis of the effect of curcumin and/or PEITC supplemented diet on the proliferation marker PCNA and apoptosis. A, B PCNA expression followed by treatment with curcumin and/or PEITC for a period of 10 and 16 weeks, respectively. Asterisk Significantly different from the control. Data represented is the mean value and the error bars represent ±SE. C, D Apoptotic cells counted under ×40 were quantified and presented as a percentage of apoptotic cells followed by curcumin and/or PEITC supplemented diet for periods of 10 and 16 weeks, respectively. Asterisk Significantly different from the control. Double dagger Significantly different from curcumin treatment, Tukey’s test. Data represented is the mean value and the error bars represent ±SE.
The percentage of apoptotic cells in the dorso-lateral prostates of the animals fed curcumin or curcumin and PEITC supplemented diet for 10 weeks was significantly higher than control animals (P=0.0072 and 0.00523 respectively.) Although the number of apoptotic cells in the PEITC fed animals was higher than the control, this difference was not found to be statistically significant. In contrast, all the long term treatment groups—curcumin and/or PEITC demonstrated significantly higher percentage of apoptotic cells as compared to the control (P=0.001656, 0.000159, 0.0163 respectively). Results are as shown in Fig. 3C and D.
Effect of Curcumin and/or PEITC Supplemented Diet on the Expression Levels of p-PDK1, p-Akt, p-FKHR, Bad and Caspase 3 Proteins
In order to examine the molecular targets involved in curcumin and/or PEITC mediated CaP regression, we investigated the expression levels of both proliferative and pro-apoptotic biomarkers of tumor progression—PDK1, Akt, FKHR, Bad and caspase 3 proteins. Results are as depicted in Fig. 4.
Fig. 4.
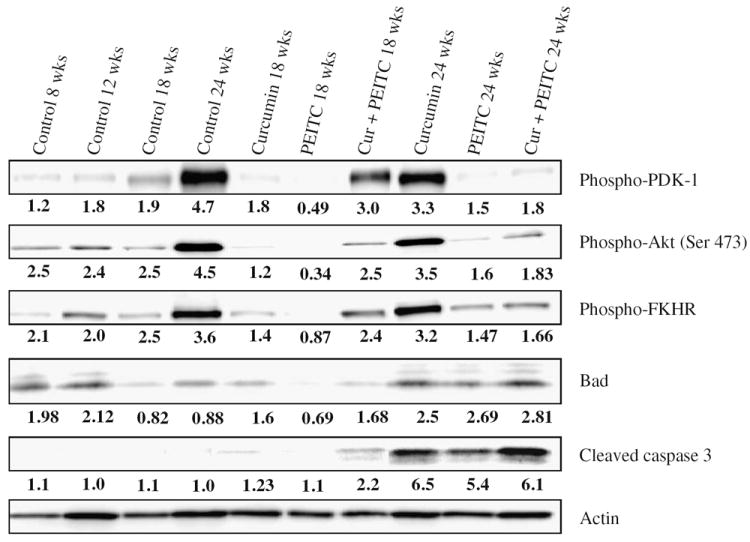
Western blot analysis of proliferative and apoptotic biomarkers—PDK-1, Akt (Ser 473), FKHR, Bad and cleaved caspase 3 in the dorsolateral prostates of mice treated with curcumin and/or PEITC. A 20 μg protein sample was loaded onto each lane. The numbers below represent the relative expression of each biomarker normalized to actin.
Western blot analysis demonstrate reduced expression levels of phospho-PDK1, phospho-Akt (Ser 473) and phospho-FKHR proteins in the prostatic tissues of the treated animals as compared to the control. From the figure, it is clear that as age of the TRAMP mouse advances the levels of the proliferative biomarkers p-PDK-1, p-Akt and p-FKHR progressively increase while that of the pro-apoptotic biomarkers either progressively decrease as in the case of Bad, or are undetectable in the case of cleaved caspase-3. Interestingly, curcumin and PEITC treatment alone for a period of 10 weeks suppressed the levels of PDK-1, Akt and FKHR however, a combination diet did not suppress these levels as much on the other hand did increase levels of pro-apoptotic biomarkers Bad and cleaved caspase 3. The most interesting observation is that curcumin when fed for a period of 16 weeks, did not effectively suppress the proliferative biomarkers though it did significantly increase the levels of both Bad and cleaved caspase 3 proteins. Thus it may be inferred that a diet supplemented with curcumin and/or PEITC can impede the progression of PIN at least in part by downregulating the Akt pro-survival pathway.
DISCUSSION
Earlier studies from our laboratory have demonstrated that intraperitoneal injections of curcumin and/or PEITC retarded the growth of PC3 xenografts in immunodeficient mice. This study clearly demonstrated the efficacy of the two agents as chemopreventive rather than cancer therapeutic. Hence, as an extension to the above study, we evaluated the chemopreventive properties of curcumin and PEITC in a genetically engineered mouse model which has revolutionized the process of investigating molecular mechanisms of prostate carcinogenesis, the Transgenic Adenocarcinoma of Mouse Prostate (TRAMP) model. The major strength of this model is that cancer arises from normal prostate epithelial cells in their natural tissue microenvironment and progresses through multiple stages thus exhibiting both histological and molecular features similar to human prostate cancer.
It is known that by approximately 6 weeks of age, TRAMP mice exhibit low-grade PIN which progresses to high grade PIN by 12–16 weeks. Focal adenocarcinoma develops between 12–18 weeks and progresses to poorly differentiated carcinoma within 24 weeks. By 28 weeks of age, 100% of these transgenic mice harbor metastatic prostate cancer in liver, lymph nodes and lungs. In the current study we demonstrate that by 24 weeks of age, six out of nine animals had developed palpable tumors and demonstrated metastasis mainly to the lymph nodes. Curcumin or PEITC either alone or in combination were found to be highly effective in decreasing the incidence of tumor formation in these mice.
Histologic analysis of the prostate of control animals clearly displayed major epithelial proliferation, hyperchromatic nuclei and mitotic figures. In contrast the prostate sections of mice fed curcumin and/or PEITC diets displayed no indication of neoplasia. Analysis of other tissues such as lungs, liver and lymph nodes showed normal morphology thus indicating that curcumin and/or PEITC did not have any deleterious effects on these organs (data not shown).
For cancer progression it is vital that cell survival signals are amplified and pro-apoptotic signals are minimalized. The Akt is one such pathway that is often involved in amplifying cell-survival signals. Besides, there is also a rich body of evidence suggesting that prostate cancer progression may be mediated by the Akt signaling pathway (19-22).
The PTEN gene encodes a dual specificity phosphatase active against protein and lipid substrates and is known to be a direct antagonist to the PI3K signaling pathway. The most important downstream target of the PI3K is serine-threonine Akt kinase family. Activation of PI3K caused by suppression of PTEN leads to an increase in its lipid products that facilitate the recruitment of Akt and PDK-1 to the plasma membrane. Once recruited, to the plasma membrane, Akt is phosphorylated by PDK-1 (22). From the data we provide, it is clear that the progression from prostatic lesions to highly invasive carcinoma in the control animals relates well with the gradual increase in levels of activated Akt, thus reinforcing the fact that cell-survival signals are amplified in cancer progression. Treatment with curcumin or PEITC lowers the levels of activated Akt and its upstream kinase PDK-1. Slightly aberrant from the trend is the observation that curcumin treatment for a period of 16 weeks did not significantly alter levels of Akt and PDK-1. This observation is truly intriguing, although it is also suggestive that in vivo long term treatment with PEITC may prove better than curcumin, in suppressing the Akt signaling pathway. Curcumin and PEITC put together demonstrated significant inhibition of the Akt signaling pathway again when treated for longer periods of time. A short term treatment with the combination did not alter the Akt pathway which is consistent with the observation that such treatment did not affect levels of high grade PIN as much as individual agents did. Since immunohistochemical analysis demonstrates that such treatment did decrease the levels of proliferation marker-PCNA and induce apoptosis in the cells, it is alluring to make a conjecture that other cell-survival amplifying pathways may play a role.
Akt is known to possess two phosphorylation sites at Thr 308 and Ser 473/474. PDK-1 is known to phosphorylate Akt at the Thr 308 position. Although our results demonstrate modulation of Akt activity by curcumin and/or PEITC at Ser473/474 position, we did not observe a similar modulation at the Thr 308 position. Prior studies in our lab have demonstrated that in human PC3 cells a combination of curcumin and PEITC can modulate PDK-1 and Akt Ser 473/474 as well as Thr 308 activity (16). However lack of phosphorylation of Akt at the Thr 308 position in vivo suggests that studies aimed to correlate observations made in vitro with those made in vivo are imminent. The ability of curcumin and PEITC to modulate Akt activity at Ser 473/474 site directly without the involvement of its upstream kinase PDK-1 might also in part explain such observation. Activated Akt is known to promote cell survival by inactivating its downstream targets including Bad and members of the forkhead family. From figures it is clear that activated Akt in the untreated controls led to suppression of Bad and activation of phosphorylated forkhead proteins while curcumin and/or PEITC treatment could effectively activate pro-apoptotic protein Bad and suppress phosphorylated forkhead proteins. The finding that curcumin when treated for a period of 16 weeks does induce pro-apoptotic Bad without suppressing the Akt signaling pathway is indicative of the intervention of certain other signal transduction pathways. Another possible explanation for this finding could be that as the levels of Akt increase with invasiveness of the carcinoma, curcumin alone may not be able to suppress Akt and thus proliferation. It would do so in a much better fashion when combined with other chemopreventive agents mainly PEITC. This also explains the findings that a combination of curcumin and PEITC was far more effective in suppressing tumor incidence as well as activating cleaved caspase 3—a hallmark feature of apoptosis. Hence, though we did not observe any synergistic effects of combined therapy with curcumin and PEITC when administered for a shorter period of time (10 weeks), a long term treatment definitely could prove beneficial.
In summary, we show that dietary agents—curcumin and PEITC can suppress levels of high grade PIN in the TRAMP mouse model by downregulating the Akt signaling pathway and upregulating members of the pro-apoptotic family ultimately resulting in inhibited prostate tumor formation. To the best of our knowledge, this study is the first to demonstrate the efficacy of these agents both alone and in combination in the TRAMP mouse model.
Acknowledgments
This study is supported in part by the National Institutes of Health grants R01-CA094828 and R01-CA118947 to A.-N. T. Kong.
References
- 1.Denis L, Morton MS, Griffiths K. Diet and its preventive role in prostatic disease. Eur Urol. 1999;35:377–387. doi: 10.1159/000019912. [DOI] [PubMed] [Google Scholar]
- 2.Abate-Shen C, Shen MM. Mouse models of prostate carcinogenesis. Trends Genet. 2002;18:S1–S5. doi: 10.1016/s0168-9525(02)02683-5. [DOI] [PubMed] [Google Scholar]
- 3.Gingrich JR, Barrios RJ, Morton RA, Boyce BF, DeMayo FJ, Finegold MJ, et al. Metastatic prostate cancer in a transgenic mouse. Cancer Res. 1996;56:4096–102. [PubMed] [Google Scholar]
- 4.Greenberg NM, DeMayo FJ, Sheppard PC, Barrios R, Lebovitz R, Finegold M, et al. The rat probasin gene promoter directs hormonally and developmentally regulated expression of a heterologous gene specifically to the prostate in transgenic mice. Mol Endocrinol. 1994;8:230–239. doi: 10.1210/mend.8.2.8170479. [DOI] [PubMed] [Google Scholar]
- 5.Xiao D, Singh SV. Phenethyl isothiocyanate-induced apoptosis in p53-deficient PC-3 human prostate cancer cell line is mediated by extracellular signal-regulated kinases. Cancer Res. 2002;62:3615–619. [PubMed] [Google Scholar]
- 6.Huang C, Ma WY, Li J, Hecht SS, Dong Z. Essential role of p53 in phenethyl isothiocyanate-induced apoptosis. Cancer Res. 1998;58:4102–4106. [PubMed] [Google Scholar]
- 7.Chen YR, Han J, Kori R, Kong AN, Tan TH. Phenylethyl isothiocyanate induces apoptotic signaling via suppressing phosphatase activity against c-Jun N-terminal kinase. J Biol Chem. 2002;277:39334–39342. doi: 10.1074/jbc.M202070200. [DOI] [PubMed] [Google Scholar]
- 8.Xiao D, Zeng Y, Choi S, Lew KL, Nelson JB, Singh SV. Caspase-dependent apoptosis induction by phenethyl isothiocyanate, a cruciferous vegetable-derived cancer chemopreventive agent, is mediated by Bak and Bax. Clin Cancer Res. 2005;11:2670–2679. doi: 10.1158/1078-0432.CCR-04-1545. [DOI] [PubMed] [Google Scholar]
- 9.Xu C, Shen G, Yuan X, Kim JH, Gopalakrishnan A, Keum YS, et al. ERK and JNK signaling pathways are involved in the regulation of activator protein 1 and cell death elicited by three isothiocyanates in human prostate cancer PC-3 cells. Carcinogenesis. 2006;27:437–445. doi: 10.1093/carcin/bgi251. [DOI] [PubMed] [Google Scholar]
- 10.Sharma OP. Antioxidant activity of curcumin and related compounds. Biochem Pharmacol. 1976;25:1811–1812. doi: 10.1016/0006-2952(76)90421-4. [DOI] [PubMed] [Google Scholar]
- 11.Huang MT, Wang ZY, Georgiadis CA, Laskin JD, Conney AH. Inhibitory effects of curcumin on tumor initiation by benzo [a]pyrene and 7,12-dimethylbenz[a]anthracene. Carcinogenesis. 1992;13:2183–186. doi: 10.1093/carcin/13.11.2183. [DOI] [PubMed] [Google Scholar]
- 12.Huang MT, Lou YR, Ma W, Newmark HL, Reuhl KR, et al. Inhibitory effects of dietary curcumin on forestomach, duodenal, and colon carcinogenesis in mice. Cancer Res. 1994;54:5841–5847. [PubMed] [Google Scholar]
- 13.Deeb D, Jiang H, Gao X, Hafner MS, Wong H, Divine G, et al. Curcumin sensitizes prostate cancer cells to tumor necrosis factor-related apoptosis-inducing ligand/Apo2L by inhibiting nuclear factor-kappaB through suppression of IkappaBalpha phosphorylation. Mol Cancer Ther. 2004;3:803–812. [PubMed] [Google Scholar]
- 14.Hong JH, Ahn KS, Bae ES, Jeon SS, Choi HY. The effects of curcumin on the invasiveness of prostate cancer in vitro and in vivo. Prostate Cancer Prostatic Dis. 2006;9:147–152. doi: 10.1038/sj.pcan.4500856. [DOI] [PubMed] [Google Scholar]
- 15.Khor TO, Keum YS, Lin W, Kim JH, Hu R, Shen G, et al. Combined inhibitory effects of curcumin and phenethyl isothiocyanate on the growth of human PC-3 prostate xenografts in immunodeficient mice. Cancer Res. 2006;66:613–621. doi: 10.1158/0008-5472.CAN-05-2708. [DOI] [PubMed] [Google Scholar]
- 16.Kim JH, Xu C, Keum YS, Reddy BS, Conney A, Kong AN. Inhibition of EGFR signaling in human prostate cancer PC-3 cells by combination treatment with beta-phenylethyl isothiocyanate and curcumin. Carcinogenesis. 2006;27:475–482. doi: 10.1093/carcin/bgi272. [DOI] [PubMed] [Google Scholar]
- 17.Park JH, Walls JE, Galvez JJ, Kim M, Abate-Shen C, Shen MM, et al. Prostatic intraepithelial neoplasia in genetically engineered mice. Am J Pathol. 2002;161:727–735. doi: 10.1016/S0002-9440(10)64228-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Gupta S, Hastak K, Ahmad N, Lewin JS, Mukhtar H. Inhibition of prostate carcinogenesis in TRAMP mice by oral infusion of green tea polyphenols. Proc Natl Acad Sci U S A. 2001;98:10350–10355. doi: 10.1073/pnas.171326098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Aziz MH, Nihal M, Fu VX, Jarrard DF, Ahmad N. Resveratrol-caused apoptosis of human prostate carcinoma LNCaP cells is mediated via modulation of phosphatidylinositol 3’-kinase/Akt pathway and Bcl-2 family proteins. Mol Cancer Ther. 2006;5:1335–1341. doi: 10.1158/1535-7163.MCT-05-0526. [DOI] [PubMed] [Google Scholar]
- 20.Pommery N, Henichart JP. Involvement of PI3K/Akt pathway in prostate cancer—potential strategies for developing targeted therapies. Mini Rev Med Chem. 2005;5:1125–1132. doi: 10.2174/138955705774933356. [DOI] [PubMed] [Google Scholar]
- 21.Pommery N, Henichart JP. Involvement of PI3K/Akt pathway in prostate cancer. Potential strategies for developing targeted therapies. Ann Pharm Fr. 2005;63:69–75. doi: 10.1016/s0003-4509(05)82253-9. [DOI] [PubMed] [Google Scholar]
- 22.Shukla S, Maclennan GT, Marengo SR, Resnick MI, Gupta S. Constitutive activation of P I3 K-Akt and NF-kappaB during prostate cancer progression in autochthonous transgenic mouse model. Prostate. 2005;64:224–239. doi: 10.1002/pros.20217. [DOI] [PubMed] [Google Scholar]