

Abstract
After partial hepatectomy (PH) the initial mass of the organ is restored through a complex network of cellular interactions that orchestrate both proliferative and hepatoprotective signalling cascades. Among agonists involved in this network many of them drive Ca2+ movements. During liver regeneration in the rat, hepatocyte cytosolic Ca2+ signalling has been shown on the one hand to be deeply remodelled and on the other hand to enhance progression of hepatocytes through the cell cycle. Mechanisms through which cytosolic Ca2+ signals impact on hepatocyte cell cycle early after PH are not completely understood, but at least they include regulation of immediate early gene transcription and ERK and CREB phosphorylation. In addition to cytosolic Ca2+, there is also evidence that mitochondrial Ca2+ and also nuclear Ca2+ may be critical for the regulation of liver regeneration. Finally, Ca2+ movements in hepatocytes, and possibly in other liver cells, not only impact hepatocyte progression in the cell cycle but more generally may regulate cellular homeostasis after PH.
1. Introduction
After partial hepatic destruction in experimental or clinical context, the initial mass of the organ is restored through compensatory growth of the remnant liver. A complex and yet incompletely elucidated network of cellular interactions (including paracrine, autocrine, endocrine, or nervous pathways) orchestrates the regulation of regeneration, through both proliferative and hepatoprotective signalling cascades [1]. A number of agonists constituting this network drive intracellular Ca2+ movements, in particular through the formation of inositol 1,4,5 trisphosphate which binds on its receptor in the membrane of the endoplasmic reticulum, and release the Ca2+ stored in this organelle. Such agonists include some of the main comitogenic—as noradrenalin [2], arginine vasopressin (AVP) [3], and adenosine triphosphate (ATP) [4]—and mitogenic factors, as epidermal growth factor (EGF), hepatocyte growth factor (HGF) [5], and insulin [6]. The resulting increase in ionized cytosolic calcium concentration generally consists of a regular succession of Ca2+ peaks (oscillations) [7] that can be transmitted to other cells (intercellular calcium waves) which mechanisms and functions are not fully known [8, 9]. The impact of calcium signalling on liver regeneration has, however, only been scarcely studied.
2. Hepatocyte Ca2+ Signalling: Mechanisms and Functions
The first calcium oscillations were reported in hepatocytes [10], and numerous studies, both experimental and theoretical, have been conducted after that to decipher, in the hepatocyte, the machinery by which agonists generated cytosolic calcium signals. However, the functions of hepatocyte calcium signalling remain far less understood.
In hepatocytes, as in most nonexcitable cells, Ca2+ oscillations originate from the periodic opening of Ca2+ channels located in the ER membrane, following activation of the phosphoinositide cascade. The binding of an agonist to a membrane-bound receptor activates the Gα-subunit of a G-protein complex coupled to the receptor. This activated G protein in turn stimulates phospholipase C (PLC) activity. The latter enzyme catalyzes the hydrolysis of the membrane-bound phosphatidyl-inositol bisphosphate (PIP2) into diacyl-glycerol and inositol trisphosphate (InsP3). Ca2+ release from the internal stores is ensured by the InsP3R, an homotetramer that can bind up to 4 InsP3 molecules, forming a Ca2+ channel which equilibrium open probability presents a bell-shaped dependence on cytosolic Ca2+ [11]. The decrease of [Ca2+]i in the cytosol is due to the activity of the Ca2+ ATPases (SERCA pumps), which actively transports Ca2+ from the cytosol into the ER. Ca2+-regulated InsP3Rs and Ca2+ ATPases are together sufficient to generate Ca2+ oscillations [12]. In most cases, hormone-induced Ca2+ oscillations in hepatocytes take the form of repetitive, sharp spikes sometimes preceded by a slower, pacemaker-like elevation in the cytosolic Ca2+ concentration. These periodic increases in the level of free Ca2+ in the cytosol from about 0.1 μM up to 1 μM have been observed in hepatocytes in response to stimulation by a large number of agonists such as noradrenalin, vasopressin, phenylephrin, angiotensin II, adenosine triphosphate (ATP), histamine, and thrombin, the shape of the oscillations being agonist dependent [7]. The oscillation frequency increases with the agonist concentration, a phenomenon known as “frequency encoding”, and is affected by external [Ca2+]—and thus by the rate of Ca2+ entry into the cell through plasma membrane Ca2+ channels. Intracellular Ca2+ waves do not result from a simple diffusion of Ca2+ itself—which is quickly buffered—but from the spreading of InsP3—which is more soluble in the cytosol—that mobilizes Ca2+ from storage compartments throughout the cell. A particular spatial pattern of subcellular InsP3R distribution was reported to support the direction of intracellular Ca2+ waves, starting from the canalicular region containing the most abundant and affine InsP3R isoform (type II InsP3R) and spreading toward the other regions of the cytosol, less sensitive to InsP3 [13]. As in many other cell types, intracellular movements of Ca2+ in hepatocytes, induced by hormones and neurotransmitters, may be propagated from cell to cell. Our group demonstrated in multicellular rat hepatocyte systems (couplets and triplets) that agonists such as vasopressin or noradrenalin induce tightly coordinated and sequentially ordered intracellular Ca2+ increases [8, 14–16]. Such signals were also observed in the intact perfused liver in which vasopressin elicits waves of [Ca2+]i increase running along hepatocyte plates across the lobules [17–19]. We demonstrated that unidirectional Ca2+ waves resulted from a gradually decreasing cellular sensitivity to hormonal stimuli from the first to the last responding cell, and that this cell to cell heterogeneity was due to a lobular gradual distribution of hormonal receptors density [8]. Moreover, InsP3 has been shown to flow through gap junctions and thereby coordinate Ca2+ spiking among adjacent hepatocytes [20]. Such a configuration in which the most responsive hepatocytes drive the response of the less sensitive cells is similar to the cell to cell triggering of cardiac pacemaker cells [8, 21, 22].
In general terms, Ca2+ oscillations in hepatocytes optimize the effect of hormonal stimulation, thanks to enzymes decoding [23] their frequency [24, 25], as proposed for glycogen metabolism in hepatocytes [26]. Since the early data showing that production of glucose by the liver was at least in part mediated by hormone-induced intracellular Ca2+ increases [27], most recent studies have deciphered the molecular mechanisms linking intracellular Ca2+ to glucose metabolism in hepatocytes [28–30]. In particular, the serine-threonine kinase “calcium calmodulin-dependent kinase II” (CaMKII), a major mediator of Ca2+ signalling in different cell types, has been found to play essential roles in the regulation of glycogenolysis and gluconeogenesis in hepatocytes, not only during physiological fasting, but also in the pathophysiological setting of obesity [28, 30]. It is moreover well established that Ca2+ oscillations in hepatocytes coordinate intramitochondrial ATP synthesis with cellular energy demand, maintaining cell homeostasis and viability [31]. It has also been demonstrated that [32] the temporal pattern of calcium signals was of major impact as to the expression of transcription factors in lymphocytes, but this aspect has never been investigated in hepatocytes. Many events related to bile secretion are also regulated by cytosolic Ca2+, such as vesicular trafficking and canalicular exocytosis of bile acid transporters [33, 34], permeability of tight junctions [35], or canalicular contraction [9, 36]. Intracellular calcium waves, as described above, starting from the canaliculus to the basolateral poles may have physiological impact on secretion, as it has been shown in pancreatic acinar cells [37], although direct evidence in hepatocytes is lacking. Moreover, interhepatocyte calcium waves have been reported to support canalicular peristaltism and thereby to regulate bile flow, in the normal and regenerating rat liver [3, 38]. As emphasized in the following, all these Ca2+-regulated physiological processes may impact the course of liver regeneration.
3. Intracellular Calcium and Hepatocyte Proliferation: Liver Regeneration
It is well established that intracellular Ca2+ is crucial for tissue homeostasis through regulation of cell cycle and apoptosis [7]. In particular, intracellular calcium has been reported to regulate cell proliferation at multiple steps of the cell cycle, from immediate early genes activation, toward G1-S and G2-M transitions, as well as during mitosis [39]. Pioneer studies have shown that extracellular calcium was crucial for liver regeneration [40]. Also, modifications of intracellular calcium homeostasis during liver regeneration have been reported, concerning Ca2+-binding proteins [41], membrane Ca2+-ATPases [42], or the InsP3 receptor [43, 44]. It has been also suggested that the alteration of the InsP3 and Ca2+mobilisation pathway could alter liver regeneration in the rat [45]. Subsequently, it has been shown in nonhepatocytic cell lines that the spatiotemporal organisation of Ca2+ signals was determined for the activation of transcription factors like CREB, NF-κB, or NF-AT, and for immediate early genes like c-fos or c-jun [46–48]. It is also well established that the activation of the RAS pathway is controlled by [Ca2+]i oscillations [49]. Moreover, intra-nuclear calcium signals, which have been well documented [50], have a major impact on gene transcription [51–53] and can result either from the diffusion of cytosolic calcium to the nucleus, or from an InsP3 -mediated calcium release in the nucleus itself (see below) [6, 54]. Finally, our previous work demonstrated that hepatocyte calcium signalling was deeply remodelled during liver regeneration in the rat, contributing to the regulation of bile flow and cell proliferation [3, 55].
In a recent study, we examined the physiological involvement of cytosolic calcium during liver regeneration in the rat [56]. We interfered with calcium signalling before PH by expressing parvalbumin (PV) in the liver, a calcium-binding protein expressed in muscle cells and neurons but absent from the liver [57], using adenoviruses coding for PV targeted to the cytosol, to selectively buffer Ca2+ in this compartment [53, 54]. We found that expression of PV efficiently buffers agonist-induced calcium oscillations in the cytosol and inhibits primary hepatocyte proliferation in vitro as well as in vivo during liver regeneration.
We found that immediate early gene transcription, early phosphorylation of ERK and CREB, and hepatocyte progression in the cell cycle after PH were inhibited in rats expressing cytosolic PV [56]. These data were in line with previous reports describing these pathways and genes as dependant on cytosolic and/or nuclear calcium signalling [46, 58–60]. We thus suggested that attenuated [Ca2+]i oscillations in calcium-buffered hepatocytes resulted in impaired activation of these pathways. A potential reduction in CaM-kinase activation, as previously reported [61], or reduced ERK1/2 activation that we observed in PV-NES expressing hepatocytes may have also contributed to altered CREB phosphorylation. Since CaM-kinase II [24], as well as ERK1/2 [60] activity, is sensitive to Ca2+ oscillation frequency, an attractive hypothesis would be that cytosolic PV expression, by attenuating agonist-generated Ca2+ signals, resulted in impaired phosphorylation of CREB.
Cytosolic calcium signalling impacts most likely the early triggering of hepatocyte progression from G0 to G1 and S phases. In line with this view, a rise in concentration—in the liver and in the plasma—is observed early after PH for several Ca2+-mobilizing agonists, suggesting these agonists might be involved in initiating the regeneration process. In particular, EGF and HGF elicit cytosolic Ca2+ oscillations in hepatocytes, the physiological impact of which has never been specifically addressed [5]. Also, extracellular ATP [4], arginine vasopressin [3], and noradrenalin [2], which are comitogenic Ca2+-mobilizing agonists, have been individually reported to contribute to early phases of liver regeneration. Our study thus suggested that buffering hepatocyte calcium signals, potentially generated by these agonists in the minutes after PH, result in delaying hepatocyte cell cycle progression.
There is evidence in the literature for the crucial role of mitochondrial calcium in the regulation of apoptotic processes. It is well known in particular, that mitochondrial calcium overload can be one of the pathways leading to the swelling of mitochondria and to the rupture of the outer membrane, in turn releasing proapoptotic molecules in the cytosol. Mechanisms for excessive calcium transfer to mitochondria are debated and include mainly interactions between proteins of the Bcl2 family and the InsP3-R. Anti-apoptotic members appeared as reducing calcium transfer from the ER to the mitochondria, whereas proapoptotic factors were reported to enhance this flux [62]. In this context, recent data suggest that mitochondrial Ca2+, as well as cytosolic Ca2+, may be critical for the regulation of liver regeneration after PH in the rat [63]. The authors suggested that buffering calcium in the mitochondria resulted in a shift in the balance between pro- and antiapoptotic factors, thereby protecting hepatocytes from apoptosis, in vitro in an hepatoma cell line, as well as in vivo in the rat liver after PH.
4. Nuclear Calcium Signalling and Liver Cell Proliferation
Previous studies have established that growth factors important for liver regeneration such as HGF [54] and insulin [6] can differentially affect cytosolic and nuclear calcium in hepatocytes. It has been reported that agonist-induced calcium movements in the nucleus can schematically result from the diffusion of cytosolic calcium to the nucleus and/or from an autonomous InsP3 generation and calcium mobilization from local, intra, or perinuclear Ca2+ stores [64, 65]. Nuclear InsP3 may again come from the cytosol or be generated in the nucleus [6]. Indeed, the nucleus, its envelope and the nucleoplasmic reticulum, has been shown to possess several crucial molecules involved in Ca2+ storage, InsP3 production, and calcium release, therefore opening the possibility that local nuclear Ca2+ signals may occur independently from the cytosol [64, 65]. Although these two views may coexist in the same cells according to circumstances and cell types, it has been shown in SkHep cells that an InsP3-sensitive intranuclear calcium compartment (i.e., the “nucleoplasmic reticulum”) exists [50]. PLCβ, PIP2, and InsP3R have been found in the nucleus, allowing a local InsP3 production and providing the machinery necessary to generate autonomous Ca2+ signals [64, 65]. We also know that calcium signals in these two compartments—cytosol and nucleus—can have different effects [51, 58]. Recent works revealed that buffering calcium in the nucleus, but not in the cytosol, in a hepatoma cell line, resulted in an inhibition of cell proliferation, suggesting that nuclear Ca2+ was necessary for centrosome separation and cell progression through early prophase [53]. Gomes et al. further showed that the HGF receptor (c-met) can translocate (upon agonist stimulation) from the plasma membrane to the nucleus and generate an InsP3 production and calcium elevation in the nucleus, independently of cytosolic calcium, in a hepatoma cell line [54]. Very similarly, it was shown by the same group that insulin can induce nuclear calcium signals through a translocation of its receptor to the nucleus, in primary rat hepatocytes [6]. Importantly, the nucleoplasmic reticulum as an intranuclear calcium compartment has not been shown in primary hepatocytes, and some authors claimed that it was not essential for calcium signalling [66].
Important cellular functions are thought to be regulated by nuclear calcium signals, including nuclear pore permeability, transcription factor activity and protein kinase translocation, thereby controlling gene expression [65]. In particular, the transcriptional activity of CREB [58], NFAT [46], and DREAM [67] has been well described as dependent on nuclear Ca2+. Therefore, agonist-induced nuclear Ca2+ movements are potentially expected to impact both hepatocyte progression in the cell cycle and more generally cellular homeostasis after PH.
It is important to realize that liver regeneration, seen as a process involving the whole organ—and even the entire organism—cannot be restrained to the sole hepatocyte division. In that view, after PH, Ca2+ signalling may also regulate physiological processes unrelated directly to cell cycle control, although they may ultimately interfere with cell cycle progression, not only in hepatocytes but also in other liver cell types (cholangiocytes, endothelial and Kupffer cells). Whereas we do not have any direct data about the impact of Ca2+ signals in nonhepatocytic cells after PH, we can easily anticipate about Ca2+-dependent physiological processes that may be crucial in hepatocytes after PH. For example, glucose homeostasis, which regulation is critical for hepatocyte exit from quiescence [68], may depend on Ca2+ signals early after PH. Also, biliary homeostasis, which adaptation after PH is critical for liver regeneration [69, 70], may involve Ca2+-dependent regulation [3, 71].
An integrated full picture of the “liver calcium signalling”, which is obviously lacking, may improve our knowledge on the interaction network that regulates liver regeneration processes (Figure 1).
Figure 1.
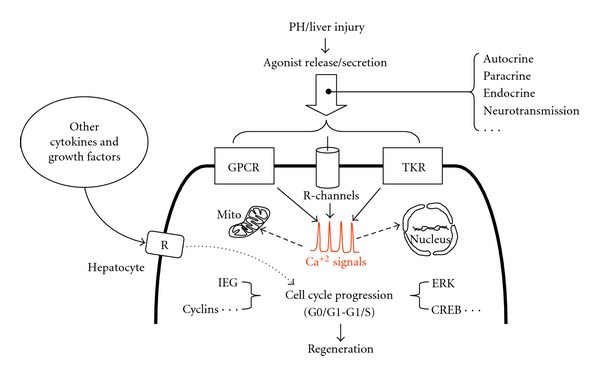
A simplified view of the impact of hepatocyte calcium signals during liver regeneration. After PH or toxic liver injury, a number of calcium mobilizing agonists are released inside or outside the liver, interacting with hepatocytes through autocrine, paracrine, and endocrine pathways. Diverse membrane receptors, either G protein coupled receptors (GPCR), tyrosine kinase receptors (TKR), or receptor channels (e.g., ionotropic purinergic receptors), can be involved in the generation of cytosolic calcium signals. These calcium movements in the cytosol can be transferred to other crucial compartments like the mitochondrion (Mito) or the nucleus, in which they could regulate respectively apoptosis and gene transcription. Previous studies have shown that cytosolic calcium signals regulate cell cycle progression from G0 to G1 and from G1 to S phases in hepatocytes after PH, at least in part through an impact on immediate early genes transcription, cyclin expression, and ERK and CREB phosphorylation.
References
- 1.Taub R. Liver regeneration: from myth to mechanism. Nature Reviews Molecular Cell Biology. 2004;5(10):836–847. doi: 10.1038/nrm1489. [DOI] [PubMed] [Google Scholar]
- 2.Cruise JL, Muga SJ, Lee YS, Michalopoulos GK. Regulation of hepatocyte growth: Alpha-1 adrenergic receptor and ras p21 changes in liver regeneration. Journal of Cellular Physiology. 1989;140(2):195–201. doi: 10.1002/jcp.1041400202. [DOI] [PubMed] [Google Scholar]
- 3.Nicou A, Serriere V, Prigent S, et al. Hypothalamic vasopressin release and hepatocyte Ca2+ signaling during liver regeneration: an interplay stimulating liver growth and bile flow. The FASEB Journal. 2003;17:1901–1903. doi: 10.1096/fj.03-0082fje. [DOI] [PubMed] [Google Scholar]
- 4.Gonzales E, Julien B, Serrière-Lanneau V, et al. ATP release after partial hepatectomy regulates liver regeneration in the rat. Journal of Hepatology. 2010;52(1):54–62. doi: 10.1016/j.jhep.2009.10.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Mine T, Kojima I, Ogata E, Nakamura T. Comparison of effects of HGF and EGF on cellular calcium in rat hepatocytes. Biochemical and Biophysical Research Communications. 1991;181(3):1173–1180. doi: 10.1016/0006-291x(91)92062-o. [DOI] [PubMed] [Google Scholar]
- 6.Rodrigues MA, Gomes DA, Andrade VA, Leite MF, Nathanson MH. Insulin induces calcium signals in the nucleus of rat hepatocytes. Hepatology. 2008;48(5):1621–1631. doi: 10.1002/hep.22424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Berridge MJ, Bootman MD, Roderick HL. Calcium signalling: dynamics, homeostasis and remodelling. Nature Reviews Molecular Cell Biology. 2003;4(7):517–529. doi: 10.1038/nrm1155. [DOI] [PubMed] [Google Scholar]
- 8.Tordjmann T, Berthon B, Jacquemin E, et al. Receptor-oriented intercellular calcium waves evoked by vasopressin in rat hepatocytes. The EMBO Journal. 1998;17(16):4695–4703. doi: 10.1093/emboj/17.16.4695. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Serrière V, Tran D, Stelly N, et al. Vasopressin-induced morphological changes in polarized rat hepatocyte multiplets: dual calcium-dependent effects. Cell Calcium. 2008;43(1):95–104. doi: 10.1016/j.ceca.2007.04.008. [DOI] [PubMed] [Google Scholar]
- 10.Woods NM, Cuthbertson KSR. Repetitive transient rises in cytoplasmic free calcium in hormone-stimulated hepatocytes. Nature. 1986;319(6054):600–602. doi: 10.1038/319600a0. [DOI] [PubMed] [Google Scholar]
- 11.Taylor CW. Inositol trisphosphate receptors: Ca2+-modulated intracellular Ca2+ channels. Biochimica et Biophysica Acta. 1998;1436(1-2):19–33. doi: 10.1016/s0005-2760(98)00122-2. [DOI] [PubMed] [Google Scholar]
- 12.Dupont G, Swillens S, Clair C, Tordjmann T, Combettes L. Hierarchical organization of calcium signals in hepatocytes: From experiments to models. Biochimica et Biophysica Acta. 2000;1498(2-3):134–152. doi: 10.1016/s0167-4889(00)00090-2. [DOI] [PubMed] [Google Scholar]
- 13.Hernandez E, Leite MF, Guerra MT, et al. The spatial distribution of inositol 1,4,5-trisphosphate receptor isoforms shapes Ca2+ waves. Journal of Biological Chemistry. 2007;282(13):10057–10067. doi: 10.1074/jbc.M700746200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Nathanson MH, Burgstahler AD. Coordination of hormone-induced calcium signals in isolated rat hepatocyte couplets: demonstration with confocal microscopy. Molecular Biology of the Cell. 1992;3(1):113–121. doi: 10.1091/mbc.3.1.113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Combettes L, Tran D, Tordjmann T, Laurent M, Berthon B, Claret M. Ca2+-mobilizing hormones induce sequentially ordered Ca2+ signals in multicellular systems of rat hepatocytes. Biochemical Journal. 1994;304, part 2:585–594. doi: 10.1042/bj3040585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Tordjmann T, Berthon B, Claret M, Combettes L. Coordinated intercellular calcium waves induced by noradrenaline in rat hepatocytes: dual control by gap junction permeability and agonist. The EMBO Journal. 1997;16(17):5398–5407. doi: 10.1093/emboj/16.17.5398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Nathanson MH, Burgstahler AD, Mennone A, Fallon MB, Gonzalez CB, Saez JC. Ca2+ waves are organized among hepatocytes in the intact organ. American Journal of Physiology—Gastrointestinal and Liver Physiology. 1995;269(1):G167–G171. doi: 10.1152/ajpgi.1995.269.1.G167. [DOI] [PubMed] [Google Scholar]
- 18.Robb-Gaspers LD, Thomas AP. Coordination of Ca2+ signaling by intercellular propagation of Ca2+ waves in the intact liver. Journal of Biological Chemistry. 1995;270(14):8102–8107. doi: 10.1074/jbc.270.14.8102. [DOI] [PubMed] [Google Scholar]
- 19.Patel S, Robb-Gaspers LD, Stellato KA, Shon M, Thomas AP. Coordination of calcium signalling by endothelial-derived nitric oxide in the intact liver. Nature Cell Biology. 1999;1(8):467–471. doi: 10.1038/70249. [DOI] [PubMed] [Google Scholar]
- 20.Clair C, Chalumeau C, Tordjmann T, et al. Investigation of the roles of Ca2+ and InsP3 diffusion in the coordination of Ca2+ signals between connected hepatocytes. Journal of Cell Science. 2001;114, part 11:1999–2007. doi: 10.1242/jcs.114.11.1999. [DOI] [PubMed] [Google Scholar]
- 21.Dupont G, Tordjmann T, Clair C, Swillens S, Claret M, Combettes L. Mechanism of receptor-oriented intercellular calcium wave propagation in hepatocytes. The FASEB Journal. 2000;14(2):279–289. doi: 10.1096/fasebj.14.2.279. [DOI] [PubMed] [Google Scholar]
- 22.Leite MF, Hirata K, Pusl T, et al. Molecular basis for pacemaker cells in epithelia. Journal of Biological Chemistry. 2002;277(18):16313–16323. doi: 10.1074/jbc.M109207200. [DOI] [PubMed] [Google Scholar]
- 23.Meyer T, Stryer L. Calcium spiking. Annual Review of Biophysics and Biophysical Chemistry. 1991;20:153–174. doi: 10.1146/annurev.bb.20.060191.001101. [DOI] [PubMed] [Google Scholar]
- 24.de Koninck P, Schulman H. Sensitivity of CaM kinase II to the frequency of Ca2+ oscillations. Science. 1998;279(5348):227–230. doi: 10.1126/science.279.5348.227. [DOI] [PubMed] [Google Scholar]
- 25.Hanson PI, Meyer T, Stryer L, Schulman H. Dual role of calmodulin in autophosphorylation of multifunctional CaM kinase may underlie decoding of calcium signals. Neuron. 1994;12(5):943–956. doi: 10.1016/0896-6273(94)90306-9. [DOI] [PubMed] [Google Scholar]
- 26.Gall D, Baus E, Dupont G. Activation of the liver glycogen phosphorylase by Ca2+ oscillations: a theoretical study. Journal of Theoretical Biology. 2000;207(4):445–454. doi: 10.1006/jtbi.2000.2139. [DOI] [PubMed] [Google Scholar]
- 27.Exton JH. Mechanisms of hormonal regulation of hepatic glucose metabolism. Diabetes/Metabolism Reviews. 1987;3(1):163–183. doi: 10.1002/dmr.5610030108. [DOI] [PubMed] [Google Scholar]
- 28.Ozcan L, Wong CC, Li G, et al. Calcium signaling through CaMKII regulates hepatic glucose production in fasting and obesity. Cell Metabolism. 2012;15(5):739–751. doi: 10.1016/j.cmet.2012.03.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Wang Y, Li G, Goode J, et al. Inositol-1, 4, 5-trisphosphate receptor regulates hepatic gluconeogenesis in fasting and diabetes. Nature. 2012;485(7396):128–132. doi: 10.1038/nature10988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Anderson KA, Lin F, Ribar TJ, et al. Deletion of CaMKK2 from the liver lowers blood glucose and improves whole-body glucose tolerance in the mouse. Molecular Endocrinology. 2012;26(2):281–291. doi: 10.1210/me.2011-1299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Robb-Gaspers LD, Rutter GA, Burnett P, Hajnóczky G, Denton RM, Thomas AP. Coupling between cytosolic and mitochondrial calcium oscillations: role in the regulation of hepatic metabolism. Biochimica et Biophysica Acta. 1998;1366(1-2):17–32. doi: 10.1016/s0005-2728(98)00118-2. [DOI] [PubMed] [Google Scholar]
- 32.Dolmetsch RE, Xu K, Lewis RS. Calcium oscillations increase the efficiency and specificity of gene expression. Nature. 1998;392(6679):933–936. doi: 10.1038/31960. [DOI] [PubMed] [Google Scholar]
- 33.Beuers U, Nathanson MH, Isales CM, Boyer JL. Tauroursodeoxycholic acid stimulates hepatocellular exocytosis and mobilizes extracellular Ca++ mechanisms defective in cholestasis. Journal of Clinical Investigation. 1993;92(6):2984–2993. doi: 10.1172/JCI116921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Beuers U, Bilzer M, Chittattu A, et al. Tauroursodeoxycholic acid inserts the apical conjugate export pump, Mrp2, into canalicular membranes and stimulates organic anion secretion by protein kinase C-dependent mechanisms in cholestatic rat liver. Hepatology. 2001;33(5):1206–1216. doi: 10.1053/jhep.2001.24034. [DOI] [PubMed] [Google Scholar]
- 35.Nathanson MH, Gautam A, Ng OC, Bruck R, Boyer JL. Hormonal regulation of paracellular permeability in isolated rat hepatocyte couplets. American Journal of Physiology—Gastrointestinal and Liver Physiology. 1992;262(6):G1079–G1086. doi: 10.1152/ajpgi.1992.262.6.G1079. [DOI] [PubMed] [Google Scholar]
- 36.Kitamura T, Brauneis U, Gatmaitan Z, Arias IM. Extracellular ATP, intracellular calcium and canalicular contraction in rat hepatocyte doublets. Hepatology. 1991;14(4):640–647. doi: 10.1016/0270-9139(91)90051-v. [DOI] [PubMed] [Google Scholar]
- 37.Ito K, Miyashita Y, Kasai H. Micromolar and submicromolar Ca2+ spikes regulating distinct cellular functions in pancreatic acinar cells. The EMBO Journal. 1997;16(2):242–251. doi: 10.1093/emboj/16.2.242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Serrière V, Berthon B, Boucherie S, et al. Vasopressin receptor distribution in the liver controls calcium wave propagation and bile flow. The FASEB Journal. 2001;15(8):1484–1486. doi: 10.1096/fj.00-0659fje. [DOI] [PubMed] [Google Scholar]
- 39.Kahl CR, Means AR. Regulation of cell cycle progression by calcium/calmodulin-dependent pathways. Endocrine Reviews. 2003;24(6):719–736. doi: 10.1210/er.2003-0008. [DOI] [PubMed] [Google Scholar]
- 40.Whitfield JF, Boynton AL, MacManus JP, et al. The roles of calcium and cyclic AMP in cell proliferation. Annals of the New York Academy of Sciences. 1980;339:216–240. doi: 10.1111/j.1749-6632.1980.tb15980.x. [DOI] [PubMed] [Google Scholar]
- 41.Katsumata T, Yamaguchi M. Inhibitory effect of calcium-binding protein regucalcin on protein kinase activity in the nuclei of regenerating rat liver. Journal of Cellular Biochemistry. 1998;71:569–576. doi: 10.1002/(sici)1097-4644(19981215)71:4<569::aid-jcb11>3.0.co;2-z. [DOI] [PubMed] [Google Scholar]
- 42.Takahasi H, Yamaguchi M. Enhancement of plasma membrane (Ca2+-Mg2+)-ATPase activity in regenerating rat liver: involvement of endogenous activating protein regucalcin. Molecular and Cellular Biochemistry. 1996;162(2):133–138. doi: 10.1007/BF00227540. [DOI] [PubMed] [Google Scholar]
- 43.Diaz-Munoz M, Canedo-Merino R, Gutierrez-Salinas J, Hernandez-Munoz R. Modifications of intracellular calcium release channels and calcium mobilization following 70% hepatectomy. Archives of Biochemistry and Biophysics. 1998;349:105–112. doi: 10.1006/abbi.1997.0396. [DOI] [PubMed] [Google Scholar]
- 44.Magnino F, St-Pierre M, Lüthi M, Hilly M, Mauger JP, Dufour JF. Expression of intracellular calcium channels and pumps after partial hepatectomy in rat. Molecular Cell Biology Research Communications. 2000;3(6):374–379. doi: 10.1006/mcbr.2000.0242. [DOI] [PubMed] [Google Scholar]
- 45.Zhang BH, Horsfield BP, Farrell GC. Chronic ethanol administration to rats decreases receptor-operated mobilization of intracellular ionic calcium in cultured hepatocytes and inhibits 1,4,5-inositol trisphosphate production: relevance to impaired liver regeneration. Journal of Clinical Investigation. 1996;98(5):1237–1244. doi: 10.1172/JCI118907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Dolmetsch RE, Lewis RS, Goodnow CC, Healy JI. Differential activation of transcription factors induced by Ca2+ response amplitude and duration. Nature. 1997;386(6627):855–858. doi: 10.1038/386855a0. [DOI] [PubMed] [Google Scholar]
- 47.Graef IA, Mermelstein PG, Stankunas K, et al. L-type calcium channels and GSK-3 regulate the activity of NF-ATc4 in hippocampal neurons. Nature. 1999;401(6754):703–708. doi: 10.1038/44378. [DOI] [PubMed] [Google Scholar]
- 48.Sée V, Rajala NKM, Spiller DG, White MRH. Calcium-dependent regulation of the cell cycle via a novel MAPK-NF-κB pathway in Swiss 3T3 cells. Journal of Cell Biology. 2004;166(5):661–672. doi: 10.1083/jcb.200402136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Cullen PJ, Lockyer PJ. Integration of calcium and Ras signalling. Nature Reviews Molecular Cell Biology. 2002;3(5):339–348. doi: 10.1038/nrm808. [DOI] [PubMed] [Google Scholar]
- 50.Echevarria W, Leite MF, Guerra MT, Zipfel WR, Nathanson MH. Regulation of calcium signals in the nucleus by a nucleoplasmic reticulum. Nature Cell Biology. 2003;5(5):440–446. doi: 10.1038/ncb980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Pusl T, Wu JJ, Zimmerman TL, et al. Epidermal growth factor-mediated activation of the ETS domain transcription factor Elk-1 requires nuclear calcium. Journal of Biological Chemistry. 2002;277(30):27517–27527. doi: 10.1074/jbc.M203002200. [DOI] [PubMed] [Google Scholar]
- 52.Dolmetsch R. Excitation-transcription coupling: signaling by ion channels to the nucleus. Science’s STKE. 2003;2003(166):p. PE4. doi: 10.1126/stke.2003.166.pe4. [DOI] [PubMed] [Google Scholar]
- 53.Rodrigues MA, Gomes DA, Leite MF, et al. Nucleoplasmic calcium is required for cell proliferation. Journal of Biological Chemistry. 2007;282(23):17061–17068. doi: 10.1074/jbc.M700490200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Gomes DA, Rodrigues MA, Leite MF, et al. c-Met must translocate to the nucleus to initiate calcium signals. Journal of Biological Chemistry. 2008;283(7):4344–4351. doi: 10.1074/jbc.M706550200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Nicou A, Serrière V, Hilly M, et al. Remodelling of calcium signalling during liver regeneration in the rat. Journal of Hepatology. 2007;46(2):247–256. doi: 10.1016/j.jhep.2006.08.014. [DOI] [PubMed] [Google Scholar]
- 56.Lagoudakis L, Garcin I, Julien B, et al. Cytosolic calcium regulates liver regeneration in the rat. Hepatology. 2010;52(2):602–611. doi: 10.1002/hep.23673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Pauls TL, Cox JA, Berchtold MW. The Ca2+-binding proteins parvalbumin and oncomodulin and their genes: new structural and functional findings. Biochimica et Biophysica Acta. 1996;1306(1):39–54. doi: 10.1016/0167-4781(95)00221-9. [DOI] [PubMed] [Google Scholar]
- 58.Hardingham GE, Chawla S, Johnson CM, Bading H. Distinct functions of nuclear and cytoplasmic calcium in the control of gene expression. Nature. 1997;385(6613):260–265. doi: 10.1038/385260a0. [DOI] [PubMed] [Google Scholar]
- 59.Deisseroth K, Tsien RW. Dynamic multiphosphorylation passwords for activity-dependent gene expression. Neuron. 2002;34(2):179–182. doi: 10.1016/s0896-6273(02)00664-5. [DOI] [PubMed] [Google Scholar]
- 60.Kupzig S, Deaconescu D, Bouyoucef D, et al. GAP1 family members constitute bifunctional Ras and Rap GTPase-activating proteins. Journal of Biological Chemistry. 2006;281(15):9891–9900. doi: 10.1074/jbc.M512802200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Kornhauser JM, Cowan CW, Shaywitz AJ, et al. CREB transcriptional activity in neurons is regulated by multiple, calcium-specific phosphorylation events. Neuron. 2002;34(2):221–233. doi: 10.1016/s0896-6273(02)00655-4. [DOI] [PubMed] [Google Scholar]
- 62.Giorgi C, Baldassari F, Bononi A, et al. Mitochondrial Ca+ and apoptosis. Cell Calcium. 2012;52:36–43. doi: 10.1016/j.ceca.2012.02.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Guerra MT, Fonseca EA, Melo FM, et al. Mitochondrial calcium regulates rat liver regeneration through the modulation of apoptosis. Hepatology. 2011;54(1):296–306. doi: 10.1002/hep.24367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Mauger JP. Role of the nuclear envelope in calcium signalling. Biology of the Cell. 2012;104(2):70–83. doi: 10.1111/boc.201100103. [DOI] [PubMed] [Google Scholar]
- 65.Gomes DA, Leite MF, Bennett AM, Nathanson MH. Calcium signaling in the nucleus. Canadian Journal of Physiology and Pharmacology. 2006;84(3-4):325–332. doi: 10.1139/y05-117. [DOI] [PubMed] [Google Scholar]
- 66.Bezin S, Fossier P, Cancela JM. Nucleoplasmic reticulum is not essential in nuclear calcium signalling mediated by cyclic ADPribose in primary neurons. Pflugers Archiv European Journal of Physiology. 2008;456(3):581–586. doi: 10.1007/s00424-007-0435-8. [DOI] [PubMed] [Google Scholar]
- 67.Carrión AM, Link WA, Ledo F, Mellström B, Naranjo JR. DREAM is a Ca2+-regulated transcriptional repressor. Nature. 1999;398(6722):80–84. doi: 10.1038/18044. [DOI] [PubMed] [Google Scholar]
- 68.Weymann A, Hartman E, Gazit V, et al. P21 is required for dextrose-mediated inhibition of mouse liver regeneration. Hepatology. 2009;50(1):207–215. doi: 10.1002/hep.22979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Huang W, Ma K, Zhang J, et al. Nuclear receptor-dependent bile acid signaling is required for normal liver regeneration. Science. 2006;312(5771):233–236. doi: 10.1126/science.1121435. [DOI] [PubMed] [Google Scholar]
- 70.Csanaky IL, Aleksunes LM, Tanaka Y, Klaassen CD. Role of hepatic transporters in prevention of bile acid toxicity after partial hepatectomy in mice. American Journal of Physiology—Gastrointestinal and Liver Physiology. 2009;297(3):G419–G433. doi: 10.1152/ajpgi.90728.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Cruz LN, Guerra MT, Kruglov E, et al. Regulation of multidrug resistance-associated protein 2 by calcium signaling in mouse liver. Hepatology. 2010;52(1):327–337. doi: 10.1002/hep.23625. [DOI] [PMC free article] [PubMed] [Google Scholar]