

Abstract
Lexical-semantic access is affected by the phonological structure of the lexicon. What is less clear is whether such effects are the result of continuous activation between lexical form and semantic processing, or whether they arise from a more modular system in which the timing of accessing lexical form determines the timing of semantic activation. This study examined this issue using the visual world paradigm by investigating the time course of semantic priming as a function of the number of phonological competitors. Critical trials consisted of high or low density auditory targets (e.g. horse) and a visual display containing a target, a semantically related object (e.g. saddle), and two phonologically and semantically unrelated objects (e.g. chimney, bikini). Results showed greater magnitude of priming for semantically related objects of low than of high density words and no differences for high and low density word targets in the time course of looks to the word semantically related to the target. This pattern of results is consistent with models of cascading activation, which predict that lexical activation has continuous effects on the level of semantic activation, with no delays in the onset of semantic activation for phonologically-competing words.
Keywords: Spoken word recognition, Lexical competition, Semantic priming, Neighborhood density
Spoken word recognition is typically framed in terms of two processes: the activation of the word-form and the subsequent access of its meaning. However, the timing of these processes is uncertain. They could occur simultaneously, with activation of meaning building simultaneously with word-form activation; or sequentially, with activation of meaning beginning only after word-form activation reaches a threshold. The current study contrasts the former continuous cascade approach, from the latter modular approach.
Early theories of word recognition viewed lexical and semantic access as autonomous processes, with activation of the word-form completing before accessing meaning (Forster, 1981; Tanenhaus, Carlson & Seidenberg, 1985). This grew out of theories of modularity, suggesting that language operates as a distinct module from other cognitive systems, and that the levels of language are informationally encapsulated. In this view, word recognition is distinct from and precedes the access of meaning.
Marslen-Wilson’s (1990) cross-modal priming work provides evidence against encapsulation. He showed that words with phonological cohort competitors exhibited less semantic priming than words without competitors. These findings suggest that the degree of semantic activation depends on the degree of lexical activation of the target (which is reduced by phonological competitors). More broadly, this predicts that the number of competitors determines the strength of semantic access: words from denser phonological neighborhoods will more poorly activate semantic representations (Gaskell & Marslen-Wilson, 1997, 1999).
Zwitserlood (1989), using a gating paradigm, found that semantic associates of the words “captain” and “captive” were simultaneously activated after hearing /kæpt/, even when sentence context strongly favored one completion. This finding suggests parallel access of semantic interpretations during lexical access (Gaskell & Marslen-Wilson, 1997, 1999). Listeners do not need to hear the entire word (or even up to the uniqueness point) to begin accessing meaning, as strict autonomous perspectives predict. Instead, the meanings of both words are activated prior to disambiguation.
These studies imply that lexical competition need not be resolved before accessing meaning, and variations in activation due to competition cascade to semantic activation. While interactive activation models of spoken word recognition (e.g. TRACE, McClelland & Elman, 1986) do not attempt to model semantics, the framework underlying these models suggests cascading activation. In this view, semantic activation is a continuous function of the degree of lexical activation. All words access their semantic networks as soon as lexical access begins, with more active word-forms yielding greater semantic activation. Thus, words with greater competition (e.g. words in dense neighborhoods) will show decreased word-form activation, and thereby decreased semantic activation and decreased priming, compared to words with less competition (e.g. words in sparse neighborhoods).
The distributed cohort model (DCM: Gaskell & Marslen-Wilson, 1997, 1999) captures these notions explicitly. This model uses activation across distributed semantic and phonological units to signal lexical access rather than activation of localist lexical nodes. In this model, words with greater phonological competition do not activate their semantic representations as effectively because the overlapping word-forms result in overlapping semantic representations. Thus both word-form and semantic activation (which are not separate in the DCM) should decrease with more active cohorts, producing continuous differences in semantic activation between words in dense and sparse neighborhoods.
As a result of these empirical findings and the theoretical approaches they led to, the autonomous view has been largely abandoned in word recognition, and even many of the authors of studies favoring autonomous processing have since advanced interactive theories (e.g. Tanenhaus, Spivey-Knowlton, Eberhart & Sedivy, 1995; Joanisse & Seidenberg, 1998). However, autonomous theories are still influential in other areas of language, like speech production (Levelt, Roelofs & Meyer, 1999). Moreover, while the Marslen-Wilson (1990) and Zwitserlood (1989) findings are suggestive of continuous cascade models, such effects can be explained without continuous interaction between lexical access and semantics. Zwitserlood’s (1989) gating task used word fragments that stopped before enough information was heard to identify the target. This cessation of auditory information could cause an encapsulated word-form recognition system to pass on whatever information it currently has. What appears to be semantic activation during the early portions of the word (e.g. capt in captain) could be an artifact of unnaturally truncated word segments.
Similarly, Marslen-Wilson’s (1990) results could be accounted for by an autonomous model if all words activate their semantic networks with equal strength, but this activation is delayed until activation of the prime word reaches some threshold. While the magnitude of priming would be the same for all words, the delay in when the target word with a competitor reaches its threshold would result in later onset for semantic priming. Because reaction time in semantic priming tasks measures the time taken to reach the end-state of processing, not the time course of processing, this cannot be ruled out.
What is needed is a way to measure the ongoing amount of semantic priming. We employed the visual world paradigm (VWP) to measure semantic priming as a function of phonological neighborhood density (Tanenhaus et al, 1995). This paradigm offers a continuous estimate of the degree of lexical activation and has shown effects of phonological competition (Allopenna, Magnuson & Tanenhaus, 1998; Magnuson, Tanenhaus, Aslin & Dahan, 2003, Magnuson, Dixon, Tanenhaus & Aslin, 2007), and semantic priming (Huettig & Altmann, 2005; Yee & Sedivy, 2006; Yee, Blumstein & Sedivy, 2008). Thus, the VWP should reveal whether such differences are continuous, reflecting dynamic interaction of lexical and semantic processing; whether the magnitude of semantic activation varies as a function of lexical density; and whether semantic activation is delayed by phonological competition.
Considering the time-course of word-form recognition, a number of studies have examined the effect of phonological density on lexical access in the VWP (Magnuson et al, 2003a, 2007; Sweeney, Blumstein & Apfelbaum, 2009). They find that low density (LD) words are fixated earlier than high density (HD) words, and often exhibit more fixations than HD words (Figure 1). This suggests that decreased phonological competition leads to faster activation growth and increased total activation. These findings give rise to multiple predictions for the time course of semantic priming. Based on Marslen-Wilson’s (1990) finding of decreased priming for words sharing onsets, the patterns of semantic activation should differ between density conditions. Thus, some difference in semantic priming as a function of phonological competition is likely.
Figure 1.

Timecourse of target fixation for words low density and high density words.
Yet importantly, how these conditions differ depends on the link between word recognition and semantic access (see Figure 2). An autonomous account of the Marslen-Wilson (1990) results predicts that semantic activation is delayed until a threshold of word-form activation has been reached, while the magnitude of semantic activation increases identically for all words after this point. Figure 2A presents such a modular pattern of results – HD items access their semantic network later than LD items because of the delay in reaching threshold (as shown in Figure 1), however both sets of words ultimately show similar priming.
Figure 2.

A) The predicted magnitude of priming given a hypothesis in which competition delays semantic access, but does not affect magnitude of activation. B) The predicted magnitude of priming if semantic access is continuously dependent on degree of lexical activation. C) The predicted magnitude of priming if semantic access is both delayed and dependent on degree of lexical activation. D) The predicted magnitude of priming if semantic activation always begins at the same time, but peak magnitude of activation is dependent on degree of lexical activation.
Alternatively, Figure 2B presents a continuous cascade account, in which semantic access begins at the onset of lexical activation, but the degree of semantic activation is continuously dependent on lexical activation levels. The continuous greater activation for LD words yields faster and stronger semantic activation than for HD words.
Figure 2C combines the two approaches, with modular timing but cascading peak magnitude: semantic access does not initiate until a recognition threshold is reached, but the magnitude of semantic access thereafter depends on the level of lexical activation. HD words begin semantic access later than LD words, and show a decreased peak magnitude of semantic access. Finally, in Figure 2D semantic access begins at the same time for words regardless of their lexical access profiles, however the peak magnitude of semantic activation is dependent on degree of lexical access. While logically possible, this hypothesis lacks strong theoretical support.
Each of these accounts predicts slower reaction times for HD words in semantic priming lexical decision tasks (e.g. Marslen-Wilson, 1990) as each possesses periods when activation for the semantically-related items in LD conditions exceeds that in HD conditions. The VWP can help disentangle these predictions by measuring semantic priming over time..
METHODS
Design
Neighborhood density (Luce & Pisoni, 1998) was used to manipulate phonological competition. Target stimuli were 72 monosyllabic words, divided evenly into two non-overlapping sets of lexical density distributions. Frequency-weighted neighborhood density was measured from phonological transcriptions of the Kucera and Francis (1967) database (the sum of the log frequencies of all neighbors formed by a one-phoneme change; Luce & Pisoni, 1998). LD words had a density between 0 and 42 (M=19.47) and HD words were between 58 and 197 (M=97.12). The distributions also differed significantly by raw number of neighbors (t(36)=1.99, p<.0001). LD words had between 1 and 30 neighbors (only one word had more than 19 neighbors; M=9.25), and HD words had between 13 and 35 neighbors (M=23.97). The words did not differ on average frequency (LD=29.83, HD=38.11; t<1).
For each target item, a semantically related word was selected based on association strength values of the South Florida Free Association Norms (Nelson, McEvoy & Schreiber, 1998). There was no difference in association strength between the semantically related and target words between density conditions (t<1), nor was there a difference in the frequency (t(70)=1.52; p=.13) or number of syllables of the semantically related word (t<1). An additional 144 items were chosen as unrelated fillers. Filler items were semantically and phonologically unrelated to the target and semantically related stimuli.
Previous work suggests that conceptually similar words show greater priming than associatively related items (Huettig & Altmann, 2005). It was important to ensure that our effects could not result from differences in type of semantic relations between words across the HD and LD conditions, as it is possible that priming of associatively related words and conceptually related words may unfold via different time courses. Thus, we analyzed the similarity of the words using the WordNet::Similarity (Pederson, Patwardhan & Michelizzi, 2004) software, which measures similarity based on the number of edges between senses of words (Fellbaum, 1998). There was no difference in semantic similarity between the semantically related words in the HD and LD conditions (t<1).
Two types of control trials were randomly interwoven with experimental trials: 36 trials contained pairs of semantically-related items, but with neither as the target; and an additional 36 did not contain any semantically-related items.
Stimuli
Visual stimuli were color illustrations of objects taken from a commercial Clipart database. Images were initially selected by committee to be prototypical representations of their lexical targets. All images of targets and semantic associates were then presented to 10 naïve participants, who identified these items in a free response task. Only images appropriately named by 7 of 10 subjects were included.
A male native speaker of American English recorded the auditory stimuli. Words did not differ in overall length between density conditions (LD=372 ms; HD=360 ms; t(40)<1).
Subjects
Sixteen participants from the University of Iowa participated in exchange for partial course credit. None had participated in the free response picture norming test. All were native speakers of American English, had no history of speech or neurological disorders, and reported normal vision and hearing.
Procedure
Participants were tested in a single one-hour session. Eye movements were tracked using a head-mounted SMI EyeLink II. At the beginning of the experiment, the eye-tracker was calibrated using the standard nine-point calibration procedure. Drift correction was performed every 36 trials to account for shifts in head position or tracker movement. Fixations were monitored at 250 Hz and automatically parsed into saccades, fixations and blinks using the default Eyelink parameters. Adjacent saccades and fixations were combined into a “look”, starting at saccade onset, and ending at the fixation offset. (McMurray, Tanenhaus & Aslin, 2002).
Each trial presented four images, one in each corner of the screen, and a blue fixation dot at the center of the screen. This dot turned red after 500 ms, at which point the participant clicked it to initiate the trial. Once clicked, the dot disappeared and an auditory stimulus identified the target. Participants then clicked the named picture.
Critical trials contained a target (e.g. ‘horse’), an item semantically related to this target (e.g. ‘saddle’), and two unrelated items (see Appendix). Placement of the target, competitor, and unrelated foils on the screen was randomized across trials. Unrelated items were counterbalanced between subjects, such that unrelated items in high density trials for one half of subjects were then used as low density filler items for the other half of subjects. Thus effects of density could not arise from differential salience of unrelated items in one condition.
Appendix.
Stimuli for Experimental Trials
| Low Density | High Density | ||||||
|---|---|---|---|---|---|---|---|
| Target | Relative | Control | Control | Target | Relative | Control | Control |
| beard | mustache | pick | rug | badge | police | cart | hopscotch |
| bridge | tunnel | lips | dart | bee | honey | veterinarian | stove |
| broom | mop | robin | tailor | beer | party | teacher | skate |
| brush | comb | helmet | jam | bird | nest | drill | matchbook |
| cliff | mountain | pillow | nurse | boot | cowboy | pencil | rock |
| clown | circus | onion | ram | bow | arrow | sundae | pope |
| cord | phone | mug | shuttle | bowl | spoon | chin | tack |
| crutch | cast | horn | medal | bus | taxi | frame | dice |
| desk | lamp | frog | wallet | cake | chocolate | paddle | radar |
| dog | cat | baggage | yolk | chair | table | penny | road |
| dress | girl | fin | plane | chick | egg | batter | ice |
| film | camera | chalk | general | cone | ice cream | jeep | pill |
| glove | baseball | fireplace | pitchfork | deer | antler | pumpkin | shack |
| grill | hamburger | nickel | robe | door | window | pedal | referee |
| harp | angel | rabbit | curtains | feet | shoes | radio | pole |
| hook | fish | clover | punch | goat | sheep | cork | knot |
| horse | saddle | chimney | bikini | gun | bullet | football | water |
| ink | pen | root | soccer | hose | garden | kick | waffles |
| jar | cookie | snowman | puck | hut | tent | peacock | berry |
| knife | fork | parachute | whip | knight | soldier | tadpole | blocks |
| milk | cow | checkbook | surfer | leaf | plant | moat | racket |
| noose | rope | picnic | steam | lock | key | pizza | blood |
| pipe | cigar | spatula | wig | moon | sun | beans | captain |
| plug | outlet | grasshopper | bib | nail | hammer | chess | windmill |
| priest | collar | basketball | soup | net | volleyball | pepper | whisk |
| raft | boat | cabbage | peanut | nun | church | pacifier | race |
| scarf | hat | carrier | alligator | patch | jeans | lightbulb | cable |
| shield | sword | braid | fan | pie | apple | rocker | thread |
| shrimp | lobster | bandaid | gate | plate | cup | ladder | geek |
| skull | brain | muffin | pot | rake | leaves | sandal | check |
| snake | grass | jet | radiator | rice | corn | grave | package |
| swing | park | raspberry | toad | shoe | sock | tail | restaurant |
| thorn | rose | hanger | fireworks | suit | tie | bug | palm tree |
| throne | king | beetle | rain | tire | car | dove | lettuce |
| truck | van | hammock | pin | tooth | dentist | bale | hamster |
| wine | cheese | scissors | rocking chair | vase | flower | dynamite | note |
Participants completed four practice trials prior to the experiment. The items in the practice trials were all semantically and phonologically unrelated.
RESULTS
One subject received only 139 trials due to a computer error. All others completed 144 trials. Trials in which the subject did not select the target were excluded. Across all subjects, this totaled 33 trials (of 1149 experimental trials, 2.9%).
The proportion of trials in which each picture type (target, semantically related, and unrelated) was fixated was computed every 4 ms for each density condition. Since it takes approximately 200 ms to program a saccade (Matin, Shao & Boff, 1993), and the auditory target was preceded by 100 ms of silence, only fixations initiated 300 ms after stimulus onset were examined. Analysis ended 1000 ms after trial onset (average length of the auditory stimuli was 529 ms). This was selected because looks to the target declined after this window, indicating completed lexical access.
Throughout the time course of processing, semantically related words were fixated more than unrelated words for both density conditions (see Figure 3). There were also fewer looks to the semantically related item in the HD than LD condition, suggesting a reduction in the magnitude of semantic priming in this condition.
Figure 3.
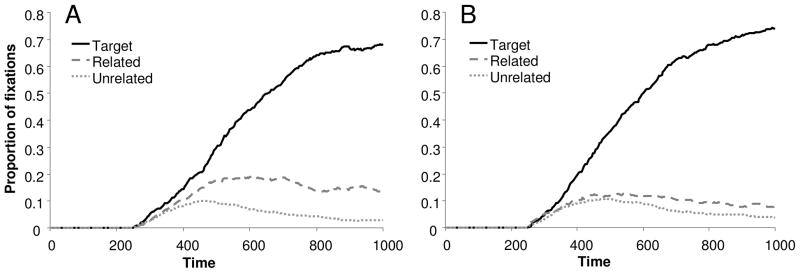
The proportion of trials on which subjects were fixating the target, semantically related item and unrelated pictures as a function of time. A) Trials in which the stimulus was a low density word. B) High density trials.
To examine the time course of semantic priming, the magnitude of priming (MoP) was computed by subtracting the proportion of looks to the average of the two unrelated items from the proportion of looks to the semantically related item. This parallels the MoP measure used in traditional semantic priming paradigms. Logistic functions were fit to MoP as a function of time (see McMurray, Samelson, Lee & Tomblin, 2010). Logistic functions are defined by four values: the minimum, maximum (upper asymptote of MoP), slope (how rapidly the function approaches its asymptote), and crossover point (the midpoint of the rising portion of the curve – this gives a measure of the timing of MoP). In terms of the hypotheses outlined in Figure 2, the factors of primary interest are the maximum, which will reveal if the MoP differs between conditions (predicted by cascade accounts); the crossover, which will reveal if activation of the semantic associate is delayed for one condition (predicted by autonomous accounts); and the slope, which will reveal if one condition more rapidly approaches its peak MoP.
Slope differences may be difficult to disentangle as a shallower slope can arise from averaging across subjects with steep but variable boundaries, or from a genuinely shallower slope. Moreover, our analysis relied on a jackknife procedure (see below) that makes differentiating these sources difficult. However, the predictions of the non-autonomous accounts (Figures 2B, D) predict a lower maximum MoP for the HD condition; and the autonomous accounts (Figures 2A,C) predict a later crossover point for HD items, as they predict that phonological competition delays semantic access. Thus, these two factors alone may be sufficient.
Because each subject completed very few trials per condition (36), it was difficult to obtain accurate fits for individual subjects1. Thus, we adapted the jackknife method of Miller, Paterson and Ulrich (1998; see McMurray, Clayards, Tanenhaus & Aslin, 2008, for a VWP application) by first computing the average MoP in the entire dataset, excluding a single subject. We then fit the logistic function to this data and extracted the slope, crossover and asymptotes. This was repeated, excluding each subject in turn. The slope, crossover and asymptotes can be compared across conditions using standard T statistics, with conservative adjusted error terms..
Figure 4 shows the MoP data plotted along with the mean logistic fits. Curve fits were excellent (mean R2=.97; range: .93 to .99). Both conditions showed significant priming, as indicated by a maximum MoP greater than 0 (HD: t1(15)=5.4, p<.01; t2(35)=20.1, p<.01; LD: t1(15)=7.2, p<.01; t2(35)=31.2, p<.01). However, the LD condition attained a higher overall magnitude of priming than the HD condition across time (t1(15)=4.3, p<.01; t2(70)=2.9, p<.01). The crossover point did not differ significantly (t1<1, t2<1), though there was a slight trend toward later activation for the high density items (HD: M=510, SD=125, LD: M=479, SD=28).Similarly, the slope did not differ significantly (t1(15)=1.08, p=.30; t2(70)=1.29, p=.20), though differences trended in the direction predicted by the continuous cascade account, with steeper activation increases in the LD condition (HD: M=.00022, SD=.00033; LD: M=.00081, SD=.00036). Overall, the primary effect of neighborhood density was in the magnitude of semantic priming, while the timing and rate of semantic activation was less affected.
Figure 4.

Time course of the magnitude of the semantic priming effect in each density condition. Mean logistic functions are overlaid on the raw data.
DISCUSSION
The current study demonstrated that the size or density of a word’s phonological neighborhood affects the magnitude of semantic access. HD target stimuli elicited fewer looks to pictures that were semantically related than did LD stimuli. The timing of these looks did not differ between conditions, yet LD words showed a greater peak level of activation. Thus, phonological competition affects the strength of activation of semantically-related items across time during word recognition. These findings support earlier work (Marslen-Wilson, 1990; Zwiterlood, 1989) and provide evidence for cascading activation between phonological and semantic processes during lexical access.
Considering our results in light of the predictions presented in Figure 2, at least two can be ruled out. Our data cannot be accounted for by autonomous accounts predicting that phonological competition delays the onset of semantic activation (Figure 2A), but does not affect the magnitude, as the peak MoP differed, while the timing did not. The combination of delay in onset of activation and decreased peak activation (Figure 2C) also fails to account for the similar timing of activation between conditions.
This leaves the accounts predicting similar timing across conditions, but different maximum levels of priming (Figures 2B and 2D). The nonsignificant slope difference seems to support the predictions of 2D, that semantic access always begins at the same time, while the peak MoP reflects degree of lexical activation. However, this account may be untenable. First, it predicts that the onset of priming is delayed a fixed amount for all words. However, as seen in Figure 4, semantic priming begins very quickly after the onset of the word, well before the entire word can be processed (given the 200ms oculomotor delay). Thus semantic access does not appear to have a fixed delay, but instead appears to begin almost simultaneously with lexical access. The lack of a sound theoretical backing for this account further suggests that the nonsignificant slope effect may be an issue of statistical power, rather than an argument against the cascade approach (as previously discussed, the jackknife analysis may have obscured true slope effects). Finally, given the theoretical backing for the hypothesis of activation continuously cascading from lexical to semantic levels and the trend toward different slopes between the conditions, this hypothesis appears the most tenable; even if the slope effect proves illusory, some form of cascading activation appears necessary to explain the peak MoP results.
These results challenge autonomous models in which gradations in activation affect only the current level of processing and have limited influence on later stages (e.g. Levelt et al, 1999). They are consistent with interactive activation approaches to word recognition (McClelland & Elman, 1986) in which activation levels continuously influence other layers. They are especially consistent with the predicted interaction between activation levels and semantic access in DCM (Gaskell & Marslen-Wilson, 1997, 1999). Differences in the degree of phonological competition affect the activation of a target item and have a cascading effect on the semantic activation, with words in dense neighborhoods showing decreases semantic priming.
However, while our results are largely consistent with DCM, one area shows discrepancy. DCM employs distributed lexical/semantic representations which create limitations on the number of candidates that can be effectively evaluated. As such, increasing the number of coactive candidates weakens semantic activation, and with a large enough cohort, semantic activation disappears.. When lexical information is ambiguous between two words, activation across these nodes is averaged between the words’ semantic representations. If many lexical candidates with different semantic representations are active, the activation in semantic space becomes uninformative due to this averaging. While the current study showed decreased priming for HD words, in accordance with the predicted decrease in activation in DCM, even our HD items yielded significant semantic priming. This suggests that either the ceiling for competition is beyond the mean of 24 competitors in the HD condition, or the mechanisms of competition in DCM are not appropriately structured to explain semantic activation in dense neighborhoods. A sparser (or even localist) representation may be required.
This study demonstrates that phonological competition has cascading effects on semantic access, such that words with high degrees of phonological competition show decreased access to their semantic networks compared to words from sparser neighborhoods. This adds to the body of evidence arguing for real-time interactions between lexical processes and other domains of cognition and perception (e.g. Dahan & Tanenhaus, 2004; Magnuson, McMurray, Tanenhaus & Aslin, 2003; Revill, Tanenhaus & Aslin, 2008; Levy, Bicknell, Slattery & Rayner, 2009).
Acknowledgments
This research was supported in part by NIH Grant DC00314 to Sheila Blumstein and DC008089 to Bob McMurray. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institute on Deafness and Other Communication Disorders or the National Institutes of Health. Many thanks to James Magnuson for providing estimates of neighborhood density.
Footnotes
We attempted to fit logistics to individual subject’s data, but the sparseness of the data for each subject led to very poor fits; we further attempted a non-linear mixed-effects model, and still found poor fits.
Contributor Information
Keith S. Apfelbaum, Dept. of Psychology, University of Iowa
Sheila E. Blumstein, Dept. of Cognitive and Linguistics Sciences, Brown University
Bob McMurray, Dept. of Psychology, University of Iowa.
References
- Allopenna PD, Magnuson JS, Tanenhaus MK. Tracking the time course of spoken word recognition using eye movements: Evidence for continuous mapping models. Journal of Memory and Language. 1998;38(4):419–439. [Google Scholar]
- Dahan D, Tanenhaus MK. Continuous mapping from sound to meaning in spoken-language comprehension: Immediate effects of verb-based thematic constraints. Journal of Experimental Psychology: Learning, Memory and Cognition. 2004;30(2):493–513. doi: 10.1037/0278-7393.30.2.498. [DOI] [PubMed] [Google Scholar]
- Fellbaum C, editor. WordNet: An Electronic Lexical Database. Cambridge, MA: MIT Press; 1998. [Google Scholar]
- Forster KI. Priming and the effects of sentence and lexical contexts on naming time: Evidence for autonomous lexical processing. Quarterly Journal of Experimental Psychology. 1981;33(A):465–495. [Google Scholar]
- Gaskell MG, Marslen-Wilson WD. Integrating form and meaning: a distributed model of speech perception. Language and Cognitive Processes. 1997;12:613–656. [Google Scholar]
- Gaskell MG, Marslen-Wilson WD. Ambiguity, competition, and blending in spoken word recognition. Cognitive Science. 1999;23:439–462. [Google Scholar]
- Huettig F, Altmann GTM. Word meaning and the control of eye fixation: Semantic competitor effects and the visual world paradigm. Cognition. 2005;96:B23–B32. doi: 10.1016/j.cognition.2004.10.003. [DOI] [PubMed] [Google Scholar]
- Joanisse MF, Seidenberg MS. Specific language impairment: A deficit in grammar or processing? Trends in Cognitive Sciences. 1998;2(7):240–247. doi: 10.1016/S1364-6613(98)01186-3. [DOI] [PubMed] [Google Scholar]
- Kucera H, Francis WN. Computational Analysis of Present Day American English. Providence, RI: Brown University Press; 1967. [Google Scholar]
- Levelt WJM, Roelofs A, Meyer AS. A theory of lexical access in speech production. Behavioral and Brain Sciences. 1999;22:1–75. doi: 10.1017/s0140525x99001776. [DOI] [PubMed] [Google Scholar]
- Levy R, Bicknell K, Slattery T, Rayner K. Eye movement evidence that readers maintain and act on uncertainty about past linguistic input. Proceedings of the National Academy of Sciences. 2009;106(50):21086–21090. doi: 10.1073/pnas.0907664106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luce PA, Pisoni DB. Recognizing spoken words: the neighborhood activation model. Ear and Hearing. 1998;19(1):1–36. doi: 10.1097/00003446-199802000-00001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Magnuson J, McMurray B, Tanenhaus M, Aslin R. Lexical effects on compensation for coarticulation: The ghost of Christmash past. Cognitive Science. 2003;27(2):285–298. [Google Scholar]
- Magnuson JS, Tanenhaus MK, Aslin RN, Dahan D. The time course of spoken word learning and recognition: Studies with artificial lexicons. Journal of Experimental Psychology: General. 2003a;132(2):202–227. doi: 10.1037/0096-3445.132.2.202. [DOI] [PubMed] [Google Scholar]
- Magnuson JS, Dixon JA, Tanenhaus MK, Aslin RN. The dynamics of lexical competition during spoken word recognition. Cognitive Science. 2007;31(1):133–156. doi: 10.1080/03640210709336987. [DOI] [PubMed] [Google Scholar]
- Marslen–Wilson WD. Activation, competition, and frequency in lexical access. In: Altmann GTM, editor. Cognitive models of speech processing: Psycholinguistic and computational perspectives. Cambridge, MA: MIT Press; 1990. pp. 148–172. [Google Scholar]
- Matin E, Shao KC, Boff KR. Saccadic overhead: information-processing time with and without saccades. Perception and Psychophysics. 1993;53(4):372–380. doi: 10.3758/bf03206780. [DOI] [PubMed] [Google Scholar]
- McClelland JL, Elman JL. The TRACE model of speech perception. Cognitive Psychology. 1986;18:1–86. doi: 10.1016/0010-0285(86)90015-0. [DOI] [PubMed] [Google Scholar]
- McMurray B, Clayards M, Tanenhaus M, Aslin R. Tracking the timecourse of phonetic cue integration during spoken word recognition. Psychonomic Bulletin and Review. 2008;15(6):1064–1071. doi: 10.3758/PBR.15.6.1064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McMurray B, Samelson V, Lee S, Tomblin JB. Eye-movements reveal the time-course of online spoken word recognition language impaired and normal adolescents. Cognitive Psychology. 2010;60(1):1–39. doi: 10.1016/j.cogpsych.2009.06.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McMurray B, Tanenhaus MK, Aslin RN. Gradient effects of within-category phonetic variation on lexical access. Cognition. 2002;86(2):B33–B42. doi: 10.1016/s0010-0277(02)00157-9. [DOI] [PubMed] [Google Scholar]
- Miller J, Patterson T, Ulrich R. Jackknife-based method for measuring LRP onset latency differences. Psychophysiology. 1998;35(1):99–115. [PubMed] [Google Scholar]
- Nelson DL, McEvoy CL, Schreiber TA. The University of South Florida word association, rhyme, and word fragment norms. 1998 doi: 10.3758/bf03195588. http://www.usf.edu/FreeAssociation/ [DOI] [PubMed]
- Pederson T, Patwardhan S, Michelizzi J. WordNet::Similarity – Measuring the Relatedness of Concepts. Proceedings of the the Fifth Annual Meeting of the North American Chapter of the Association for Computational Linguistics,; 2004. pp. 38–41. [Google Scholar]
- Revill KP, Tanenhaus MK, Aslin RN. Context and spoken word recognition in a novel lexicon. Journal of experimental psychology: Learning, memory, and cognition. 2008;34(5):1207–1223. doi: 10.1037/a0012796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sweeney C, Blumstein SE, Apfelbaum KS. The neural bases of phonological competition: Evidence from aphasia. Poster presented at the Annual Meeting of the Academy of Aphasia; Boston, MA. 2009. [Google Scholar]
- Tanenhaus MK, Carlson GN, Seidenberg MS. Do listeners compute linguistic representations? In: Dowty DR, Kartunnen L, Zwicky AM, editors. Natural language parsing: Psycholinguistic, theoretical and computational perspectives. Cambridge: Cambridge University Press; 1985. pp. 359–408. [Google Scholar]
- Tanenhaus MK, Spivey-Knowlton MJ, Eberhard KM, Sedivy JC. Integration of visual and linguistic information in spoken language comprehension. Science. 1995;268:1632–1634. doi: 10.1126/science.7777863. [DOI] [PubMed] [Google Scholar]
- Yee E, Sedivy JC. Eye movements to pictures reveal transient semantic activation during spoken word recognition. Journal of Expermental Psychology: Learning, Memory, and Cognition. 2006;32:1–14. doi: 10.1037/0278-7393.32.1.1. [DOI] [PubMed] [Google Scholar]
- Yee E, Blumstein SE, Sedivy JC. Lexical-semantic activation in Broca’s and Wernicke’s aphasia: Evidence from eye movements. Journal of Cognitive Neuroscience. 2008;20(4):592–612. doi: 10.1162/jocn.2008.20056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zwitserlood P. The locus of the effects of sentential-semantic content in spoken-word processing. Cognition. 1989;32(1):25–64. doi: 10.1016/0010-0277(89)90013-9. [DOI] [PubMed] [Google Scholar]