

Abstract
In multicellular organisms, cell behavior is dictated by interactions with the extracellular matrix. Consequences of matrix-engagement range from regulation of cell migration and proliferation, to secretion and even differentiation. The signals underlying each of these complex processes arise from the molecular interactions of extracellular matrix receptors on the surface of the cell. Integrins are the prototypic receptors and provide a mechanical link between extracellular matrix and the cytoskeleton, as well as initiating some of the adhesion-dependent signaling cascades. However, it is becoming increasingly apparent that additional transmembrane receptors function alongside the integrins to regulate both the integrin itself and signals downstream. The most elegant of these examples is the transmembrane proteoglycan, syndecan-4, which cooperates with α5β1-integrin during adhesion to fibronectin. In vivo models demonstrate the importance of syndecan-4 signaling, as syndecan-4-knockout mice exhibit healing retardation due to inefficient fibroblast migration1,2. In wild-type animals, migration of fibroblasts toward a wound is triggered by the appearance of fibronectin that leaks from damaged capillaries and is deposited by macrophages in injured tissue. Therefore there is great interest in discovering strategies that enhance fibronectin-dependent signaling and could accelerate repair processes.
The integrin-mediated and syndecan-4-mediated components of fibronectin-dependent signaling can be separated by stimulating cells with recombinant fibronectin fragments. Although integrin engagement is essential for cell adhesion, certain fibronectin-dependent signals are regulated by syndecan-4. Syndecan-4 activates the Rac1 protrusive signal3, causes integrin redistribution1, triggers recruitment of cytoskeletal molecules, such as vinculin, to focal adhesions4, and thereby induces directional migration3. We have looked for alternative strategies for activating such signals and found that low-intensity pulsed ultrasound (LIPUS) can mimic the effects of syndecan-4 engagement5. In this protocol we describe the method by which 30 mW/cm2, 1.5 MHz ultrasound, pulsed at 1 kHz (Fig. 1) can be applied to fibroblasts in culture (Fig. 2) to induce Rac1 activation and focal adhesion formation. Ultrasound stimulation is applied for a maximum of 20 minutes, as this combination of parameters has been found to be most efficacious for acceleration of clinical fracture repair6. The method uses recombinant fibronectin fragments to engage α5β1-integrin, without engagement of syndecan-4, and requires inhibition of protein synthesis by cycloheximide to block deposition of additional matrix by the fibroblasts., The positive effect of ultrasound on repair mechanisms is well documented7,8, and by understanding the molecular effect of ultrasound in culture we should be able to refine the therapeutic technique to improve clinical outcomes.
Keywords: Biomedical Engineering, Issue 63, Ultrasound, LIPUS, Focal Adhesion, Syndecan-4, Wound Healing, Extracellular Matrix, Rac1, bioengineering
Protocol
1. Coating Surfaces with Matrix Ligand
Ultrasound will be applied to cells in 3.5-cm wells, either as individual dishes or as a 6-well plate. For biochemical assays, coat ligand directly onto the plastic. For immunofluorescence, boil glass coverslips three times in MilliQ water and place 4 coverslips into the bottom of each well.
Glass surfaces must be derivatized to ensure efficient ligand coating. Dissolve sulpho-m-maleimidobenzoyl-N-hydrosuccinimide ester in PBS at 1 mM. Add 2 ml to each well and incubate for 30 minutes at room temperature.
Wash the derivatized surfaces 3 times with PBS.
Coat either derivatized glass or untreated plastic with integrin ligand. Apply 2 ml of 10 ug/ml integrin ligand9, dissolved in PBS containing Ca2+ and Mg2+ to each well and incubate overnight at 4 °C.
Prepare monoparticulate BSA by heating 1% BSA, dissolved in PBS, to 85 °C for 13 minutes. Allow to cool and then pass through a 0.45-μm filter.
Wash ligand-coated surfaces 3 times with PBS and block with the monoparticulate BSA for 30 minutes.
2. Preparation of Cells
To control the matrix environment throughout the experiment, protein synthesis should be blocked by addition of 25 μg/ml cycloheximide to 80% confluent fibroblasts for 2 hours.
Rinse cycloheximide-treated cells with PBS and detach from the culture flask using 0.5 mg/ml trypsin/EDTA.
Harvest detached cells by centrifugation at 500x g for 5 minutes.
Resuspend cells in DMEM/25 mM HEPES, 25 μg/ml cycloheximide.
Count cell density of the suspension using a haemocytometer and dilute to 105 ml-1.
Incubate cells at 37 °C for 20 minutes to recover surface integrin.
Rinse the ligand-coated, BSA-blocked wells 3 times with PBS.
Seed 2 ml of the cell suspension into each well.
Allow cells to spread for 2 hours at 37 °C, 5% CO2.
3. Ultrasound Stimulation
Warm the ultrasound emitter array and coupling gel to 37 °C.
Apply a pea-sized blob of coupling gel to each emitter.
Test that each emitter is functional using a ultrasound indicator diode detector.
Place the wells on the emitter array. Return array to 37 °C, 5% CO2 and allow temperature to equilibrate for 10 minutes.
Switch on the ultrasound signal.
The ultrasound signal will deactivate after 20 minutes, mimicking the therapeutic regime. If shorter stimulation times are required, plates should be removed from the array at the appropriate time-points. In all cases use unstimulated plates as the negative control.
4. Focal Adhesion Analysis by Immunofluorescence
For analysis of focal adhesion formation, apply the 20-minute ultrasound signal and then allow structures to develop for a further 40 minutes by maintaining cells at 37 °C, 5% CO2.
Aspirate media and fix cells by applying 2 ml 4% paraformaldehyde in PBS. Incubate for 14 minutes at room temperature.
Aspirate paraformaldehyde and rinse three times with PBS, applying the PBS gently down the edge of the well to avoid generation of shear forces.
Transfer each coverslip from the 3.5 cm well to a 24-well plate for subsequent immunofluorescent staining.
Quench residual paraformaldehyde by applying 0.5 ml 0.1 M glycine in PBS to each of the 24 wells. Incubate for 20 minutes at room temperature.
Aspirate glycine and rinse three times with PBS.
Permeabilise cells by applying 0.5 ml 0.5% Triton X-100 (w/v) in PBS. Incubate for 4 minutes at room temperature.
Aspirate Triton X-100 and rinse three times with PBS.
Block by applying 0.5 ml 3% (w/v) BSA in PBS. Incubate overnight at 4 °C.
Stain vinculin-containing focal adhesion with hVIN-1 antibody diluted 1:400 in BSA block. Incubate for one hour at room temperature.
Rinse three times with PBS.
Apply fluorophore (DyLight 488)-conjugated secondary antibody (diluted 1:200) and stain actin with TRITC-conjugated phalloidin (diluted 1:200) in BSA block. Incubate for 30 minutes at room temperature.
Rinse three times with PBS, and once with water.
Mount on a glass slide in an inverted position using Prolong Gold mountant.
5. Quantification of Rac1 Activation by Pull-down Assay
For analysis of Rac1 activation, apply the 20-minute ultrasound signal to cells at 37 °C, 5% CO2. When assessing response times longer than 20 minutes, apply the ultrasound signal for 20 minutes and maintain cells at 37 °C, 5% CO2 for remaining time to allow the response to develop.
Aspirate media and place wells on ice. Rinse wells with 2 ml cold PBS and aspirate. Ensure any residual PBS is removed by leaving wells tilted for 1 minute followed by thorough aspiration.
Lyse cells on ice with 35 μl lysis buffer (10% glycerol, 20 mM Hepes pH 7.4, 140 mM NaCl, 1% NP40, 0.5% Sodium Deoxycholate, 4 mM EGTA, 4 mM EDTA, 1 × cOmplete protease inhibitor) per well. In this example, 6 wells are used per time point to generate 210 μl of cell lysate. More or fewer wells can be used but total lysate volume should total ~200 μl.
Scrape cells thoroughly with cell scraper to ensure complete cell lysis and leave wells tilted for 1 minute to allow lysate to pool.
Transfer lysate to a ice-cold 1.5-ml microfuge tube (pooling lysates from same treatment regime) and spin at 21000 × g for 2 minutes at 4 °C to pellet cellular debris.
Transfer 180 μl of lysate to ice-cold 1.5-ml microfuge tubes containing 130 μl of lysis buffer and 25 μl PAK-glutathione agarose beads10. Keep remaining crude lysate at -20 °C for subsequent gel separation.
Capture active Rac1 on the PAK-glutathione agarose beads by rotating tubes for 40 minutes at 4 °C.
Harvest agarose bead conjugates by centrifugation at 2000 × g for 1 minute at 4 °C.
Wash bead conjugates three times with 500 μl lysis buffer, harvesting beads by centrifugation at 2000x g and discarding supernatant after each wash. Carefully remove any remaining lysis wash buffer with a 200 μl pipette.
Elute each sample by adding 35 μl SDS-PAGE loading buffer and incubate at 85 °C on a shaking heat block for 5 minutes. Remove beads by centrifugation at 21000 × g for 1 minute.
Resolve bead eluate by SDS-PAGE, transfer to nitrocellulose and probe for Rac1.
6. Representative Results
In this protocol we describe the induction of vinculin-stained focal adhesions and Rac1 activity by ultrasound. For the focal adhesion experiment, the baseline position is that fibroblasts spread on a ligand of α5β1-integrin do not form vinculin-containing adhesions (Fig. 3A) unless a second fibronectin receptor, syndecan-4, is engaged by addition of soluble ligand (Fig. 3B) 3,4. However, stimulation with ultrasound induces focal adhesion formation to the same extent as engagement of syndecan-4 (Fig. 3C), indicating that ultrasound stimulation can substitute for certain components of the fibronectin-dependent signaling pathway 5. The key hurdle with this protocol is eliminating focal adhesion formation in the absence of stimulant. Incomplete inhibition of protein synthesis by cycloheximide will allow matrix deposition that is sufficient for the cells to auto-stimulate. Overcrowding of cells can also lead to low-level focal adhesion formation and a poor response to ultrasound as crosstalk between cell-cell and cell-matrix contacts will complicate an experiment that is designed to isolate a single stimulus. As a development of the basic assay we show the same experiment, repeated with fibroblasts lacking syndecan-4 (Fig. 4). These fibroblasts still respond to ultrasound (Fig. 4C), but fail to respond to syndecan-4 ligand (Fig. 4B). The experiment using Sdc4 -/- fibroblasts demonstrates that ultrasound can indeed bypass the need for certain fibronectin receptors, and illustrates how the simple protocol can be developed to gain extra information about the signaling pathway.
For the biochemical experiment we demonstrate that ultrasound can induce activation of Rac1, using a pull-down assay that is an adaptation of the method described by Del Pozo et al. 10. Precipitation of active Rac1 0, 10, 30 and 60 minutes after initiation of ultrasound stimulation reveals that Rac1 activity peaks at 10-30 minutes, before returning to the baseline (Fig. 5). Blotting crude lysates for vinculin ensures equivalent loading between time points. The result resembles activation of Rac1 by engagement of syndecan-43, but is slightly more protracted than activation by fibronectin. Therefore 8-10 repeats are typically required to achieve significant data. Activation of Rac1 is responsible for commitment of cells to focal adhesion formation, and nascent focal adhesions start to form at 10-30 minutes, coinciding with Rac1 activation. Once initiated, those adhesions will continue to mature into the large adhesion plaques shown in Figures 3 and 4.
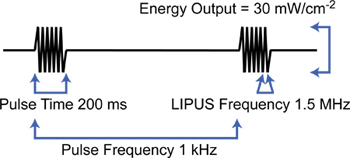 Figure 1. The ultrasound wave form. Ultrasound comprises an intermittent 1.5 MHz signal that due to low amplitude (30 mW/cm2, spatial average, temporal average) has no heating effect.
Figure 1. The ultrasound wave form. Ultrasound comprises an intermittent 1.5 MHz signal that due to low amplitude (30 mW/cm2, spatial average, temporal average) has no heating effect.
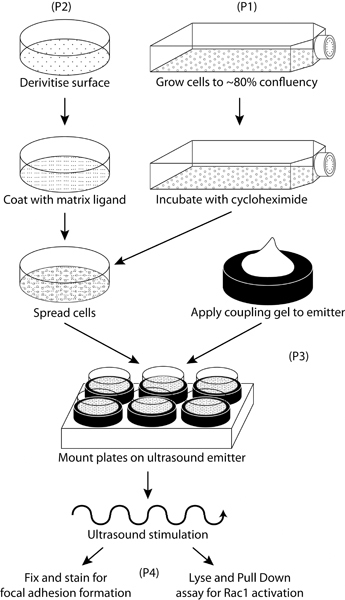 Figure 2. Schematic representation of the workflow for preparation and stimulation of cells with ultrasound.
Figure 2. Schematic representation of the workflow for preparation and stimulation of cells with ultrasound.
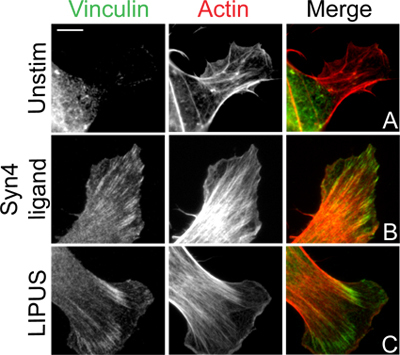 Figure 3. Ultrasound induction of focal adhesion formation in fibroblasts. Fibroblasts were spread on the integrin-binding fragment of fibronectin (A) before stimulation with the syndecan-4-binding fragment of fibronectin (B) or ultrasound (C). Following 60-minute stimulation, cells were fixed, stained for vinculin and actin, and imaged by epifluorescence. Bar = 10 μm.
Figure 3. Ultrasound induction of focal adhesion formation in fibroblasts. Fibroblasts were spread on the integrin-binding fragment of fibronectin (A) before stimulation with the syndecan-4-binding fragment of fibronectin (B) or ultrasound (C). Following 60-minute stimulation, cells were fixed, stained for vinculin and actin, and imaged by epifluorescence. Bar = 10 μm.
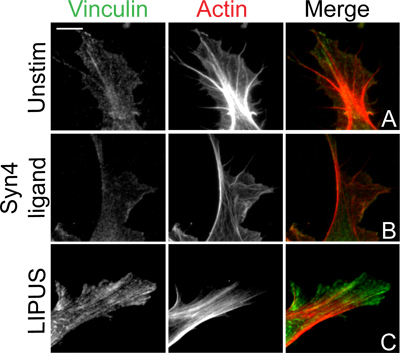 Figure 4. Ultrasound induction of focal adhesion formation in fibroblasts lacking syndecan-4. Sdc4 -/- fibroblasts were spread on the integrin-binding fragment of fibronectin (A) before stimulation with the syndecan-4-binding fragment of fibronectin (B) or ultrasound (C). Following 60-minute stimulation, cells were fixed, stained for vinculin and actin, and imaged by epifluorescence. Although Sdc4 -/- fibroblasts were fibronectin-insensitive, ultrasound still induced focal adhesion formation, indicating that ultrasound bypasses matrix receptor engagement. Bar = 10 μm.
Figure 4. Ultrasound induction of focal adhesion formation in fibroblasts lacking syndecan-4. Sdc4 -/- fibroblasts were spread on the integrin-binding fragment of fibronectin (A) before stimulation with the syndecan-4-binding fragment of fibronectin (B) or ultrasound (C). Following 60-minute stimulation, cells were fixed, stained for vinculin and actin, and imaged by epifluorescence. Although Sdc4 -/- fibroblasts were fibronectin-insensitive, ultrasound still induced focal adhesion formation, indicating that ultrasound bypasses matrix receptor engagement. Bar = 10 μm.
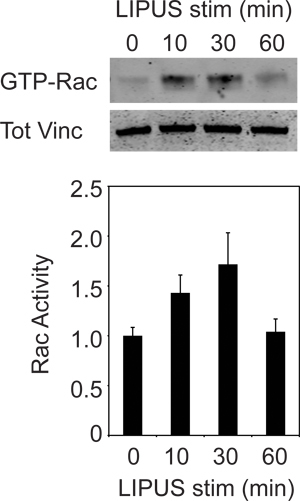 Figure 5. Activation of Rac1 by ultrasound. Cells were stimulated for 0, 10, 30 and 60 minutes with 6 wells used per time point. The amount of GTP-Rac1 present in samples from a Rac1 pull-down assay time course was visualized by incubation with anti-Rac1 antibody on a western blot (top image). Quantification of the band intensity (average of 8-10 replicates) reveals increased Rac1 activity resulting from ultrasound signal at 10 and 30 minutes, before returning to baseline levels at 60 minutes (graph). Error bars represent s.e.m.
Figure 5. Activation of Rac1 by ultrasound. Cells were stimulated for 0, 10, 30 and 60 minutes with 6 wells used per time point. The amount of GTP-Rac1 present in samples from a Rac1 pull-down assay time course was visualized by incubation with anti-Rac1 antibody on a western blot (top image). Quantification of the band intensity (average of 8-10 replicates) reveals increased Rac1 activity resulting from ultrasound signal at 10 and 30 minutes, before returning to baseline levels at 60 minutes (graph). Error bars represent s.e.m.
Discussion
In this protocol we describe the method by which a treatment that is normally applied to human patients can be used in cell-based experiments. The ultimate goal is to understand the molecular mechanism of ultrasound action so that the therapy can be refined. In this protocol, we use mouse embryonic fibroblasts (MEFs) as a model cell system, but ultrasound has been also been found to be effective in primary human foreskin fibroblasts 5, mesenchymal stem cells, osteoblasts and chondrocytes 6. We use Rac1 activation as an example biochemical assay, but the method could be used equally to test regulation of protein phosphorylation or formation of protein complexes by immunoprecipitation. For the immunofluorescent example we demonstrate an effect ultrasound on focal adhesion formation, but redistribution of any molecule could be examined. For example, one could test recruitment of cytosolic factors to the plasma membrane, colocalisation of proteins in trafficking vesicles or examine the effect of ultrasound on mitotic spindle organization. So far we have confined ourselves to testing fibronectin-dependent pathways, by the effect of ultrasound on cells on collagen, or in the presence of growth factors could be tested to build up a picture of how ultrasound affects cell behavior in a complex environment that more closely resembles an in vivo situation. A greater challenge will be to test the effect of ultrasound using time-lapse imaging, where the presence of the emitter blocking the light path and vibrations from the ultrasound present additional technical obstacles. However, the fact that therapeutic application requires only 20 minutes of stimulation per day suggests that analysis of cell behavior after a burst of stimulation may well be productive.
Disclosures
Andrew Harrison is an employee of Smith & Nephew UK Limited.
Acknowledgments
This work was supported by Wellcome Trust grant 088419 to MDB and sponsorship by Smith & Nephew UK Ltd.
References
- Bass MD. A syndecan-4 hair trigger initiates wound healing through caveolin- and RhoG-regulated integrin endocytosis. Dev. Cell. 2011;21:681–693. doi: 10.1016/j.devcel.2011.08.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Echtermeyer F. Delayed wound repair and impaired angiogenesis in mice lacking syndecan-4. J. Clin. Invest. 2001;107:R9–R14. doi: 10.1172/JCI10559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bass MD. Syndecan-4-dependent Rac1 regulation determines directional migration in response to the extracellular matrix. J. Cell Biol. 2007;177:527–538. doi: 10.1083/jcb.200610076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woods A, Couchman JR, Johansson S, Hook M. Adhesion and cytoskeletal organisation of fibroblasts in response to fibronectin fragments. Embo. J. 1986;5:665–670. doi: 10.1002/j.1460-2075.1986.tb04265.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mahoney CM, Morgan MR, Harrison A, Humphries MJ, Bass MD. Therapeutic ultrasound bypasses canonical syndecan-4 signaling to activate rac1. J. Biol. Chem. 2009;284:8898–8909. doi: 10.1074/jbc.M804281200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Claes L, Willie B. The enhancement of bone regeneration by ultrasound. Prog. Biophys. Mol. Biol. 2007;93:384–398. doi: 10.1016/j.pbiomolbio.2006.07.021. [DOI] [PubMed] [Google Scholar]
- Heckman JD, Ryaby JP, McCabe J, Frey JJ, Kilcoyne RF. Acceleration of tibial fracture-healing by non-invasive, low-intensity pulsed ultrasound. J. Bone Joint Surg. Am. 1994;76:26–34. doi: 10.2106/00004623-199401000-00004. [DOI] [PubMed] [Google Scholar]
- Kristiansen TK, Ryaby JP, McCabe J, Frey JJ, Roe LR. Accelerated healing of distal radial fractures with the use of specific, low-intensity ultrasound. A multicenter, prospective, randomized, double-blind, placebo-controlled study. J. Bone Joint Surg. Am. 1997;79:961–973. doi: 10.2106/00004623-199707000-00002. [DOI] [PubMed] [Google Scholar]
- Danen EH. Requirement for the synergy site for cell adhesion to fibronectin depends on the activation state of integrin alpha 5 beta 1. J. Biol. Chem. 1995;270:21612–21618. doi: 10.1074/jbc.270.37.21612. [DOI] [PubMed] [Google Scholar]
- Del Pozo MA, Price LS, Alderson NB, Ren XD, Schwartz MA. Adhesion to the extracellular matrix regulates the coupling of the small GTPase Rac to its effector PAK. Embo. J. 2000;19:2008–2014. doi: 10.1093/emboj/19.9.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Makarem R. Competitive binding of vascular cell adhesion molecule-1 and the HepII/IIICS domain of fibronectin to the integrin alpha 4 beta 1. J. Biol. Chem. 1994;269:4005–4011. [PubMed] [Google Scholar]