

Abstract
Bordetella avium continues to be an economic issue in the turkey industry as the causative agent of bordetellosis, which often leads to serious secondary infections. This study presents a broad characterization of the antibiotic resistance patterns in this diverse collection of B. avium strains collected over the past thirty years. In addition, the plasmid basis for the antibiotic resistance was characterized. The antibiotic resistance pattern allowed the development of a novel enrichment culture method that was subsequently employed to gather new isolates from diseased turkeys and a healthy sawhet owl. While a healthy turkey flock was shown to seroconvert by four weeks-of-age, attempts to culture B. avium from healthy turkey poults were unsuccessful. Western blot of B. avium strains using pooled serum from diseased and healthy commercial turkey flocks revealed both antigenic similarities and differences between strains. In sum, the work documents the continued exposure of commercial turkey flocks to B. avium and the need for development of an effective, inexpensive vaccine to control spread of the disease.
Keywords: Bordetellosis incidence, serology, antibiotic resistance, Bordetella avium, poultry
Introduction
Bordetella avium, the causative agent of bordetellosis in turkeys, continues to have significant, negative economic impacts in the turkey industry (Jackwood and Saif, 2008). Bordetellosis is a highly infectious, upper respiratory infection resulting in high morbidity and low mortality (Jackwood and Saif, 2008). Diseased turkeys fail to gain weight and are more susceptible to secondary infections. A 2009 report names B. avium the third infectious agent in a list of most challenging health issues for the US turkey industry as reported by poultry growers and veterinarians (Clark. 2009). B. avium has been surveyed in a number of studies (Clubb et al., 1994; Hinz and Glunder, 1985; Hopkins et al., 1990; Malik et al., 2005; McBride et al., 1991; Raffel et al., 2002; van den Hurk. 1992), and has been consistently detected by culture or specific antibodies in commercial turkey flocks, non-commercially raised turkeys, and other wild and domesticated avian species. B. avium can also infect chickens, but is considered to be an opportunistic pathogen in that host (Jackwood et al., 1995; Jackwood and Saif, 2008). Although B. avium has been associated with disease in humans (Harrington et al., 2009; Spilker et al., 2008), it does not appear to present a public health problem.
As a major turkey-growing region in the United States, turkeys in the Shenandoah Valley of Virginia are routinely monitored by serology for the presence of B. avium. An existing vaccine (ArtVax©) offers some protection against severe disease, but does not limit infection or spread (Jackwood and Saif, 2008), and is used by 10% or less of farmers in the Shenandoah Valley. This study explores the antibiotic resistance profiles of a variety of isolates, provides a novel enrichment culture protocol for B. avium, and further explores the presence of B. avium in the commercial turkeys in the Shenandoah Valley, with the ultimate goal of a successful attenuated vaccine strain.
Methods
Growth and maintenance of bacterial cultures
Our main laboratory strain, B. avium 197N, was derived from a spontaneous Nalr isolate of Gobl-124 (Gentry-Weeks et al., 1995), as described previously (Temple et al., 1998). B. avium was routinely grown and maintained on brain heart infusion (BHI) agar or broth (Fisher Scientific) with addition of 10 μg/ml aztreonam and 20 μg/ml ampicillin (Sigma-Aldrich) after our discovery of the differential antibiotic resistance described herein. For phenotypic tests, MacConkey agar and Bordet-Gengou with 12% sheep blood agar plates were used. B. avium strains discovered and/or used in this study are listed in Table 1.
Table 1.
Bacterial isolates used
| Isolate | Source | Year of Isolation |
|---|---|---|
| 4143 | diseased turkey | pre-1979 |
| Gobl-151, Ba169, Gobl-146, Gobl-142A, BA177 | diseased turkey | pre-1990 |
| Gobl-124 (also known as 197) (Gentry-Weeks et al., 1995) | 197N parent; Tetr Nals | pre-1990 |
| 197N (Temple et al., 1998) | laboratory strain; Tets Nalr | 1993 |
| T4 (Raffel et al., 2002) | healthy wild turkey | 1998 |
| D10, D23 (Raffel et al., 2002) | healthy duck | 1998 |
| G24 (Raffel et al., 2002) | healthy goose | 1998 |
| JBBA (NCSU) | diseased turkey | 2001 |
| VT-1, VT-4 (Virginia Tech) | diseased turkey | pre-2004 |
| SV-1 | diseased turkey | 2004 |
| Owl19 | healthy owl | 2006 |
| VDAC4219A, VDAC2499, VDAC22CD5, 15 others | diseased turkey (VDACS) | 2008–2010 |
| 197N+pRAM | 197N with pRAM plasmid; Tetr Nalr | 2008 |
| B. hinzii WC3-4 | diseased turkey | 2001 |
| B. hinzii SV-2 | diseased turkey | 2004 |
| E. coli+pRK2013 (Figurski and Helinski, 1979) | laboratory strain, Tra+ | n/a |
Antibiotic Resistance Testing
Non-fermenting Sensititre plates (Trek Diagnostics, Cleveland, OH) were used according to manufacturer’s instructions, to establish the sensitivity/resistance of various B. avium strains to a panel of antibiotics. To further establish the antibiotic resistance profile, several colonies from overnight cultures grown on BHI agar plates were added to 5 ml of dH2O to match the McFarland standard absorbance of 0.115, and 10 μl of the bacterial suspension was then added to 10 ml Mueller-Hinton broth (22 mg/ml) and vortexed. This broth was applied to each well of the plate (50 μl/well) and incubated at 37 °C for 18–24 hours before being analyzed for absorbance at 600 nm with a BioRad microplate absorbance reader and interpreted according to manufacturer’s directions. High levels of resistance were defined as equal to or greater than the maximum dose (supplementary data). Antimicrobial resistance in bacteria isolated from animals has been extensively studied, and the methodologies are inconsistent ((Schwarz et al., 2010), and our observations). The Sensititre system we utilized provides information and data that can be repeated by others using the same kit. Our goal in determining the resistance/sensitivity patterns of our isolates was to find an improved in vitro isolation method. However, we include in Table 2 and the supplementary data the entire panel for readers interested in these patterns for different reasons. Since most isolates showed some resistance to aztreonam and ampicillin, the resistance of B. avium to these antibiotics was studied in greater detail by plating equal numbers of colony forming units (CFU) on BHI plates containing 5, 10, 20, or 50 μg/ml of each antibiotic. Media concentrations of 20 μg/ml ampicillin and 10 μg/ml aztreonam were used for all future bacterial screening.
Table 2.
Antibiotic resistance of select B. avium and B. hinzii isolates*
| Antibiotic | Bacterial Isolate
|
||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 197N | 197N+pRAM | Gobl-124 | BA177 | Gobl-146 | VT-4 | 4143 | Gobl-151 | Gobl-142A | T4 | BA169 | G24 | B. hinzii SV-2 | |
| cefotaxime | ● | ● | ● | ||||||||||
| ceftriaxone | ○ | ○ | ○ | ○ | ○ | ● | |||||||
| ticarcillin | ● | ○ | ○ | ||||||||||
| carbenicillin | ● | ||||||||||||
| ampicillin/sulbactam 2:1 | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ||||||
|
| |||||||||||||
| ticarcillin/clavulonic acid | ○ | ||||||||||||
| lomefloxacin | ● | ○ | ○ | ○ | |||||||||
| ciprofloxacin | ○ | ○ | ○ | ○ | |||||||||
| levofloxacin | ○ | ||||||||||||
| imipenem | ● | ||||||||||||
|
| |||||||||||||
| aztreonam | ● | ● | ● | ● | ● | ○ | ● | ● | ● | ● | ● | ○ | ● |
| tobramycin | ○ | ● | |||||||||||
| sulfamethox/trimethoprim | ● | ● | ● | ||||||||||
| sulfisoxazole | ● | ● | ● | ● | ● | ||||||||
| tetracycline | ● | ● | |||||||||||
| chloramphenicol | ○ | ○ | ○ | ○ | |||||||||
● = complete resistance
○ = intermediate resistance
no circle = sensitive to antibiotic in Sensititre assay conditions
Not all isolates listed in Table 1 were tested in this system
Tetracycline resistance was determined in select B. avium strains using tubes with 2 ml of BHI broth containing tetracycline concentrations of 12, 20, 36, 48, 60, 72, 84 and 96 μg/ml, respectively. Duplicate tubes were inoculated with each strain of interest and incubated overnight at 37 °C in a shaking incubator, and quantified by spectroscopy at 600 nm. Additionally, triparental conjugal matings were performed between E. coli containing plasmid pRK2013 (Tra+), B. avium strain Gobl-124 [tetracycline resistant (Tetr)], and B. avium strain 197N [tetracycline sensitive (Tets), naladixic acid resistant (Nalr)] (Temple et al., 1998). To select for B. avium strains transformed to Tetr via a conjugable plasmid, matings were plated on BHI agar containing 50 μg/ml tetracycline, 10 μg/ml aztreonem, and 30 μg/ml naladixic acid.
PCR amplification and genetic confirmation
PCR for specific identification of B. avium was performed as previously described using primers (Register and Yersin, 2005; Savelkoul et al., 1993); presence of the amplicon identified B. avium. For 16S-rDNA amplification, two universal primers were used to produce a 1484 bp fragment [Bac8f: 5′-AGA GTT TGA TCC TGG CTC AG-3′ and Univ1492r: 5′-GGT TAC CTT GTT ACG ACT T-3′ (Baker et al., 2003)]. PCR was performed using a MyCycler thermocycler (Biorad). Gel electrophoresis for PCR product confirmation was carried out using routine procedures (Sambrook and Russell, 2001). Sequencing of 16S-rDNA amplicons was performed by Elim Biopharmaceuticals (Hayward, CA), and comparisons to known sequences were made using BLAST (McGinnis and Madden, 2004). Samples were considered positive if published B. avium sequences were the highest scoring matches.
Cloning of DNA fragments from pRAM
The naturally occurring plasmid, pRAM, was isolated from B. avium Gobl-124 (Temple et al., 1998), using a plasmid purification kit (Qiagen). Amplification of pRAM segments was performed using random primers, and amplicons were cloned into the pCR2.1 TOPO vector (Invitrogen). Resulting plasmids were prepared with the Qiagen kit and sequenced at Elim Biopharmaceuticals (Hayward, CA).
Sample collections from turkey poults and owls
In the fall of 2007, blood samples and tracheal swabs were collected at a commercial turkey farm from 5 day-old birds (n=10–12 birds per collection date) at approximately two week intervals through eight weeks-of-age. Additional blood samples and tracheal swabs were obtained from different commercial turkey farms in 2010 as part of routine serological examinations in the Shenandoah Valley. In December of 2006 and 2007, tracheal swabs were collected from more than 200 migrating sawhet owls captured in mist nets and banded. Additionally, 25 blood samples were collected from owls in December 2007. Blood samples were centrifuged after clotting 2000 xg for 10 min, and serum was transferred to microtubes and stored at −20 °C until testing. Tracheal swabs were cultured according to our standard protocol for isolation of B. avium from mixed cultures.
Serology
The ProFLOK BA-T ELISA kit (Synbiotics, Kansas City, KS) was used to detect and quantify B. avium antibodies according to manufacturer’s instructions, each sample performed in triplicate. For statistical analysis, a one-tailed, unpaired t-test was performed on the calculated titer values. Significance was determined using a critical p value of 0.05. Additionally, serology reports from commercial turkey farms were provided by the veterinarians of those farms.
Western blots
Western blots were performed using selected B. avium isolates and turkey serum from diseased turkeys from the Shenandoah Valley, some of which had been collected by the College of Veterinary Medicine at VA Tech (kind gift of Dr. F. William Pierson) and others by the Virginia Department of Agriculture and Consumer Services (VDACS) regional laboratory (kind gift of Dr. David Brown). Bacteria were grown overnight in BHI broth at 37°C with shaking. Membrane-enriched fractions were prepared by lysing the cells from overnight cultures with shearing (French pressure cell) and centrifugation to collect the membrane fragments. Since finding antigenic proteins was the goal, absolute protein content was not important at this stage; thus, relatively similar amounts of protein were loaded by harvesting overnight cultures at identical optical densities. Ten microliters of each denatured sample was added to wells of 4–12% gradient SDS polyacrylamide gels (Invitrogen) and run according to manufacturer’s instruction. Transfer to nitrocellulose membranes was accomplished using a semi-dry transfer apparatus (BioRad). Proteins immobilized on nitrocellulose were probed with turkey serum diluted 1:100 with Tris-buffered saline with Tween-20 (TBS-T) and 3% casein from three sources: (1) serum from healthy commercial turkey poults (collected during this study), (2) serum from turkeys experimentally infected with the laboratory strain 197N, (3) serum collected from commercial turkey poults following a disease outbreak (two different farms). Membranes were incubated with antisera overnight at room temperature, followed by 3 washes with TBS-T, and 2 hour incubation with 1:5000 diluted horseradish peroxidase labeled rabbit anti-turkey/chicken IgG antibody (Invitrogen). After six washes with TBS-T, reactive proteins were detected using diaminobenzidene (Sigma).
Results
Antibiotic sensitivity of B. avium isolates
In order to determine antibiotic resistance characteristics of B. avium, 12 B. avium isolates and 1 B. hinzii isolate were tested using the Sensititre assay (Table 2). Five B. avium isolates were found to be sensitive to all of the antibiotics tested in the Sensititre assay: SV-1-2006, D10, JBBA, D23, and Owl19 (supplementary data, which defines the antibiotic concentration designated high, medium, and low). High or intermediate levels of resistance to the third generation cephalosporins (cefotaxime or ceftriaxone) were observed in 197N, 197N+pRAM, and B. hinzii SV-2 (Table 2). A high level of resistance to penicillins (ticarcillin and carbenicillin) was seen only in Gobl-124 (Table 2). Intermediate resistance to the ampicillin/sulbactam combination was found in B. hinzii SV-2 and 6/17 B. avium isolates tested, while all but one isolate were sensitive to a ticarcillin/clavulonic acid mixture (Table 2). Since the Sensititre assay did not test ampicillin alone, we used the agar plate method to test all 17 B. avium isolates in the collection; all 17 were determined to be resistant to 20 μg/ml ampicillin (data not shown). High or intermediate levels of resistance to the second generation fluoroquinolones (lomefloxacin and ciprofloxacin) were observed in 197N, 197N+pRAM, and T4, while only a single isolate (BA177) was resistant to the carbapenem imipenem (Table 2). A high or intermediate level of resistance to the monobactam aztreonam was found in all isolates except D10, Owl19, JBBA, D23, and SV-1-2006 (Supplemental data). When tested for aztreonam resistance in a plate resistance assay, all 17 B. avium isolates were able to grow on BHI agar plates containing 10 μg/ml aztreonam (data not shown). High levels of resistance to the sulfonamides (sulfamethoxazole/trimethoprim and sulfisoxazole) were seen in Gobl-124, 197N+pRAM, and T4 (Table 2). A high level of resistance to tetracycline was observed in Gobl-124 and 197N+pRAM. Intermediate levels of resistance to chloramphenicol were found for strains 197N+pRAM, Gobl-146, T4, and B. hinzii SV-2 (Table 2). All isolates tested were sensitive to gentamicin, cefoperazone, cefepime, ceftazidime, piperacillin, and amikacin (supplemental data).
Mechanism of antibiotic resistance in B. avium
Tetracycline resistance has been associated with a limited number of B. avium strains (Cutter and Luginbuhl, 1991), including the parent strain of our common laboratory strain, Gobl124 (Table 1). A plasmid was successfully isolated from strain Gobl124 (pRAM), and restriction fragment analysis indicated an approximate size of 12–13 kb (data not shown). During subculture of 197N for a naladixic acid resistant phenotype, the pRAM plasmid was lost. Using standard triparental mating between 197N, Gobl124, and E. coli pRK2013, pRAM was transferred successfully via conjugation, transferring the tetracycline resistance phenotype to strain 197N (197N+pRAM). Based on the Sensititre data, pRAM also carries resistance genes to two sulfonamides, sulfisoxazole and sulfamethoxazole/trimethoprim (Table 2). Partial DNA sequence analysis of pRAM (GenBank accession # HQ908281) revealed two genes for conjugation, similar to P-type conjugative transfer ATPase TrbB and TrbC of Enterobacter aerogenes (Thorsted et al., 1998). The predicted amino acids from the sequence were 99% identical to 133 residues (out of a total of 320 residues) at the C-terminus of trbB and 93% identical to the first 100 amino acid residues (out of a total of 154 residues).
New B. avium isolates
Since the plate assays for ampicillin and aztreonam showed B. avium isolates to have consistent resistance patterns, including those sensitive to higher concentrations measured in the Sensititre test, the laboratory began routine culturing of environmental samples with these two antibiotics. Colonies that grew in broth or on plates containing 10 μg/ml aztreonam and 20 μg/ml ampicillin, followed by phenotypic (lack of hemolysis on blood agar, non-fermenting growth on McConkey agar) and genotypic verification by species-specific PCR testing, a total of 18 B. avium isolates (VDACS isolates, Table 1) were cultured from 596 tracheal samples from commercial turkeys with respiratory disease of unknown etiology (~3%). These samples were collected over a period of over 3 years in the Shenandoah Valley and represent birds from different flocks. Another B. avium isolate (Owl19, Table 1) was recovered from sampling of more than 200 sawhet owls (~0.5%) being banded in a migration survey in the Allegheny Mountains. This is the first reported B. avium isolated from an owl, although we have previously reported two seropositive owls (Raffel et al., 2002). None of the 25 owl serum samples collected in this study tested positive for B. avium antibodies. Owl19, isolated in 2006, did not have serological testing performed on it since blood was not drawn from that sampled owl. Because of the ongoing perception that B. avium can survive for long periods and is carried in water (Raffel et al., 2002), we have initiated efforts to identify the organism using culture and/or PCR methods. However, in two collections of 2–3 liters of stream or standing water near poultry farms in our area, we were unsuccessful in finding evidence of the organism. Efforts to screen larger volumes of water in areas suspected to have B. avium present to isolate the organism are planned.
Detection of B. avium-specific antibodies in serum of turkey poults at commercial farms
To observe the kinetics of flock seroconversion to B. avium in a local turkey farm, serum samples and tracheal swabs were collected from a single flock at approximately two-week intervals in 2007. Swabs were expressed onto MacConkey plates with no antibiotics, and cultured for 48 hours at 37 °C. All poults tested at 5 days and 15 days of age were sero-negative, while seroconversion was observed in the flock at 30 days old, with all birds having detectable B. avium-specific antibodies (Figure 1A). Despite the clear seroconversion of the flock, B. avium was not isolated from any of the tracheal swabs collected and the birds showed no sign of disease.
Figure 1. Bordetella avium-specific antibodies detected in serum of commercial turkey poults by ELISA.
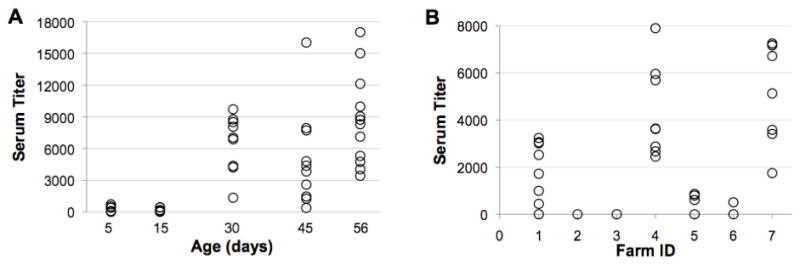
(A) Time-course study of B. avium-specific antibody titers at a single commercial turkey farm. Blood samples were randomly collected at a commercial turkey farm at approximately two-week intervals (n=10–12 birds per collection date) and tested for B. avium-specific antibodies using a commercially available ELISA kit (Synbiotics). (B) Distribution of B. avium-specific antibodies detected in the serum of turkey poults at different commercial turkey farms. Blood samples were randomly collected from 8–10 week-old turkey poults (n=8 birds per farm) at seven different commercial turkey farms in the Shenandoah Valley, Virginia, and tested for B. avium-specific antibodies using a commercially available ELISA kit (Synbiotics).
Routine screening of 8–10 week-old turkeys at seven commercial turkey farms in our region from summer and fall of 2010, confirm that B. avium exposure is widespread. ELISA results from the commercial veterinarians showed that 3/7 flocks had moderate to high levels of serum antibodies, while only 2/7 flocks were apparently seronegative (Figure 1B).
Antigenic similarity of diverse isolates of B. avium
To determine the immunological profile of multiple isolates of B. avium, western blotting was used. Pooled antisera from several sources were used as a probes, including serum from healthy (Fig. 2A), experimentally infected (Fig. 2B), and diseased commercial turkey flocks (Fig. 2C & Fig. 2D). Multiple proteins of various sizes were common among the different samples tested, including ones at 70, 52, 22, 19, 14, and 12 kDa (Figure 2). An unidentified non-Bordetella bacterium isolated from a tracheal swab from a diseased turkey (sharing no culture characteristics with B. avium) was included as a negative control (Lane 1 in each panel). Each serum pool detected similar proteins in the B. avium samples, which include isolates from diseased turkeys as well as a healthy goose and owl. These isolates were collected over a 30 year period and included two isolates collected in the past two years (VDAC4219A and 2499). The 22 kDa band seems to be universally present, and the 70, 19, and 14 kDa bands are frequently detected, dependent on the pool of antiserum used. A total of four B. hinzii isolates were tested, and shown to be indistinguishable from B. avium by western blot (Figure 2 and data not shown).
Figure 2. Western blots of membrane-enriched bacterial protein preparations using reactive antiserum from commercial turkeys.
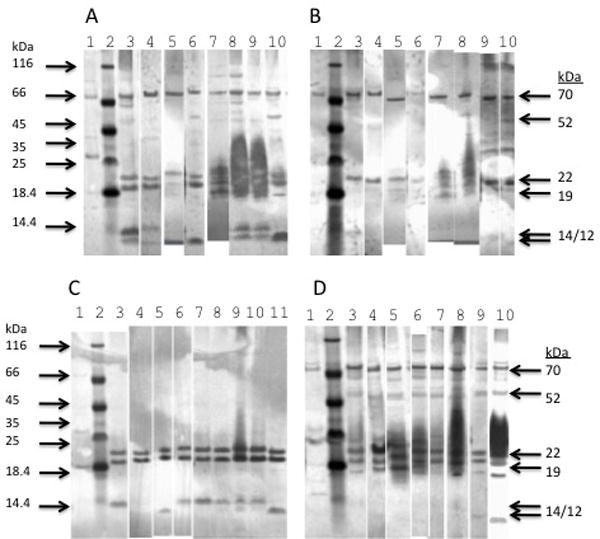
Membrane-enriched protein samples were prepared from overnight cultures of VT-1 (lane 3), VDAC4219A (lane 4), 197N (lane 5), B. hinzii WC3-4 (lane 6), G24 (lane 7), SV-1 (lane 8), Owl19 (lane 9), VDAC2499 (lane10), and VDAC22CD5 (panel C only), and SDS-PAGE was performed with denaturing conditions. A non-Bordetella isolated from turkey trachea was used as a negative control (lane 1). Molecular weight standards (BioRad) were included in each panel (lane 2), with size (kDa) of molecular weight standards displayed on left. After transfer to nitrocellulose membrane, proteins were probed with pooled serum from four sources: Panel A: pooled serum from clinically healthy turkeys at a single commercial turkey farm in the Shenandoah Valley, 2007 (Figure 1A); Panel B: pooled serum from turkeys experimentally infected with laboratory strain 197N; Panel C & D: pooled serum from diseased turkeys at two different commercial farms in the Shenandoah Valley. Sizes of most common B. avium antigenic proteins shown on the right. Panel lanes are combined from three gels to optimize the exposure and illustrate the banding patterns.
Discussion
Bordetellosis, caused by B. avium, continues to cause economic and animal welfare problems in the commercial turkey industry. Quantitative data on the incidence of outbreaks and strain characterization is essential for vaccine development; however, the isolation and identification of B. avium is seldom carried out in a rigorous manner because bordetellosis is not a reportable disease. This study describes a new systematic approach to the examination of B. avium infection in commercial turkey populations. The enrichment and isolation scheme we have developed exploits the bacterium’s natural resistance to the antibiotics aztreonam and ampicillin. Growth of mixed bacterial populations, including those from environmental samples and tracheal swabs, in rich media supplemented with aztreonam and ampicillin led to the successful counter-selection of many Pseudomonads and enteric bacteria. Following aztreonam enrichment, B. avium isolates were further distinguished from B. hinzii, via hemolysis and hemagglutination phenotyping. Isolates were ultimately confirmed by PCR with B. avium-specific primers (Register and Yersin, 2005). It is important to emphasize that the concentration of aztreonam used in the plate isolation method (10 μg/ml) is much lower than the concentration tested in the Sensititre assay, and all isolates that were “sensitive” to aztreonam in the Sensititre test were capable of growth at this concentration.
In this study, our antibiotic enrichment scheme was used to successfully isolate B. avium from tracheal swabs of sick birds, samples of which were obtained from Virginia diagnostics laboratories (VDACS isolates, Table 1), as well as an apparently healthy sawhet owl (Owl19). Attempts to isolate B. avium from farm run-off waterways and tracheal swabs of healthy turkey poults were unsuccessful, most likely due to low sample sizes. We were able to document B. avium seroconversion in a clinically healthy flock of turkey poults (Figure 1A), demonstrating that although clinical bordetellosis may not be observed in a population, the bacterium is present at levels sufficient to elicit an immune response in young birds.
Antibiotic resistance in B. avium has been studied previously in several reports. In 1998–99, four B. avium isolated in Minnesota tested sensitive to ampicillin and tetracycline (among others), and all four were resistant to erythromycin (Malik et al., 2005). Also in Minnesota, reports over five years (1998 to 2002) show decreasing resistance to tetracycline (100% to 0%) and variable resistance to ampicillin (14% to 50%) (Malik, Y.S., Olsen, K., Chander, Y., Goyal, S.M. 2003). Mortensen reported MIC levels for 10 B. avium isolates with similar variability from Australian isolates (Mortensen et al., 1989). MICs were determined for 18 B. avium isolates that had low to no resistance to ampicillin and tetracycline (Blackall et al., 1995). In an effort to optimize our procedures for isolating bacteria from environmental samples, we established resistance for a panel of antibiotics, using the Sensititre assay to characterize several B. avium isolates from a variety of sources. This screening revealed consistent resistance to aztreonam, a monobactam, which we have successfully utilized to exclude other bacteria from mixed cultures. Previous reports did not include this particular antibiotic. In addition to resistance of all B. avium isolates tested at concentrations up to 20 μg/ml aztreonam, we also found that all B. hinzii isolates tested are resistant to the same concentration (data not shown). We speculate that resistance of the isolates may be due to a lack of the penicillin binding protein 3 (PBP3) gene (Georgopapadakou et al., 1982, American Society of Hospital Pharmacists. 1998) shown in the complete genome sequence of B. avium strain 197N (Sebaihia et al., 2006). The mechanism of action for aztreonam involves the drug binding to this protein to inhibit cell wall synthesis. In addition to a high affinity for PBP3, aztreonam also has a low binding affinity to PBP1a (Georgopapadakou et al., 1982). The presence of genes for PBP1a or PBP3 in other strains may explain any variation in aztreonam resistance or sensitivity. Mutagenesis testing is required to confirm or deny these hypotheses.
Tetracycline resistance has been associated with a limited number of B. avium isolates (Cutter and Luginbuhl, 1991), including Gobl-124, the parent strain of our common laboratory strain 197N. A plasmid was successfully isolated from Gobl-124 (pRAM), and restriction fragment analysis indicated an approximate size of 12–13 kb (data not shown). Using standard triparental mating between 197N, Gobl-124, and E. coli +pRK2013, pRAM was transferred successfully via conjugation, transferring the tetracycline resistance phenotype to strain 197N (197N+pRAM). Based on the Sensititre data, pRAM also carries resistance genes to two sulfonamides, sulfisoxazole and sulfamethoxazole/trimethoprim (Table 2). Plasmids carrying tetracycline and sulfonamide resistance have been isolated from B. avium in the past. Cutter et al. studied five plasmids varying in size from 16 to 51.5 kb (Cutter and Luginbuhl, 1991). All of these plasmids conferred resistance to sulfonamides, and three of them also conferred tetracycline resistance. It is not thought that the plasmid isolated in this study is one of the ones isolated in the previous studies because pRAM is much smaller (12–13 kb) than those studied previously (Cutter and Luginbuhl, 1991; Morris and Luginbuhl, 1992). It is possible that previous B. avium isolates that are tetracycline resistant also harbor a plasmid similar or identical to pRAM. We have shown that this plasmid is conjugable, both by empirical evidence and by identification of conjugation machinery in a partial sequence from the plasmid.
Humoral immunity appears to be the mechanism of disease resolution and protection from recurrence (Jackwood and Saif, 2008). However, the actual antigenic proteins have not been previously characterized. The overall antigenic banding pattern of several B. avium isolates had bands in common when tested with pooled serum from a single flock (Fig. 2); however, there were some differences in the reactivity of the serum pools used. Specifically, the higher molecular weight bands (52 and 70 kDa) were not visualized using the serum from one of the diseased turkey flocks (Fig. 2C) while there were seemingly a larger number of bands visualized in general by the serum from the other diseased flock (Fig. 2D). It is possible that differences in banding patterns are due to differences in the strain infecting the turkeys, or simply variation among the turkeys during seroconversion. However, prominent reactive bands were seen at 19–22kDa in all of the blots, and are likely fimbrial proteins. Protective immunity elicited by purified fimbriae (Fim) has been documented (Akeila and Saif, 1988). There are 11 putative fimbrial subunit genes (fim) in the genome of B. avium 197N (Sebaihia et al., 2006), and expression of several fim genes has been shown by reverse transcriptase PCR (Loker et al., 2011) and data not shown. Considering the intense specific staining and relative conservation of the 19–22 kDa fimbrial proteins across all isolates, it is likely that all B. avium isolates share these common antigens. This data supports the possible utility of a single live-attenuated B. avium vaccine strain that would be capable of eliciting a broadly-protective immune response in turkeys. Future work in our laboratory will focus on the development and characterization of such a vaccine.
Supplementary Material
Acknowledgments
This work was supported in part by NIH R-15-AI057382-03 to Temple and Miyamoto. We thank Mr. Charles Zieginfuss who collected the sawhet owl samples and the poultry grower who assisted in our collections from his farm. We also thank Dr. F. William Pierson and Dr. David Brown for B. avium isolates and tracheal samples. Dr. has generously consulted with us on this project, and Stan Gilliland III contributed technical assistance to the work.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Akeila MA, Saif YM. Protection of Turkey Poults from Bordetella avium Infection and Disease by Pili and Bacterins. Avian Dis. 1988;32:641–649. [PubMed] [Google Scholar]
- American Society of Hospital Pharmacists. American hospital formulary service drug information. Published by authority of the Board of Directors of the American Society of Hospital Pharmacists; Bethesda, MD: 1998. Aztreonam Monograph. [Google Scholar]
- Baker GC, Smith JJ, Cowan DA. Review and Re-Analysis of Domain-Specific 16S Primers. J Microbiol Methods. 2003;55:541–555. doi: 10.1016/j.mimet.2003.08.009. [DOI] [PubMed] [Google Scholar]
- Blackall PJ, Eaves LE, Fegan M. Antimicrobial Sensitivity Testing of Australian Isolates of Bordetella avium and the Bordetella avium-Like Organism. Aust Vet J. 1995;72:97–100. doi: 10.1111/j.1751-0813.1995.tb15018.x. [DOI] [PubMed] [Google Scholar]
- Clark S. Current Health and Industry Issues Facing the Turkey Industry 2009 [Google Scholar]
- Clubb SL, Homer BL, Pisani J, Head C. Outbreaks of Bordetellosis in Psittacines and Ostriches. Proc Assoc Avian Vet. 1994:63–68. [Google Scholar]
- Cutter DL, Luginbuhl GH. Characterization of Sulfonamide Resistance Determinants and Relatedness of Bordetella avium R Plasmids. Plasmid. 1991;26:136–140. doi: 10.1016/0147-619x(91)90054-z. [DOI] [PubMed] [Google Scholar]
- Figurski DH, Helinski DR. Replication of an Origin-Containing Derivative of Plasmid RK2 Dependent on a Plasmid Function Provided in Trans. Proc Natl Acad Sci U S A. 1979;76:1648–1652. doi: 10.1073/pnas.76.4.1648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gentry-Weeks CR, Spokes J, Thompson J. Beta-Cystathionase from Bordetella avium. Role(s) of Lysine 214 and Cysteine Residues in Activity and Cytotoxicity. J Biol Chem. 1995;270:7695–7702. doi: 10.1074/jbc.270.13.7695. [DOI] [PubMed] [Google Scholar]
- Georgopapadakou NH, Smith SA, Sykes RB. Mode of Action of Azthreonam. Antimicrob Agents Chemother. 1982;21:950–956. doi: 10.1128/aac.21.6.950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harrington AT, Castellanos JA, Ziedalski TM, Clarridge JE, III, Cookson BT. Isolation of Bordetella avium and Novel Bordetella Strain from Patients with Respiratory Disease. Emerging Infectious Diseases. 2009;15:72–74. doi: 10.3201/eid1501.071677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hinz KH, Glunder G. Occurrence of Bordetella avium Sp. Nov. and Bordetella bronchiseptica in Birds. Berl Munch Tierarztl Wochenschr. 1985;98:369–373. [PubMed] [Google Scholar]
- Hopkins BA, Skeeles JK, Houghten GE, Slagle D, Gardner K. A Survey of Infectious Diseases in Wild Turkeys (Meleagridis Gallopavo Silvestris) from Arkansas. J Wildl Dis. 1990;26:468–472. doi: 10.7589/0090-3558-26.4.468. [DOI] [PubMed] [Google Scholar]
- Jackwood MW, Saif YM. Bordetellosis (Turkey Coryza) In: Saif YM, Fadly AM, Glisson JR, et al., editors. Diseases of Poultry. Blackwell Pub. Professional; Ames, Iowa: 2008. pp. 774–789. [Google Scholar]
- Jackwood MW, McCarter SM, Brown TP. Bordetella avium: An Opportunistic Pathogen in Leghorn Chickens. Avian Dis. 1995;39:360–367. [PubMed] [Google Scholar]
- Loker SB, Temple LM, Preston A. The Bordetella avium BAV1965–1962 Fimbrial Locus is Regulated by Temperature and Produces Fimbriae Involved in Adherence to Turkey Tracheal Tissue. Infect Immun. 2011;79:2423–2429. doi: 10.1128/IAI.01169-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Malik YS, Olsen K, Chander Y, Goyal SM. Antimicrobial Resistance in Bacterial Pathogens Isolated from Turkeys in Minnesota from 1998 to 2002. The International Journal of Applied Research in Veterinary Medicine. 2003;14:506–511. [Google Scholar]
- Malik YS, Chander Y, Gupta SC, Goyal SM. A Retrospective Study on Antimicrobial Resistance in Mannheimia (Pasteurella) haemolytica, Escherichia coli, Salmonella Species, and Bordetella avium from Chickens in Minnesota. Journal of Applied Poultry Research. 2005;14:506–511. [Google Scholar]
- McBride MD, Hird DW, Carpenter TE, Snipes KP, Danaye-Elmi C, Utterback WW. Health Survey of Backyard Poultry and Other Avian Species Located within One Mile of Commercial California Meat-Turkey Flocks. Avian Dis. 1991;35:403–407. [PubMed] [Google Scholar]
- McGinnis S, Madden TL. BLAST: At the Core of a Powerful and Diverse Set of Sequence Analysis Tools. Nucleic Acids Res. 2004;32:W20–W25. doi: 10.1093/nar/gkh435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morris JS, Luginbuhl GH. Conjugative Plasmids in Bordetella avium. Plasmid. 1992;27:251–255. doi: 10.1016/0147-619x(92)90029-a. [DOI] [PubMed] [Google Scholar]
- Mortensen JE, Brumbach A, Shryock TR. Antimicrobial Susceptibility of Bordetella avium and Bordetella bronchiseptica Isolates. Antimicrob Agents Chemother. 1989;33:771–772. doi: 10.1128/aac.33.5.771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raffel TR, Register KB, Marks SA, Temple L. Prevalence of Bordetella avium Infection in Selected Wild and Domesticated Birds in the Eastern USA. J Wildl Dis. 2002;38:40–46. doi: 10.7589/0090-3558-38.1.40. [DOI] [PubMed] [Google Scholar]
- Register KB, Yersin AG. Analytical Verification of a PCR Assay for Identification of Bordetella avium. J Clin Microbiol. 2005;43:5567–5573. doi: 10.1128/JCM.43.11.5567-5573.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sambrook J, Russell DW. Molecular cloning: a laboratory manual. Cold Spring Harbor Laboratory Press; Cold Spring Harbor, N.Y: 2001. [Google Scholar]
- Savelkoul PH, de Groot LE, Boersma C, Livey I, Duggleby CJ, van der Zeijst BA, Gaastra W. Identification of Bordetella avium using the Polymerase Chain Reaction. Microb Pathog. 1993;15:207–215. doi: 10.1006/mpat.1993.1071. [DOI] [PubMed] [Google Scholar]
- Schwarz S, Silley P, Simjee S, Woodford N, van Duijkeren E, Johnson AP, Gaastra W. Editorial: Assessing the Antimicrobial Susceptibility of Bacteria obtained from Animals. J Antimicrob Chemother. 2010;65:601–604. doi: 10.1093/jac/dkq037. [DOI] [PubMed] [Google Scholar]
- Sebaihia M, Preston A, Maskell DJ, Kuzmiak H, Connell TD, King ND, Orndorff PE, Miyamoto DM, Thomson NR, Harris D, Goble A, Lord A, Murphy L, Quail MA, Rutter S, Squares R, Squares S, Woodward J, Parkhill J, Temple LM. Comparison of the Genome Sequence of the Poultry Pathogen Bordetella avium with those of B. bronchiseptica, B. pertussis, and B. parapertussis Reveals Extensive Diversity in Surface Structures Associated with Host Interaction. J Bacteriol. 2006;188:6002–6015. doi: 10.1128/JB.01927-05. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spilker T, Liwienski AA, Lipuma JJ. Identification of Bordetella Spp. in Respiratory Specimens from Individuals with Cystic Fibrosis. Clinical Microbiology and Infection. 2008;14:504–506. doi: 10.1111/j.1469-0691.2008.01968.x. [DOI] [PubMed] [Google Scholar]
- Temple LM, Weiss AA, Walker KE, Barnes HJ, Christensen VL, Miyamoto DM, Shelton CB, Orndorff PE. Bordetella avium Virulence Measured in Vivo and in Vitro. Infect Immun. 1998;66:5244–5251. doi: 10.1128/iai.66.11.5244-5251.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thorsted PB, Macartney DP, Akhtar P, Haines AS, Ali N, Davidson P, Stafford T, Pocklington MJ, Pansegrau W, Wilkins BM, Lanka E, Thomas CM. Complete Sequence of the IncPbeta Plasmid R751: Implications for Evolution and Organisation of the IncP Backbone. J Mol Biol. 1998;282:969–990. doi: 10.1006/jmbi.1998.2060. [DOI] [PubMed] [Google Scholar]
- van den Hurk JV. Characterization of the Structural Proteins of Hemorrhagic Enteritis Virus. Arch Virol. 1992;126:195–213. doi: 10.1007/BF01309695. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.