

Abstract
α-Enolase is a key glycolytic enzyme in the cytoplasm of prokaryotic and eukaryotic cells and is considered a multifunctional protein. α-enolase is expressed on the surface of several cell types, where it acts as a plasminogen receptor, concentrating proteolytic plasmin activity on the cell surface. In addition to glycolytic enzyme and plasminogen receptor functions, α-Enolase appears to have other cellular functions and subcellular localizations that are distinct from its well-established function in glycolysis. Furthermore, differential expression of α-enolase has been related to several pathologies, such as cancer, Alzheimer's disease, and rheumatoid arthritis, among others. We have identified α-enolase as a plasminogen receptor in several cell types. In particular, we have analyzed its role in myogenesis, as an example of extracellular remodelling process. We have shown that α-enolase is expressed on the cell surface of differentiating myocytes, and that inhibitors of α-enolase/plasminogen binding block myogenic fusion in vitro and skeletal muscle regeneration in mice. α-Enolase could be considered as a marker of pathological stress in a high number of diseases, performing several of its multiple functions, mainly as plasminogen receptor. This paper is focused on the multiple roles of the α-enolase/plasminogen axis, related to several pathologies.
1. Introduction
Enolase, also known as phosphopyruvate hydratase, was discovered in 1934 by Lohman and Mayerhof. It is one of the most abundantly expressed cytosolic proteins in many organisms. It is a key glycolytic enzyme that catalyzes the dehydratation of 2-phosphoglycerate to phosphoenolpyruvate, in the last steps of the catabolic glycolytic pathway [1] (Figure 1). It is a metalloenzyme that requires the metal ion magnesium (Mg2+) to be catalytically active. Enolase is found from archaebacteria to mammals, and its sequence is highly conserved [2]. In vertebrates, the enzyme occurs as three isoforms: α-enolase (Eno1) is found in almost all human tissues, whereas β-enolase (Eno3) is predominantly found in muscle tissues, and γ-enolase (Eno2) is only found in neuron and neuroendocrine tissues [3]. The three enolase isoforms share high-sequence identity and kinetic properties [4–6]. Enzymatically active enolase which exists in a dimeric (homo- or heterodimers) form is composed of two subunits facing each other in an antiparallel fashion [6, 7]. The crystal structure of enolase from yeast and human has been determined and catalytic mechanisms have been proposed [8–10].
Figure 1.
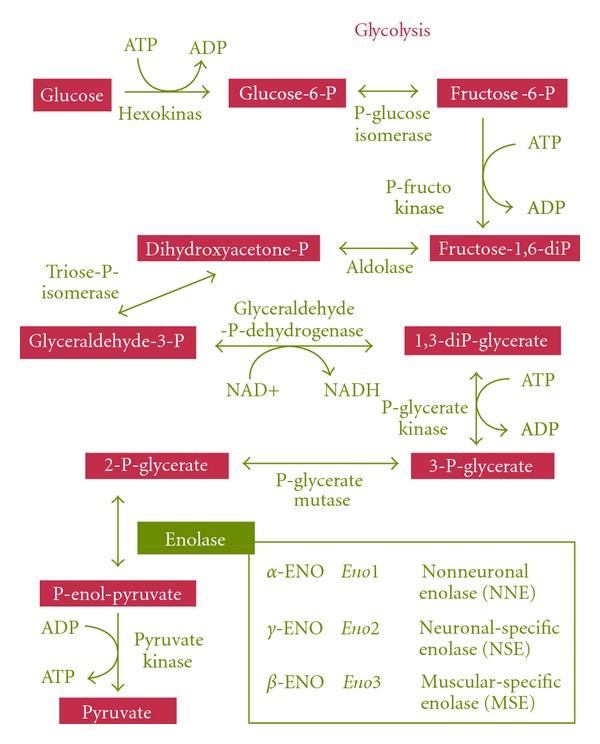
Summary of glycolytic metabolic pathway. Metabolic chain reactions of glycolysis, the central pathway for the catabolism of carbohydrates that takes place in the cytoplasm of almost all prokaryotic and eukaryotic cells. The insert shows different enolase isoenzymes in vertebrates.
Although it is expressed in most of the cells, the gene that encodes enolase is not considered a housekeeping gene since its expression varies according to the pathophysiological, metabolic, or developmental conditions of cells [11]. α-Enolase mRNA translation which is primarily under developmental control is significantly upregulated during cellular growth and practically undetectable during quiescent phases [12, 13].
Recent accumulation of evidence revealed that, in addition to its innate glycolytic function, α-enolase plays an important role in several biological and pathophysiological processes: by using an alternative stop codon, the α-enolase mRNA can be translated into a 37 kDa protein which lacks the first 96 amino acid residues. This protein, named c-myc promoter-binding protein 1 (MBP-1) is localized in the nucleus and can bind to the c-myc P2 promoter and negatively regulates transcription of the protooncogene [14]. α-Enolase has been detected on the surface of hematopoietic cells such as monocytes, T cells and B cells, neuronal cells, and endothelial cells as a strong plasminogen receptor, modulating pericellular fibrinolytic activity. The expression of α-enolase on the surface of a variety of eukaryotic cells has been found to be dependent on the pathophysiological conditions of these cells [15–19].
α-Enolase has also been described as a neurotrophic factor [20], a heat-shock protein (HSP48) [21], and a hypoxic stress protein [22]. Furthermore, α-enolase is part of the crystallin lens of vertebrates [23], binds to fragments of F-actin and tubulin [24], and has been detected associated to centrosomes in HeLa cells [25]. α-Enolase also binds with high affinity to other glycolytic enzymes: pyruvate kinase, phosphoglycerate mutase, which are adjacent to enolase in the glycolytic pathway, and to aldolase, which is known to associate with cytoskeletal proteins [26] (Figure 2).
Figure 2.
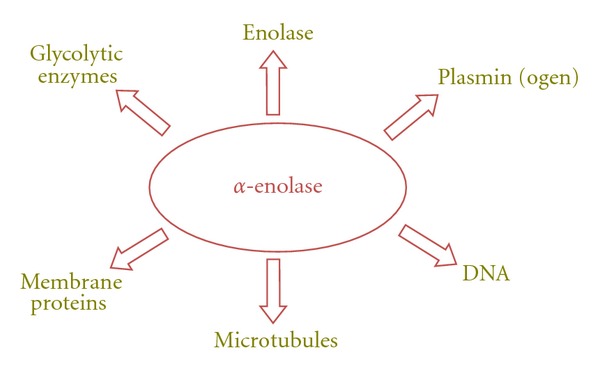
Interaction of α-enolase with other nuclear, cytoplasmic, or membrane molecules. α-Enolase can directly interact with other enolase isoforms (α, β, and γ) to form enzymatically active dimers, other glycolytic enzymes as pyruvate kinase, phosphoglycerate mutase and aldolase. It can also bind to microtubules network proteins, as F-actin and tubulin, and it is localized on the cell surface, interacting with other membrane proteins, where it binds to plasminogen and plasmin.
It has also been suggested that upregulation of α-enolase contributes to hypoxia tolerance through nonglycolytic mechanisms [27]. Increased expression of α-enolase has been reported to correlate with progression of tumors, neuroblastoma, and lung cancer, and enolase has been considered to be a potential diagnostic markers for many tumors [28–32].
Thus, α-enolase appears to be a “moonlighting protein,” one of a growing list of proteins that are recognized as identical gene products exhibiting multiple functions at distinct cellular sites through “gene sharing” [33, 34]. This paper is focused on the multiple roles of the α-enolase/plasminogen axis, related to several pathologies.
2. The Plasminogen Activation System
In multicellular organisms, extracellular proteolysis is important to many biological processes involving a dynamic rearrangement of cell-cell and cell-matrix interactions, being the plasminogen activation (PA) system among the most important extracellular proteases. The PA system comprises an inactive proenzyme, plasminogen, and ubiquitous in body fluid, that can be converted into the active enzyme, plasmin, by two physiological activators (PAs): tissue-type plasminogen activator (tPA) and urokinase-type plasminogen activator (uPA). Inhibition of the plasminogen system occurs at the level of the PA, by specific inhibitors (PAI-1 and PAI-2), or at the level of plasmin, by α2-antiplasmin (reviewed in [35]). The PA/plasmin system is a key regulator in extracellular matrix (ECM) remodeling directly by its ability to degrade ECM components, such as laminin or fibronectin, and indirectly via activation of matrix metalloproteinases (MMPs), which will degrade collagen(s) subsequently. Furthermore, plasmin is able to activate latent growth factors, such as transforming growth factor β (TGFβ) and basic fibroblast growth factor (bFGF) (reviewed in [35]).
Work from numerous groups has clearly demonstrated that the localization of plasminogen and its activators uPA and tPA on the cell surface, though association to specific cell membrane receptors, provides a mechanism for cells to harness and regulate the activities of these proteases [36, 37]. Binding sites for plasminogen, tPA, and uPA have been identified on a variety of cell types, including monocytes, fibroblasts, and endothelial cells [38, 39]. uPA is recruited to the cell membrane immediately after its secretion via a specific uPA receptor (uPAR, CD87), expressed on the cell surface, that localize extracellular proteolysis and induces cell migration, cell adhesion, and proliferation (reviewed in [40, 41]).
Described binding sites for plasminogen include α-enolase [18, 42], annexin A2 [43], p11 [44], histone H2B [45, 46], actin [47], gp330 [48], cytokeratin 8 [49], histidine-proline rich glycoprotein [50], glyceraldehide-3-phosphate dehydrogenase [51] gangliosides [18], and Plg-RTK [52]. α-Enolase and most of these proteins have C-terminal lysines predominantly responsible for plasminogen binding/activation [53]. Notably, most of these proteins have other described functions than plasminogen receptors, and lack a transmembrane domain, Plg-RTK being an exception, as it is a transmembrane receptor [52].
3. α-Enolase as a Plasminogen Receptor
We and others have previously identified α-enolase as a plasminogen receptor on the surfaces of several diverse cell types including carcinoma cells [42], monocytoid cells [15, 18], leukocytic cell lines [54], rat neuronal cells [16], and pathogenic streptococci [1].
On the cell surface, interaction of plasminogen with α-enolase enhances its activation by PAs, concentrates protease activity pericellularly [55–57], and protects plasmin from inhibition by α2-antiplasmin [18, 58] (Figure 3). In order to examine the role of α-enolase in the pericellular generation of plasmin activity, we produced a monoclonal antibody, MAb11G1, that specifically blocked plasminogen binding to purified α-enolase [54]. MAb11G1 allowed us to demonstrate that α-enolase occupancy by plasminogen on leukocytoid cells and on peripheral blood neutrophils is required for pericellular plasminogen activation and plasmin generation [54].
Figure 3.
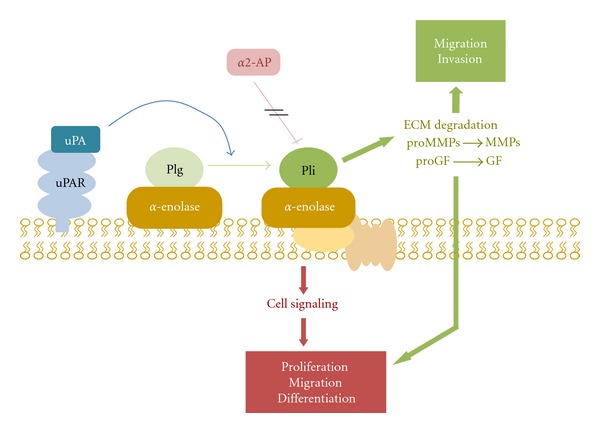
Schematic overview represents α-enolase/plasminogen interaction on the cell surface. α-Enolase enhances plasminogen activation on the cell surface, concentrates plasmin proteolytic activity on the pericellular area and protects plasmin from its inhibitor α2-antiplasmin. Once activated, plasmin can degrade most of the components of the extracellular matrix, directly or indirectly by activating metalloproteases. It is also capable to activate prohormones of progrowing factors. Abbreviations: Plg, plasminogen; Pli, plasmin, α2-AP, α2-antiplasmin; uPA, urokinase-type plasminogen activator, uPAR, urokinase-type plasminogen activator; ECM, extracellular matrix; MMPs, metalloproteases; GF, growing factors.
Considering the extraordinarily high number of plasminogen binding sites/cells that have been described in different cell types, no single surface protein can account for all plasminogen binding sites, suggesting that different receptors coexist on the cell surface [18]. Evidence from monocytoid cells suggested that α-enolase was only one of several plasminogen receptors and its contribution to plasmin activation was only modest [18, 54]. Posterior studies have emphasized the role of annexin A2 and histone H2B as plasminogen receptors in the same cells [46], suggesting a minor contribution of α-enolase as plasminogen receptor. In more recent studies, the role of α-enolase has been resurrected, showing a central role for α-enolase in monocyte recruitment in inflammatory lung disease [59]. These results imply that different plasminogen receptors could be targeted to regulate inflammatory cell recruitment in a temporal-specific manner.
The α-enolase-plasminogen interaction is mediated by binding of plasminogen kringle domains to the C-terminal residues of α-enolase (K434) [15, 18]. Furthermore, interaction of plasminogen lysine binding sites with α-enolase depends upon recognition of C-terminal lysines K420, K422, and K434, suggesting that amino acid residues upstream and/or secondary structure may be responsible for the high affinity of α-enolase for plasminogen [15, 18]. Another putative plasminogen-binding motif has been proposed in view of its crystal structure at position, 250FFRSGKY256, that remains exposed when α-enolase forms a dimer, necessary for its glycolytic activity [10]. Human α-enolase structure has been determined and it has been found that it exhibits specific surface properties that are distinct from those of other enolases despite high-sequence similarity. These differences in structure explain its various activities, including plasmin(ogen) and DNA binding [10].
The mechanism by which α-enolase, that lacks a signal sequence, is associated with the cell membrane remains unknown. Some authors have speculated that a hydrophobic domain within α-enolase might serve as an internal signal sequence [60], while others suggest that posttranslational acetylation [61] or phosphorylation [62] may control membrane association. Nevertheless, α-enolase forms part of a growing list of proteins that lack signal sequences, but are transported to the cell surface by a yet unknown mechanism.
4. α-Enolase in Myogenesis and Muscle Regeneration
Proteolysis associated with the cell surface is a usual mechanism in several physiological processes involving tissue remodeling. Myogenesis is an example of tissue remodeling in which massive extracellular matrix degradation takes place. Components of the PA system play important, yet distinct roles in muscle regeneration after injury. Using genetically modified mice for uPA and plasminogen, we and others have shown that loss of uPA-mediated plasmin activity blunts muscle repair in vivo [63–66]. In contrast, a negative role for PAI-1 in muscle regeneration was suggested [65]. The PA system has also been shown to have an increasingly important role in muscular dystrophies. For example, greater expression of uPA has been found in mdx muscle, the mouse model for Duchenne muscle dystrophy (DMD). Conversely, genetic loss of uPA exacerbated dystrophy and reduced muscle function in mdx mice [66]. Satellite cells derived from human DMD patients produce more uPAR and PAI-1 and less uPA than normal satellite cells [67]. uPA and plasmin appear to be required for infiltration of macrophages into the damaged or dystrophic muscle in mdx mice. However, an interesting observation underpinning these results was that genetic loss of the uPAR in mdx mice failed to exacerbate muscular dystrophy, suggesting that uPA exerts its proteolytic effects independently of its cell surface receptor uPAR [66].
β-enolase is considered the specific muscular enolase isoform, it is expressed in proliferating adult myoblasts as well as in differentiated myotubes [68]. It is upregulated in muscle during embryogenic development and it is considered an early marker of myogenesis [69]. The increase of the β-isoform is accompanied by a decrease of the α and γ isoform; the γ-isoform is completely absent in the adult muscle, but the expression of the α-isoform is maintained in the adult muscle and in muscular cells [70, 71]. Furthermore, we have described that α-enolase is upregulated in murine myoblasts C2C12 differentiation in vitro and in muscle regeneration in vivo [72], thus raising the question of whether plasminogen receptors may also function in myogenesis and skeletal regeneration as a mechanism for regulating plasmin activity.
We have investigated the role of α-enolase plasminogen receptor in muscle regeneration after injury, a process involving extensive cell infiltration and ECM remodeling. Injured wild-type mice and dystrophic mdx mice were treated with inhibitors of α-enolase/plasminogen binding: MAb11G1 (an inhibitory monoclonal antibody against α-enolase) and ε-aminocaproic acid (EACA, a lysine analogue). These treatments had negative impacts on muscle repair by impairing adequate inflammatory cell infiltration and promoting extracellular matrix deposition, which resulted in persistent degeneration. Furthermore, satellite cell-derived myoblasts (i.e., MPCs) expressed α-enolase on the cell surface, and this expression was upregulated during myogenic differentiation, correlating with an increase of plasminogen binding to the cell surface. We found that both MAb11G1 and EACA treatments impaired satellite cell-derived myoblasts functions in vitro in agreement with blunted growth of new myofibers in vivo (Diaz-Ramos et al., unpublished results).
Loss of uPAR in vivo did not affect the degeneration/regeneration process; in addition, cultured myoblasts from uPAR-deficient mice showed efficient myoblast differentiation and fusion [66, 73], indicating that uPAR is dispensable for efficient muscle repair. This reinforces the idea that α-enolase is the main functional plasminogen receptor during muscle tissue remodeling. Altogether, these results demonstrate the novel requirement of α-enolase for restoring homeostasis of injured muscle tissue, by concentrating plasmin activity on the cell surface of inflammatory and myogenic cells.
5. The α-Enolase Expression in Injured Cardiac Muscle
The actuation of the PA system in tissue healing after a cardiac failure, driving the degradation the ECM and scar tissue after an ischemic injury and allowing the inflammatory cell invasion, has been extensively demonstrated.
The regulation of α-enolase in cardiac tissue as regulator of glucose metabolism has been analyzed by several authors. A decrease of α-enolase expression in the aging heart of old male monkeys has been described, paralleling left ventricular dysfunction, and could be involved in the mechanism for the cardiomyopathy of aging [74]. α-Enolase expression has been identified as a strongly induced factor in response to ischemic hypoxia and reoxygenation in rat hearts subjected to ischemia-reperfusion [75]. Furthermore, α-Enolase improved the contractility of cardiomyocytes impaired by ischemic hypoxia [76]. α-Enolase has also been proposed as a marker for early diagnosis for acute myocardial infarction [77].
On the other hand, recent evidences indicates an involvement of proteinases, including the PAs and MMPs systems, in the process of extracellular matrix degradation and cell migration during cardiac wound healing [78]. In a recent study, Heymans et al. demonstrated that uPA-deficient mice showed impaired infarct healing and were completely protected against cardiac rupture after induction of a myocardial infarction [79]. Wound healing after infarct was abolished in plasminogen-deficient mice, indicating that the plasminogen system is required for the repair process of the heart after infarction. In the absence of plasminogen, inflammatory cells did not migrate into the infarcted myocardium, necrotic cardiomyocytes were not removed and there was no formation of granulation tissue and fibrous tissue [80]. Furthermore, PAI-1, which has been shown to be expressed in mammalian cardiomyocytes [81], has been implicated in the process of the cardiac remodeling by inhibiting activation of MMPs as well as plasmin generation. A dramatic induction of PAI-1 in a mouse model of infarct has been described [82]. Experiments using mice deficient in PAI-1 suggest that increased expression of cardiac PAI-1 may contribute to the development of fibrous change after acute myocardial infarction (AMI). In vivo studies also showed that PAI-1 expression was induced in hearts under pathological conditions as ventricular hypertrophy [83].
All these results demonstrate that the PA system plays a role in ECM remodeling after a cardiac injury and allows inflammatory cell invasion. Furthermore, it can also play a role in cardiomyocyte survival. Cardiomyocytes, which are terminally differentiated cells, cannot proliferate, even when they are damaged; the damage can lead to cell death in the case of serious diseases such as acute myocardial infarction and myocarditis [84]. Recent studies have identified myocyte apoptosis in the failing human heart [85, 86]. Plasminogen could also drive cardiomyocyte apoptosis, because plasmin induces cell detachment and apoptosis of smooth muscle cells through its binding to the cell surface, although the receptor responsable for plasminogen binding has not yet been identified [87].
Knowing that the PA system has been associated with cardiac remodeling, and that α-enolase is upregulated in cardiac infarction, it is tempting to speculate that α-enolase could act as plasminogen receptor, regulating PA activity on cardiac cells. Previous results from our laboratory have shown that plasmin activity is concentrated on the cell surface of cardiac fibroblasts in a lysine-dependent manner, and this binding capacity is increased by hypoxic conditions. Furthermore, plasminogen binding drives the activation of fibroblasts to myofibroblasts, the main cells responsible of tissue remodeling after a cardiac injury (Garcia-Melero et al., unpublished results).
6. α-Enolase/Plasmin Role in Apoptosis
It has been described that plasminogen binding to the cell surface and its further activation to plasmin induces cell detachment and apoptosis in smooth muscle cells, neurons and vascular myofibroblasts [88–90], although the molecular responsible for plasminogen interaction with the cell surface has not been identified.
Externalization of glycolytic enzymes is a common and early aspect of cell death in different cell types triggered to die with different suicidal stimuli [91]. Apoptotic cells are recognized by phagocytes and trigger an active immunosuppressive response. The lack of inflammation associated normally with the clearance of apoptotic cells has been linked to inflammatory and autoimmune disease as systemic lupus erythematosis and rheumatic diseases [92–95]. Regarding apoptotic cell surface proteins, a new concept has been defined, SUPER, referring to Surface-exposed (during apoptotic cell death), Ubiquitously expressed, Protease sensitive, Evolutionary-conserved, and Resident normally in viable cells (SUPER), to emphasize defining properties of apoptotic determinants for recognition and immune modulation. Ucker et al. have recently demonstrated that almost all members of the glycolytic pathway are enriched among apoptotic cell membranes, with α-enolase being the more abundant enzyme in the cell membrane, and considered the most paradigmatic SUPER protein [91]. In the cell membrane of apoptotic cells, α-enolase has lost its glycolytic activity, but it acts as plasminogen receptor, coinciding with the description of the association of plasminogen binding with apoptotic cell death [96]. In contrast to α-enolase, other molecular plasminogen receptors as annexin A2 [97] or H2B [46], were not preferentially enriched on the apoptotic cell surface.
7. α-Enolase in Cancer
Several reports have shown an upregulation of α-enolase in several types of cancer [98–100]. The role of α-enolase as a plasminogen receptor on cancer cells has been extensively documented, where it acts as a key protein, promoting cellular metabolism in anaerobic conditions, and driving tumor invasion through plasminogen activation and ECM degradation (reviewed in [101]).
Recently, an analysis of disease-specific gene network identified desmin, interleukin 8, and α-enolase as central elements for colon cancer tumorogenesis [102]. Knockdown of α-enolase expression in different tumor cell lines caused a dramatic increase in their sensitivity to microtubule targeted drugs (e.g., taxanes and vincristine), probably due to α-enolase-tubulin interactions [103], suggesting a role for α-enolase in modulating the microtubule network. Downregulation of α-enolase gene product deceased invasiveness of the follicular thyroid carcinoma cell lines [104]. α-Enolase overexpression has been associated with head and neck cancer cells, and this increase associated not only with cancer progression but also with poor clinical outcomes. Furthermore, exogenous α-enolase expression promoted cell proliferation, migration, invasion, and tumorogenesis [105].
During tumor formation and expansion, tumor cells must increase glucose metabolism [106]. Hypoxia is common feature of solid tumors. Consistent with this, overexpression of glycolytic genes has been found in a myriad of human cancers [107]. In tumor cells, α-enolase is upregulated and supports anaerobic proliferation (Warburg effect), and it is expressed on the cell surface, where it promotes cancer invasion. Thus, it seems that α-enolase is playing a pleitropic role on cancer cell progression. Furthermore, it has been demonstrated that α-enolase is upregulated in pancreatic ductal adenocarcinoma, where it is subjected to a array of posttranslational modifications, namely acetylation, methylation, and phosphorylation [108]. Both, α-enolase expression and posttranslational modifications could be of diagnostic and prognostic value in cancer (reviewed in [101]).
8. Posttranslational Modifications of α-Enolase
Posttranslational protein modifications, such as phosphorylation, acetylation, and methylation are common and important mechanisms of acute and reversible regulation of protein function in mammalian cells, and largely control cellular signaling events that orchestrate biological functions. Several posttranslational modifications have been described for α-enolase. α-Enolase phosphorylation has been associated with pancreatic cancer, and induces specific autoantibody production in pancreatic ductal adenocarcinoma patients with diagnostic value [109]. Lysine acetylated α-enolase has been detected in mouse brain [110]. Nitration of tyrosine residues in α-enolase has been detected in diabetic rat hearts, contributing to the impaired glycolytic activity in diabetic cardiomyopathy [111]. Phosphorylated α-enolase has been detected in gastrocnemious muscle, and phosphorylation decreased with age [112]. Furthermore, carbonylation of α-enolase has been detected on human myoblasts under oxidative stress [113].
It remains to be determined how the posttranslational modifications of α-enolase can affect its catalytic activity, localization of the cell, protein stability, and the ability to dimerize or form a complex with other molecules. Investigations of these modifications patterns in different pathologies will provide insights into its important role in pathophysiological processes.
9. α-Enolase in Rheumatoid Arthritis
The overexpression of α-enolase has also been found associated with chronic autoimmune diseases like rheumatoid arthritis [19, 114], systemic sclerosis [115], and primary nephropathies [116]. Autoantibodies to α-enolase, are present in the sera of patients with very early rheumatoid arthritis and have potential diagnostic and prognostic value [117]. Recently, citrullinated proteins have been considered the main autoantigen of rheumatoid arhritis. Citrullination, also termed deimination, is a modification of arginine side chains catalyzed by peptidylarginine deaminase. This posttranscriptional modification has the potential to alter the structure, antigenicity, and function of proteins. α-Enolase is abundantly expressed in the sinovial membrane, and antibodies against citrullinated α-enolase were specific for rheumatoid arthritis. Citrullination changes the conformation of α-enolase and interferes with the noncovalent interaction involved in the formation of the enolase dimer, then results in an altered glycolytic activity and plasminogen binding. It is likely that citrullination of cell-surface α-enolase abrogates its plasminogen binding and activating function and contributes to the decreased fibrinolysis observed in rheumatoid arthritis [118]. Curiously, other glycolytic enzymes such as glucose phosphate isomerase and aldolase also promote rheumatoid arthritis autoimmunity by acting as autoantigens [119].
10. α-Enolase in Alzheimer's Disease
Although γ-enolase is the specific neuronal enolase isoform, α-isoform is also present in neurological tissues. Plasmin formation enhanced by α-enolase has been proposed to enhance neuritogenesis [16, 120]. Furthermore, cathepsin X cleavage of C-terminal lysine of α-enolase impaired survival and neuritogenesis of neuronal cells [121]. α-Enolase has been reported as a strong plasminogen receptor within the brain; it is known to be upregulated in the Alzheimer's disease brain and has been proposed as a promising therapeutic target for this disease (reviewed in [122]). Glucose hypometabolism and upregulation of glycolytic enzymes is a predominant feature in Alzheimer's disease [123], but accumulating results suggest that α-enolase may have other functions that just metabolic processing of glucose: plasminogen bound to α-enolase stimulates plasmin activation of mitogen-activated protein kinase (MAPK)/extracellular-signal regulated kinase 1/2 (ERK1/2) prosurvival factor and also can drive plasmin degradation of amyloid-β (Aβ) protein, the main component of amyloid plaques. Thus, α-enolase might play a neuroprotective role through its multiple functions (reviewed in [122]).
Recently, several posttranslational modifications to α-enolase have been found in Alzheimer's disease. Elevated levels of glycosylated-α-enolase [124], oxidized [123], or glutathionylated [125] have been found related to Alzheimer's disease. These modifications would render enolase catallyticaly inactive, related to the metabolic deficit associated to Alzheimer's disease. The effect of these modifications in other multiple functions of α-enolase is a subject of ongoing experiments, but it is possible that α-enolase modifications alter not only glucose metabolism, but also its role as plasminogen receptor, controlling neuronal survival and Aβ degradation.
11. Plasmin and Intracellular Signaling
Other than its role in concentrating proteolytic activity on the cell surface, several recent studies have shown that plasmin is able to activate several intracellular signaling pathways, that led to the activation of several transcription factors, in a cell surface binding dependent way. In most of the cases, the molecular mechanism responsible remains unknown: it could be due to the proteolytic activation of a second factor or due to direct binding of plasmin(ogen) to a specific receptor. Several pieces of work show that the plasmin proteolytic activity is essential for the induction of an intracellular response, as in monocytes, where plasmin bound to the cell surface proteolytically activates annexin A2 and stimulates MMP-1 production through the activation of ERK and p38 pathways [126]. The phosphorylation of Janus Kinase 1 (JAK1)/Tyrosin Kinase 2 (TYK2) that drives to the activation to the transcription factors AP-1 and Nuclear Factor κB (NFκB), and the expression of several cytokines: interleukin-1α and-1β (IL-1α and IL-1β), tissue factor (TF), and the Tumoral Necrosis Factor-α (TNF-α), are a consequence of plasmin interaction with the cell surface [127–129]. Plasmin promotes p38 and p44/42 MAPK activation and fibroblast proliferation through Protease Activated Receptor-1 (PAR-1) [130, 131]. Other authors have described that plasminogen and plasmin regulate the gene transcription of genes as c-fos, erg-1, and Eno1 in mononucleated blood cells and fibroblasts, by activating the MEK/ERK pathways [132, 133].
In most of the cases, the receptor responsible for this cellular response remains to be identified. Most of the protein candidates for plasminogen receptors are small proteins that lack a transmembrane domain and are not able to induce directly an intracellular response. Some work suggests an association between the plasminogen receptor and other membrane proteins, that could serve as molecular collaborators to induce the activation of intracellular signaling pathways. Several proteins have been identified as such molecular collaborators. For instance, plasmin can activate PAR-1 in fibroblasts, by the phosphorylation of Erk [130]; plasminogen and plasmin activate the expression of several genes in fibroblasts and monocytes through G-Protein Coupled Receptors, (GPCR) [132, 133]; some integrins such as α9β1 integrin in Chinese Hamster Ovary (CHO) cells [134] and αvβ3 integrin, in vascular endothelial cells [135], participate actively in plasmin-induced cell migration.
In none of these cases, the plasmin receptor associated with these proteins have been identified. Some work have identified annexin A2 as the receptor that concentrates plasmin activity to the cell surface and drives a subsequent intracellular response [127–129]. Other authors have described a collaboration between α-enolase and GPCR in fibroblasts and mononucleated blood cells [132, 133]. Plasmin induces smooth muscle cell proliferation through extracellular transactivation of the epidermal growth factor receptor (EGFR) by a MMP-mediated, heparin binding—epidermal growth factor (HB-EGF-) dependent process [136]. Future studies will be necessary to determine the molecular mechanism of the plasminogen receptor on several cell types and the putative proteins associated with it.
We have shown that plasmin activity is able to activate MAPK/ERK and phosphatidyl-inositol 3-kinase (PI3K)/Akt pathways in C2C12 murine myoblast cell lines and in primary cultures of muscle precursor cells, and that intracellular activation depends on plasmin activity, but also on plasmin(ogen) binding to the cell surface in a lysine binding sites dependent way (Roig-Borrellas et al., unpublished results), although the receptor responsible and the molecular mechanism remains to be elucidated.
12. Concluding Remarks
Recently, a proteomic meta-analysis of 169 published articles, including differently expressed 4700 proteins, based on 2-dimensional electrophoresis analysis of human, mouse, and rat tissues, identified α-enolase as the first protein differentially expressed in mice and the second in human pathologies, regardless of the tissue used and experiment performed [137], suggesting that α-enolase could be part of a group of universal cellular sensors that respond to multiple different stimuli. Thus, α-enolase could be considered as a marker of pathological stress in a high number of diseases. The importance of α-enolase as plasminogen receptor has been determined in several pathologies such as cancer, skeletal myogenesis, Alzheimer's disease, and rheumatoid arthritis, among others. α-Enolase upregulation has also been described in a myriad of other pathologies, as inflammatory bowel disease [138, 139], autoimmune hepatitis [140], or membranous glomerulonephritis [141], not discussed in this paper, although its role on concentrating plasmin activity on the cell surface has not always been stablished. It will not be surprising that in many of these pathologies, α-enolase could exert one of its multiple functions, mainly as a plasminogen receptor, focalizing plasmin activity on the cell membrane and promoting ECM degradation/remodeling, but also activating intracellular survival pathways and controlling survival/apoptosis of cells.
Further studies of posttranslational modifications of α-enolase and its implications on α-enolase subcellular distribution and function, especially interaction with other proteins will be necessary. Also, the role of α-enolase as activator of intracellular signaling pathways, probably in collaboration with other membrane proteins, will serve to elucidate the multiples roles of this functionally complex protein.
Unexpectedly, other glycolytic enzymes have been described as having other nonglycolytic functions in transcriptional regulation (hexokinase-2, HK; lactate dehydrogenase A, LDH; glyceraldehydes-3-phosphate dehydrogenase, GAPDH), stimulation of cell motility (glucose-6-phosphate isomerase), and regulation of apoptosis (glucokinase, HK and GAPDH), indicating that they are more complicated, multifunctional proteins rather than simply components of the glycolytic pathway (reviewed in [142]).
Some of the more interesting and challenging issues, regarding α-enolase multifunction that need to be addressed are (i) the mechanism of its export to the cell surface, (ii) the role of α-enolase as an inductor of intracellular signaling pathways, and (iii) the role of posttranslational modifications of α-enolase and implications on its subcellular distribution and function. Investigations of these subjects in different human pathologies will provide insights into its important role on pathophysiological processes and it would make this protein an interesting drug target for different diseases.
Acknowledgments
This study was supported by the Ministerio de Ciencia e Innovación, Spain (SAF2004-04717, SAF2007-63596) and Association Française contre les Myopathies, France (AFM 9869). A. Díaz-Ramos was supported by an AFM predoctoral fellowship. A. Roig-Borrellas was supported by a F.P.U. Fellowship. A. García-Melero was supported by a Marató-TV3 Foundation fellow. A. Roig-Borrellas is a registered student in the PhD Doctorate Program in Biochemistry, Molecular Biology and Biomedicine of the Universitat Autònoma de Barcelona (UAB), Spain.
References
- 1.Pancholi V. Multifunctional α-enolase: its role in diseases. Cellular and Molecular Life Sciences. 2001;58(7):902–920. doi: 10.1007/PL00000910. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Piast M, Kustrzeba-Wójcicka I, Matusiewicz M, Banaś T. Molecular evolution of enolase. Acta Biochimica Polonica. 2005;52(2):507–513. [PubMed] [Google Scholar]
- 3.Marangos PJ, Parma AM, Goodwin FK. Functional properties of neuronal and glial isoenzymes of brain enolase. Journal of Neurochemistry. 1978;31(3):727–732. doi: 10.1111/j.1471-4159.1978.tb07847.x. [DOI] [PubMed] [Google Scholar]
- 4.Feo S, Oliva D, Barbieri G, Xu W, Fried M, Giallongo A. The gene for the muscle-specific enolase is on the short arm of human chromosome 17. Genomics. 1990;6(1):192–194. doi: 10.1016/0888-7543(90)90467-9. [DOI] [PubMed] [Google Scholar]
- 5.Giallongo A, Feo S, Moore R, Croce CM, Showe LC. Molecular cloning and nucleotide sequence of a full-length cDNA for human α enolase. Proceedings of the National Academy of Sciences of the United States of America. 1986;83(18):6741–6745. doi: 10.1073/pnas.83.18.6741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Fletcher L, Rider CC, Taylor CB. Enolase isoenzymes. III. Chromatographic and immunological characteristics of rat brain enolase. Biochimica et Biophysica Acta. 1976;452(1):245–252. doi: 10.1016/0005-2744(76)90077-2. [DOI] [PubMed] [Google Scholar]
- 7.Kato K, Okagawa Y, Suzuki F, Shimizu A, Mokuno K, Takahashi Y. Immunoassay of human muscle enolase subunit in serum: a novel marker antigen for muscle diseases. Clinica Chimica Acta. 1983;131(1-2):75–85. doi: 10.1016/0009-8981(83)90354-6. [DOI] [PubMed] [Google Scholar]
- 8.Lebioda L, Stec B. Crystal structure of enolase indicates that enolase and pyruvate kinase evolved from a common ancestor. Nature. 1988;333(6174):683–686. doi: 10.1038/333683a0. [DOI] [PubMed] [Google Scholar]
- 9.Zhang E, Brewer JM, Minor W, Carreira LA, Lebioda L. Mechanism of enolase: the crystal structure of asymmetric dimer enolase- 2-phospho-D-glycerate/enolase-phosphoenolpyruvate at 2.0 A resolution. Biochemistry. 1997;36(41):12526–12534. doi: 10.1021/bi9712450. [DOI] [PubMed] [Google Scholar]
- 10.Kang HJ, Jung SK, Kim SJ, Chung SJ. Structure of human α-enolase (hENO1), a multifunctional glycolytic enzyme. Acta Crystallographica D. 2008;64(6):651–657. doi: 10.1107/S0907444908008561. [DOI] [PubMed] [Google Scholar]
- 11.McAlister L, Holland MJ. Targeted deletion of a yeast enolase structural gene. Identification and isolation of yeast enolase isozymes. Journal of Biological Chemistry. 1982;257(12):7181–7188. [PubMed] [Google Scholar]
- 12.Holland JP, Labieniec L, Swimmer C, Holland MJ. Homologous nucleotide sequences at the 5’ termini of messenger RNAs synthesized from the yeast enolase and glyceraldehyde-3-phosphate dehydrogenase gene families. The primary structure of a third yeast glyceraldehyde-3-phosphate dehydrogenase gene. Journal of Biological Chemistry. 1983;258(8):5291–5299. [PubMed] [Google Scholar]
- 13.Giallongo A, Oliva D, Calì L, Barba G, Barbieri G, Feo S. Structure of the human gene for alpha-enolase. European Journal of Biochemistry. 1990;190(3):567–573. doi: 10.1111/j.1432-1033.1990.tb15611.x. [DOI] [PubMed] [Google Scholar]
- 14.Feo S, Arcuri D, Piddini E, Passantino R, Giallongo A. ENO1 gene product binds to the c-myc promoter and acts as a transcriptional repressor: relationship with Myc promoter-binding protein 1 (MBP-1) FEBS Letters. 2000;473(1):47–52. doi: 10.1016/s0014-5793(00)01494-0. [DOI] [PubMed] [Google Scholar]
- 15.Miles LA, Dahlberg CM, Plescia J, Felez J, Kato K, Plow EF. Role of cell-surface lysines in plasminogen binding to cells: identification of α-enolase as a candidate plasminogen receptor. Biochemistry. 1991;30(6):1682–1691. doi: 10.1021/bi00220a034. [DOI] [PubMed] [Google Scholar]
- 16.Nakajima K, Hamanoue M, Takemoto N, Hattori T, Kato K, Kohsaka S. Plasminogen binds specifically to α-enolase on rat neuronal plasma membrane. Journal of Neurochemistry. 1994;63(6):2048–2057. doi: 10.1046/j.1471-4159.1994.63062048.x. [DOI] [PubMed] [Google Scholar]
- 17.Dudani AK, Cummings C, Hashemi S, Ganz PR. Isolation of a novel 45 kDa plasminogen receptor from human endothelial cells. Thrombosis Research. 1993;69(2):185–196. doi: 10.1016/0049-3848(93)90044-o. [DOI] [PubMed] [Google Scholar]
- 18.Redlitz A, Fowler BJ, Plow EF, Miles LA. The role of an enolase-related molecule in plasminogen binding to cells. European Journal of Biochemistry. 1995;227(1-2):407–415. doi: 10.1111/j.1432-1033.1995.tb20403.x. [DOI] [PubMed] [Google Scholar]
- 19.Fontan PA, Pancholi V, Nociari MM, Fischetti VA. Antibodies to streptococcal surface enolase react with human α-enolase: implications in poststreptococcal sequelae. Journal of Infectious Diseases. 2000;182(6):1712–1721. doi: 10.1086/317604. [DOI] [PubMed] [Google Scholar]
- 20.Takei N, Kondo J, Nagaike K, Ohsawa K, Kato K, Kohsaka S. Neuronal survival factor from bovine brain is identical to neuron-specific enolase. Journal of Neurochemistry. 1991;57(4):1178–1184. doi: 10.1111/j.1471-4159.1991.tb08277.x. [DOI] [PubMed] [Google Scholar]
- 21.Iida H, Yahara I. Yeast heat-shock protein of M(r) 48,000 is an isoprotein of enolase. Nature. 1985;315(6021):688–690. [Google Scholar]
- 22.Aaronson RM, Graven KK, Tucci M, McDonald RJ, Farber HW. Non-neuronal enolase is an endothelial hypoxic stress protein. Journal of Biological Chemistry. 1995;270(46):27752–27757. doi: 10.1074/jbc.270.46.27752. [DOI] [PubMed] [Google Scholar]
- 23.Mathur RL, Reddy MC, Yee S, Imbesi R, Groth-Vasselli B, Farnsworth PN. Investigation of lens glycolytic enzymes: species distribution and interaction with supramolecular order. Experimental Eye Research. 1992;54(2):253–260. doi: 10.1016/s0014-4835(05)80215-5. [DOI] [PubMed] [Google Scholar]
- 24.Walsh JL, Keith TJ, Knull HR. Glycolytic enzyme interactions with tubulin and microtubules. Biochimica et Biophysica Acta. 1989;999(1):64–70. doi: 10.1016/0167-4838(89)90031-9. [DOI] [PubMed] [Google Scholar]
- 25.Johnstone SA, Waisman DM, Rattner JB. Enolase is present at the centrosome of HeLa cells. Experimental Cell Research. 1992;202(2):458–463. doi: 10.1016/0014-4827(92)90099-t. [DOI] [PubMed] [Google Scholar]
- 26.Merkulova T, Lucas M, Jabet C, et al. Biochemical characterization of the mouse muscle-specific enolase: developmental changes in electrophoretic variants and selective binding to other proteins. Biochemical Journal. 1997;323(3):791–800. doi: 10.1042/bj3230791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Subramanian A, Miller DM. Structural analysis of α-enolase: mapping the functional domains involved in down-regulation of the c-myc protooncogene. Journal of Biological Chemistry. 2000;275(8):5958–5965. doi: 10.1074/jbc.275.8.5958. [DOI] [PubMed] [Google Scholar]
- 28.Eriksson B, Öberg K, Stridsberg M. Tumor markers in neuroendocrine tumors. Digestion. 2000;62(supplement 1):33–38. doi: 10.1159/000051853. [DOI] [PubMed] [Google Scholar]
- 29.Niklinski J, Furman M. Clinical tumour markers in lung cancer. European Journal of Cancer Prevention. 1995;4(2):129–138. doi: 10.1097/00008469-199504000-00002. [DOI] [PubMed] [Google Scholar]
- 30.Cooper EH. Neuron-specific enolase. International Journal of Biological Markers. 1994;9(4):205–210. doi: 10.1177/172460089400900401. [DOI] [PubMed] [Google Scholar]
- 31.Ledermann JA. Serum neurone-specific enolase and other neuroendocrine markers in lung cancer. European Journal of Cancer A. 1994;30(5):574–576. doi: 10.1016/0959-8049(94)90519-3. [DOI] [PubMed] [Google Scholar]
- 32.Takashima M, Kuramitsu Y, Yokoyama Y, et al. Overexpression of alpha enolase in hepatitis C virus-related hepatocellular carcinoma: association with tumor progression as determined by proteomic analysis. Proteomics. 2005;5(6):1686–1692. doi: 10.1002/pmic.200401022. [DOI] [PubMed] [Google Scholar]
- 33.Jeffery CJ. Moonlighting proteins. Trends in Biochemical Sciences. 1999;24(1):8–11. doi: 10.1016/s0968-0004(98)01335-8. [DOI] [PubMed] [Google Scholar]
- 34.Piatigorsky J. Multifunctional lens crystallins and corneal enzymes: more than meets the eye. Annals of the New York Academy of Sciences. 1998;842:7–15. doi: 10.1111/j.1749-6632.1998.tb09626.x. [DOI] [PubMed] [Google Scholar]
- 35.Irigoyen JP, Muñoz-Cánoves P, Montero L, Koziczak M, Nagamine Y. The plasminogen activator system: biology and regulation. Cellular and Molecular Life Sciences. 1999;56(1-2):104–132. doi: 10.1007/PL00000615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Miles LA, Plow EF. Binding and activation of plasminogen on the platelet surface. Journal of Biological Chemistry. 1985;260(7):4303–4311. [PubMed] [Google Scholar]
- 37.Hajjar KA, Harpel PC, Jaffe EA, Nachman RL. Binding of plasminogen to cultured human endothelial cells. Journal of Biological Chemistry. 1986;261(25):11656–11662. [PubMed] [Google Scholar]
- 38.Plow EF, Freaney DE, Plescia J, Miles LA. The plasminogen system and cell surfaces: evidence for plasminogen and urokinase receptors on the same cell type. Journal of Cell Biology. 1986;103(6):2411–2420. doi: 10.1083/jcb.103.6.2411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Miles LA, Plow EF. Receptor mediated binding of the fibrinolytic components, plasminogen and urokinase, to peripheral blood cells. Thrombosis and Haemostasis. 1987;58(3):936–942. [PubMed] [Google Scholar]
- 40.Blasi F, Carmeliet P. uPAR: a versatile signalling orchestrator. Nature Reviews Molecular Cell Biology. 2002;3(12):932–943. doi: 10.1038/nrm977. [DOI] [PubMed] [Google Scholar]
- 41.Ossowski L, Aguirre-Ghiso JA. Urokinase receptor and integrin partnership: coordination of signaling for cell adhesion, migration and growth. Current Opinion in Cell Biology. 2000;12(5):613–620. doi: 10.1016/s0955-0674(00)00140-x. [DOI] [PubMed] [Google Scholar]
- 42.Lopez-Alemany R, Correc P, Camoin L, Burtin P. Purification of the plasmin receptor from human carcinoma cells and comparison to α-enolase. Thrombosis Research. 1994;75(4):371–381. doi: 10.1016/0049-3848(94)90252-6. [DOI] [PubMed] [Google Scholar]
- 43.Hajjar KA, Jacovina AT, Chacko J. An endothelial cell receptor for plasminogen/tissue plasminogen activator. I. Identity with annexin II. Journal of Biological Chemistry. 1994;269(33):21191–21197. [PubMed] [Google Scholar]
- 44.Kassam G, Le BH, Choi KS, et al. The p11 subunit of the annexin II tetramer plays a key role in the stimulation of t-PA-dependent plasminogen activation. Biochemistry. 1998;37(48):16958–16966. doi: 10.1021/bi981713l. [DOI] [PubMed] [Google Scholar]
- 45.Herren T, Burke TA, Das R, Plow EF. Identification of histone H2B as a regulated plasminogen receptor. Biochemistry. 2006;45(31):9463–9474. doi: 10.1021/bi060756w. [DOI] [PubMed] [Google Scholar]
- 46.Das R, Burke T, Plow EF. Histone H2B as a functionally important plasminogen receptor on macrophages. Blood. 2007;110(10):3763–3772. doi: 10.1182/blood-2007-03-079392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Dudani AK, Ganz PR. Endothelial cell surface actin serves as a binding site for plasminogen, tissue plasminogen activator and lipoprotein(a) British Journal of Haematology. 1996;95(1):168–178. doi: 10.1046/j.1365-2141.1996.7482367.x. [DOI] [PubMed] [Google Scholar]
- 48.Kanalas JJ, Makker SP. Identification of the rat Heymann nephritis autoantigen (GP330) as a receptor site for plasminogen. Journal of Biological Chemistry. 1991;266(17):10825–10829. [PubMed] [Google Scholar]
- 49.Hembrough TA, Li L, Gonias SL. Cell-surface cytokeratin 8 is the major plasminogen receptor on breast cancer cells and is required for the accelerated activation of cell- associated plasminogen by tissue-type plasminogen activator. Journal of Biological Chemistry. 1996;271(41):25684–25691. doi: 10.1074/jbc.271.41.25684. [DOI] [PubMed] [Google Scholar]
- 50.Borza DB, Morgan WT. Acceleration of plasminogen activation by tissue plasminogen activator on surface-bound histidine-proline-rich glycoprotein. Journal of Biological Chemistry. 1997;272(9):5718–5726. doi: 10.1074/jbc.272.9.5718. [DOI] [PubMed] [Google Scholar]
- 51.Winram SB, Lottenberg R. The plasmin-binding protein Plr of group A streptococci is identified as glyceraldehyde-3-phosphate dehydrogenase. Microbiology. 1996;142(8):2311–2320. doi: 10.1099/13500872-142-8-2311. [DOI] [PubMed] [Google Scholar]
- 52.Andronicos NM, Chen EI, Baik N, et al. Proteomics-based discovery of a novel, structurally unique, and developmentally regulated plasminogen receptor, Plg-RKT, a major regulator of cell surface plasminogen activation. Blood. 2010;115(7):1319–1330. doi: 10.1182/blood-2008-11-188938. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Felez J. Plasminogen binding to cell surfaces. Fibrinolysis and Proteolysis. 1998;12(4):183–189. [Google Scholar]
- 54.López-Alemany R, Longstaff C, Hawley S, et al. Inhibition of cell surface mediated plasminogen activation by a monoclonal antibody against α-enolase. American Journal of Hematology. 2003;72(4):234–242. doi: 10.1002/ajh.10299. [DOI] [PubMed] [Google Scholar]
- 55.Longstaff C, Merton RE, Fabregas P, Felez J. Characterization of cell-associated plasminogen activation catalyzed by urokinase-type plasminogen activator, but independent of urokinase receptor (uPAR, CD87) Blood. 1999;93(11):3839–3846. [PubMed] [Google Scholar]
- 56.Sinniger V, Mertont RE, Fabregas P, Felez J, Longstaff C. Regulation of tissue plasminogen activator activity by cells: domains responsible for binding and mechanism of stimulation. Journal of Biological Chemistry. 1999;274(18):12414–12422. doi: 10.1074/jbc.274.18.12414. [DOI] [PubMed] [Google Scholar]
- 57.Félez J, Miles LA, Fábregas P, Jardí M, Plow EF, Lijnen RH. Characterization of cellular binding sites and interactive regions within reactants required for enhancement of plasminogen activation by tPA on the surface of leukocytic cells. Thrombosis and Haemostasis. 1996;76(4):577–584. [PubMed] [Google Scholar]
- 58.Miles LA, Fless GM, Scanu AM, et al. Interaction of Lp(a) with plasminogen binding sites on cells. Thrombosis and Haemostasis. 1995;73(3):458–465. [PubMed] [Google Scholar]
- 59.Wygrecka M, Marsh LM, Morty RE, et al. Enolase-1 promotes plasminogen-mediated recruitment of monocytes to the acutely inflamed lung. Blood. 2009;113(22):5588–5598. doi: 10.1182/blood-2008-08-170837. [DOI] [PubMed] [Google Scholar]
- 60.von Heijne G, Liljestrom P, Mikus P, Andersson H, Ny T. The efficiency of the uncleaved secretion signal in the plasminogen activator inhibitor type 2 protein can be enhanced by point mutations that increase its hydrophobicity. Journal of Biological Chemistry. 1991;266(23):15240–15243. [PubMed] [Google Scholar]
- 61.Bottalico LA, Kendrick NC, Keller A, Li Y, Tabas I. Cholesteryl ester loading of mouse peritoneal macrophages is associated with changes in the expression or modification of specific cellular proteins, including increase in an α-enolase isoform. Arteriosclerosis and Thrombosis. 1993;13(2):264–275. doi: 10.1161/01.atv.13.2.264. [DOI] [PubMed] [Google Scholar]
- 62.Cooper JA, Esch FS, Taylor SS, Hunter T. Phosphorylation sites in enolase and lactate dehydrogenase utilized by tyrosine protein kinase in vivo and in vitro. Journal of Biological Chemistry. 1984;259(12):7835–7841. [PubMed] [Google Scholar]
- 63.Lluís F, Roma J, Suelves M, et al. Urokinase-dependent plasminogen activation is required for efficient skeletal muscle regeneration in vivo. Blood. 2001;97(6):1703–1711. doi: 10.1182/blood.v97.6.1703. [DOI] [PubMed] [Google Scholar]
- 64.Suelves M, López-Alemany R, Lluís F, et al. Plasmin activity is required for myogenesis in vitro and skeletal muscle regeneration in vivo. Blood. 2002;99(8):2835–2844. doi: 10.1182/blood.v99.8.2835. [DOI] [PubMed] [Google Scholar]
- 65.Suelves M, Vidal B, Ruiz V, et al. The plasminogen activation system in skeletal muscle regeneration: antagonistic roles of Urokinase-type Plasminogen Activator (UPA) and its inhibitor (PAI-1) Frontiers in Bioscience. 2005;10(3):2978–2985. doi: 10.2741/1754. [DOI] [PubMed] [Google Scholar]
- 66.Suelves M, Vidal B, Serrano AL, et al. uPA deficiency exacerbates muscular dystrophy in MDX mice. Journal of Cell Biology. 2007;178(6):1039–1051. doi: 10.1083/jcb.200705127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Fibbi G, Barletta E, Dini G, et al. Cell invasion is affected by differential expression of urokinase plasminogen activator/urokinase plasminogen activator receptor system in muscle satellite cells from normal and dystrophic patients. Laboratory Investigation. 2001;81(1):27–39. doi: 10.1038/labinvest.3780209. [DOI] [PubMed] [Google Scholar]
- 68.Taylor JM, Davies JD, Peterson CA. Regulation of the myoblast-specific expression of the human β-enolase gene. Journal of Biological Chemistry. 1995;270(6):2535–2540. doi: 10.1074/jbc.270.6.2535. [DOI] [PubMed] [Google Scholar]
- 69.Fougerousse F, Edom-Vovard F, Merkulova T, et al. The muscle-specific enolase is an early marker of human myogenesis. Journal of Muscle Research and Cell Motility. 2001;22(6):535–544. doi: 10.1023/a:1015008208007. [DOI] [PubMed] [Google Scholar]
- 70.Keller A, Peltzer J, Carpentier G, et al. Interactions of enolase isoforms with tubulin and microtubules during myogenesis. Biochimica et Biophysica Acta. 2007;1770(6):919–926. doi: 10.1016/j.bbagen.2007.01.015. [DOI] [PubMed] [Google Scholar]
- 71.Merkulova T, Dehaupas M, Nevers MC, Créminon C, Alameddine H, Keller A. Differential modulation of α, β and γ enolase isoforms in regenerating mouse skeletal muscle. European Journal of Biochemistry. 2000;267(12):3735–3743. doi: 10.1046/j.1432-1327.2000.01408.x. [DOI] [PubMed] [Google Scholar]
- 72.López-Alemany R, Suelves M, Muñoz-Cánoves P. Plasmin generation dependent on α-enolase-type plasminogen receptor is required for myogenesis. Thrombosis and Haemostasis. 2003;90(4):724–733. doi: 10.1160/TH03-04-0291. [DOI] [PubMed] [Google Scholar]
- 73.Bryer SC, Koh TJ. The urokinase-type plasminogen activator receptor is not required for skeletal muscle inflammation or regeneration. American Journal of Physiology. 2007;293(3):R1152–R1158. doi: 10.1152/ajpregu.00132.2007. [DOI] [PubMed] [Google Scholar]
- 74.Yan L, Ge H, Li H, et al. Gender-specific proteomic alterations in glycolytic and mitochondrial pathways in aging monkey hearts. Journal of Molecular and Cellular Cardiology. 2004;37(5):921–929. doi: 10.1016/j.yjmcc.2004.06.012. [DOI] [PubMed] [Google Scholar]
- 75.Zhu LA, Fang NY, Gao PJ, Jin X, Wang HY. Differential expression of α-enolase in the normal and pathological cardiac growth. Experimental and Molecular Pathology. 2009;87(1):27–31. doi: 10.1016/j.yexmp.2009.05.002. [DOI] [PubMed] [Google Scholar]
- 76.Mizukami Y, Iwamatsu A, Aki T, et al. ERK1/2 regulates intracellular ATP levels through α-enolase expression in cardiomyocytes exposed to ischemic hypoxia and reoxygenation. Journal of Biological Chemistry. 2004;279(48):50120–50131. doi: 10.1074/jbc.M402299200. [DOI] [PubMed] [Google Scholar]
- 77.Mair J. Progress in myocardial damage detection: new biochemical markers for clinicians. Critical Reviews in Clinical Laboratory Sciences. 1997;34(1):1–66. doi: 10.3109/10408369709038215. [DOI] [PubMed] [Google Scholar]
- 78.Kim HE, Dalal SS, Young E, Legato MJ, Weisfeldt ML, D’Armiento J. Disruption of the myocardial extracellular matrix leads to cardiac dysfunction. Journal of Clinical Investigation. 2000;106(7):857–866. doi: 10.1172/JCI8040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Heymans S, Luttun A, Nuyens D, et al. Inhibition of plasminogen activators or matrix metalloproteinases prevents cardiac rupture but impairs therapeutic angiogenesis and causes cardiac failure. Nature Medicine. 1999;5(10):1135–1142. doi: 10.1038/13459. [DOI] [PubMed] [Google Scholar]
- 80.Creemers E, Cleutjens J, Smits J, et al. Disruption of the plasminogen gene in mice abolishes wound healing after myocardial infarction. American Journal of Pathology. 2000;156(6):1865–1873. doi: 10.1016/S0002-9440(10)65060-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Macfelda K, Weiss TW, Kaun C, et al. Plasminogen activator inhibitor 1 expression is regulated by the inflammatory mediators interleukin-1α, tumor necrosis factor-α, transforming growth factor-β and oncostatin M in human cardiac myocytes. Journal of Molecular and Cellular Cardiology. 2002;34(12):1681–1691. doi: 10.1006/jmcc.2002.2117. [DOI] [PubMed] [Google Scholar]
- 82.Takeshita K, Hayashi M, Iino S, et al. Increased expression of plasminogen activator inhibitor-1 in cardiomyocytes contributes to cardiac fibrosis after myocardial infarction. American Journal of Pathology. 2004;164(2):449–456. doi: 10.1016/S0002-9440(10)63135-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Carroll SM, Nimmo LE, Knoepfler PS, White FC, Bloor CM. Gene expression in a swine model of right ventricular hypertrophy: intercellular adhesion molecule, vascular endothelial growth factor and plasminogen activators are upregulated during pressure overload. Journal of Molecular and Cellular Cardiology. 1995;27(7):1427–1441. doi: 10.1006/jmcc.1995.0135. [DOI] [PubMed] [Google Scholar]
- 84.Pasumarthi KBS, Field LJ. Cardiomyocyte cell cycle regulation. Circulation Research. 2002;90(10):1044–1054. doi: 10.1161/01.res.0000020201.44772.67. [DOI] [PubMed] [Google Scholar]
- 85.Olivetti G, Abbi R, Quaini F, et al. Apoptosis in the failing human heart. The New England Journal of Medicine. 1997;336(16):1131–1141. doi: 10.1056/NEJM199704173361603. [DOI] [PubMed] [Google Scholar]
- 86.Guerra S, Leri A, Wang X, et al. Myocyte death in the failing human heart is gender dependent. Circulation Research. 1999;85(9):856–866. doi: 10.1161/01.res.85.9.856. [DOI] [PubMed] [Google Scholar]
- 87.Ho-Tin-Noé B, Rojas G, Vranckx R, Lijnen HR, Anglés-Cano E. Functional hierarchy of plasminogen kringles 1 and 4 in fibrinolysis and plasmin-induced cell detachment and apoptosis. FEBS Journal. 2005;272(13):3387–3400. doi: 10.1111/j.1742-4658.2005.04754.x. [DOI] [PubMed] [Google Scholar]
- 88.Meilhac O, Ho-Tin-Noé B, Houard X, Philippe M, Michel JB, Anglés-Cano E. Pericellular plasmin induces smooth muscle cell anoikis. The FASEB Journal. 2003;17(10):1301–1303. doi: 10.1096/fj.02-0687fje. [DOI] [PubMed] [Google Scholar]
- 89.Ho-Tin-Noé B, Enslen H, Doeuvre L, Corsi JM, Lijnen HR, Anglés-Cano E. Role of plasminogen activation in neuronal organization and survival. Molecular and Cellular Neuroscience. 2009;42(4):288–295. doi: 10.1016/j.mcn.2009.08.001. [DOI] [PubMed] [Google Scholar]
- 90.Kochtebane N, Choqueux C, Passefort S, et al. Plasmin induces apoptosis of aortic valvular myofibroblasts. Journal of Pathology. 2010;221(1):37–48. doi: 10.1002/path.2681. [DOI] [PubMed] [Google Scholar]
- 91.Ucker DS, Jain MR, Pattabiraman G, Palasiewicz K, Birge RB, Li H. Externalized glycolytic enzymes are novel, conserved, and early biomarkers of apoptosis. Journal of Biological Chemistry. 2012;287:10625–10343. doi: 10.1074/jbc.M111.314971. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Herrmann M, Voll RE, Zoller OM, Hagenhofer M, Ponner BB, Kalden JR. Impaired phagocytosis of apoptotic cell material by monocyte-derived macrophages from patients with systemic lupus erythematosus. Arthritis and Rheumatism. 1998;41:1241–1250. doi: 10.1002/1529-0131(199807)41:7<1241::AID-ART15>3.0.CO;2-H. [DOI] [PubMed] [Google Scholar]
- 93.Cohen PL, Caricchio R, Abraham V, et al. Delayed apoptotic cell clearance and lupus-like autoimmunity in mice lacking the c-mer membrane tyrosine kinase. Journal of Experimental Medicine. 2002;196(1):135–140. doi: 10.1084/jem.20012094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Rothlin CV, Ghosh S, Zuniga EI, Oldstone MBA, Lemke G. TAM receptors are pleiotropic inhibitors of the innate immune response. Cell. 2007;131(6):1124–1136. doi: 10.1016/j.cell.2007.10.034. [DOI] [PubMed] [Google Scholar]
- 95.Thorp EB. Mechanisms of failed apoptotic cell clearance by phagocyte subsets in cardiovascular disease. Apoptosis. 2010;15(9):1124–1136. doi: 10.1007/s10495-010-0516-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.O’Mullane MJ, Baker MS. Loss of cell viability dramatically elevates cell surface plasminogen binding and activation. Experimental Cell Research. 1998;242(1):153–164. doi: 10.1006/excr.1998.4067. [DOI] [PubMed] [Google Scholar]
- 97.Cesarman GM, Guevara CA, Hajjar KA. An endothelial cell receptor for plasminogen/tissue plasminogen activator (t-PA). II. Annexin II-mediated enhancement of t-PA-dependent plasminogen activation. Journal of Biological Chemistry. 1994;269(33):21198–21203. [PubMed] [Google Scholar]
- 98.Chang GC, Liu KJ, Hsieh CL, et al. Identification of α-enolase as an autoantigen in lung cancer: its overexpression is associated with clinical outcomes. Clinical Cancer Research. 2006;12(19):5746–5754. doi: 10.1158/1078-0432.CCR-06-0324. [DOI] [PubMed] [Google Scholar]
- 99.López-Pedrera C, Villalba JM, Siendones E, et al. Proteomic analysis of acute myeloid leukemia: identification of potential early biomarkers and therapeutic targets. Proteomics. 2006;6(supplement 1):S293–S299. doi: 10.1002/pmic.200500384. [DOI] [PubMed] [Google Scholar]
- 100.Katayama M, Nakano H, Ishiuchi A, et al. Protein pattern difference in the colon cancer cell lines examined by two-dimensional differential in-gel electrophoresis and mass spectrometry. Surgery Today. 2006;36(12):1085–1093. doi: 10.1007/s00595-006-3301-y. [DOI] [PubMed] [Google Scholar]
- 101.Capello M, Ferri-Borgogno S, Cappello P, Novelli F. α-enolase: a promising therapeutic and diagnostic tumor target. FEBS Journal. 2011;278(7):1064–1074. doi: 10.1111/j.1742-4658.2011.08025.x. [DOI] [PubMed] [Google Scholar]
- 102.Jiang W, Li X, Rao S, et al. Constructing disease-specific gene networks using pair-wise relevance metric: application to colon cancer identifies interleukin 8, desmin and enolase 1 as the central elements. BMC Systems Biology. 2008;2, article 72 doi: 10.1186/1752-0509-2-72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Georges E, Bonneau AM, Prinos P. RNAi-mediated knockdown of alpha-enolase increases the sensitivity of tumor cells to antitubulin chemotherapeutics. International Journal of Biochemistry and Molecular Biology. 2011;2:303–308. [PMC free article] [PubMed] [Google Scholar]
- 104.Trojanowicz B, Winkler A, Hammje K, et al. Retinoic acid-mediated down-regulation of ENO1/MBP-1 gene products caused decreased invasiveness of the follicular thyroid carcinoma cell lines. Journal of Molecular Endocrinology. 2009;42(3):249–260. doi: 10.1677/JME-08-0118. [DOI] [PubMed] [Google Scholar]
- 105.Tsai ST, Chien IH, Shen WH, et al. ENO1, a potential prognostic head and neck cancer marker, promotes transformation partly via chemokine CCL20 induction. European Journal of Cancer. 2010;46(9):1712–1723. doi: 10.1016/j.ejca.2010.03.018. [DOI] [PubMed] [Google Scholar]
- 106.Jin S, DiPaola RS, Mathew R, White E. Metabolic catastrophe as a means to cancer cell death. Journal of Cell Science. 2007;120(3):379–383. doi: 10.1242/jcs.03349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Abiko Y, Nishimura M, Kusano K, et al. Expression of MIP-3α/CCL20, a macrophage inflammatory protein in oral squamous cell carcinoma. Archives of Oral Biology. 2003;48(2):171–175. doi: 10.1016/s0003-9969(02)00167-x. [DOI] [PubMed] [Google Scholar]
- 108.Zhou W, Capello M, Fredolini C, et al. Mass spectrometry analysis of the post-translational modifications of r-enolase from pancreatic ductal adenocarcinoma cells. Journal of Proteome Research. 2010;9(6):2929–2936. doi: 10.1021/pr901109w. [DOI] [PubMed] [Google Scholar]
- 109.Tomaino B, Cappello P, Capello M, et al. Circulating autoantibodies to phosphorylated α-enolase are a hallmark of pancreatic cancer. Journal of Proteome Research. 2011;10(1):105–112. doi: 10.1021/pr100213b. [DOI] [PubMed] [Google Scholar]
- 110.Iwabata H, Yoshida M, Komatsu Y. Proteomic analysis of organ-specific post-translational lysine-acetylation and -methylation in mice by use of anti-acetyllysine and -methyllysine mouse monoclonal antibodies. Proteomics. 2005;5(18):4653–4664. doi: 10.1002/pmic.200500042. [DOI] [PubMed] [Google Scholar]
- 111.Lu N, Zhang Y, Li H, Gao Z. Oxidative and nitrative modifications of α-enolase in cardiac proteins from diabetic rats. Free Radical Biology and Medicine. 2010;48(7):873–881. doi: 10.1016/j.freeradbiomed.2010.01.010. [DOI] [PubMed] [Google Scholar]
- 112.Gannon J, Staunton L, O’Connell K, Doran P, Ohlendieck K. Phosphoproteomic analysis of aged skeletal muscle. International Journal of Molecular Medicine. 2008;22(1):33–42. [PubMed] [Google Scholar]
- 113.Baraibar MA, Hyzewicz J, Rogowska-Wrzesinska A, et al. Oxidative stress-induced proteome alterations target different cellular pathways in human myoblasts. Free Radical Biology and Medicine. 2011;51:1522–1532. doi: 10.1016/j.freeradbiomed.2011.06.032. [DOI] [PubMed] [Google Scholar]
- 114.Kinloch A, Tatzer V, Wait R, et al. Identification of citrullinated alpha-enolase as a candidate autoantigen in rheumatoid arthritis. Arthritis Research & Therapy. 2005;7(6):R1421–1429. doi: 10.1186/ar1845. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Pratesi F, Moscato S, Sabbatini A, Chimenti D, Bombardieri S, Migliorini P. Autoantibodies specific for α-enolase in systemic autoimmune disorders. Journal of Rheumatology. 2000;27(1):109–115. [PubMed] [Google Scholar]
- 116.Wakui K, Tanemura M, Suzumori K, et al. Clinical applications of two-color telomeric fluorescence in situ hybridization for prenatal diagnosis: identification of chromosomal translocation in five families with recurrent miscarriages or a child with multiple congenital anomalies. Journal of Human Genetics. 1999;44(2):85–90. doi: 10.1007/s100380050115. [DOI] [PubMed] [Google Scholar]
- 117.Saulot V, Vittecoq O, Charlionet R, et al. Presence of autoantibodies to the glycolytic enzyme α-enolase in sera from patients with early rheumatoid arthritis. Arthritis and Rheumatism. 2002;46(5):1196–1201. doi: 10.1002/art.10252. [DOI] [PubMed] [Google Scholar]
- 118.Wegner N, Lundberg K, Kinloch A, et al. Autoimmunity to specific citrullinated proteins gives the first clues to the etiology of rheumatoid arthritis. Immunological Reviews. 2010;233(1):34–54. doi: 10.1111/j.0105-2896.2009.00850.x. [DOI] [PubMed] [Google Scholar]
- 119.Chang X, Wei C. Glycolysis and rheumatoid arthritis. International Journal of Rheumatic Diseases. 2011;14(3):217–222. doi: 10.1111/j.1756-185X.2011.01598.x. [DOI] [PubMed] [Google Scholar]
- 120.Jacovina AT, Zhong F, Khazanova E, Lev E, Deora AB, Hajjar KA. Neuritogenesis and the nerve growth factor-induced differentiation of PC-12 cells requires annexin II-mediated plasmin generation. Journal of Biological Chemistry. 2001;276(52):49350–49358. doi: 10.1074/jbc.M106289200. [DOI] [PubMed] [Google Scholar]
- 121.Obermajer N, Doljak B, Jamnik P, Fonović UP, Kos J. Cathepsin X cleaves the C-terminal dipeptide of alpha- and gamma-enolase and impairs survival and neuritogenesis of neuronal cells. International Journal of Biochemistry and Cell Biology. 2009;41(8-9):1685–1696. doi: 10.1016/j.biocel.2009.02.019. [DOI] [PubMed] [Google Scholar]
- 122.Butterfield DA, Lange MLB. Multifunctional roles of enolase in Alzheimer’s disease brain: beyond altered glucose metabolism. Journal of Neurochemistry. 2009;111(4):915–933. doi: 10.1111/j.1471-4159.2009.06397.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Castegna A, Aksenov M, Thongboonkerd V, et al. Proteomic identification of oxidatively modified proteins in Alzheimer’s disease brain. Part II: dihydropyrimidinase-related protein 2, α-enolase and heat shock cognate 71. Journal of Neurochemistry. 2002;82(6):1524–1532. doi: 10.1046/j.1471-4159.2002.01103.x. [DOI] [PubMed] [Google Scholar]
- 124.Owen JB, Domenico FD, Suitana R, et al. Proteomics-determined differences in the concanavalin-A-fractionated proteome of hippocampus and inferior parietal lobule in subjects with alzheimer’s disease and mild cognitive impairment: implications for progression of AD. Journal of Proteome Research. 2009;8(2):471–482. doi: 10.1021/pr800667a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Newman SF, Sultana R, Perluigi M, et al. An increase in S-glutathionylated proteins in the Alzheimer’s disease inferior parietal lobule, a proteomics approach. Journal of Neuroscience Research. 2007;85(7):1506–1514. doi: 10.1002/jnr.21275. [DOI] [PubMed] [Google Scholar]
- 126.Zhang Y, Zhou ZH, Bugge TH, Wahl LM. Urokinase-type plasminogen activator stimulation of monocyte matrix metalloproteinase-1 production is mediated by plasmin-dependent signaling through annexin A2 and inhibited by inactive plasmin. Journal of Immunology. 2007;179(5):3297–3304. doi: 10.4049/jimmunol.179.5.3297. [DOI] [PubMed] [Google Scholar]
- 127.Syrovets T, Jendrach M, Rohwedder A, Schüle A, Simmet T. Plasmin-induced expression of cytokines and tissue factor in human monocytes involves AP-1 and IKKβ-mediated NF-κB activation. Blood. 2001;97(12):3941–3950. doi: 10.1182/blood.v97.12.3941. [DOI] [PubMed] [Google Scholar]
- 128.Burysek L, Syrovets T, Simmet T. The serine protease plasmin triggers expression of MCP-1 and CD40 in human primary monocytes via activation of p38 MAPK and Janus kinase (JAK)/STAT signaling pathways. Journal of Biological Chemistry. 2002;277(36):33509–33517. doi: 10.1074/jbc.M201941200. [DOI] [PubMed] [Google Scholar]
- 129.Li Q, Laumonnier Y, Syrovets T, Simmet T. Plasmin triggers cytokine induction in human monocyte-derived macrophages. Arteriosclerosis, Thrombosis, and Vascular Biology. 2007;27(6):1383–1389. doi: 10.1161/ATVBAHA.107.142901. [DOI] [PubMed] [Google Scholar]
- 130.Pendurthi UR, Ngyuen M, Andrade-Gordon P, Petersen LC, Rao LVM. Plasmin induces Cyr61 gene expression in fibroblasts via protease-activated receptor-1 and p44/42 mitogen-activated protein kinase-dependent signaling pathway. Arteriosclerosis, Thrombosis, and Vascular Biology. 2002;22(9):1421–1426. doi: 10.1161/01.atv.0000030200.59331.3f. [DOI] [PubMed] [Google Scholar]
- 131.Mandal SK, Rao LVM, Tran TTT, Pendurthi UR. A novel mechanism of plasmin-induced mitogenesis in fibroblasts. Journal of Thrombosis and Haemostasis. 2005;3(1):163–169. doi: 10.1111/j.1538-7836.2004.01054.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Sousa LP, Silva BM, Brasil BSAF, et al. Plasminogen/plasmin regulates α-enolase expression through the MEK/ERK pathway. Biochemical and Biophysical Research Communications. 2005;337(4):1065–1071. doi: 10.1016/j.bbrc.2005.09.154. [DOI] [PubMed] [Google Scholar]
- 133.De Sousa LP, Brasil BSAF, Silva BM, et al. Plasminogen/plasmin regulates c-fos and egr-1 expression via the MEK/ERK pathway. Biochemical and Biophysical Research Communications. 2005;329(1):237–245. doi: 10.1016/j.bbrc.2005.01.123. [DOI] [PubMed] [Google Scholar]
- 134.Majumdar M, Tarui T, Shi B, Akakura N, Ruf W, Takada Y. Plasmin-induced migration requires signaling through protease-activated receptor 1 and integrin α9β1. Journal of Biological Chemistry. 2004;279(36):37528–37534. doi: 10.1074/jbc.M401372200. [DOI] [PubMed] [Google Scholar]
- 135.Tarui T, Majumdar M, Miles LA, Ruf W, Takada Y. Plasmin-induced migration of endothelial cells: a potential target for the anti-angiogenic action of angiostatin. Journal of Biological Chemistry. 2002;277(37):33564–33570. doi: 10.1074/jbc.M205514200. [DOI] [PubMed] [Google Scholar]
- 136.Roztocil E, Nicholl SM, Galaria II, Davies MG. Plasmin-induced smooth muscle cell proliferation requires epidermal growth factor activation through an extracellular pathway. Surgery. 2005;138(2):180–186. doi: 10.1016/j.surg.2005.06.014. [DOI] [PubMed] [Google Scholar]
- 137.Petrak J, Ivanek R, Toman O, et al. Déjà vu in proteomics. A hit parade of repeatedly identified differentially expressed proteins. Proteomics. 2008;8(9):1744–1749. doi: 10.1002/pmic.200700919. [DOI] [PubMed] [Google Scholar]
- 138.Roozendaal C, Zhao MH, Horst G, et al. Catalase and α-enolase: two novel granulocyte autoantigens in inflammatory bowel disease (IBD) Clinical and Experimental Immunology. 1998;112(1):10–16. doi: 10.1046/j.1365-2249.1998.00528.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Vermeulen N, Arijs I, Joossens S, et al. Anti-α-enolase antibodies in patients with inflammatory bowel disease. Clinical Chemistry. 2008;54(3):534–541. doi: 10.1373/clinchem.2007.098368. [DOI] [PubMed] [Google Scholar]
- 140.Ballot E, Bruneel A, Labas V, Johanet C. Identification of rat targets of anti-soluble liver antigen autoantibodies by serologic proteome analysis. Clinical Chemistry. 2003;49(4):634–643. doi: 10.1373/49.4.634. [DOI] [PubMed] [Google Scholar]
- 141.Bruschi M, Carnevali ML, Murtas C, et al. Direct characterization of target podocyte antigens and auto-antibodies in human membranous glomerulonephritis: alfa-enolase and borderline antigens. Journal of Proteomics. 2011;74(10):2008–2017. doi: 10.1016/j.jprot.2011.05.021. [DOI] [PubMed] [Google Scholar]
- 142.Kim JW, Dang CV. Multifaceted roles of glycolytic enzymes. Trends in Biochemical Sciences. 2005;30(3):142–150. doi: 10.1016/j.tibs.2005.01.005. [DOI] [PubMed] [Google Scholar]