

Abstract
Atherogenesis is potentiated by metabolic abnormalities that contribute to a heightened state of systemic inflammation resulting in endothelial dysfunction. However, early functional changes in endothelium that signify an individual's level of risk are not directly assessed clinically to help guide therapeutic strategy. Moreover, the regulation of inflammation by local hemodynamics contributes to the non-random spatial distribution of atherosclerosis, but the mechanisms are difficult to delineate in vivo. We describe a lab-on-a-chip based approach to quantitatively assay metabolic perturbation of inflammatory events in human endothelial cells (EC) and monocytes under precise flow conditions. Standard methods of soft lithography are used to microfabricate vascular mimetic microfluidic chambers (VMMC), which are bound directly to cultured EC monolayers.1 These devices have the advantage of using small volumes of reagents while providing a platform for directly imaging the inflammatory events at the membrane of EC exposed to a well-defined shear field. We have successfully applied these devices to investigate cytokine-,2 lipid-3, 4 and RAGE-induced5 inflammation in human aortic EC (HAEC). Here we document the use of the VMMC to assay monocytic cell (THP-1) rolling and arrest on HAEC monolayers that are conditioned under differential shear characteristics and activated by the inflammatory cytokine TNF-α. Studies such as these are providing mechanistic insight into atherosusceptibility under metabolic risk factors.
Keywords: Biomedical Engineering, Issue 65, Bioengineering, Immunology, Molecular Biology, Genetics, endothelial cell, monocyte arrest, microfluidics, shear stress, cytokine, atherosclerosis, inflammation
Protocol
1. Cell Culture and Substrate Preparation
Cut 3-inch circular substrates from a 100 x 20 mm tissue culture dish (BD Falcon) using a lathe. Sterilize substrates by submersion in 70% ethanol. Place in a Petri dish and coat with 4 ml type-I collagen (100 μg/ml) for 1 hr at room temperature, then rinse with 4 ml 1 x PBS.
Suspend Human Aortic Endothelial Cells (HAEC, passage 4-6) at 6.5x105 cells/ml and seed by applying 1 ml directly to substrate. Place in a 37 °C, 5% CO2 incubator and allow cells to adhere to the substrate for 1 hr.
Add 9 ml of Endothelial Growth Medium-2 (EGM-2) supplemented with 1x antibiotic-antimycotic solution to the cells. Change media every 2 days until the cells reach 90% confluency.
Maintain THP-1 cells in RPMI 1640 medium supplemented with 10% FBS and 1x antibiotic-antimycotic solution. Passage cells when they reach a density of 106 cells/ml.
2. Cell Shearing Protocol
Cells are conditioned in a custom cone-and-plate Cell Shearing Device (CSD). Device details and design specifications are documented in Table 1 and Figure 4.
Prepare the CSD. Sterilize the chamber housing with ethanol and rinse with sterile PBS. Heat the housing to 37 °C and level to ensure that the cone is perpendicular to the cell monolayer. Assemble the substrate with the cells from the bottom of the housing and set the cone to the desired height.
Use Leibovitz-15 (L-15) medium supplemented with Endothelial BulletKit as the shearing medium to maintain proper pH in the absence of CO2. To stimulate inflammation, add tumor necrosis factor-α (TNF-α, 0.5 ng/ml) to the shearing medium and inject 6 ml in the device via a syringe connected to an inlet port, taking care not to introduce air bubbles in the chamber.
Shear cells at a high magnitude steady shear stress (HSS, 15 dyne/cm2) or an oscillatory shear stress (OSS, 0 ± 5 dyne/cm2, 1 Hz) for 4 hr. The desired SS is achieved by rotating the cone above the cells, which is precisely controlled with a micro-stepping motor as previously described.6 The wall SS at the surface (τw) is approximated by:
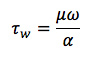 where μ is the viscosity of the medium, ω is the angular velocity of the cone, and α is the cone angle (0.5°). The cone alignment with the substrate is adjusted to within 0.05° through precision leveling and the gap height set at 20 ± 10 μm using a depth gauge to minimize circumferential and radial variation in τw. The device has been validated to ensure laminar flow characteristics. Remove cells from the device and prepare for analysis of monocyte adhesion.
where μ is the viscosity of the medium, ω is the angular velocity of the cone, and α is the cone angle (0.5°). The cone alignment with the substrate is adjusted to within 0.05° through precision leveling and the gap height set at 20 ± 10 μm using a depth gauge to minimize circumferential and radial variation in τw. The device has been validated to ensure laminar flow characteristics. Remove cells from the device and prepare for analysis of monocyte adhesion.
3. Fabrication of PDMS Microfluidic Chamber
Obtain master mold for desired microfluidic channel number and channel height depending on the application. The characteristics of the specific design used in this protocol are detailed in Table 1. The method for creating these is already well documented in the literature.1 Briefly, the network of channels is designed using CAD software (Autodesk Inventor) and printed at 5000 dpi on a transparency. Negative photoresist (SU8) is spun onto a silicon wafer to a thickness of 100 μm. The transparency is overlaid and exposed to UV light. Non-polymerized photoresist is removed to generate the positive replica master.
To make PDMS solution mix silicone base with silicone curing agent 10:1 by weight in a weigh boat and stir with a 5 ml serological pipet. Be careful not to scrape the walls of the weigh boat to avoid adding plastic flakes to the solution.
Place the master wafer in a 100x15mm Petri dish and pour the PDMS solution into the dish.
Cover the dish and place it into a vacuum container for 15 minutes to remove bubbles. After 15 minutes take the Petri dish out and remove any excess bubbles by gently blowing air over the surface.
Place the Petri dish in an oven for 1 hour at 70 °C. Remove the dish and cut out the microfluidic chambers using a sharp blade.
4. Assembly of Device on Microscope
Turn on light source to inverted microscope and set heater to 37 °C. Attach tubing to a 10 ml syringe and fill with distilled water such that there are no bubbles. Place the syringe in the syringe pump and set flow rate to induce a wall SS of 1 dyne/cm2. The wall shear stress (τw) generated along the centerline of the channel is approximated using the standard equation for a parallel-plate flow chamber:
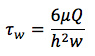 where μ is the viscosity of the medium, Q is the volumetric flow rate, h is the channel height, and w is the channel width. Note: since the channels are of finite width the actual τw will vary with w depending on the channel aspect ratio, and measurements are avoided at the 25% nearest the side walls.1,7
where μ is the viscosity of the medium, Q is the volumetric flow rate, h is the channel height, and w is the channel width. Note: since the channels are of finite width the actual τw will vary with w depending on the channel aspect ratio, and measurements are avoided at the 25% nearest the side walls.1,7Place PDMS chamber in water and use a 100μl micropipette to remove air from the individual channels. Attach vacuum adapter to chamber making sure that the valve is set to off.
Place Petri dish with HAEC monolayer over the objective. Place the PDMS chamber over the Petri dish and carefully set it on the HAEC monolayer while assuring that no air bubbles form between the device and the monolayer. Turn the valve so that the vacuum is on and the chamber is firmly attached.
Cut 19 gauge needles blunt such that there is only 5 mm of metal left. Fill the plastic reservoir with buffer and put into PDMS chamber entrance. Repeat for appropriate number of channels to be used. Attach tubing from syringe pumps to PDMS channel exits.
5. Monocyte Adhesion Assay Under Shear Stress
Suspend THP-1 cells at 2x106 cells/ml in HBSS + Ca2+/Mg2+ + 0.1% HSA. Activate THP-1 cells by incubating with stromal derived factor-1 (SDF-1, 50 μg/ml) for 15 min at room temperature.
Turn on syringe pump to establish flow, then add 50 μl of cell suspension to reservoir without introducing air bubbles. Once flow is fully developed, begin data acquisition.
Run for 5 min at appropriate flow rate, adding buffer to the reservoir if the solution gets low. Continue flowing buffer into the channels to wash out any unbound cells in the flow stream.
6. Data Acquisition and Analysis
Acquire digital image sequences at 3 frames/s for 2 min following fully developed flow along the longitudinal axis of the channel at 3 locations in the channel, taking care not to record immediately after entrance or before exit ports or in the 25% nearest the side walls.
Count the number of arrested cells per field by identifying their phase-bright appearance in the same focal plane as the monolayer.
Firm cell arrest is defined by movement < ½ cell diameter in 10 s. Average data over 3 random fields / channel and quantify using ImageJ software.
7. Representative Results
Although the vascular endothelium is uniformly exposed to agonists that contribute to systemic inflammation, atherosclerosis is a focal disease that develops preferentially at sites of flow disturbance in arteries, implicating spatial heterogeneity in endothelial function.8 Circulating inflammatory mediators such as TNF-α are elevated in human serum under metabolic stress and correlate with impaired endothelial function.9, 10 TNF-α induces a well-characterized response in EC that includes up-regulation of cell adhesion molecules (CAMs; e.g. VCAM-1, ICAM-1, E-selectin) and chemokines that recruit monocytes from the circulation, a hallmark of the early atherosclerotic lesion.11 Hemodynamics is an important regulator of endothelial inflammatory phenotype.12 High magnitude shear stress (HSS; e.g. 15 dynes/cm2) or pulsatile shear stress (PSS; e.g. 15 ± 5 dynes/cm2) associated with sites of resistance to atherosclerosis in arteries down-regulate TNF-α-induced responses. Conversely, low magnitude shear stress (LSS; e.g. 2 dynes/cm2) or oscillatory shear (OSS; e.g. 0 ± 5 dynes/cm2), characteristic of susceptible sites in vivo, exacerbate cytokine-induced inflammation.13, 14 Our microfluidic PDMS devices provide a platform for conducting mechanistic studies to test hypotheses related to the superposition of metabolic stress with hydrodynamic factors in the regulation of endothelial function and pathology. The utility and versatility of the approach is broadly illustrated through the following representative results.
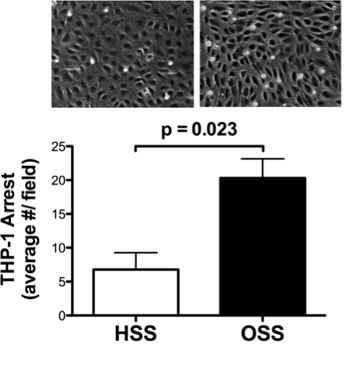 Figure 1. Shear stress differentially modulates THP-1 cell adhesion to TNF-α inflamed endothelial monolayers. These data were acquired using the detailed protocol described above and documented in the video. HAEC monolayers were simultaneously exposed to a low dose of the inflammatory cytokine TNF-α (0.5 ng/ml, ~EC50 for VCAM-1 up-regulation) and either HSS (15 dynes/cm2) or OSS (0 ± 5 dynes/cm2, 1 Hz) in a custom CSD for 4 hr as described in the detailed protocol. THP-1 cell arrest was quantified under shear flow in a PDMS flow chamber. This cell line that mimics the phenotype of human monocytes was chosen for its reproducibility and ease of use. HSS attenuated the inflammatory response compared to OSS, resulting in decreased THP-1 cell arrest. Data were analyzed by Student's t-test and are represented as mean ± SEM of 3 independent experiments. Adapted with permission from DeVerse et al.5
Figure 1. Shear stress differentially modulates THP-1 cell adhesion to TNF-α inflamed endothelial monolayers. These data were acquired using the detailed protocol described above and documented in the video. HAEC monolayers were simultaneously exposed to a low dose of the inflammatory cytokine TNF-α (0.5 ng/ml, ~EC50 for VCAM-1 up-regulation) and either HSS (15 dynes/cm2) or OSS (0 ± 5 dynes/cm2, 1 Hz) in a custom CSD for 4 hr as described in the detailed protocol. THP-1 cell arrest was quantified under shear flow in a PDMS flow chamber. This cell line that mimics the phenotype of human monocytes was chosen for its reproducibility and ease of use. HSS attenuated the inflammatory response compared to OSS, resulting in decreased THP-1 cell arrest. Data were analyzed by Student's t-test and are represented as mean ± SEM of 3 independent experiments. Adapted with permission from DeVerse et al.5
The following data demonstrate the versatility of our approach using examples from our published work. Departures from the above protocol are indicated where applicable.
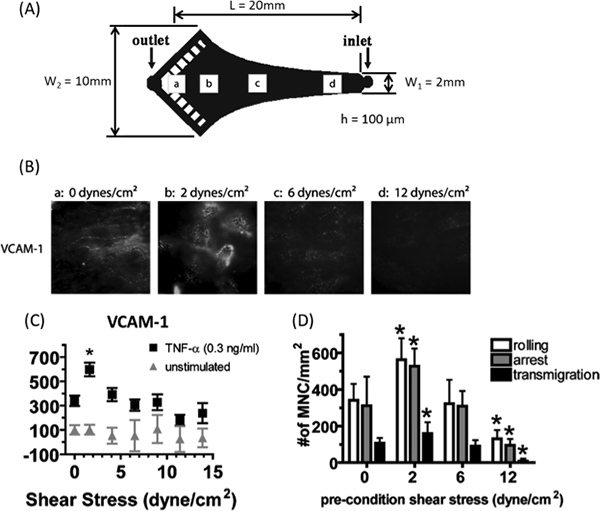 Figure 2. Modulation of cytokine-induced VCAM-1 expression and monocyte recruitment in a shear stress gradient. A) HAEC monolayers were stimulated with TNF-α (0.3 ng/ml) for 4 hr under flow in a VMMC designed based on Hele-Shaw flow theory.15 A linear decrease in SS magnitude along the centerline of the channel was achieved by designing the sidewalls of the chamber to coincide with the streamlines of a two dimensional stagnation flow and shaping the end of the channel to match the iso-potential lines. The wall shear stress (τw) generated along the centerline of this channel at an axial distance from the entrance (x) is given by:
Figure 2. Modulation of cytokine-induced VCAM-1 expression and monocyte recruitment in a shear stress gradient. A) HAEC monolayers were stimulated with TNF-α (0.3 ng/ml) for 4 hr under flow in a VMMC designed based on Hele-Shaw flow theory.15 A linear decrease in SS magnitude along the centerline of the channel was achieved by designing the sidewalls of the chamber to coincide with the streamlines of a two dimensional stagnation flow and shaping the end of the channel to match the iso-potential lines. The wall shear stress (τw) generated along the centerline of this channel at an axial distance from the entrance (x) is given by:
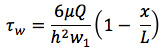
where μ is the viscosity of the medium, Q is the volumetric flow rate, h is the channel height, w1 is the channel entrance width, and L is the total channel length. Q was chosen to generate a gradient in SS magnitude from 16 dynes/cm2 at the inlet to 0 at the stagnation point "a" just proximal to the outlet. B)-C) VCAM-1 expression was measured in situ by immunofluorescence microscopy using antibodies conjugated to quantum dots and presented as percentage median fluorescence intensity (MFI) of unstimulated static control. SS did not modulate the basal level of VCAM-1, which is low in unstimulated HAEC (grey triangles). However, VCAM-1 expression increased relative to TNF-α at LSS magnitudes, peaking at ~2 dynes/cm2 and returning to the TNF-α baseline or below at shear magnitudes > 5 dynes/cm2 (black squares). D) Monocytes (106/ml) were perfused at 2 dynes/cm2 for 5 min in parallel flow channels over monolayers preconditioned as in A). Monocyte recruitment to endothelium quantitatively correlated with the VCAM-1 expression at a given shear stress magnitude. Notably, the ability to probe VCAM-1 expression and monocyte recruitment with fine spatial resolution in a monolayer revealed significant changes over a narrow range in SS magnitude that would not be appreciated by applying only a few discrete values of shear in separate experiments. Data were analyzed by repeated measures ANOVA and differences assessed by Newman-Keuls posttest, *P < 0.05 from TNF-α under static conditions. Data are mean ± SEM from 3-6 independent experiments. Adapted with permission from Tsou et al.2
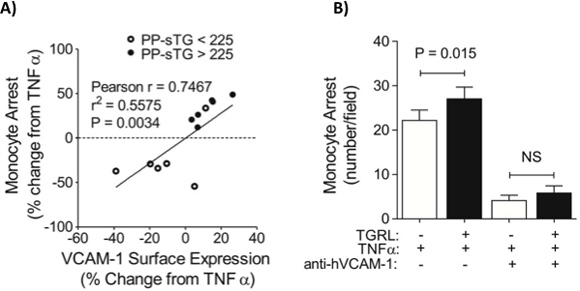 Figure 3. Triglyceride-rich lipoproteins (TGRL) modulate cytokine-induced inflammation in proportion to donor serum triglyceride levels and VCAM-1 expression. HAEC monolayers were stimulated with TNF-α (0.3 ng/ml) for 4 hr simultaneously with TGRL (10 mg/dl ApoB) isolated from human subjects after a high fat meal in static culture. A) VCAM-1 expression was measured separately in suspended EC by flow cytometry and monocyte adhesion quantified using a VMMC as described in the detailed protocol. Data are presented as % change from TNF-α. Monocyte arrest varied in direct proportion to endothelial VCAM-1 expression, which increased directly with the level of donor serum triglyceride. B) Monocyte arrest for a subset of pro-inflammatory donors was abrogated in the presence of a VCAM-1 blocking antibody. Significance was determined by paired Student's t-test, means ± SEM from 3-5 independent experiments. Adapted with permission from Wang et al.4
Figure 3. Triglyceride-rich lipoproteins (TGRL) modulate cytokine-induced inflammation in proportion to donor serum triglyceride levels and VCAM-1 expression. HAEC monolayers were stimulated with TNF-α (0.3 ng/ml) for 4 hr simultaneously with TGRL (10 mg/dl ApoB) isolated from human subjects after a high fat meal in static culture. A) VCAM-1 expression was measured separately in suspended EC by flow cytometry and monocyte adhesion quantified using a VMMC as described in the detailed protocol. Data are presented as % change from TNF-α. Monocyte arrest varied in direct proportion to endothelial VCAM-1 expression, which increased directly with the level of donor serum triglyceride. B) Monocyte arrest for a subset of pro-inflammatory donors was abrogated in the presence of a VCAM-1 blocking antibody. Significance was determined by paired Student's t-test, means ± SEM from 3-5 independent experiments. Adapted with permission from Wang et al.4
Discussion
We describe the use of microfluidic PDMS devices for the on-chip assessment of endothelial inflammatory phenotype through the real-time imaging of CAM expression and monocyte adhesion. A major advantage of our approach lies in the ability to quantify outcomes associated with endothelial dysfunction in cells exposed to inflammatory mediators such as dietary lipids and cytokines under defined hydrodynamic conditions that mirror shear stress in atherogenic vessels. The VMMC facilitate the use of small amounts of reagents or materials that may be in limited supply and have the added advantages of being inexpensive, customizable and amenable to high throughput.
The use of microfluidic devices represents an alternative to macroscale approaches that have been used for decades to investigate the response of cultured EC to shear. These include parallel-plate flow chambers, cylindrical tubes, and cone-and-plate viscometers, each of which is capable of imposing steady, laminar Poiseuille flow conditions (reviewed in 16). Modifications to these devices can produce temporal or spatial variations in wall shear stress or even recapitulate arterial waveforms.6 However, in general these devices require a large amount of media, are expensive to fabricate, less adaptable, and suffer from comparably low throughput.
Limitations also exist with the use of the VMMCs. These devices are designed based on parallel-plate flow theory, which assumes infinite width. This is a reasonable assumption for macroscale devices where width is much greater than height. However, the finite width of the microfluidic channels necessitates that studies be conducted along the center of the longitudinal axis of the channel to eliminate side wall effects, as the actual τw will vary with w depending on the channel aspect ratio.1, 7 Similarly, measurements are not taken close to the entrance and exit of channels to avoid entrance effects. We have ruled out any significant paracrine signaling effects or inflammation induced by injury or damage to EC caused by sealing the device to the monolayer as contributing to our observed responses. PDMS has a high affinity for hydrophobic molecules, which could lead to the absorption of secreted chemokines and non-specific interactions between leukocytes and the VMMC. However, we have not found this to be a problem for the duration of our studies.2
The use of flow devices with cultured cells constitutes a reductionist approach that facilitates mechanistic studies and the demonstration of cause and effect with respect to shear stress. However, a limitation of any in vitro approach is the assumption that it will adequately recapitulate the complexity of the environment in native arteries. In vivo, endothelial cells are chronically adapted to complex flow patterns and are subject to interactions with other cells, matrix, and humoral mediators that regulate their behavior.
It is noteworthy that all of the results presented above are in short-term models and as such may not be time invariant. However, they are representative of experiments which we commonly perform to investigate mechanism under an acute inflammatory challenge. A relevant strategy for our studies is to monitor changes from an inflammatory baseline of low-dose TNF-α stimulation, for which 4 hr represents the approximate peak in VCAM-1 expression. We have documented in several studies that changes in the surface expression of VCAM-1 correlate with enhanced monocyte recruitment under these conditions.2-4, 17
Moreover, the choice of SS waveform can evoke differential endothelial responses. In our representative protocol we document the use of the VMMC to quantify monocyte adhesion events in HAEC conditioned under shear stresses chosen to be representative of sites of relative resistance (HSS) and susceptibility (OSS) to atherogenesis in arteries in a customized cell shearing device. Consistent with previous findings in cultured EC, HSS attenuated the response to cytokine activation relative to OSS.12, 13 The results are consistent with a greater permissiveness to inflammatory activation of EC at sites of flow disturbance in vivo. While these conditions are commonly used to represent undisturbed and disturbed flow respectively, they do not fully recapitulate the complexity of flow in arteries. However, a wealth of literature supports their use as simple approximations that adequately reproduce the salient flow features.14
Notably, our cone-and-plate-based CSD is capable of generating more complex waveforms and conditioning cells for longer periods as previously reported.6 Thus, pairing the VMMC with the CSD in the described protocol imparts a maximum degree of flexibility. A consideration of this approach lies in the orientation of the VMMC with respect to the direction of shear conditioning in the CSD, in which HAEC experience flow in the circumferential direction. In practice, the VMMC is oriented so that channels are parallel with the flow field and the inlet upstream of conditioned cells so far as possible. The channels are small enough that this represents a reasonable approximation. However, we also demonstrated in a previous study that flowing monocytes parallel to or perpendicular to the direction of shear conditioning in the VMMC did not affect the results shown in Figure 2.2
Additional considerations associated with in vitro techniques include variability induced by phenotypic drift associated with serial passaging of EC in culture and heterogeneity in isolated EC or monocytes. HAEC should typically be used by passage 6 and each lot characterized for a consistent inflammatory response to TNF-α. The use of a cell line such as THP-1 can reduce the variability associated with the use of monocytes, which vary in activity by donor and are easily activated by their isolation. In order to ensure reproducible and accurate functional readouts, cell preparation and transfer times should be minimized. The careful selection of controls is necessary to meaningfully interpret the results. Depending on the experiment, these might include untreated or static-cultured cells, TNF-α treatment, unactivated THP-1 cells, the use of blocking antibodies, siRNAs or pharmacological inhibitors, and others.
As testament to the versatility of the approach, we have applied these devices to investigate cytokine-,2 lipid-3, 4 and RAGE-induced5 inflammation in HAEC. Our studies have included measuring the inflammatory potential of serum-derived lipid particles from human subjects on endothelial cells3, 4 and monocytes.17 In an alternative strategy, monocytes may be presented with substrates derivatized with immobilized molecules such as VCAM-1 to examine adhesion mechanisms. Using this approach, we previously demonstrated a functional role for the adhesion molecule CD11c, which cooperates with very late antigen (VLA)-4 in monocyte adhesion to VCAM-1 during dyslipidemia in humans17 and mice.18 Since monocytes can be easily activated by their isolation, we also developed a whole blood assay for use with our lab-chip device to allow for more sensitive and reproducible detection.17
Ultimately, in addition to providing insight into the early inflammatory events underlying atherosclerosis, we envision that this technology development will lead to faithful ex vivo approaches for assessing an individual's risk of inflammation-mediated cardiovascular disease, paving the way for the development of personalized diagnostics that report on monocyte and EC activation.
| Cone-and-Plate Cell Shearing Device | ||
| Cone Radius | 0.06985 | m |
| Cone Angle | 0.00877 (0.5°) | rad (degrees) |
| Gap Height | 20 ± 10 | μm |
| Max Angular Velocity (for experiments outlined here) | 18.85 | rad/s |
| Kinematic Viscosity | 0.00077 | m2/s |
| Max R number (for experiments outlined here) | 0.000758 | dimensionless |
| VMMC Specifications | ||
| Channel Length | 8.0 | mm |
| Channel Height | 60 | μm |
| Channel Width | 2 | mm |
| Reynolds Number | 1.67 | dimensionless |
Table 1. CSD and VMMC Specifications.
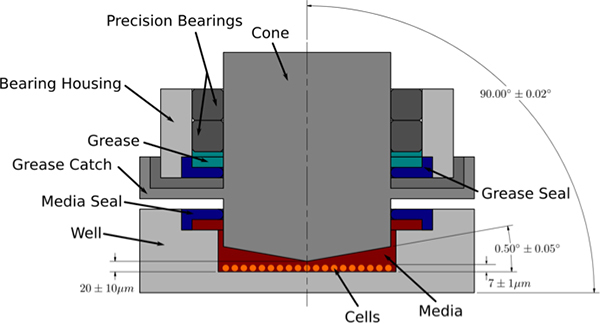 Figure 4. Cross-sectional view of cone-and-plate cell shearing device (CSD). The CSD consists of a stainless steel cylindrical cone that rotates above a stationary cell culture substrate. The cone angle and gap height are critical parameters that define the nature of the shear field produced in the presence of culture medium. A modified Reynolds number showing the relationship of centrifugal to viscous forces is given by:
Figure 4. Cross-sectional view of cone-and-plate cell shearing device (CSD). The CSD consists of a stainless steel cylindrical cone that rotates above a stationary cell culture substrate. The cone angle and gap height are critical parameters that define the nature of the shear field produced in the presence of culture medium. A modified Reynolds number showing the relationship of centrifugal to viscous forces is given by:
![]()
where r is cone radius, ω is angular velocity, α is cone angle, and ν is the kinematic velocity of the fluid. When R is << 1 centrifugal forces are low, flow is laminar and purely azimuthal, and the induced shear stress is constant over the monolayer.19 In our CSD, the substrate with cultured cells is loaded from the bottom into the chamber that houses the cone. Medium is added to the chamber and used as the shearing fluid. The cone is mounted with pre-loaded precision bearings and the rotation driven by a micro-stepping motor. A precision leveling system maintains alignment perpendicular to the cell substrate. The gap height (defined by the distance from the apex of the cone to the cell substrate) is set with a high-precision depth gauge. A grease catch and media seal are in place to ensure that the chamber housing the cells is kept clean and free of oil and debris.
Disclosures
No conflicts of interest declared.
Acknowledgments
This work was supported by NIH/NHLBI grant R01 HL082689 to Scott I. Simon and Anthony G. Passerini.
References
- Schaff UY, Xing MM, Lin KK, Pan N, Jeon NL, Simon SI. Vascular mimetics based on microfluidics for imaging the leukocyte--endothelial inflammatory response. Lab Chip. 2007;7:448–456. doi: 10.1039/b617915k. [DOI] [PubMed] [Google Scholar]
- Tsou JK, Gower RM, Ting HJ, Schaff UY, Insana MF, Passerini AG, Simon SI. Spatial regulation of inflammation by human aortic endothelial cells in a linear gradient of shear stress. Microcirculation. 2008;15:311–323. doi: 10.1080/10739680701724359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ting HJ, Stice JP, Schaff UY, Hui DY, Rutledge JC, Knowlton AA, Passerini AG, Simon SI. Triglyceride-rich lipoproteins prime aortic endothelium for an enhanced inflammatory response to tumor necrosis factor-alpha. Circ. Res. 2007;100:381–390. doi: 10.1161/01.RES.0000258023.76515.a3. [DOI] [PubMed] [Google Scholar]
- Wang YI, Schulze J, Raymond N, Tomita T, Tam K, Simon SI, Passerini AG. Endothelial inflammation correlates with subject triglycerides and waist size after a high-fat meal. Am. J. Physiol. Heart Circ. Physiol. 2011;300:H784–H791. doi: 10.1152/ajpheart.01036.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deverse JS, Bailey KA, Jackson KN, Passerini AG. Shear stress modulates rage-mediated inflammation in a model of diabetes-induced metabolic stress. Am. J. Physiol. Heart Circ. Physiol. 2012. [DOI] [PMC free article] [PubMed]
- Dai G, Kaazempur-Mofrad MR, Natarajan S, Zhang Y, Vaughn S, Blackman BR, Kamm RD, Garcia-Cardena G, Gimbrone MA. Distinct endothelial phenotypes evoked by arterial waveforms derived from atherosclerosis-susceptible and -resistant regions of human vasculature. Proc. Natl. Acad. Sci. U.S.A. 2004;101:14871–14876. doi: 10.1073/pnas.0406073101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Young EW, Wheeler AR, Simmons CA. Matrix-dependent adhesion of vascular and valvular endothelial cells in microfluidic channels. Lab Chip. 2007;7:1759–1766. doi: 10.1039/b712486d. [DOI] [PubMed] [Google Scholar]
- Davies PF, Civelek M, Fang Y, Guerraty MA, Passerini AG. Endothelial heterogeneity associated with regional athero-susceptibility and adaptation to disturbed blood flow in vivo. Semin Thromb Hemost. 2008;36:265–275. doi: 10.1055/s-0030-1253449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burdge GC, Calder PC. Plasma cytokine response during the postprandial period: A potential causal process in vascular disease. Br. J. Nutr. 2005;93:3–9. doi: 10.1079/bjn20041282. [DOI] [PubMed] [Google Scholar]
- Hotamisligil GS. Inflammation and metabolic disorders. Nature. 2006;444:860–867. doi: 10.1038/nature05485. [DOI] [PubMed] [Google Scholar]
- Libby P. Inflammation in atherosclerosis. Nature. 2002;420:868–874. doi: 10.1038/nature01323. [DOI] [PubMed] [Google Scholar]
- Nigro P, Abe J, Berk BC. Flow shear stress and atherosclerosis: A matter of site specificity. Antioxid Redox Signal. 2011;15:1405–1414. doi: 10.1089/ars.2010.3679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chiu JJ, Lee PL, Chen CN, Lee CI, Chang SF, Chen LJ, Lien SC, Ko YC, Usami S, Chien S. Shear stress increases icam-1 and decreases vcam-1 and e-selectin expressions induced by tumor necrosis factor-[alpha] in endothelial cells. Arterioscler. Thromb. Vasc. Biol. 2004;24:73–79. doi: 10.1161/01.ATV.0000106321.63667.24. [DOI] [PubMed] [Google Scholar]
- Li YS, Haga JH, Chien S. Molecular basis of the effects of shear stress on vascular endothelial cells. J. Biomech. 2005;38:1949–1971. doi: 10.1016/j.jbiomech.2004.09.030. [DOI] [PubMed] [Google Scholar]
- Usami S, Chen HH, Zhao Y, Chien S, Skalak R. Design and construction of a linear shear stress flow chamber. Ann. Biomed. Eng. 1993;21:77–83. doi: 10.1007/BF02368167. [DOI] [PubMed] [Google Scholar]
- Helmke BP. Molecular control of cytoskeletal mechanics by hemodynamic forces. Physiology (Bethesda) 2005;20:43–53. doi: 10.1152/physiol.00040.2004. [DOI] [PubMed] [Google Scholar]
- Gower RM, Wu H, Foster GA, Devaraj S, Jialal I, Ballantyne CM, Knowlton AA, Simon SI. Cd11c/cd18 expression is upregulated on blood monocytes during hypertriglyceridemia and enhances adhesion to vascular cell adhesion molecule-1. Arterioscler. Thromb. Vasc. Biol. 2011;31:160–166. doi: 10.1161/ATVBAHA.110.215434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu H, Gower RM, Wang H, Perrard XY, Ma R, Bullard DC, Burns AR, Paul A, Smith CW, Simon SI, Ballantyne CM. Functional role of cd11c+ monocytes in atherogenesis associated with hypercholesterolemia. Circulation. 2009;119:2708–2717. doi: 10.1161/CIRCULATIONAHA.108.823740. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bussolari SR, Dewey CF, Gimbrone MA. Apparatus for subjecting living cells to fluid shear stress. The Review of scientific instruments. 1982;53:1851–1854. doi: 10.1063/1.1136909. [DOI] [PubMed] [Google Scholar]