

Abstract
Identification of cardiomyocyte nuclei has been challenging in tissue sections as most strategies rely only on cytoplasmic marker proteins1. Rare events in cardiac myocytes such as proliferation and apoptosis require an accurate identification of cardiac myocyte nuclei to analyze cellular renewal in homeostasis and in pathological conditions2. Here, we provide a method to isolate cardiomyocyte nuclei from post mortem tissue by density sedimentation and immunolabeling with antibodies against pericentriolar material 1 (PCM-1) and subsequent flow cytometry sorting. This strategy allows a high throughput analysis and isolation with the advantage of working equally well on fresh tissue and frozen archival material. This makes it possible to study material already collected in biobanks. This technique is applicable and tested in a wide range of species and suitable for multiple downstream applications such as carbon-14 dating3, cell-cycle analysis4, visualization of thymidine analogues (e.g. BrdU and IdU)4, transcriptome and epigenetic analysis.
Keywords: Medicine, Issue 65, Stem Cell Biology, Cardiology, Physiology, Tissue Engineering, cardiomyocyte, post mortem, nuclei isolation, flow cytometry, pericentriolar material 1, PCM-1
Protocol
1. Isolation of the Cardiac Nuclei
Coat ultracentrifuge tubes (Beckman Centrifuge Tubes #363664) with 10 ml of 1%BSA/PBS coating solution. Cap the tubes and let them rotate for 30 min in a tube rotator. Remove the coating solution and let the centrifuge tubes air dry (one tube per mouse heart is required for the analysis of single mouse hearts, alternately up to 5 mouse hearts or 1 g of heart tissue from a different species (e.g. human) can be processed in one tube).
All following steps should be performed on ice. Dissect the left ventricle from fresh or snap-frozen mouse heart with a scalpel. Note, this protocol is optimized for mouse heart but can also be adapted to rat or human heart. Alternatively, use up to 1 g of heart tissue from a different species.
Trim the specimen into small cubicles with a scalpel.
Transfer the tissue pieces into a 50 ml Falcon tube filled with 15 ml of lysis buffer.
Homogenize the heart tissue with a T-25 Ultra-Turrax probe homogenizer (IKA) at 24,000 rpm for 10 sec.
Dilute the homogenate with equal volume of lysis buffer to 30 ml.
Use a glass douncer (40 ml) to further homogenize the tissue and free the nuclei. Perform eight strokes with a large clearance pestle.
Pass the crude nuclei isolate through a 100 μm and 70 μm nylon mesh cell strainer (BD Biosciences), consecutively.
Spin down the crude nuclei isolate in a refrigerated centrifuge (4 °C) at 700 x g for 10 min.
Remove the supernatant carefully by inverting the tubes and wipe the inside of the tube with paper towel. Be careful not to disturb the nuclei pellet.
Dissolve the crude nuclei isolate in 5 ml of sucrose buffer by pipetting the solution several times up and down. Add a further 25 ml of sucrose buffer to the dissolved pellet.
Add 10 ml of freshly prepared sucrose buffer to the coated ultracentrifuge tube (see step 1.1).
Carefully overlay the added 10 ml of sucrose buffer with the resuspended nuclei pellet dissolved in sucrose buffer from step 1.9.
Balance the centrifuge tubes before placing them into a JS13.1 free swinging rotor and place the rotor into a high-speed centrifuge (Beckman Avanti S-25).
Spin the nuclei sample at 13,000 x g at 4 °C for 60 min.
When the spin has completed, remove the tubes carefully from the rotor and discard the supernatant by inverting the tubes and wiping the remaining debris from the inside of the tubes with paper towel.
Dissolve the nuclei pellet in 1 ml of nuclei storage buffer (NSB plus buffer). Note: NSB plus contains 1.5 mM spermine as a DNA stabilizer.
Proceed with step 2.1, immunostaining of cardiomyocyte nuclei.
2. Immunostaining for Flow Cytometry
Prepare the negative control for immunostaining. Take an aliquot of 20 μl out of the nuclei sample and add 980 μl of NSB plus buffer.
Add anti-pericentriolar material 1 antibody (rabbit anti-PCM-1, Atlas Antibodies) to the nuclei sample in a dilution of 1:500 to immunolabel cardiomyocyte nuclei. Add the isotype antibody in the same dilution as the anti-PCM-1 antibody to the negative control, prepared in step 2.1.
Incubate negative control and sample tube at 4 °C overnight.
Wash negative control and sample at least once with NSB plus buffer (spin down tubes in a refrigerated centrifuge (4 °C) at 700 x g for 10 min. Discard supernatant and dissolve the nuclei pellet in 1 ml of NSB plus buffer).
Add anti-rabbit fluorescent secondary antibody (FITC or APC) to negative control and sample tube in a dilution of 1:1000.
Incubate negative control and sample tube at 4 °C for 1 h.
Wash negative control and sample at least once with NSB plus buffer (spin down tubes in a refrigerated centrifuge (4 °C) at 700 x g for 10 min. Discard supernatant and dissolve the nuclei pellet in 1 ml of NSB plus buffer).
Proceed with the flow cytometric analysis and sorting.
3. Flow Cytometry
Coat nuclei collection tubes (Falcon 15 ml) with 1%BSA/PBS solution before you start flow cytometry sorting as described in step 1.1.
Filter the sample and the negative control through a 30 μm cell strainer and load first the negative control to the flow cytometer (BD Influx). Define the first and second gate to define nuclei and singlets (single nuclei), based on forward scatter (FSC), forward scatter pulse width (FS pulse width) and side scatter (SSC) (Fig. 2a and b). Adding a DNA stain (DRAQ5 (1:500)) to the sample can help to identify the nuclei population initially.
Load the immunolabeled sample and define the third gate to isolate cardiomyocyte nuclei (PCM-1-positve) from non-cardiomyocyte nuclei (PCM-1-negative). Start the sorting (Fig. 2c and d).
Optional: In order to analyze the nuclear DNA content (ploidy) and to perform cell cycle analysis add an appropriate DNA stain to the nuclei (e.g. Hoechst 33342 or DRAQ5) (Fig. 2e).
After flow cytometry sorting, place the nuclei on ice and re-analyze to determine the sorting purity (Fig. 3a and b).
Spin down the sorted nuclei in the collection tubes at 1500 x g in a refrigerated centrifuge for 15 min.
Dissolve the nuclei pellet in a buffer compatible with the downstream application.
4. Representative Results
Nuclei morphology and integrity can be assessed by DNA stains and visualized by microscopy (Fig. 1). Successful PCM-1 labeling can be assessed by epifluorescence microscopy and by flow cytometry (Fig. 1 and Fig. 2c and d). PCM-1-positive and negative populations should be well separated from each other (Fig. 2c and d). In murine left ventricle about 30% of all nuclei should be cardiomyocyte nuclei (Fig. 2d). Sorting purity can be assessed by re-analyzing the sorted nuclei (Fig. 3a and b). Both nuclei populations should have a sorting purity exceeding 95%.
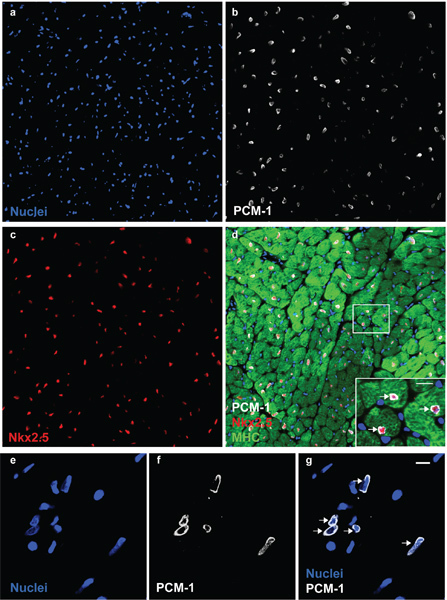 Figure 1. PCM-1 identifies cardiomyocyte nuclei. Cardiac nuclei (a) are stained with antibodies to PCM-1 (b) and to Nkx2.5 (c) in an adult mouse heart. (d) PCM-1-labeled nuclei are surrounded by cardiomyocyte cytoplasm (myosin heavy chain (MHC)) and express the transcription factor Nkx2.5, documenting the accurate identification of cardiomyocyte nuclei by PCM-1 staining (scale bars 20 μm and 10 μm (d, inset)). (e) Cardiac nuclei isolates visualized with the DNA stain DRAQ5. (f and g) Cardiomyocyte nuclei are labeled with antibodies against PCM-1 (scale bar 10 μm). Note, the epinuclear staining pattern of PCM-1 in myocyte nuclei in tissue section and in isolated nuclei (arrows).
Figure 1. PCM-1 identifies cardiomyocyte nuclei. Cardiac nuclei (a) are stained with antibodies to PCM-1 (b) and to Nkx2.5 (c) in an adult mouse heart. (d) PCM-1-labeled nuclei are surrounded by cardiomyocyte cytoplasm (myosin heavy chain (MHC)) and express the transcription factor Nkx2.5, documenting the accurate identification of cardiomyocyte nuclei by PCM-1 staining (scale bars 20 μm and 10 μm (d, inset)). (e) Cardiac nuclei isolates visualized with the DNA stain DRAQ5. (f and g) Cardiomyocyte nuclei are labeled with antibodies against PCM-1 (scale bar 10 μm). Note, the epinuclear staining pattern of PCM-1 in myocyte nuclei in tissue section and in isolated nuclei (arrows).
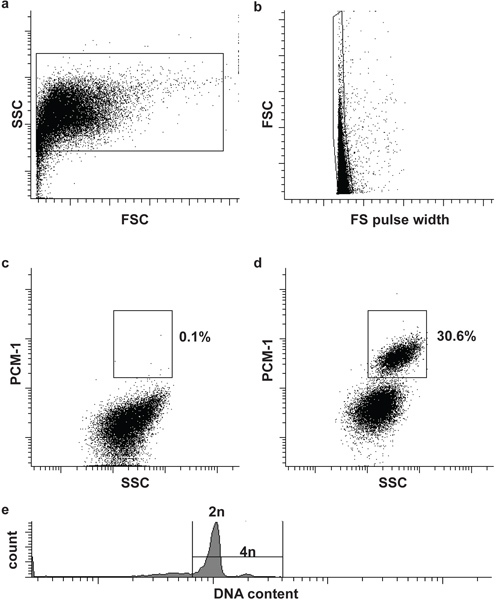 Figure 2. Flow cytometric sorting of cardiomyocyte nuclei. (a) Cardiac nuclei are identified by forward scatter (FSC) and side scatter (SSC). (b) A second gate identifies single nuclei by FSC and FS pulse width5. (c, d) Fluorescent gating allows the separation of cardiomyocyte nuclei (PCM-1-positive) and non-cardiomyocyte (PCM-1-negative) nuclei from heart tissue. (e) Mouse cardiomyocyte are mostly (>80%) diploid (2n), only a small subset is tetraploid (4n)6. Note, human cardiomyocytes contain a higher frequency of polyploidy nuclei (>2n)7,8.
Figure 2. Flow cytometric sorting of cardiomyocyte nuclei. (a) Cardiac nuclei are identified by forward scatter (FSC) and side scatter (SSC). (b) A second gate identifies single nuclei by FSC and FS pulse width5. (c, d) Fluorescent gating allows the separation of cardiomyocyte nuclei (PCM-1-positive) and non-cardiomyocyte (PCM-1-negative) nuclei from heart tissue. (e) Mouse cardiomyocyte are mostly (>80%) diploid (2n), only a small subset is tetraploid (4n)6. Note, human cardiomyocytes contain a higher frequency of polyploidy nuclei (>2n)7,8.
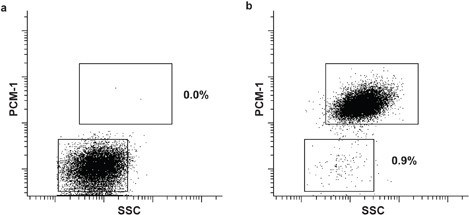 Figure 3. Purity analysis of sorted cardiomyocyte and non-cardiomyocyte nuclei. Re-analysis of sorted non-cardiomyocyte (a) and cardiomyocyte nuclei (b). Both populations show a sorting purity exceeding 99%.
Figure 3. Purity analysis of sorted cardiomyocyte and non-cardiomyocyte nuclei. Re-analysis of sorted non-cardiomyocyte (a) and cardiomyocyte nuclei (b). Both populations show a sorting purity exceeding 99%.
Discussion
Accurate identification of cardiomyocyte nuclei is crucial for the analysis of regenerative processes in the myocardium2,3. Conventional techniques to isolate cardiomyocytes from fresh tissue are mainly based on enzymatic digestion of extracellular matrix proteins and the subsequent purification from interstitial cells by low speed centrifugation. Further purification of living cardiomyocytes from embryonic stem cells (ESC) can be performed by immunolabeling with surface markers such as SIRPA9 or mitochondrial dyes10, fixed cardiomyocytes can be identified by cytoplasmic markers such as myosin heavy chain (MHC) or nuclear proteins such as GATA4 or Nkx2.5. However, the expression of Nkx2.5 and GATA4 is not restricted to mature cardiomyocytes, but can be also detected in progenitor cells during development and after cardiac infarction11,12. Genetic strategies to identify cardiomyocytes show the highest specificity, but cannot be applied in large animal models or humans1. The limitations of the existing strategies have led to controversial conclusions relying on the quantification of rare events such as proliferation, apoptosis and chimerism 1,13. Thus, there is a need to establish an accurate strategy to identify and purify mature differentiated cardiomyocyte nuclei.
Here, we describe a purification technique based on pericentriolar material 1 (PCM-1). Previously, we strengthened the validity of our nuclei isolation by showing that three independent nuclear markers, PCM-1, cardiac troponin T and I, identify the same cardiomyocyte population.2,3 The purified population showed a high enrichment of cardiomyocyte specific gene products such as cardiac troponins, myosin heavy chain protein and the transcription factor Nkx2.5 and GATA42,3.
The current protocol, based on PCM-1, has been successfully applied for the analysis of human and mouse cardiomyocyte turnover2,4 as well as for the analysis of the cardiomyocyte transcriptional profile2. We anticipate that the presented strategy can also be adapted to the isolation of cardiomyocytes from other species. PCM-1, a centrosome protein, re-localizes to the nuclear membrane where it accumulates and forms an insoluble matrix only in differentiated myocytes 2,14 (Fig. 1). Flow cytometry allows the rapid unbiased analysis of a high number of cells, which is required to quantify rare events in cardiomyocytes such as apoptosis and cell cycle re-entry15. Furthermore, nucleotide analogues (e.g. BrdU) based proliferation assays can be readily applied using our presented technique4. Our technique has the advantage that the DNA content of single cardiomyocyte nuclei (nuclear ploidy) can be analyzed in multinuclear cardiomyocytes16,17, which provides important information to discriminate between cardiomyocyte duplication and polyploidisation6-8. However, due to the nature of our strategy, a characterization of cardiomyocyte multinucleation is not possible.
Further applications of the presented method are studies focused on epigenetic changes of chromatin, nucleoproteins, protein-DNA/RNA interactions and on the nuclear transcriptome 18,19.
Because of the stability of nuclei in freeze-thawed tissue, the presented nuclei isolation strategy can also be applied to frozen tissues, making it possible to utilize archived material from biobanks.
Disclosures
No conflicts of interest declared.
Acknowledgments
We like to acknowledge Marcelo Toro for the assistance with flow cytometry. This study was supported by Swedish Heart- and Lung Foundation, EU Commission FP7 "CardioCell", Swedish Research Council, AFA insurances and ALF. O.B. was supported by Deutsche Forschungsgemeinschaft.
References
- Ang KL. Limitations of conventional approaches to identify myocyte nuclei in histologic sections of the heart. American journal of physiology. Cell physiology. 2010. pp. 298–1603. [DOI] [PMC free article] [PubMed]
- Bergmann O. Identification of cardiomyocyte nuclei and assessment of ploidy for the analysis of cell turnover. Experimental cell research. 2011;327:188–194. doi: 10.1016/j.yexcr.2010.08.017. [DOI] [PubMed] [Google Scholar]
- Bergmann O. Evidence for Cardiomyocyte Renewal in Humans. Science. 1126;324:98–102. doi: 10.1126/science.1164680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walsh S. Cardiomyocyte cell cycle control and growth estimation. Cardiovascular Research. 2010. pp. 1–31. [DOI] [PubMed]
- Spalding K, Bhardwaj RD, Buchholz B, Druid H, Frisén J. Retrospective birth dating of cells in humans. Cell. 2005;122:133–143. doi: 10.1016/j.cell.2005.04.028. [DOI] [PubMed] [Google Scholar]
- Adler CP, Friedburg H, Herget GW, Neuburger M, Schwalb H. Variability of cardiomyocyte DNA content, ploidy level and nuclear number in mammalian hearts. Virchows Arch. 1996;429:159–164. doi: 10.1007/BF00192438. [DOI] [PubMed] [Google Scholar]
- Herget GW, Neuburger M, Plagwitz R, Adler CP. DNA content, ploidy level and number of nuclei in the human heart after myocardial infarction. Cardiovascular Research. 1997;36:45–51. doi: 10.1016/s0008-6363(97)00140-5. [DOI] [PubMed] [Google Scholar]
- Adler CP, Friedburg H. Myocardial DNA content. ploidy level and cell number in geriatric hearts: postmortem examinations of human myocardium in old age. Mol. Cell Cardiol. 1986;18:3953–39. doi: 10.1016/s0022-2828(86)80981-6. [DOI] [PubMed] [Google Scholar]
- Dubois NC. SIRPA is a specific cell-surface marker for isolating cardiomyocytes derived from human pluripotent stem cells. Nature. 2011;29:1011–1018. doi: 10.1038/nbt.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hattori F. Nongenetic method for purifying stem cell-derived cardiomyocytes. Nature Methods. 2010;7:61–66. doi: 10.1038/nmeth.1403. [DOI] [PubMed] [Google Scholar]
- Fransioli J. Evolution of the c-kit-Positive Cell Response to Pathological Challenge in the Myocardium. Stem Cells. 2008;26:1315–1324. doi: 10.1634/stemcells.2007-0751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elliott DA. NKX2-5(eGFP/w) hESCs for isolation of human cardiac progenitors and cardiomyocytes. Nature Methods. 2011;8:1037–1040. doi: 10.1038/nmeth.1740. [DOI] [PubMed] [Google Scholar]
- Laflamme MA. Evidence for Cardiomyocyte Repopulation by Extracardiac Progenitors in Transplanted Human Hearts. Circulation Research. 2002;90:634–640. doi: 10.1161/01.res.0000014822.62629.eb. [DOI] [PubMed] [Google Scholar]
- Srsen V, Fant X, Heald R, Rabouille C, Merdes A. Centrosome proteins form an insoluble perinuclear matrix during muscle cell differentiation. BMC cell biology. 2009;10:28. doi: 10.1186/1471-2121-10-28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spoelgen R. A novel flow cytometry-based technique to measure adult neurogenesis in the brain. Journal of neurochemistry. 2011;119:165–175. doi: 10.1111/j.1471-4159.2011.07413.x. [DOI] [PubMed] [Google Scholar]
- Soonpaa MH, Kim KK, Pajak L, Franklin M, Field LJ. Cardiomyocyte DNA synthesis and binucleation during murine development. The American journal of physiology. 1996;271:H2183–H2189. doi: 10.1152/ajpheart.1996.271.5.H2183. [DOI] [PubMed] [Google Scholar]
- Olivetti G. Aging, cardiac hypertrophy and ischemic cardiomyopathy do not affect the proportion of mononucleated and multinucleated myocytes in the human heart. J Mol Cell Cardiol. 1996;28:1463–1477. doi: 10.1006/jmcc.1996.0137. [DOI] [PubMed] [Google Scholar]
- Okada S. Flow cytometric sorting of neuronal and glial nuclei from central nervous system tissue. Journal of cellular physiology. 2011;226:552–558. doi: 10.1002/jcp.22365. [DOI] [PubMed] [Google Scholar]
- Matevossian A, Akbarian S. Neuronal Nuclei Isolation from Human Postmortem Brain Tissue. J. Vis. Exp. 2008. p. e914. [DOI] [PMC free article] [PubMed]