

Abstract
The Aux/IAA family genes encode short-lived nuclear proteins that function as transcriptional regulators in auxin signal transduction. Aux/IAA genes have been reported to control many processes of plant development. Our recent study showed that down-regulation of SlIAA15 in tomato reduced apical dominance, altered pattern of axillary shoot development, increased lateral root formation and leaves thickness. The SlIAA15 suppressed lines display strong reduction of trichome density, suggesting that SlIAA15 is involved in trichome formation. Here, we reported that SlIAA15-suppressed transgenic lines display increased number of xylem cells compared to wild-type plants. Moreover, the monoterpene content in trichome exudates are significantly reduced in SlIAA15 down-regulated leaves. The results provide the roles of SlIAA15 in production of volatile compounds in leaf exudates and xylem development, clearly indicating that members of the Aux/IAA gene family can play distinct and specific functions.
Keywords: SlIAA15, Aux/IAA genes, Trichome, auxin, xylem
Auxin plays important roles in many aspects of plant growth and development, including cell division, vascular differentiation, apical dominance, lateral/adventitious root formation, fruit set and development, and embryogenesis.1 Aux/IAA family genes are early auxin-induced genes that encode short-lived nuclear proteins. Aux/IAA proteins can act as transcriptional repressors through interaction with auxin response factor (ARF) proteins. Ubiquitin-dependent degradation of the Aux/IAAs activates ARF, which can be either transcriptional activators or repressors of primary/early auxin-responsive genes.2
It has been reported that tomato AUX/IAA genes control many processes of plant development. In potato, downregulation of StIAA2 resulted in altered phenotypes including petiole hyponasty, curvature of developing leaf primordia and increased plant height.3 In tomato, genome-wide analysis identifies 26 putative AUX/IAA genes.4 Suppression of SlIAA3 produced auxin and ethylene-related developmental defects, such as reduced apical dominance and exaggerated hook in etiolated seedlings.5 Downregulation of SlIAA9 altered apical dominance and leaf architecture, and resulted in parthenocarpic fruit.6,7 SlIAA15 was first cloned from tomato fruit using Aux/IAAs family-specific degenerate primers.8 Expression studies revealed SlIAA15 transcripts can be detected in all the tissues including roots, stems, leaves, seedlings, flowers and fruit. In a recent publication, downregulationg of SlIAA15 in tomato results in pleiotropic phenotypes including reduced apical dominance, altered pattern of axillary shoot development, increased lateral root formation, and increased leaves thickness.9 Our results indicate that SlIAA15 has a common role in maintaining growth and developmental processes.
In this work, the effect of downregulation of SlIAA15 on anatomical characterization of stem was investigated in embedded sections. The SlIAA15-suppressed transgenic lines display increased number of xylem cells compared with wild-type plants (WT) (Fig. 1). This result indicates that SlIAA15 is involved in stem xylem development in tomato. It has been reported that auxin is a key signal for stem xylem development.10 Moyle et al.11 found hybrid aspen five Aux/IAA genes (PttIAA1, 2, 3, 4, and 8) are upregulated in xylem. In Arabidopsis, eight of the Aux/IAA genes (IAA19, IAA28, IAA22, IAA2, IAA12, IAA8, IAA13, and IAA26) were upregulated in xylem.12 The data describe the roles of SlIAA15 gene in xylem development and provide new evidence supporting that xylem formation requires a functional auxin signaling pathway.

Figure 1. The increased number of xylem cells in SlIAA15-suppressed transgenic plants. Samples (fifth internode from top) from ten-weeks-old WT and transgenic plants were embedded in LR white resin. Semi-thin sections were stained with toluidine blue and viewed with the light microscope. WT, wild-type plants. AS, SlIAA15 downregulated plants. C: Cambia, X: Xylem. Bars, 40 μm.
Tomato trichomes are a variety of multicellular glandular and non-glandular structures which act as both physical and chemical barriers against biotic and abiotic stresses.13 Tomato trichomes are categorized as types I to VII, with types I, IV, VI and VII being glandular while types II, III and V are non-glandular.14 In our recent study, the SlIAA15-suppressed transgenic plants display a remarkable reduction in numbers of type I, V and VI trichomes, suggesting that auxin-dependent transcriptional regulation is involved in trichome formation.9 Since trichome glands are an important site of volatile compounds production, in this study the content of volatile terpenes in trichome exudates was analyzed by gas chromatography-mass spectrometry (GC-MS) following extraction with 3 mL of tert-butyl methyl ether (MTBE) for 5 min. Important differences were found between WT and SlIAA15-suppressed transgenic plants in the content of five compounds identified as monoterpenes α-Terpinene, β-Phellandrene, γ-Elemene, β-Caryophyllene, and α-Humulene (Fig. 2). Notably, these compounds were shown previously to be major volatile components of tomato leaf aroma.15,16 These data indicate that monoterpene content in trichome exudates are significantly reduced in SlIAA15 downregulated leaves compared with wild-type leaves.
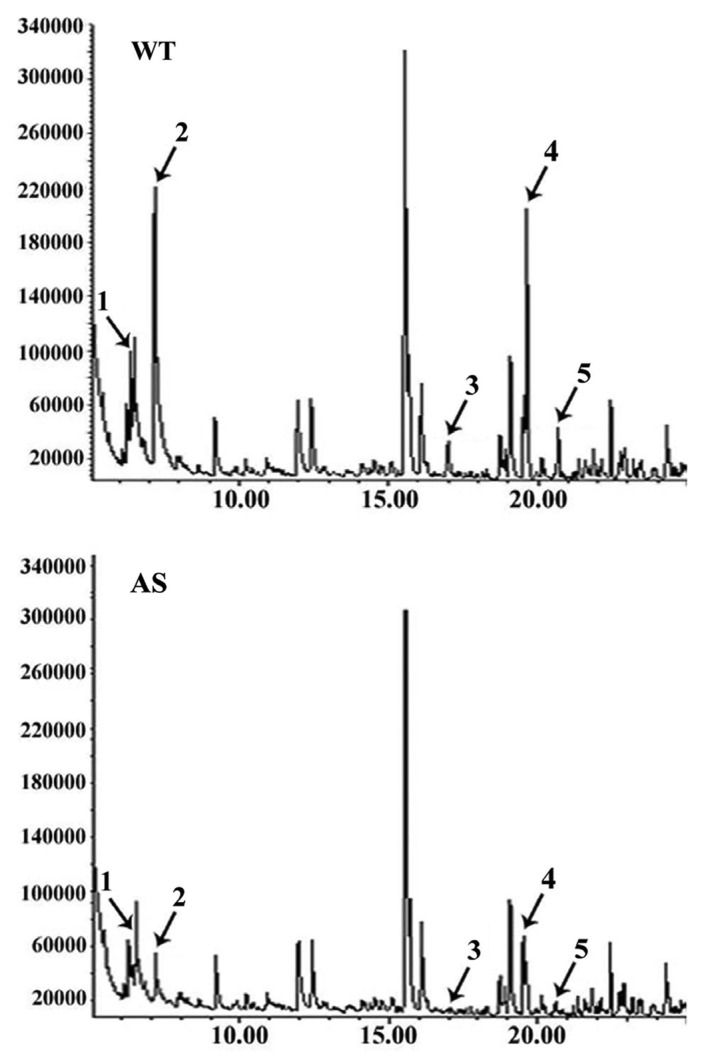
Figure 2. The reduced contents of volatile compounds in leaf exudates of transgenic plants. Gas chromatography-mass spectrometry (GC-MS) was used to measure the young-leaf exudates from ten-weeks-old WT and transgenic plants. The exudates were obtained by brief extraction with tert-methyl butyl ether buffer (MTBE). WT, wild-type plants. AS, SlIAA15 downregulated plants. Numbers 1–5 represent the different monoterpenes respectively: 1, α-Terpinene, 2, β-Phellandrene, 3, γ-Elemene, 4, β-Caryophyllene, 5, α-Humulene.
Li et al.17 reported that a tomato homolog of CORONATINE-INSENSITIVE1 (COI1), an F-box protein required for JA-signaled processes, positively regulates both glandular trichome development and the production of monoterpenes stored in this structure. In this study, the reduction in monoterpene content of SlIAA15-suppressed transgenic plants is consistent with our finding that downregulation of SlIAA15 expression significantly reduced the density of type I and VI glandular trichomes.9 We speculate that the reduction of glandular trichomes’ density may lead to the low exudation of monoterpene from tomato leaves. This data provide the functions of SlIAA15 in production of volatile compounds in leaf exudates, clearly demonstrating that members of the Aux/IAA genes can play distinct and specific functions.
Acknowledgments
This work was supported by a Project from the National Natural Science Foundation of China (30700544, 30471214 and 31071798), and the Natural Science Foundation of Chongqing (2011BA1024).
Footnotes
Previously published online: www.landesbioscience.com/journals/psb/article/20723
References
- 1. Friml J. Auxin transport - shaping the plant. Curr Opin Plant Biol. 2003;6:7–12. doi: 10.1016/S1369526602000031. [DOI] [PubMed] [Google Scholar]
- 2.Gray WM, Kepinski S, Rouse D, Leyser O, Estelle M. Auxin regulates SCF(TIR1)-dependent degradation of AUX/IAA proteins. Nature. 2001;414:271–6. doi: 10.1038/35104500. [DOI] [PubMed] [Google Scholar]
- 3.Kloosterman B, Visser RGF, Bachem CWB. Isolation and characterization of a novel potato Auxin/Indole-3-Acetic Acid family member (StIAA2) that is involved in petiole hyponasty and shoot morphogenesis. Plant Physiol Biochem. 2006;44:766–75. doi: 10.1016/j.plaphy.2006.10.026. [DOI] [PubMed] [Google Scholar]
- 4.Wu J, Peng Z, Liu S, He Y, Cheng L, Kong F, et al. Genome-wide analysis of Aux/IAA gene family in Solanaceae species using tomato as a model. Mol Genet Genomics. 2012;287:295–11. doi: 10.1007/s00438-012-0675-y. [DOI] [PubMed] [Google Scholar]
- 5.Chaabouni S, Jones B, Delalande C, Wang H, Li Z, Mila I, et al. Sl-IAA3, a tomato Aux/IAA at the crossroads of auxin and ethylene signalling involved in differential growth. J Exp Bot. 2009;60:1349–62. doi: 10.1093/jxb/erp009. 60/4/1349 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Wang H, Jones B, Li ZG, Frasse P, Delalande C, Regad F, et al. The tomato Aux/IAA transcription factor IAA9 is involved in fruit development and leaf morphogenesis. Plant Cell. 2005;17:2676–92. doi: 10.1105/tpc.105.033415. 17/10/2676 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Wang H, Schauer N, Usadel B, Frasse P, Zouine M, Hernould M, et al. Regulatory features underlying pollination-dependent and -independent tomato fruit set revealed by transcript and primary metabolite profiling. Plant Cell. 2009;21:1428–52. doi: 10.1105/tpc.108.060830. 21/5/1428 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Jones B, Frasse P, Olmos E, Zegzouti H, Li ZG, Latché A, et al. Down-regulation of DR12, an auxin-response-factor homolog, in the tomato results in a pleiotropic phenotype including dark green and blotchy ripening fruit. Plant J. 2002;32:603–13. doi: 10.1046/j.1365-313X.2002.01450.x. [DOI] [PubMed] [Google Scholar]
- 9.Deng W, Yang YW, Ren ZX, Audran-Delalande C, Mila I, Wang XY, et al. The tomato SlIAA15 is involved in trichome formation and axillary shoot development. New Phytol. 2012;194:379–90. doi: 10.1111/j.1469-8137.2012.04053.x. [DOI] [PubMed] [Google Scholar]
- 10.Sundberg B, Ugglar C, Tuominen H. Cambial growth and auxin gradient. In: Savidge RA, Barnett JR, Napier R, eds. Cell and molecular biology of wood formation. Oxford, UK: Bios Scienti®c Publishers Ltd 2000; slu/v11n1/11n1a14
- 11.Moyle R, Schrader J, Stenberg A, Olsson O, Saxena S, Sandberg G, et al. Environmental and auxin regulation of wood formation involves members of the Aux/IAA gene family in hybrid aspen. Plant J. 2002;31:675–85. doi: 10.1046/j.1365-313X.2002.01386.x. [DOI] [PubMed] [Google Scholar]
- 12. Oh S, Park S, Han KH. Transcriptional regulation of secondary growth in Arabidopsis thaliana. J Exp Bot. 2003;54:2709–22. doi: 10.1093/jxb/erg304. [DOI] [PubMed] [Google Scholar]
- 13.Kennedy GG. Tomato, pests, parasitoids, and predators: tritrophic interactions involving the genus Lycopersicon. Annu Rev Entomol. 2003;48:51–72. doi: 10.1146/annurev.ento.48.091801.112733. abs/10.1146/annurev.ento.48.091801.112733 [DOI] [PubMed] [Google Scholar]
- 14.Luckwill LC. The Genus Lycopersicon: Historical, biological, and taxonomic survey of the wild and cultivated tomatoes. Aberdeen University Press, Aberdeen, Scotland 1943; pp 44 [Google Scholar]
- 15. Andersson BA, Holman RT, Lennart L, Stenhagen G. Capillary gas chromatograms of leaf volatiles. A possible aid to breeders for pest and disease resistance. J Agric Food Chem. 1980;28:985–9. doi: 10.1021/jf60231a018. [DOI] [Google Scholar]
- 16.Buttery RG, Ling LC, Light DM. Tomato leaf volatile aroma components. J Agric Food Chem. 1987;35:1039–43. doi: 10.1021/jf00078a043. [DOI] [Google Scholar]
- 17.Li L, Zhao Y, McCaig BC, Wingerd BA, Wang J, Whalon ME, et al. The tomato homolog of CORONATINE-INSENSITIVE1 is required for the maternal control of seed maturation, jasmonate-signaled defense responses, and glandular trichome development. Plant Cell. 2004;16:126–43. doi: 10.1105/tpc.017954. [DOI] [PMC free article] [PubMed] [Google Scholar]