

Abstract
Proteomic analysis of the hemocytic proteins of Penaeus monodon (Pm) has previously shown that alpha-2-macroglobulin (A2M) was among the proteins that showed substantially altered expression levels upon Vibrio harveyi infection. Therefore, in this study its potentially important role in the response of shrimp to bacterial infection was further characterized. The yeast two-hybrid system revealed that the receptor binding domain of PmA2M interacted with the carboxyl-terminus of one or both of the transglutaminase type II isoforms, which are key enzymes involved in the shrimp clotting system. In accord with this, PmA2M was found to be localized on the extracellular blood clots and to colocalize with clottable proteins. RNA interference (RNAi)-mediated knockdown of A2M transcript levels reduced the PmA2M transcript levels (∼94%) and significantly reduced the bacterial seizing ability of the clotting system, resulting in an up to 3.3-fold higher number of V. harveyi that systemically disseminated into the circulatory system at 5 min post-infection before subsequent clearance by the immune system. Furthermore, an appearance of PmA2M depleted clots in the presence of V. harveyi strikingly demonstrated fibrinolysis zones surrounding the bacteria. This study provides the first evidence of the vital role of PmA2M in enhancing bacterial sequestration by protecting blood clots against fibrinolysis.
Introduction
Infection of Vibrio harveyi, one of the major species categorized in the genus Vibrio [1], in penaeid shrimp causes necrosis of the basal tissues followed by the systemic dissemination of the bacteria [2]. The characteristic aspects of V. harveyi diseased penaeid shrimp are a glowing in the dark (luminous vibriosis) and the occurrence of developmental disorders of the epidermal tissues obstructing the digestive tract (bolitas negrican). Although the pathogenic mechanisms of V. harveyi and its associated phages have not been clearly elucidated, extracellular products, such as proteases, phospholipase and hemolysin, are regarded as the major virulent factors produced by the bacteria [3]. Indeed, proteases secreted from diverse pathogenic organisms have been recognized as a significant factor that plays a variety of crucial roles in facilitating the invasion and dissemination of the pathogen into the host, and in evasion of the immune system and alteration of housekeeping functions [4]. In addition to many cysteine proteases, considered as the major toxins, three alkaline metal ion sensitive proteases have also been characterized from V. harveyi, and these exotoxins exert lethal pathogenesis to penaeid shrimp, and in particular to the black tiger shrimp, Penaeus monodon [5]–[10].
Consequentially, as expected, specific protease inhibitors are a vital element of the host to defend and neutralize against invasion by protease-secreting pathogens so as to diminish the illness and death among the hosts. From these, a number of protease inhibitors that are ubiquitously found in microorganisms and multicellular organisms have been found and they serve essential roles in controlling both the endogenous and exogenous proteases [11]. These protease inhibitors can be divided into two main classes; (i) active site inhibitors, which directly interact with the active site of the endopeptidase, and (ii) the alpha-2-macroglobulins (A2M), which inhibit the protease activity via a conformation change to engage the targeted protease inside the A2M molecule conformation and so result in an irreversible inhibition called the physical entrapment mechanism [12], [13].
In invertebrates, A2M is a broad range proteinase inhibitor and a highly abundant plasma protein [14] that is crucially involved in many immune responses, such as phagocytosis in the hard tick, Ixodes ricinus [15], and in the mosquito, Anopheles gambiae [16], the prophenoloxidase activating system in the crayfish, Pacifastacus leniusculus [17], and the blood clotting system in P. leniusculus [18] and the horseshoe crab, Limulus polyphemus [19]. In penaeid shrimp, the cloning, biochemical characterization, binding studies and expression at both the transcription and translation levels of A2M have been reported [20]–[28]. The protein expression profile in the major immune tissues of V. harveyi infected P. monodon revealed that PmA2M was the most strongly altered protein in hemocytes [28] and lymphoid organ [29], following V. harveyi infection. However, to date there is no direct evidence concerning the biological role(s) of A2M in response to severe pathogens, and especially in response of shrimp to V. harveyi.
This research, therefore, focused on which immune pathways A2M is involved in and how A2M functions against V. harveyi infections in P. monodon. The yeast two-hybrid (Y2H) technology was firstly conducted to identify interacting partners of the receptor binding domain (RBD) of PmA2M. Since the RBD is exposed upon protease inactivation, it should be noted that this study reveals only one part of the roles of PmA2M in the immune response against V. harveyi. The immunofluorescent technique was later performed in order to localize PmA2M in different hemocyte types and the hemolymph clots of shrimp. Furthermore, based on the fact that the pathogen entrapment within hemolymph clots acts as an early immune mechanism in other invertebrates [30], the biological roles of PmA2M in sequestration of V. harveyi within the shrimp blood clots were also investigated using RNA interference (RNAi).
Materials and Methods
Experimental animals and V. harveyi 639 cultivation
Healthy black tiger shrimp, P. monodon, each weighing approximately either 15 grams or 1–3 g body weight were used in the analysis of PmA2M protein expression levels as well as the localization of PmA2M protein and in RNAi based gene silencing experiments, respectively. Shrimp were acclimatized in artificially aerated sea water (15 ppt salinity) for a week before experimental use. The shrimp pathogenic V. harveyi strain 639 was freshly prepared, as previously reported [31].
Ethics statement
The experimental shrimp were obtained from the Broodstock and Larval Development Research Center at Walailuk University, Nakornsrithammarat Province, Thailand, where is not privately-owned or protected. No specific permits were required for the described field studies and for these locations. We confirm that these field studies did not involve endangered or protected species.
The manual of Standard Operating Procedures (SOPs) for the Ethical Use of Live Aquatic Animals approved by the Museum Victoria Animal Ethics Committee on 12th November 2010 was applied to our studies, since there have been no official recommendations for the experimental use of invertebrates for scientific purposes in Thailand. The transporting, raising and experimental processing of the animals were strictly performed as described in the manual of SOPs. Also, the humane killing at the end of experiments was carried out by first completely anaesthetizing the experimental shrimp and then placing them into a −20°C freezer, and all efforts were taken in order to minimize animal suffering, as pointed out in the SOPs.
Screening for the PmA2M interacting protein(s) from shrimp hemocytes using a Y2H assay
The RBD of PmA2M was amplified from the hemocyte cDNA of unchallenged shrimp using the gene specific forward and reverse primers (Table 1), with 5′ flanking NdeI and BamHI sequences (underlined), respectively, and the Advantage®2 Polymerase Mix (Clontech). The purified amplicon was then cloned into the bait vector, pGBKT7 (Clontech). The resulting recombinant plasmid (pGBKT7-RBD-A2M) was introduced into the yeast Saccharomyces cerevisiae strain Gold (Clontech) using the Yeastmaker™ Yeast Transformation System 2 (Clontech). The cDNA library of V. harveyi-infected shrimp hemocytes used to screen for the interaction with the RBD of A2M was constructed using the Make Your Own “Mate & Plate™” Library system (Clontech). In brief, the hemocytes from five individual shrimp previously infected with 106 CFU of V. harveyi were harvested at 6 or 48 h post-infection. The total RNA was extracted and cDNA was synthesized as described previously [32]. The library was constructed according to the manufacturer's protocol. Yeast mating and selection of positive transformants were carried out according to the Matchmaker™ Gold Y2H System user manual (Clontech). The prey plasmids were rescued as previously reported [33] and later confirmed the interaction by the co-transformation assay, as described in the instruction manual. The prey plasmid which truly interacts with the RBD of A2M was subjected to DNA sequencing. The obtained sequences were searched against the GenBank database using the BlastX program.
Table 1. DNA sequences of the primers used in experiments.
| Primers | Primer sequences (5′-3′) |
| Yeast two-hybrid | |
| PmA2M-F | 5′-GCCGATCATATGCTTGTCCACGAATTCACC-3′ |
| PmA2M-R | 5′-ATCGAGGATCCTTTTGCTGCCGTCCAC-3′ |
| Protein production | |
| PmA2M-F | 5′-CGCGGCCATATGCTTGTCCACGAATTCACC-3′ |
| PmA2M-R | 5′-CATATGCTCGAGTTTGCTGCCGTCCACCTCGTA-3′ |
| Gene silencing | |
| PmA2M-F | 5′-CCATGGAGGGGCAGGGATGC-3′ |
| PmA2M-R | 5′-ATCGCACCCTTCGAGGTACG-3′ |
| T7PmA2M-F | 5′-GGATCCTAATACGACTCACTATAGGCCATGGAGGGGCAGGGATGC-3′ |
| T7PmA2M-R | 5′-GGATCCTAATACGACTCACTATAGGATCGCACCCTTCGAGGTACG-3′ |
| Semiquantitative RT-PCR analysis | |
| PmA2M-F | 5′-CCATGGAGGGGCAGGGATGC-3′ |
| PmA2M-R | 5′-ATCGCACCCTTCGAGGTACG-3′ |
| β-actin-F | 5′-GCTTGCTGATCCACATCTGCT-3′ |
| β-actin-R | 5′-ATCACCATCGGCAACGAGA-3′ |
Production of recombinant PmRBD-A2M and anti-PmRBD-A2M Abs
The forward and reverse primers (Table 1), with 5′ flanking NdeI and XhoI sequences (underlined), respectively, were designed to amplify the RBD of A2M from the pGBKT7-RBD-A2M plasmid using the Advantage®2 Polymerase Mix (Clontech). The purified PCR product was digested with NdeI and XhoI, and then ligated to the pET-22b (+) expression vector (Novagen) cut with the same restriction enzymes. The obtained recombinant plasmid (pET-RBD-A2M) was then transformed into E. coli strain BL21(DE3). The transformed E. coli was cultured and the protein expression was induced by addition of 1 mM IPTG (Isopropyl β-D-1-thiogalactopyranoside). At 4 h after IPTG induction, the cells were harvested, resuspended in phosphate buffered saline pH 7.4 (PBS) and then sonicated to break the cells. The inclusion bodies were collected, solubilized with 50 mM sodium carbonate pH 10.3, and dialyzed against 20 mM sodium phosphate pH 8.0 overnight at 4°C to refold the protein. The protein was purified using a Ni-Sepharose™ 6 Fast Flow affinity column (GE Healthcare), in accordance with the manufacture's manual. SDS-PAGE and western blotting using the anti-HIS antibody (GE Healthcare) were performed to analyze the purity of the recombinant protein. The polyclonal Abs against the recombinant Pm-RBD-A2M were commercially raised in rabbits and mice at the Biomedical Technology Research Unit, Chiang Mai University, Thailand.
Localization of PmA2M protein in shrimp hemocytes by immunofluorescence
To examine the PmA2M protein localized in hemocytes, hemolymph was collected from three individual shrimp and then pooled. Hemocytes were isolated by centrifugation (800× g; 10 min; 4°C), with the hemocyte pellet being immediately fixed in 4% (w/v) paraformaldehyde in PBS and kept at 4°C until use.
Hemocytes (1×105 cells/ml) were attached onto polylysine-coated microscope slides by cytospin centrifugation. Based on immunofluorescence staining procedure previously described [34], PmA2M was probed using purified rabbit anti-Pm-RBD-A2M polyclonal IgG Ab diluted 1∶500 in PBSF (PBS plus 1% (v/v) FBS) followed by a 1∶500 dilution of Alexa Fluor® 488 goat anti-rabbit IgG Ab (Invitrogen) in PBSF. Hemocyte nuclei were finally stained. The microscope slides were kept in the dark at 4°C until they were observed under a confocal fluorescence microscope.
Activation of shrimp blood clotting and preparation of a clot matrix
Two hundred µl of shrimp hemolymph was collected from a ventral haemoceol using a sterile syringe with a 23G needle containing 50 µl of anticoagulant (10% (w/v) sodium citrate). To activate the blood coagulation, 5 µl of 1 M CaCl2 was added to the plasma, gently mixed and incubated at ambient temperature for 5 min then stopping the clotting reaction by the addition of 25 µl of 0.5 M EDTA. The clot matrix was fixed onto a polylysine-coated microscope slide using the hanging drop technique, as previously pointed out [35].
Co-localization of PmA2M and clottable proteins (CPs) on the blood clot by in vitro immunohistochemical staining
After preparing clot matrices on the polylysine-coated slides, the samples were air-dried at 37°C in an oven. For immunohistochemical staining [34], the samples were incubated with the primary Abs (purified mouse anti-Pm-RBD-A2M polyclonal IgG Ab at 1∶150 dilution in PBSF and the purified rabbit anti-CP polyclonal IgG Ab at 1∶250 dilution in PBSF), simultaneously introduced to the samples, and subsequently probed with 1∶500 dilution of Alexa Fluor®568 goat anti-mouse (Invitrogen) and Alexa Fluor®647 goat anti-rabbit Abs (Invitrogen) in PBSF, respectively. The samples were stored in the dark at 4°C until observed under a confocal fluorescence microscope.
RNAi-mediated PmA2M gene silencing and semi-quantitative determination of PmA2M transcript levels
Clone number HC-N-N01-1034 from contig 711 of the P. monodon EST database (http://pmonodon.biotec.or.th) was found to be similar to the RBD of PmA2M and so was used as the template for PmA2M dsRNA production. Two sets of primers specific to the RBD of PmA2M were designed as shown in Table 1, plus the same primers but 5′ flanked by the T7 promoter sequence (underlined) to form T7PmA2M-F and T7PmA2M-R. Otherwise, dsRNA of GFP, the negative control, was prepared as previously described [36]. The two PCR products were separately amplified using those primer pairs with the following conditions; 94°C for 5 min followed by 30 cycles of 94°C for 30 s, 57°C for 30 s and 72°C for 30 s, and then a final 72°C for 10 min.
Experimental shrimp, each of approximately 1–3 g body weight, were divided into three groups. The first control group was injected with 50 µl of sterile saline solution (SSS) alone. The second and third groups (control and knockdown groups) were injected with 10 µg of either GFP-dsRNA or A2M-dsRNA in 50 µl of SSS, respectively. The injection was repeated as above after 24 h, and the hemolymph was collected from each shrimp in all three groups after a further 24 h (48 h after the first injection). The hemocytes were isolated by centrifugation. Total RNA was extracted and used for cDNA synthesis [32]. Using semi-quantitative RT-PCR and 1.5% (w/v) agarose-TBE gel electrophoresis, the PmA2M gene knockdown level was assessed by comparison between the transcript levels of the PmA2M gene relative to that of the internal housekeeping control gene, β-actin (Forward and reverse primers, shown in Table 1). The band densitometry software (GeneTools) was carried out to quantify the band intensity.
Clot observation under the scanning electron microscope (SEM)
Scanning electron microscopy (SEM) was used to observe the effect of A2M-dsRNA-mediated knockdown on the physiological changes of the clot in the presence of V. harveyi. As previously reported [35], [37], the bacterial suspension in PBS was prepared and 1.5 µl of the suspension was added into the plasma from the GFP-dsRNA and A2M-dsRNA treated groups. Blood coagulation was activated as described above and the hanging drop technique [35] was then performed.
To prepare the samples for SEM analysis, the surface of the partially clotted plasma was attached upside down to a new small glass slide avoiding making contact between the two slides. The samples were prepared as described previously [34] and examined under a scanning electron microscope.
Invading bacteria count in the hemolymph of the PmA2M depleted (dsRNA-mediated knockdown) shrimp
1.5×105 CFU/ml of freshly prepared V. harveyi suspended in 50 µl of SSS was intramuscularly injected into the GFP-dsRNA and A2M-dsRNA knockdown groups. Three independent individuals were evaluated per group, and for each shrimp 100 µl of hemolymph was collected using a sterile syringe without any anticoagulant solution- at 5, 15 or 30 min after injection. Ten µl from each collected hemolymph sample was immediately dotted on TCBS (thiosulfate–citrate–bile–sucrose) agar plates, a Vibrio species selective medium, with five replicates performed per sample. After air-drying the dotted hemolymph, the plates were then incubated upside down at 30°C overnight. The number of invading bacteria was counted by a total plate count method, as previously pointed out [38].
Results
Carboxyl terminus of transglutaminase type II (PmSTG II) identified as an interacting partner of the receptor binding domain (RBD) of PmA2M
Previously, proteomic analysis revealed the significant down-regulation of shrimp A2M protein levels in response to V. harveyi infection [28], [29]. It is evident that the RBD of A2M plays necessary roles in subsequent clearance of the protease-reacted A2M from the blood circulation. Thus, the protein partners that likely interact with the RBD of PmA2M were identified using the Y2H system to further investigate the potential role of A2M in shrimp immunity. A DNA fragment encoding the RBD of PmA2M was initially cloned into pGBKT7. Before performing the Y2H assay, the recombinant PmA2M in the pGBKT7 backbone was tested for autoactivation of yeast reporter genes and toxicity to host yeast cells. No activation of the reporter genes or differences in yeast cell sizes were observed between the control and experimental groups (data not shown).
After the screening, six independent positive clones (SB1–4, PB1 and PB2) were discovered. The prey plasmids from these clones were isolated and subjected to DNA sequencing. Four of the positive clones (SB1–4) were identified by sequence identity as being the carboxyl-terminus of transglutaminase type II, PmSTG II (accession number AAV49005.1). The other two clones, PB1 and PB2, were identified by sequence similarity to be the homologs of the voltage-dependent anion-selective channel isoform 1 from Tribolium castaneum (accession number XP_967480.1) and the 40S ribosomal protein S20 from Rimicaris exoculata (accession number ACR78697.1), respectively.
These six purified prey plasmids were then ascertained if they authentically interact with the bait by the co-transformation assay. Like the positive control, all the transformants containing the SB1–4 and PB1–2 prey and bait plasmids seemed to grow well and appeared as blue colonies on both selective media, as a result of reporter gene activations. Correspondingly, no growth was observed in the SB1–4 prey and empty bait plasmids co-transformed yeast cells on the high stringency media, similar to the negative control (Fig. 1). On the other hand, the PB1 and PB2 prey and empty bait plasmids containing transformants grew on the high stringency media, suggesting that the PB1 and PB2 clones were false positives (Fig. 1). Altogether, the results support that the carboxyl-terminal region of the PmSTG II protein(s) interact(s) with the RBD of PmA2M.
Figure 1. Y2H based co-transformation assay to confirm positive prey proteins.

The RBD of A2M was used as the bait protein. The prey plasmids encoding for the carboxyl-terminus of the P. monodon transglutaminase type II (PmSTG II) (SB1, SB2, SB3 and SB4), and the homologs of the T. castaneum voltage-dependent anion-selective channel isoform 1 (PB1) and R. exoculata 40S ribosomal protein S20 (PB2), were co-transformed into the S. cerevisiae strain Gold with either an empty bait plasmid or a bait plasmid containing the RBD of A2M. The yeast transformants grown on selective media were restreaked onto DDO/X media (double drop-out media lacking leucine and tryptophan but supplemented with X-α-Gal) and QDO/X/A (quadruple drop-out media lacking adenine, histidine, leucine and tryptophan but supplemented with X-α-Gal and Aureobasidin A, a high stringency medium) and evaluated for growth. The interaction between murine p53 and the SV40 large T antigen served as a positive control, whereas that between the murine p53 and lamin was used as the negative control.
PmA2M is mainly contained in semigranular hemocytes
According to our proteomic study concerning responsive proteins in hemocytes of V. harveyi infected P. monodon [28] suggesting that hemocytes are one of the principle sources of PmA2M involving in bacterial defense, the localization of PmA2M was then determined in three main types of shrimp hemocytes (hyaline, semigranular, and granular hemocytes). Immunofluorescent staining of the PmA2M was conducted to assess the amount of PmA2M protein in hemocytes. The hemocyte nuclei were visualized by staining with TO-PRO-3 iodine (red color in Fig. 2), while anti-rPmA2M Ab and Alexa Fluor® 488 conjugated second Ab were applied to detect the subcellular PmA2M protein (green color in Fig. 2).
Figure 2. Confocal microscope analysis of A2M protein content in shrimp hemocytes using immunohistochemical staining.

Three different types of the hemocytes consisting of hyaline, semigranular, and granular hemocytes are indicated and pooled hemocytes from three normal shrimp (1×105 cells/ml) are shown. The A2M proteins and hemocytic nuclei are visualized as green (Alexa Fluor® 488) and red (TO-PRO-3 Iodine) colors, respectively (colocalization of the nucleus and A2M is not seen in the merged image). The scale bar corresponds to 5 µm. Images shown are representative of at least 3 such fields of view per sample.
Fig. 2 demonstrated the aspect of shrimp hemocytes which are mainly classified into three types; hyaline (agranular), semigranular (small-granular), and granular (large-granular) hemocytes, based on the existence or relative size of granules [39]. The strongest immunostaining for PmA2M was noticed in the semigranular hemocytes, and was contained within the secretory granules of those cells. There were also a few immunostaining granules within the granular hemocytes, but positive PmA2M staining hyaline cells were rarely observed (Fig. 2). The result indicated that the semigranular hemocyte is a major component among other types which might play crucial roles in PmA2M regulation against the bacterial infection.
The incorporation of PmA2M in shrimp clots
Since the Y2H screening revealed that PmA2M was able to interact with PmSTG II, a key enzyme that plays a crucial role in the blood coagulation system [40], [41], it is possible that PmA2M is one of PmSTG II's interacting partners and can be associated with the clotting system. Immunofluorescence using confocal microscopy was then conducted to examine the presence of PmA2M in the shrimp clots. CPs, known as a backbone of the fibril clot, were investigated for colocalization with PmA2M to demonstrate the location of hemolymph clot matrix.
Using the hanging drop technique with CaCl2 activated blood clotting, PmA2M and CPs were detected by anti-rPmA2M mouse IgG and anti-rCP rabbit IgG Abs, respectively, followed by Alexa Fluor® 568 conjugated goat anti-mouse and Alexa Fluor® 647 conjugated goat anti-rabbit IgG Abs, respectively. As can be seen in the bright field, in the activated clotting condition, plasma clot strands developed allowing the association of many hemocytes onto the clot fibrils (Fig. 3A; red arrows). CPs were found to be specifically dispersed on the plasma clotting fibril strands (blue color in Fig. 3B). Interestingly, PmA2M was also localized on the clot strands and the associated hemocyte cells (green color in Fig. 3C), such that the CPs and PmA2M were colocalized over much of their distribution in the extracellular fibrils (cyan color in Fig. 3D). However, the two components are not totally colocalized, with significant areas of both fibrils and hemocytes being stained for either CP and not PmA2M, or PmA2M and not CP. Nevertheless, overall PmA2M was clearly incorporated onto the extracellular blood clot matrix.
Figure 3. Co-localization of A2M and CPs in hemocytes and extracellular fibrils of blood clots.

The immunocytology and confocal microscopy were performed. Anti-rPmA2M and anti-rCP Abs were visualized by Alexa Fluor® 568 and Alexa Fluor® 647 conjugated secondary Abs and show up as green and blue colors for A2M and CP, respectively, or cyan for colocalized in the merged field. Red arrowheads demonstrate fibril clots and the scale bar corresponds to 50 µm. Images shown are representative of at least 3 such fields of view per sample.
Partially depleted PmA2M transcript levels result in the escape of V. harveyi from the clot
To elucidate the biological function of PmA2M, dsRNA interference mediated gene silencing was performed in order to suppress PmA2M transcript (and so assumed protein) expression levels in P. monodon shrimp. Ten µg of dsRNA specific to the RBD of PmA2M (A2M-dsRNA) in 50 µl of SSS was injected into each experimental shrimp twice with a 24 h interval between them. Control shrimp (two groups) were likewise injected but with either SSS alone or SSS containing dsRNA against GFP (GFP-dsRNA). A significant (∼94%) reduction in the PmA2M transcript level was observed in the hemocytes from the dsRNA-A2M knockdown shrimp at 24 h after the second injection, as determined by semi-quantitative RT-PCR, but no change was noted in the dsRNA-GFP or SSS control groups (Fig. 4). The PmA2M transcript level recovered to normal levels by day 5 (data not shown).
Figure 4. Determination of the A2M transcript in the shrimp hemocytes after in vivo RNAi-mediated gene silencing.
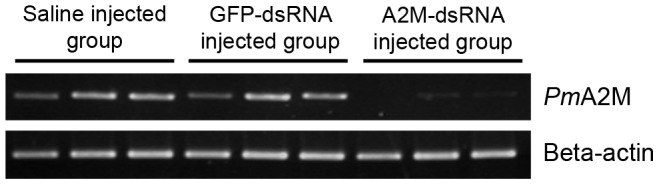
Three groups of experimental shrimp (1–3 g body weight) were injected with SSS alone or with SSS containing either GFP-dsRNA or A2M-dsRNA in a final concentration of 10 µg/g body weight. Each group contains three individuals and each lane represents an individual shrimp. The mRNA expression level of A2M was assessed by semi-quantitative RT-PCR using A2M gene specific primers and normalized relative to that for the β-actin gene, which served as the internal control.
The effect of this significantly reduced PmA2M transcript levels in the A2M gene silenced shrimp on the ability to entrap V. harveyi in the plasma clot was evaluated. To this end, hemolymph obtained from GFP-dsRNA control and A2M-dsRNA knockdown shrimp was mixed with V. harveyi and clotting was induced. Samples were then prepared, CO2 critical dried, and examined by SEM. When V. harveyi was present during the clotting, the bacteria became entrapped in the extracellular blood clots of the control group (Fig. 5A). There were some fibrils attached to the bacteria, presumably immobilizing them in the clot, and the bacteria remained in close association with the clot matrix (Fig. 5A′). Strikingly, the blood clot matrix observed in the A2M-dsRNA treated shrimp showed that the remaining bacteria were surrounded by fibrinolysis zones (Fig. 5B′), indicating the susceptibility of A2M-depleted clots to proteolysis, presumably from proteases released from the bacterial cells [42]. Moreover, some of the invading bacteria had apparently escaped from the clot matrix, as can be seen in some regions (Fig. 5B; arrows), suggesting that PmA2M bound to the clots prevented bacterial escape by inhibiting the bacterial protease-mediated fibrinolysis.
Figure 5. Observation of GFP-dsRNA and A2M-dsRNA treated clots in the presence of V. harveyi.

SEM was used at an accelerating voltage at 20 kV and a magnification of 2,000× (A, B) or 20,000× (A′, B′) were applied. Blood clots were formed by the hanging drop technique in the presence of V. harveyi from the hemolymph of (A, A′) GFP-dsRNA or (B, B′) A2M-dsRNA knockdown shrimp. Arrowheads demonstrate the fibrinolysis zones where the bacteria have escaped from the clot. Scale bars correspond to 10 µm in the upper sub-figures and 1 µm in the lower sub-figures. Images shown are representative of at least 3 such fields of view per sample.
Systemic dissemination of V. harveyi in the circulating hemolymph of PmA2M gene silenced shrimp
The number of disseminated V. harveyi in the shrimp hemolymph was counted in order to support the above finding of the apparent escape of bacteria from clots in the A2M knockdown shrimp. The hemolymph from GFP-dsRNA and A2M-dsRNA treated shrimp was collected at 5, 15 and 30 min after injection of live V. harveyi. At 5 min, the number of bacteria in the A2M-dsRNA treated group was significantly (3.3-fold) higher than that in the control group. The invading bacteria were, nevertheless, cleared out from the circulatory system in the control group by 30 min, and reduced some four-fold to <2000 CFU/ml in the A2M depleted shrimp by 15 min (Fig. 6). Additionally, there were a few of the viable bacteria detected in hemolymph of both groups at 6 h and 24 h (no significant difference) and these viable bacterial numbers kept slightly decreasing over the time (data not shown). Since the differences in the bacterial number were only seen at the most early time points of infection (5 min), it suggests that the bacterial entrapment within the plasma clots would represent the early immune mechanisms in response to the bacterial infection.
Figure 6. The evaluation of invading V. harveyi levels in the hemolymph.

Hemolymph was collected from GFP and A2M knockdown shrimp at 5, 15 and 30 min after injection with freshly prepared V. harveyi (1.5×105 CFU/ml) and then plated onto Vibrio sp. selective agar plates (TCBS) to examine the number of viable bacteria. The amount of circulating bacteria in hemolymph of the GFP knockdown group was used as control. Five trials were done in each group. The data, as CFU/ml, are shown as the mean ± S.D. * represents a significant difference between the two shrimp groups (p<0.05; Independent Sample T-test).
The results demonstrated the release of V. harveyi cells from the A2M-depleted blood clots into the circulatory system, whilst in the control shrimp, most bacteria were not able to enter the circulation system and so were presumably entrapped in the clot. This supported the important function of A2M in shrimp innate immunity by preventing the escape of bacteria from blood clots.
Discussion
In order to better understand the innate immunity of crustaceans, shrimp immune responses to several pathogens have been elucidated from a variety of proteomic, genomic, molecular genetical and biochemical approaches. From the immune responsive proteins identified and studied to date by these methods, A2M expression levels were found to be highly altered after pathogen infection in several shrimp species including P. monodon. Even though the changes in A2M expression levels in the major immune tissues were typically observed when challenged with viruses, such as white spot syndrome and yellow head viruses [43]–[46], a substantial change in both the transcript and protein levels of A2M was noticed after bacterial challenge, and particularly after V. harveyi infection [25], [28], [29]. Moreover, Gollas-Galvan et al. [20] recently revealed that the A2M from P. vannamei exhibits a broad range of protease inhibitory activities to various proteases, including members of the elastase and aminopeptidase families that include known pathogen-aiding bacterial proteases. Hence, changes in shrimp A2M expression levels might be a host defense mechanism against bacterial proteases, possibly to neutralize the severity of the bacterial infection or to prevent the bacterial dissemination throughout the shrimp hemoceol. The biological pathway to inhibit the action of bacterial proteases used by A2M in shrimp, nevertheless, remains elusive.
To unveil the defense action of PmA2M against bacteria, potential molecules that interact with PmA2M was investigated using the Y2H screening assay. The RBD of PmA2M was found to interact with the carboxyl-terminal domain of PmSTG II. Currently, at least three members of the STG enzyme family have been identified in P. monodon, which are PmSTG I (AY074924) [47] and two members of PmSTG II (AY771615; AF469484) [41]. However, the amino acid sequences of the C-terminus region of both known PmSTG II proteins are congruent and therefore further experiments are required to investigate which PmSTG II isoform is interacting with A2M, or if both isoforms are involved. Since both PmSTG IIs were previously determined to be key enzymes involved in the blood coagulation system [40], [41], the binding between PmA2M and PmSTG II suggests a possible role for PmA2M related to the hemolymph clotting. In accordance with the protein polymerization activity of STGs, which catalyses an acyl transfer reaction between lysine residues and glutamine residues, four conserved lysine residues (K1324, K1387, K1421 and K1425) were discovered in the RBD of PmA2M [22]. In addition, the A2M of the crayfish, P. leniusculus, can act as a substrate of transglutaminase and exposes free lysine and glutamine residues as does the A2M of the American horseshoe crab, Limulus polyphemus, which decorates on the clot in the presence of Ca2+ [18], [42], [48]. These data suggest that PmA2M might also be a substrate of the transglutaminase enzyme for cross-linking to other proteins serving roles in the plasma coagulation. Although there have been a number of reports on mammalian A2M revealing that the RBD can bind to the low-density lipoprotein receptor-related protein A2M receptor (LRP-A2MR) mediating the clearance of inhibitor–protease complexes from the blood circulation and the LRP receptor-associated protein (RAP) mediating a signal regulation [49], they have no report in invertebrates including shrimp. It can be proposed that the KPTVK motif which is responsible for receptor binding [50] is not present in the RBD of PmA2M; therefore, shrimp might possess other pathways for the protease clearance.
In invertebrates, the hemolymph coagulation process commonly occurs in plasma with the cooperation of a variety of hemocyte derived molecules but without the direct involvement of the hemocytes [51]. Blood coagulation in shrimp has been proposed to be similar to that in the crayfish, P. leniusculus [40], where under an external stimulus, such as LPS, STG is released from the hyaline hemocytes into the plasma to initiate the blood clotting by cross-linking the CPs, which are abundant proteins in the hemolymph that are mainly produced in the gill, central nervous system and the lymphoid organ [52], [53]. The result of this report here demonstrated that PmA2M is contained in semigranular hemocytes (Fig. 2) and possibly releases into the hemolymph via degranulation. Moreover, an increase of A2M protein was observed in the plasma upon V. harveyi infection (data not shown). We assume that the secreted A2M participates in several host immune defenses occurring in plasma including the hemolymph clotting process. This might be a reason why the A2M protein content in shrimp hemocytes was substantially decreased upon the V. harveyi infection, observed in our proteomic study [28]. As previously studied in the horseshoe crab, Limulus polyphemus [54], the exocytosis of granules packed in the blood cells was activated by endotoxin to subsequently release A2M into the plasma. This is the first report in shrimp that demonstrates the direct involvement of A2M in blood coagulation and that A2M is incorporated into the clots where it potentially serves as one of the major backbones of the clot matrix along with the CPs (Fig. 3).
To gain more insight into the biological role of PmA2M in the blood clotting, we observed the effect of PmA2M gene silencing on the entrapment of bacteria in the clots by SEM. With the partial RNAi-mediated knockdown of PmA2M gene transcript expression, the PmA2M depleted plasma clots were notably ineffective at sequestering V. harveyi, resulting in the higher systemic dissemination of the bacteria into the circulatory system (Figs. 5 and 6). Nevertheless, the invading bacteria were rapidly cleared out from the blood circulation in both control and A2M gene silenced shrimp within half an hour. It is not so surprised because an array of host immune responses actively functions against invading pathogens and is doubtlessly required for the pathogen clearance. This is the first evidence suggesting that PmA2M plays a critical role in the blood clotting system by preventing fibrinolysis of clots from V. harveyi proteases, and so in restraining the protease-mediated bacterial escape.
The physical entrapment mechanism of A2M has been proposed to be comprised of three main steps. Firstly, in the protease recognition step, a protease directly attacks some susceptible peptide bonds in the bait region followed by the compaction of the A2M molecule to entrap the protease. Secondly, in the capture step, the thiol group is cleaved and generates a reactive glutamyl group which can covalently link to the protease. Lastly, as a result of this conformation change, the RBD of A2M is exposed on the outside of the molecule and the domain is recognized by its receptor for subsequent entry to the endocytotic clearance pathway [49], [55]. Thus, it seems that only a protease reacted form of A2M, where the RBD is uncovered, will be biologically processed whilst its unreacted form is still free in both the plasma and hemocytes. The fibril bound A2M in the American horseshoe crab had already functioned and appeared as a reacted form [48], whereas, PmA2M on the shrimp clots is able to inhibit the proteolysis activity of, presumably, proteases secreted from the invading bacteria [42] (Figs. 5A and 5A′), implying that the exposure of the RBD of PmA2M occurs when linked on the clots and before entrapping any proteases. It should be noted that the reorganization of the molecular structure of PmA2M is probably distinct between the different isoforms and so this requires further investigation for clarification. Altogether, our findings clearly indicate a vital role for PmA2M in the blood coagulation system, the most efficient primitive immune response, against pathogen attack.
Acknowledgments
We would like to thank Professor Kenneth Söderhäll for providing the Ab against crayfish CP as well as Professor Takashi Aoki and Professor Ikuo Hirono for offering facilities used in the construction of V. harveyi-infected P. monodon hemocyte cDNA library. We thank Dr. Sanong Ekgasit for kindly assist on SEM analysis of blood clots and Dr. Robert Douglas John Butcher for English language correction of this manuscript.
Funding Statement
This research was financially supported by research grants from the Thailand National Center for Genetic Engineering and Biotechnology (BIOTEC), the National Research University Project of CHE and the Ratchadaphiseksomphot Endowment Fund (FW643A). VC received a student fellowship from the Royal Golden Jubilee Ph.D. program under the Thailand Research Fund (TRF) and KS received additional financial support from the TRF, the Office of the Higher Education Commission (Grant No. MRG5280119) and the Faculty of Science, Chulalongkorn University (Grant No. RESA1B1-11). The exchange of scientists (KS) under the JSPS-NRCT Program is also acknowledged, as is the provision of research equipment from the Thai Government Stimulus Package 2 (TKK2555) under the Project for the Establishment of a Comprehensive Center for Innovative Food, Health Products and Agriculture. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Dorsch M, Lane D, Stackebrandt E (1992) Towards a phylogeny of the genus Vibrio based on 16S rRNA sequences. Int J Syst Bacteriol 42: 58–63. [DOI] [PubMed] [Google Scholar]
- 2. Jiravanichpaisal P, Miyazaki T, Limsuwan C (1994) Histopathology, biochemistry, and pathogenicity of Vibrio harveyi infecting black tiger prawn Penaeus monodon . J Aquat Anim Health 6: 27–35. [Google Scholar]
- 3. Austin B, Zhang XH (2006) Vibrio harveyi: a significant pathogen of marine vertebrates and invertebrates. Lett Appl Microbiol 43: 119–124. [DOI] [PubMed] [Google Scholar]
- 4. Armstrong PB (2006) Proteases and protease inhibitors: a balance of activities in host-pathogen interaction. Immunobiology 211: 263–281. [DOI] [PubMed] [Google Scholar]
- 5. Fukasawa S, Nakamura K, Miyahira M, Kurata M (1988) Some properties of two proteinases from a luminous bacterium, Vibrio harveyi strain FLN-108. Agric Biol Chem 52: 3009–3014. [Google Scholar]
- 6. Fukasawa S, Nakamura K, Kamii A, Ohyama Y, Osumi M (1988) Purification and properties of a proteinase from a marine luminous bacterium, Vibrio harveyi strain FLA-11. Agric Biol Chem 52: 435–441. [Google Scholar]
- 7. Liu PC, Lee KK, Tu CC, Chen SN (1997) Purification and characterization of a cysteine protease produced by pathogenic luminous Vibrio harveyi . Curr Microbiol 35: 32–39. [DOI] [PubMed] [Google Scholar]
- 8. Liu PC, Lee KK (1999) Cysteine protease is a major exotoxin of pathogenic luminous Vibrio harveyi in the tiger prawn, Penaeus monodon . Lett Appl Microbiol 28: 428–430. [DOI] [PubMed] [Google Scholar]
- 9. Montero AB, Austin B (1999) Characterization of extracellular products from an isolate of Vibrio harveyi recovered from diseased post-larval Penaeus vannamei (Bonne). J Fish Dis 22: 377–386. [Google Scholar]
- 10. Harris LJ, Owens L (1999) Production of exotoxins by two luminous Vibrio harveyi strains known to be primary pathogens of Penaeus monodon larvae. Dis Aquat Organ 38: 11–22. [Google Scholar]
- 11. Laskowski M Jr, Kato I (1980) Protein inhibitors of proteinases. Ann Rev Biochem 49: 593–626. [DOI] [PubMed] [Google Scholar]
- 12. Armstrong PB, Quigley JP (1999) α2-macroglobulin: an evolutionarily conserved arm of the innate immune system. Dev Comp Immunol 23: 375–390. [DOI] [PubMed] [Google Scholar]
- 13. Kanost MR (1999) Serine proteinase inhibitors in arthropod immunity. Dev Comp Immunol 23: 291–301. [DOI] [PubMed] [Google Scholar]
- 14. Armstrong PB, Melchior R, Quigley JP (1996) Humoral immunity in long-lived arthropods. J Insect Physiol 42: 53–64. [Google Scholar]
- 15. Buresova V, Hajdusek O, Franta Z, Sojka D, Kopacek P (2009) IrAM-An α2-macroglobulin from the hard tick Ixodes ricinus: Characterization and function in phagocytosis of a potential pathogen Chryseobacterium indologenes . Dev Comp Immunol 33: 489–498. [DOI] [PubMed] [Google Scholar]
- 16. Levashina EA, Moita LF, Blandin S, Vriend G, Lagueux M, et al. (2001) Conserved role of a complement-like protein in phagocytosis revealed by dsRNA knockout in cultured cells of the mosquito, Anopheles gambiae . Cell 104: 709–718. [DOI] [PubMed] [Google Scholar]
- 17. Aspán A, Hall M, Söderhäll K (1990) The effect of endogeneous proteinase inhibitors on the prophenoloxidase activating enzyme, a serine proteinase from crayfish haemocytes. Insect Biochem 20: 485–492. [Google Scholar]
- 18. Hall M, Söderhäll K (1994) Crayfish α-macroglobulin as a substrate for transglutaminases. Comp Biochem Phys B 108: 65–72. [Google Scholar]
- 19. Armstrong PB, Levin J, Quigley JP (1984) Role of endogenous proteinase inhibitors in the regulation of the blood clotting system of the horseshoe crab, Limulus polyphemus . Thromb Haemostas 52: 117–120. [PubMed] [Google Scholar]
- 20. Gollas-Galván T, Sotelo-Mundo RR, Yepiz-Plascencia G, Vargas-Requena C, Vargas-Albores F (2003) Purification and characterization of α2-macroglobulin from the white shrimp (Penaeus vannamei). Comp Biochem Phys C 134: 431–438. [DOI] [PubMed] [Google Scholar]
- 21. Rattanachai A, Hirono I, Ohira T, Takahashi Y, Aoki T (2004) Molecular cloning and expression analysis of α2-macroglobulin in the kuruma shrimp, Marsupenaeus japonicus . Fish Shellfish Immunol 16: 599–611. [DOI] [PubMed] [Google Scholar]
- 22. Lin YC, Vaseeharan B, Ko CF, Chiou TT, Chen JC (2007) Molecular cloning and characterisation of a proteinase inhibitor, alpha 2-macroglobulin (α2-M) from the haemocytes of tiger shrimp Penaeus monodon . Mol Immunol 44: 1065–1074. [DOI] [PubMed] [Google Scholar]
- 23. Lin YC, Vaseeharan B, Chen JC (2008) Molecular cloning and phylogenetic analysis on α2-macroglobulin (α2-M) of white shrimp Litopenaeus vannamei . Dev Comp Immunol 32: 317–329. [DOI] [PubMed] [Google Scholar]
- 24. Ho PY, Cheng CH, Cheng W (2009) Identification and cloning of the α2-macroglobulin of giant freshwater prawn Macrobrachium rosenbergii and its expression in relation with the molt stage and bacteria injection. Fish Shellfish Immunol 26: 459–466. [DOI] [PubMed] [Google Scholar]
- 25. Ma H, Wang B, Zhang J, Li F, Xiang J (2010) Multiple forms of alpha-2 macroglobulin in shrimp Fenneropenaeus chinesis and their transcriptional response to WSSV or Vibrio pathogen infection. Dev Comp Immunol 34: 677–684. [DOI] [PubMed] [Google Scholar]
- 26. Tonganunt M, Phongdara A, Chotigeat W, Fujise K (2005) Identification and characterization of syntenin binding protein in the black tiger shrimp Penaeus monodon . J Biotechnol 120: 135–145. [DOI] [PubMed] [Google Scholar]
- 27. Chotigeat W, Deachamag P, Phongdara A (2007) Identification of a protein binding to the phagocytosis activating protein (PAP) in immunized black tiger shrimp. Aquaculture 271: 112–120. [Google Scholar]
- 28. Somboonwiwat K, Chaikeeratisak V, Wang HC, Lo CF, Tassanakajon A (2010) Proteomic analysis of differentially expressed proteins in Penaeus monodon hemocytes after Vibrio harveyi infection. Proteome Sci 8: 39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Chaikeeratisak V, Somboonwiwat K, Wang HC, Lo CF, Tassanakajon A (2012) Proteomic analysis of differentially expressed proteins in the lymphoid organ of Vibrio harveyi-infected Penaeus monodon . Mol Biol Rep 39: 6367–6377. [DOI] [PubMed] [Google Scholar]
- 30. Isakova V, Armstrong PB (2003) Imprisonment in a death-row cell: The fates of microbes entrapped in the Limulus blood clot. Biol Bull 205: 203–204. [DOI] [PubMed] [Google Scholar]
- 31. Ponprateep S, Somboonwiwat K, Tassanakajon A (2009) Recombinant anti- lipopolysaccharide factor isoform 3 and the prevention of vibriosis in the black tiger shrimp, Penaeus monodon . Aquaculture 289: 219–224. [Google Scholar]
- 32. Amparyup P, Sutthangkul J, Charoensapsri W, Tassanakajon A (2012) A pattern recognition protein binds to lipopolysaccharide and β-1,3-glucan and activates the shrimp prophenoloxidase system. J Biol Chem 287: 10060–10069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Robzyk K, Kassir Y (1992) A simple and highly efficient procedure for rescuing autonomous plasmids from yeast. Nucleic Acids Res 20: 3790. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Prapavorarat A, Vatanavicharn T, Söderhäll K, Tassanakajon A (2010) A novel viral responsive protein is involved in hemocyte homeostasis in the black tiger shrimp, Penaeus monodon . J Biol Chem 285: 21467–21477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Bidla G, Lindgren M, Theopold U, Dushay MS (2005) Hemolymph coagulation and phenoloxidase in Drosophila larvae. Dev Comp Immunol 29: 669–679. [DOI] [PubMed] [Google Scholar]
- 36. Amparyup P, Charoensapsri W, Tassanakajon A (2009) Two prophenoloxidases are important for the survival of Vibrio harveyi challenged shrimp Penaeus monodon . Dev Comp Immunol 33: 247–256. [DOI] [PubMed] [Google Scholar]
- 37. Wang Z, Wilhelmsson C, Hyrsl P, Loof TG, Dobes P, et al. (2010) Pathogen entrapment by transglutaminase—A conserved early innate immune mechanism. PLoS Pathog 6: e1000763 doi:10.1371/journal.ppat.1000763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Liu H, Jiravanichpaisal P, Cerenius L, Lee BL, Söderhäll I, et al. (2007) Phenoloxidase is an important component of the defense against Aeromonas hydrophila infection in a crustacean, Pacifastacus leniusculus . J Biol Chem 282: 33593–33598. [DOI] [PubMed] [Google Scholar]
- 39. Martin GG, Graves BL (1985) Fine structure and classification of shrimp hemocytes. J Morphol 185: 339–348. [DOI] [PubMed] [Google Scholar]
- 40. Chen MY, Hu KY, Huang CC, Song YL (2005) More than one type of transglutaminase in invertebrates? A second type of transglutaminase is involved in shrimp coagulation. Dev Comp Immunol 29: 1003–1016. [DOI] [PubMed] [Google Scholar]
- 41. Yeh MS, Kao LR, Huang CJ, Tsai IH (2006) Biochemical characterization and cloning of transglutaminases responsible for hemolymph clotting in Penaeus monodon and Marsupenaeus japonicus . BBA-Proteins Proteom 1764: 1167–1178. [DOI] [PubMed] [Google Scholar]
- 42. Theopold U, Schmidt O, Söderhäll K, Dushay MS (2004) Coagulation in arthropods: defence, wound closure and healing. Trends Immunol 25: 289–294. [DOI] [PubMed] [Google Scholar]
- 43. Pongsomboon S, Tang S, Boonda S, Aoki T, Hirono I, et al. (2011) A cDNA microarray approach for analyzing transcriptional changes in Penaeus monodon after infection by pathogens. Fish Shellfish Immunol 30: 439–446. [DOI] [PubMed] [Google Scholar]
- 44. Rojtinnakorn J, Hirono I, Itami T, Takahashi Y, Aoki T (2002) Gene expression in haemocytes of kuruma prawn, Penaeus japonicus, in response to infection with WSSV by EST approach. Fish Shellfish Immunol 13: 69–83. [DOI] [PubMed] [Google Scholar]
- 45. Wang HC, Wang HC, Leu JH, Kou GH, Wang AHJ, et al. (2007) Protein expression profiling of the shrimp cellular response to white spot syndrome virus infection. Dev Comp Immunol 31: 672–686. [DOI] [PubMed] [Google Scholar]
- 46. Bourchookarn A, Havanapan P, Thongboonkerd V, Krittanai C (2008) Proteomic analysis of altered proteins in lymphoid organ of yellow head virus infected Penaeus monodon . BBA-Proteins Proteom 1784: 504–511. [DOI] [PubMed] [Google Scholar]
- 47. Huang CC, Sritunyalucksana K, Söderhäll K, Song YL (2004) Molecular cloning and characterization of tiger shrimp (Penaeus monodon) transglutaminase. Dev Comp Immunol 28: 279–294. [DOI] [PubMed] [Google Scholar]
- 48. Armstrong PB, Armstrong MT (2003) The decorated clot: Binding of agents of the innate immune system to the fibrils of the Limulus blood clot. Biol Bull 205: 201–203. [DOI] [PubMed] [Google Scholar]
- 49. Armstrong PB (2010) Role of α2-macroglobulin in the immune responses of invertebrates. Inv Surv J 7: 165–180. [Google Scholar]
- 50. Lin M, Sutherland DR, Horsfall W, Totty N, Yeo E, et al. (2002) Cell surface antigen CD109 is a novel member of the α2 macroglobulin/C3, C4, C5 family of thioester-containing proteins. Blood 99: 1683–1691. [DOI] [PubMed] [Google Scholar]
- 51. Cerenius L, Söderhäll K (2011) Coagulation in invertebrates. J Innate Immun 3: 3–8. [DOI] [PubMed] [Google Scholar]
- 52.Vargas-Albores F, Hernández-López J, Gollas-Galván T, Montaño-Pérez K, Jiménez-Vega F, et al.. (1998) Activation of shrimp cellular defence functions by microbial products. In: Flegel TW, editors. Advances in shrimp biotechnology. Bangkok: National Center for Genetic Engineering and Biotechnology. pp. 161–166.
- 53. Yeh MS, Huang CJ, Cheng JH, Tsai IH (2007) Tissue-specific expression and regulation of the haemolymph clottable protein of tiger shrimp (Penaeus monodon). Fish Shellfish Immunol 23: 272–279. [DOI] [PubMed] [Google Scholar]
- 54. Armstrong PB, Quigley JP (1985) Proteinase inhibitory activity released from the horseshoe crab blood cell during exocytosis. Biochim Biophys Acta 827: 453–459. [DOI] [PubMed] [Google Scholar]
- 55. Armstrong PB (2001) The contribution of proteinase inhibitors to immune defense. Trends Immunol 22: 47–52. [DOI] [PubMed] [Google Scholar]