

Abstract
There has been a remarkable growth of papers published about purinergic signalling via ATP since 1972. I am most grateful to the wonderful PhD students and postdoctoral fellows who have worked with me over the years to pursue the purinergic hypothesis despite early opposition and to the many outstanding scientists around the world who are currently extending the story. Recently, therapeutic approaches to pathological disorders include the development of selective P1 and P2 receptor subtype agonists and antagonists, as well as of inhibitors of extracellular ATP breakdown and of ATP transport enhancers and inhibitors. Medicinal chemists are starting to develop small molecule purinergic drugs that are orally bioavailable and stable in vivo.
Keywords: ATP, adenosine, purinoceptors, pain, stroke, cancer
Sir John Gaddum was a great scientist and one of the founders of pharmacology in the UK. He was particularly interested in signalling molecules and made important contributions to our knowledge of substance P, 5-HT and histamine, as well as acetylcholine and noradrenaline. Therefore, I would like to think that he would have been interested to hear the story about another signalling molecule, namely ATP, that I am about to describe.
The beginnings
My PhD was about fish gut motility that involved simple techniques of organ bath pharmacology and histology. I needed to learn more sophisticated techniques and Wilhelm Feldberg kindly invited me to join his Department of Physiology at the Medical Research Institute, Mill Hill, to learn electrophysiology in 1957. He put me together with Ralph Straub, and together we developed the sucrose gap technique to record correlated changes in electrical and mechanical activity of smooth muscle (Burnstock and Straub, 1958). When Edith Bülbring, who ran the leading smooth muscle laboratory in the UK at that time, heard about our results, she invited me to join her group in the Department of Pharmacology, Oxford University, where they had been struggling with microelectrode recording from spontaneously active smooth muscle cells. There, I studied the effect of the classical neurotransmitters ACh and noradrenaline (NA) on the guinea pig taenia coli preparation (Burnstock 1958a,b), which was the favourite experimental model of an innervated smooth muscle preparation used by her group.
I then spent a year with Ladd Prosser at the University of Illinois on a Rockefeller Fellowship before accepting a Senior Lectureship in the Department of Zoology in Melbourne, Australia. Soon after, I set up the sucrose gap apparatus and began a very enjoyable collaboration in the Department of Physiology with Mollie Holman, whom I had met in Oxford. One day, together with my first postgraduate students, Graham Campbell and Max Bennett, we decided to transmurally stimulate the taenia coli in the presence of atropine and bretylium to block cholinergic and adrenergic neurotransmission. We were expecting to see direct stimulation of the smooth muscle resulting in depolarization and contraction. However, to our surprise and excitement, the response was hyperpolarization to single pulses and relaxation (Figure 1A). We felt that we were onto something important (see Burnstock, 2004) and the interpretation was discussed internationally. I was fortunate in having at that time, in the early 1960s, a Japanese postdoctoral fellow, whose friend in Japan was one of the authors of a paper about tetrodotoxin (TTX) extracted from the puffer fish. He sent us some TTX, which was known to block nerve conduction, but not affect smooth muscle activity. It completely blocked the hyperpolarizations (Figure 1B), so we realized that they were inhibitory junction potentials (IJPs) in response to non-adrenergic, non-cholinergic (NANC), neurotransmission (Burnstock et al., 1964). Later, during a sabbatical visit to the School of Pharmacy in London, I worked together with Mike Rand to show that the NANC responses were mediated by intrinsic inhibitory neurones that were innervated by vagal and sacral parasympathetic nerves (Burnstock et al., 1966).
Figure 1.
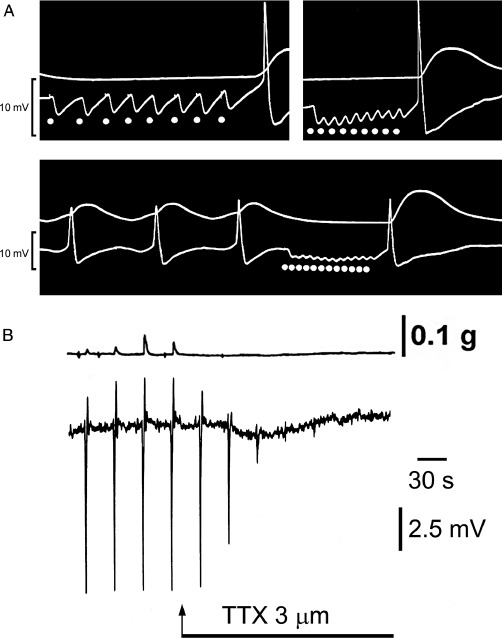
(A) IJPs recorded in smooth muscle of the atropinized guinea pig taenia coli in response to transmural stimulation of the intramural nerves remaining following degeneration of the adrenergic nerves by treatment of the animal with 6-hydroxydopamine (250 mg·kg−1 i.p. for 2 successive days) 7 days previously. Upper trace, mechanical record. Lower trace, changes in membrane potential recorded with a sucrose-gap method. The junction potentials recorded with this method are qualitatively, but not quantitatively, similar to those recorded with intracellular microelectrodes. Top left-hand panel: responses to low-frequency stimulation (l s−1). Note the long latency and slow time course of individual IJPs, and rebound excitation (spike and contraction) following cessation of stimulation. Top right-hand panel: response to a stimulation frequency of 3 s−1. Note of the summation of individual IJPs and rebound contraction. Bottom panel: response of a spontaneously active preparation to stimulation at 4 s. Note that the hyperpolarization maintained during repetitive stimulation raises the membrane potential beyond the zone of spontaneous initiation of action potentials and leads to relaxation. Rebound contraction follows cessation of stimulation. [Reproduced from Burnstock (1972), with permission from ASEPT.] (B) Sucrose gap recording of membrane potential changes in smooth muscle of guinea pig taenia coli in the presence of atropine (0.3 µM) and guanethidine (4 µM). Transmural field stimulation (0.5 ms, 0.033 Hz, 8 V) evoked transient hyperpolarizations, which were followed by rebound depolarizations. Tetrodotoxin (TTX, 3 µM) added to the superfusing Kreb's solution (applied at arrow) rapidly abolished the response to transmural field stimulation establishing these as inhibitory junction potentials in response to non-adrenergic, non-cholinergic neurotransmission. [Reproduced from Burnstock (1986), reproduced with kind permission of Blackwell Publishing.]
The next obvious question was to try to identify the transmitter involved in NANC neurotransmission. We followed the advice of Sir John Eccles that several criteria needed to be satisfied to identify a neurotransmitter: it had to be synthesized and stored in the nerve terminals; exogenous application should produce responses that mimicked those to nerve stimulation; the transmitter should be shown to be released during nerve stimulation, probably by a Ca2+-dependent mechanism; that there should be either an ectoenzymatic inactivation of the released transmitter or an uptake mechanism for the inactivation; and lastly, to identify an antagonist that blocked both the response to nerve stimulation and the exogenously applied transmitter candidate. We studied everything we could think of, neuropeptides, monoamines, amino acids, but none of them satisfied the criteria. However, I came across two important papers, one by Drury and Szent-Györgyi (1929) that described extracellular actions of purines on the heart and blood vessels and a later paper by Pamela Holton (Holton, 1959) that showed release of ATP during antidromic stimulation of sensory nerves. Together with David Satchell, we showed that ATP satisfied the criteria for both NANC inhibitory neurotransmission in the gut and also NANC excitatory neurotransmission in the urinary bladder (Figure 2; Burnstock et al., 1970). In a bold review, two years later, I invented the word ‘purinergic’, ATP being a purine nucleotide, and came up with the purinergic hypothesis (Figure 3; Burnstock, 1972).
Figure 2.

(A) Left-hand side: responses of the guinea pig taenia coli to NANC nerve stimulation (NS, 1 Hz, 0.5 ms pulse duration, for 10 s at supramaximal voltage) mimicked by ATP (2 × 10−6 M). The responses consist of a relaxation, followed by a ‘rebound contraction’. Atropine (1.5 × 10−7 M), guanethidine (5 × 10−6 M) and sodium nitrite (7.2 × 10−4 M) were present. [From Burnstock and Wong (1978), reproduced with kind permission of the Nature Publishing Group.] Right-hand side: a comparison of the NANC contractile responses of the guinea pig bladder strip to intramural nerve stimulation (NS: 5 Hz, 0.2 ms pulse duration and supramaximal voltage) mimicked by exogenous ATP (8.5 µM). Atropine (1.4 µM) and guanethidine (3.4 µM) were present throughout. [From Burnstock et al. (1978), reproduced with kind permission of the Nature Publishing Group.] (B) Effect of changing the calcium ion (Ca2+) concentration on the release of ATP (measured with the firefly luciferin/luciferase technique) from the guinea pig isolated bladder strip during stimulation of NANC nerves. Upper trace: mechanical recording of changes in tension (g) during intramural nerve stimulation (NS: 20 Hz, 0.2 ms pulse duration, supramaximal voltage for 20 s). Lower trace: concentration of ATP in consecutive 20 s fractions of the superfusate. The Ca2+ concentration in the superfusate varied as follows: (i) 2.5 mM (normal Krebs); (ii) 0.5 mM; (iii) 0.25 mM; and (iv) 2.5 mM. The successive contractions were separated by 60 min intervals as indicated by the breaks in the mechanical trace. Atropine (1.4 µM) and guanethidine (3.4 µM) were present throughout. [From Burnstock et al. (1978), reproduced with kind permission of the Nature Publishing Group.] (C) The effect of α,β-methylene ATP (α,β-meATP) receptor desensitization on the responses to nerve stimulation (↑), ATP (▵) and histamine (Hist). Atropine (1 µM) and guanethidine (3.4 µM) were present throughout. (Top panel) Control responses. (Middle panel) α,β-meATP desensitization, reached by five successive applications (▴) at approximately 4 min intervals, completely abolished nerve-mediated (↑) and ATP-induced (▵) responses. However, histamine-induced contraction is only slightly reduced. (Bottom panel) Responses 60 min after washing with Krebs. [Reproduced from Kasakov and Burnstock (1983), with permission.]
Figure 3.

Purinergic neuromuscular transmission depicting the synthesis, storage, release and inactivation of ATP. ATP, stored in vesicles in nerve varicosities, is released by exocytosis to act on post-junctional receptors for ATP on smooth muscle. ATP is broken down extracellularly by ATPases and 5′-nucleotidase to adenosine, which is taken up by varicosities to be resynthesized and reincorporated into vesicles. Adenosine is broken down further by adenosine deaminase to inosine and hypoxanthine and removed by the circulation. [From Burnstock (1972), reproduced with permission from the American Society for Pharmacology and Experimental Therapeutics.]
Unfortunately, this concept met with almost universal opposition during the following 20 years. This was perhaps not surprising because ATP was well established as an intracellular energy source and it seemed unlikely that such a ubiquitous molecule would also be involved in extracellular signalling (Figure 4). When I left Australia for University College London in 1975, at my farewell party, the Professor of Medicine introduced me as ‘the inventor of the purimagine hypothesis’. Workshops were organized at international meetings where two or three opponents were each given 10 min to explain their opposition to purinergic signalling, while I had 10 min to defend it. Ulf von Euler was with me on one occasion when a scientist said, ‘I am going to devote my life to destroying the purinergic hypothesis’. Von Euler gave me some exceptionally good advice: ‘I do not know whether your hypothesis is correct or not but firstly, negative people vanish and secondly, you must be very careful under pressure, not to make new data fit your hypothesis but look very carefully at the criticism. If it is emotion-based, forget it, but if new experiments are presented, take them seriously and be scrupulously objective in seeing if they fit yours, or any other hypothesis.’
Figure 4.
ATP communicating between cells. The image of cells with windows indicates sites of release of ATP, while doors on cells represent receptor sites, which, when opened, lead to changes in the activities of the cell.
Purinergic co-transmission
Von Euler's advice was relevant soon after, when on sabbatical leave at UCLA with Che Su and John Bevan, we discovered that ATP was released not only from NANC nerves in the taenia coli but also from sympathetic nerves supplying smooth muscle (Su et al., 1971). I was initially devastated by this new finding and stayed up all night writing a rejection of my hypothesis, but as the sun rose in the morning, I thought, could it be that ATP was being released as a co-transmitter with NA? Later, after my arrival at UCL, I published another controversial paper that challenged what was known as Dale's principle – one nerve, one transmitter (although what Dale actually proposed was that the same transmitter was released from both central and peripheral terminals of primary sensory neurones). My Commentary in Neuroscience was entitled: ‘Do some nerve cells release more than one transmitter?’ (Burnstock, 1976). It is interesting that when Mollie Holman and I recorded excitatory junction potentials (EJPs) in smooth muscle cells of the guinea pig vas deferens in response to stimulation of sympathetic nerves in 1970 (Burnstock and Holman, 1960; 1961; Figure 5A). We were puzzled that the EJPs were not abolished by adrenoceptor antagonists, NA being assumed to be the sole neurotransmitter in sympathetic nerves at that time. It was not until many years later when Peter Sneddon joined me that we showed that α,β-methylene ATP (α,β-meATP), which had been shown to desensitize the receptor (Kasakov and Burnstock, 1983), blocked the EJPs (Sneddon and Burnstock, 1984a,b). Thus, it became clear that ATP was released as a co-transmitter with NA from sympathetic nerves (Figures 5B, C and 6; Burnstock, 1990). The co-transmitter concept was also initially resisted, but it is now well established that every nerve, in both the peripheral and the central nervous system, utilizes ATP as a co-transmitter (Table 1; Burnstock, 2012b).
Figure 5.
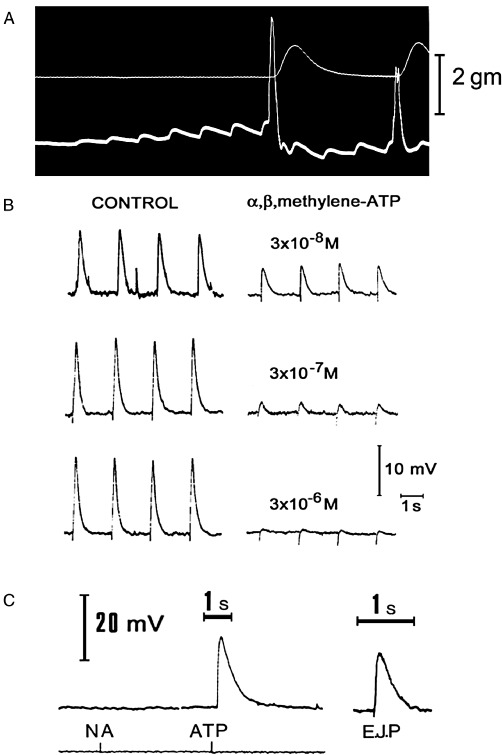
(A) Excitatory junction potentials in response to repetitive stimulation of adrenergic nerves (white dots) in the guinea pig vas deferens. The upper trace records the tension, the lower trace the electrical activity of the muscle recorded extracellularly by the sucrose gap method. Note both summation and facilitation of successive junction potentials. At a critical depolarization threshold, an action potential is initiated which results in contraction. [From Burnstock and Costa (1975), reproduced with permission of Chapman and Hall.] (B) The effect of various concentrations of α,β-methylene ATP (α,β-meATP) on excitatory junction potentials (EJPs) recorded from the guinea pig vas deferens (intracellular recording). The control responses to stimulation of the motor nerves at 0.5 Hz are shown on the left. After at least 10 min in the continuous presence of the indicated concentration of α,β-meATP, EJPs were recorded using the same stimulation parameters. The EJPs are clearly reduced in magnitude in the presence of α,β-meATP. Notice also that in the control cells, several large spontaneous EJPs were seen, whereas after α,β-meATP, no spontaneous EJPs were recorded. [Reproduced from Sneddon and Burnstock (1984a), with permission of Elsevier.] (C) Spritzed ATP, but not NA, mimicked the EJP recorded in the vas deferens. [Reproduced from Burnstock and Verkhratsky (2012) (Book Chapter 6) with permission of Springer.]
Figure 6.
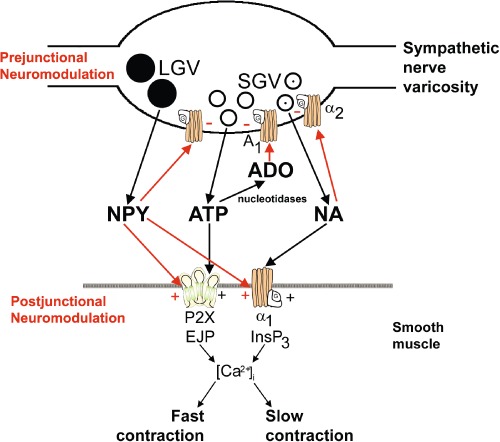
Schematic of sympathetic co-transmission. ATP and NA released from small granular vesicles (SGV) act on P2X and α1 receptors on smooth muscle respectively. ATP acting on inotropic P2X receptors evokes excitatory junction potentials (EJPs), increase in intracellular calcium ( [Ca2+]i) and fast contraction, while occupation of metabotropic α1 adrenoceptors leads to production of inositol triphosphate (InsP3), increase in [Ca2+]i and slow contraction. Neuropeptide Y (NPY) stored in large granular vesicles (LGV) acts on release both as a prejunctional inhibitory modulator of release of ATP and NA and as a post-junctional modulatory potentiator of the actions of ATP and NA. Nucleotidases are released from nerve varicosities and are also present as ectonucleotidases. [Reproduced from Burnstock and Verkhratsky (2010b) and reproduced with permission from Elsevier.]
Table 1.
Table showing co-transmitters in the peripheral and central nervous systems [Modified from Abbracchio et al. (2009), with permission]
| Co-transmitters | References | |
|---|---|---|
| Peripheral nervous system | ||
| Sympathetic nerves | ATP + NA + NPY | Westfall et al., 1978; Burnstock, 1990 |
| Parasympathetic nerves | ATP + Ach + VIP | Hoyle, 1996 |
| Sensory-motor | ATP + CGRP + SP | Burnstock, 1993 |
| NANC enteric nerves | ATP + NO + VIP | Belai and Burnstock, 1994; Burnstock, 2001 |
| Motor nerves (in early development) | ATP + ACh | Silinsky and Hubbard, 1973; Henning, 1997 |
| Central nervous system | ||
| Cortex, caudate nucleus | ATP + ACh | Richardson and Brown, 1987 |
| Hypothalamus, locus coeruleus | ATP + NA | Sperlágh et al., 1998; Poelchen et al., 2001 |
| Hypothalamus, dorsal horn, retina | ATP + GABA | Jo and Role, 2002 |
| Mesolimbic system | ATP + DA | Krügel et al., 2003 |
| Hippocampus, dorsal horn | ATP + glutamate | Mori et al., 2001; Fujii et al., 2004 |
CGRP, calcitonin gene-related peptide; VIP, vasoactive intestinal peptide.
Purinergic receptors
The next conceptual step was to identify the membrane receptors that respond to purine nucleotide and nucleoside messengers. In 1978, I recognized from hints in the literature and some simple experiments that there were different receptor families for adenosine (called P1 receptors) and for ATP and ADP (called P2 receptors) (Burnstock, 1978). P1, but not P2 receptors, were antagonized by methylxanthines. This helped resolve some of the earlier ambiguities in the literature. It was known early that there were ectoenzymes that rapidly degraded ATP to adenosine, but it was not clear whether a response to ATP was mediated by P2 or by P1 receptors after breakdown to adenosine. This is illustrated in Figure 7, which shows an update of the original model of purinergic neurotransmission. Figure 8 shows the interpretation, in retrospect, of a Loewi-inspired experiment carried out in 1966. Based on pharmacology, P2 receptors were subdivided into P2X and P2Y families (Burnstock and Kennedy, 1985). However, the turning point for widespread acceptance of purinergic signalling came in the early 1990s when the receptors to purines and pyrimidines were cloned and characterized. First, four P1 receptor subtypes were identified: A1, A2A, A2B and A3 (see Daly, 1985; Fredholm et al., 2001; Table 2). In 1993, I persuaded my old friend Eric Barnard, an expert in the cloning of nicotinic receptors, to collaborate with me to clone an ATP receptor, a G protein-coupled receptor which we named P2Y1 (Webb et al., 1993), at about the same time as David Julius and his colleagues in San Francisco cloned a P2Y2 receptor (Lustig et al., 1993). The following year, the first two P2X ion channel receptors were cloned and characterized (Brake et al., 1994; Valera et al., 1994). These P2 receptors were later divided formally into P2X ionotopic and P2Y metabotropic receptor families (Abbracchio and Burnstock, 1994). This is put into perspective with receptors to other neurotransmitters in Table 3.
Figure 7.

Purinergic neuromuscular transmembrane depicting the synthesis, storage, release and inactivation of ATP. ATP, stored in vesicles in nerve varicosities, is released by exocytosis to act on post-junctional P2 purinoceptors on smooth muscle. ATP is broken down extracellularly by ATPases and 5′-nucleotidase to adenosine, which is taken up by varicosities to be resynthesized and restored in vesicles. Adenosine acts prejunctionally on P1 purinoceptors to modulate transmitter release. If adenosine is broken down further by adenosine deaminase to inosine, it is removed by the circulation. [Modified from Burnstock (1972), reproduced with permission from the American Society for Pharmacology and Experimental Therapeutics.]
Figure 8.
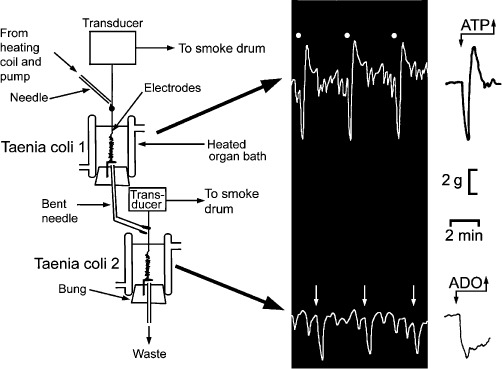
This figure shows Loewi-inspired experiments carried out in 1966. The upper guinea pig taenia coli innervated preparation was stimulated at 5 Hz for 40 s every 6 min at 50 V and 2 ms duration, in the presence of atropine and guanethidine to elicit typical non-adrenergic, non-cholinergic responses, fast relaxation followed by rebound contraction. The perfusate passed over the lower taenia coli preparation to produce slow relaxations, but not followed by rebound contractions. In later experiments, we showed that while the response of the taenia coli in the upper chamber was mimicked by ATP, the response in the lower chamber was mimicked by adenosine, the ATP released from the upper preparation being hydrolysed rapidly by ectonucleotidases to adenosine before reaching the lower preparation. [Experiments carried out by G. Burnstock and A. Smythe in 1966, but not published until 2010 (Burnstock et al., 2010) and updated with permission of the Scandinavian Physiological Society.]
Table 2.
Characterization of P1 receptors [Modified and updated from Burnstock (2003), with permission]
| P1 receptor | Agonists | Antagonists | Transduction mechanism | Patent utility |
|---|---|---|---|---|
| A1 | CCPA > R-PIA = S-ENBA | DPCPX | Gi/Go | Atrial fibrillation |
| N-0840 | cAMP ↓ | Cardioprotection | ||
| CVT-510 | MRS1754 | PLC ↑ | Diabetes, ischaemia | |
| GR79236 | WRC-0571 | |||
| A2A | HENECA > CGS21680 = CVT-3146 | KF17837 | GS | Inflammation |
| SCH 58261 | cAMP ↑ | Asthma | ||
| ZM241385 | IP3 ↑ | COPD | ||
| ATL-146e | KW 6002 | Parkinson's disease | ||
| A2B | Bay60-6583 | PBS603 | GS | Bowel and bladder abnormalities |
| MRE-2029-F20 | cAMP ↑ | |||
| PSB0788 | PLC ↑ | |||
| MRS 1706 | ||||
| PSB 1115 | ||||
| Alloxazine | ||||
| A3 | IB-MECA > 2-Cl-IB-MECA | MRS1220 | Gi/Go | Cardioprotection, cerebrovascular ischaemia, cancer, allergy |
| L-268605 | Gq/G11 | |||
| DBXRM | MRS 1191 and 1334 (h) | cAMP ↓ | ||
| VT160 | MRS 1523 (r) | IP3/DAG ↑ | ||
| HEMADO | VUF 8504 and 5574 | PLC ↑ | ||
| PSB10 |
CCPA, 2-chloro-N6-cyclopentyladenosine; COPD, chronic obstructive pulmonary disease; DBXRM, xanthine 7-riboside derivative; DPCPX, 8-cyclopentyl-1,3-dipropulxanthine; HEMADO, 2-(1-hexynyl)-N-methyladenosine; HENECA, 2-hexynyl-5′-N-ethylcarboxamidoadenosine; IB-MECA, N6-(3-iodobenzyl) adenosine-5′-N-methylcarboxamide; 2CI-1B-MECA, 2-chloro-N6-(3-iodobenzyl) adenosine-5′-N-methyluronamide; R-PIA, N6-phenylisopropyladenosine; S-ENBA, (2S)-N6-(2-endo-norbornyl) adenosine.
Table 3.
Comparison of fast ionotropic and slow metabotropic receptors for ACh, GABA, glutamate and 5-HT with those proposed for ATP [Updated and reproduced from Burnstock (1996) with permission from John Wiley and Sons]
| Receptors | ||
|---|---|---|
| Neurotransmitter | Fast ionotropic (ligand-gated ion channels) | Slow metabotropic (G protein) |
| ACh | NICOTINIC | MUSCARINIC |
| Muscle type | M1 → M5 | |
| Neuronal type | ||
| GABA | GABA A | GABA B |
| GLUTAMATE | AMPA | mGlu1 |
| Kainate | ↓ | |
| NMDA | mGlu7 | |
| 5-HT | 5-HT3 | 5-HT1A-F 5-HT2A-C |
| 5-HT4 5-HT5A-B | ||
| 5-HT6 5-HT7 | ||
| ATP | P2X | P2Y |
| P2X1 → P2X7 | P2Y1, P2Y2, P2Y4, P2Y6, P2Y11, P2Y12, P2Y13, P2Y14 | |
Outstanding studies by scientists such as Alan North, Annemarie Surprenant, Alex Verkhratsky, George Dubyak, Bal Khakh, Gary Housley, Brian King, Terry Egan and Richard Evans extended knowledge of the molecular physiology of P2X receptor subtypes (Table 4) and for P2Y receptor subtypes (Table 5), where Maria Abbracchio, Jean-Marie Boeynaems, Ken Harden and Eric Barnard were the leading contributors. The molecular structures of P1, P2X and P2Y receptors are shown in Figure 9 (Ralevic and Burnstock, 1998). A major conceptual advance was made recently when the crystal structure of P2X4 receptors was presented (Kawate et al., 2009).
Table 4.
Characterization of P2X receptors [Modified and updated from Burnstock (2003), with permission]
| Receptor | Main distribution | Agonists | Antagonists |
|---|---|---|---|
| P2X1 | Smooth muscle, platelets, cerebellum, dorsal horn spinal neurones | L-βγ-meATP ≥α,β-meATP = ATP = 2-MeSATP | TNP-ATP, IP5I, NF023, NF449 |
| PAPET-ATP (rapid desensitization) | RO1, RO 0437626, NF279, MRS2159 | ||
| P2X2 | Smooth muscle, CNS, retina, chromaffin cells, autonomic and sensory ganglia | ATP ≥ ATPγS ≥ 2-MeSATP >> α,β-meATP, β,γ-CF2ATP (pH + zinc sensitive) | Suramin, RB2, NF770, isoPPADS, PSB-1011, NF778, aminoglycoside |
| P2X3 | Sensory neurones, NTS, some sympathetic neurones | 2-MeSATP ≥ ATP ≥α,β-meATP = Ap4A, PAPET-ATP (rapid desensitization) | TNP-ATP, isoPPADS |
| A317491, NF110, RO4, RN-1838, spinorphin, AF353 | |||
| P2X4 | CNS, testis, colon | ATP > α,β-meATP, CTP | TNP-ATP and BBG (weak), BBG, Paroxetine, 5-BDBD, CORM 2, phenolphthalein |
| Ivermectin (potentiates) | |||
| P2X5 | Proliferating cells in skin, gut, bladder, thymus, spinal cord | ATPγS | Suramin, PPADS, BBG |
| Ap4A | |||
| GTP | |||
| P2X6 | CNS, motor neurones in spinal cord | Does not function as homomultimer | – |
| P2X7 | Apoptotic cells in immune cells, pancreas, skin, etc. | BzATP > ATP ≥ 2-MeSATP > α,β-meATP | KN62, KN04, MRS2427, BBG, o-ATP, Decavanadate, A-804598, RN-6189, AZD-9056, AZ10606120, A740003, A-438079, GSK-1370319 |
5-BDBD, 5-(3-bromophenyl)-1,3-dihydro-2H-benzofuro[3,2-e]-1,4-diazepine-2-one; BBG, brilliant blue G; NTS, nucleus tractus solitarii; PPADS, pyridoxalphosphate-6-azophenyl-2′,5′-disulfonic acid; TNP-ATP, 2′(3′)-O-(2,4,6-trinitrophenyl) ATP.
Table 5.
Characteristics of P2Y receptors [Modified and updated from Burnstock (2003), with permission]
| P2Y Receptor | Main distribution | Agonists | Antagonists | Transduction mechanisms |
|---|---|---|---|---|
| P2Y1 | Epithelial and endothelial cells, platelets, immune cells, osteoclasts, brain | MRS2365 > 2-MeSADP = Ap5(γB) >> ADPβS > ATP > 2-MeSATP = ADP | MRS2500 > MRS2279 > MRS2179, PIT, A3P5P | Gq/G11; PLC-β activation |
| P2Y2 | Immune cells, epithelial and endothelial cells, kidney tubules, osteoblasts | 2-thio-UTP > UTP, MRS2698 ≥ ATP, INS 365 > INS 37217, UTPγS > Ap4A > MRS 2768, Up4-phenyl ester | AR-C126313 > Suramin > RB2, PSB-716, MRS2576 | Gq/G11 and possibly Gi/Go; PLC-β activation |
| P2Y4 | Endothelial cells, placenta, spleen, thymus | 2′-azido-dUTP > UTPγS, UTP ≥ ATP ≥ Ap4A Up4U | ATP (human) > Reactive Blue 2 > Suramin, MRS2577, PPADS | Gq/G11 and possibly Gi; PLC-β activation |
| P2Y6 | Airway and intestinal epithelial cells, placenta, T cells, thymus, microglia (activated) | MRS2693 > UDPβS, PBS0474 > INS48823, Up3U, 3-phenacyl-UDP >> UDP > UTP >> ATP, α,β-meUDP | MRS2578 > Reactive Blue 2, PPADS, MRS2567, MRS2575 (human) | Gq/G11; PLC-β activation |
| P2Y11 | Spleen, intestine, granulocytes | ATPγS > AR-C67085MX > BzATP ≥ ATP, NF546, NAD+, NAADP+ | NF157 > Suramin > RB2, 5′-AMPS, NF340, AMP-α-5, | Gq/G11 and GS; PLC-β activation |
| P2Y12 | Platelets, glial cells | 2-MeSADP ≥ ADP > ATP, ADP-β-S | AR-C69931MX > AZD6140, INS50589 > RB2 > 2-MeSAMP AR-C66096, CT50547,PSB-0413, Carba-nucleosides, MRS2395, AR-C67085 | GαI; inhibition of adenylate cyclase |
| P2Y13 | Spleen, brain, lymph nodes, bone marrow, erythrocytes | ADP = 2-MeSADP > 2-MeSATP, ATP | AR-C69931MX > AR-C67085 > MRS2211, 2-MeSAMP | Gi/Go |
| P2Y14 | Placenta, adipose tissue, stomach, intestine, discrete brain regions, mast cells | MRS2690 > UDP > UDP glucose ≥ UDP-galactose, UDP-glucosamine | Gq/G11 |
PIT, 2,2′-pyridylisatogen tosylate, UDP, uridine diphosphate; UTP, uridine 5′-triphosphate.
Figure 9.
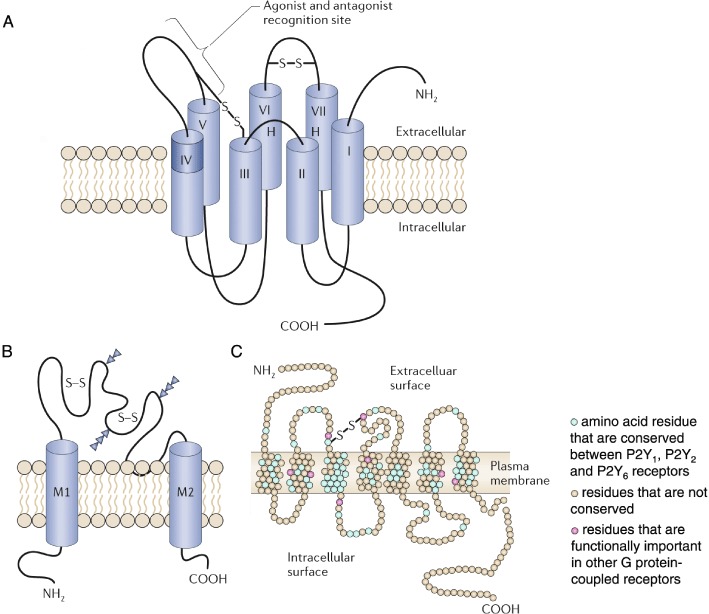
Membrane receptors for extracellular adenosine and ATP. (A) The P1 family of receptors for extracellular adenosine are G protein-coupled receptors (S–S; disulphide bond). (B) The P2X family of receptors are ligand-gated ion channels (S–S; disulphide bond; M1 and M2, transmembrane domains), and (C) the P2Y family are G protein-coupled receptors (S–S; disulphide bond). [(A) Modified from Ralevic and Burnstock (1998), reproduced with permission from the American Society for Pharmacology and Experimental Therapeutics; (B) from Brake et al. (1994), reproduced with permission from Nature; (C) modified from Barnard et al. (1994), reproduced with permission from Elsevier.]
While the initial focus was about purinergic signalling in excitable tissues, with employment of immunohistochemistry, it became clear that most non-neuronal cells in the body express multiple purinoceptor subtypes (Table 6), raising questions about the different roles of subtypes and their interactions.
Table 6.
Principal P1 and P2 receptors expressed by non-neuronal cells
| Smooth muscle | P2X1, P2X2, P2X4, P2X7, P2Y1, P2Y2 |
| Cardiac muscle | P2X1-6, P2Y2 (plus P2X7 and P2Y1 in isolated ventricle myocytes) |
| Skeletal muscle | P2X1-6, P2Y1, P2Y2, P2Y4, P2Y6 (transiently expressed during development) |
| Osteoblasts | P2X7, P2Y1, P2Y2 |
| Cartilage | P2X2, P2Y1, P2Y2, A2A, A2B |
| Keratinocytes | P2X2, P2X3, P2X5, P2X7, P2Y1, P2Y2, P2Y4, A2B |
| Fibroblasts | P2X7, P2Y1, P2Y2, A2A |
| Adipocytes | P2X1, P2Y1, P2Y2, P2Y4, A1 |
| Epithelial cells | P2X4, P2X5, P2X6, P2X7, P2Y1, P2Y2, P2Y6, P2Y11, A1, A2A, A3 |
| Hepatocytes | P2Y1, P2Y2, P2Y4, P2Y6, P2Y13, A2A, A3 |
| Glial cells | P2X1-7, P2Y1, P2Y2, P2Y4, P2Y6, P2Y11, P2Y12, P2Y13, A1, A2 |
| Sperm | P2X2, P2X7, P2Y2, A1 |
| Endothelial cells | P2X1, P2X4, P2Y1, P2Y2, P2Y4, P2Y6, A1, A2A |
| Erythrocytes | P2X2, P2X4, P2X7, P2Y1 |
| Platelets | P2X1, P2Y1, P2Y12, A2A |
| Immune cells (lymphocytes, neutrophils, macrophages, basophils, mast cells, eosinophils, osteoclasts, microglia, dendritic cells) | P2X1, P2X4, P2X7, P2Y1, P2Y2, A2A, A3 |
| Exocrine cells | P2X1, P2X4, P2X7, P2Y1, P2Y2, P2Y4, A1, A2A |
| Endocrine cells | P2X1-7, P2Y2, P2Y4, A1, A2A, A2B, A3 |
| Special senses | |
| Inner ear | P2X1, P2X2, P2X3, P2X7, P2Y2, P2Y4, A1 |
| Eye | P2X2, P2X7, P2Y2, A1, A2, A3 |
| Tongue | P2X2, P2X3, P2Y1, A1 |
| Olfactory organ | P2X2, P2X4, P2Y1, P2Y2, A2A, A3 |
Reproduced from Burnstock (2012c), with permission from Wiley.
There are papers now that show the molecular structure of ion channel receptors for ATP in primitive invertebrates such as Dictyostelium and Shistosoma, as well as green algae, which are remarkably similar to those expressing P2X receptors in mammals (Agboh et al., 2004; Fountain et al., 2007; 2008; Fountain and Burnstock, 2009). This suggests that ATP was one of the earliest extracellular messengers (see Burnstock and Verkhratsky, 2009). ATP signalling is also currently being explored in plants (see Demidchik et al., 2003; 2011; Kim et al., 2006; Clark and Roux, 2009).
Purinergic physiology
A major conceptual step was revealed when purinergic synaptic neurotransmission was reported between neurones in ganglia (Evans et al., 1992; Silinsky et al., 1992) and in the brain (Edwards et al., 1992). Another important advance was the recognition that as well as short-term purinergic signalling in neurotransmission, neuromodulation, secretion, chemoattraction, platelet aggregation and acute inflammation, purines and pyrimidines are also involved in long-term signalling of cell proliferation, differentiation and death during development and regeneration (see Abbracchio and Burnstock, 1998; Burnstock and Verkhratsky, 2010a). For example, in blood vessels, ATP, which is released from perivascular sympathetic nerves, excites smooth muscle via P2X receptors, while ATP released from endothelial cells during changes in blood flow (shear stress) and during hypoxia acts on endothelial P2Y and some P2X receptors to release nitric oxide. This results in vasodilatation, thus forming short-term dual control of vascular tone by purines (Figure 10). In addition, ATP and adenosine mediate long-term signalling during angiogenesis, restenosis following angioplasty and atherosclerosis (see Burnstock, 2002; 2008a; Erlinge and Burnstock, 2008).
Figure 10.

A schematic representation of the interactions of ATP released from perivascular nerves and from the endothelium (Endoth.). ATP is released from endothelial cells during hypoxia to act on endothelial P2Y receptors, leading to the production of endothelium-derived relaxing factor (EDRF) (NO) and subsequent vasodilation (–). In contrast, ATP released as a co-transmitter with noradrenaline (NA) from perivascular sympathetic nerves at the adventitia (Advent.)–muscle border produces vasoconstriction (+) via P2X receptors on the muscle cells. Adenosine (ADO), resulting from rapid breakdown of ATP by ectoenzymes, produces vasodilation by direct action on the muscle via P1 receptors and acts on the perivascular nerve terminal varicosities to inhibit transmitter release. [From Burnstock (1987), reproduced with permission from S. Karger AG, Basel.]
Another important concept is that there is growing recognition that purines interact synergistically with growth factors to promote growth of nerve fibres in the central nervous system (see Höpker et al., 1996; Guarnieri et al., 2004; Neary and Zimmermann, 2009) and in the differentiation of stem cells (see Burnstock and Ulrich, 2011).
For many years, it was assumed that the main source of ATP acting on purinoceptors was damaged or dying cells. However, it is now recognized that ATP is released, without causing any damage, from many cell types, including endothelial and urothelial cells, astrocytes, macrophages, osteoblasts and odontoblasts, in response to gentle mechanical disturbance, hypoxia and some agents (Bodin and Burnstock, 2001; Lazarowski et al., 2011; Lazarowski, 2012). This underlies the purinergic mechanosensory transduction that occurs in a variety of physiological events, including bone remodelling and visceral pain (Burnstock, 1999; 2007; Orriss et al., 2010). The mechanism of ATP transport from cells appears to be a combination of connexin and pannexin hemichannels and vesicular exocytosis (see Lazarowski, 2012). There is much known now about the ectoenzymes involved in the breakdown of released ATP into ADP, AMP, adenosine, inosine and hypoxanthine (see Zimmermann, 2006; Yegutkin, 2008). These enzymes include NTPDases, NPPs, alkaline phosphatases, 5′-nucleotidase and monoamine oxidase.
Purinergic pathophysiology
The autonomic nervous system shows marked plasticity of expression of co-transmitters and their receptors during development and ageing in the nerves that remain following trauma or surgery and in disease situations. For example, during embryological development of the amphibian Xenopus, we identified a P2Y8 G protein-coupled receptor that with in situ hybridization was shown to be involved in the development of the nervous system (Bogdanov et al., 1997). P2Y1 receptors were strongly expressed in the limb buds and mesonephros of the chick embryo at stage 20 (Figure 11A; Meyer et al., 1999). Mesenchyme cells isolated from the limb buds at that stage strongly proliferated in the presence of ATP (Figure 11B). In the rat brain, messenger RNA for P2X receptors revealed a sequential appearance during development: P2X3 receptors at E11; P2X2 and P2X5 receptors at E14; P2X4, P2X5 and P2X6 receptors at P1. P2X1 receptors were not expressed until later when neuromuscular junctions appeared (Cheung et al., 2005).
Figure 11.
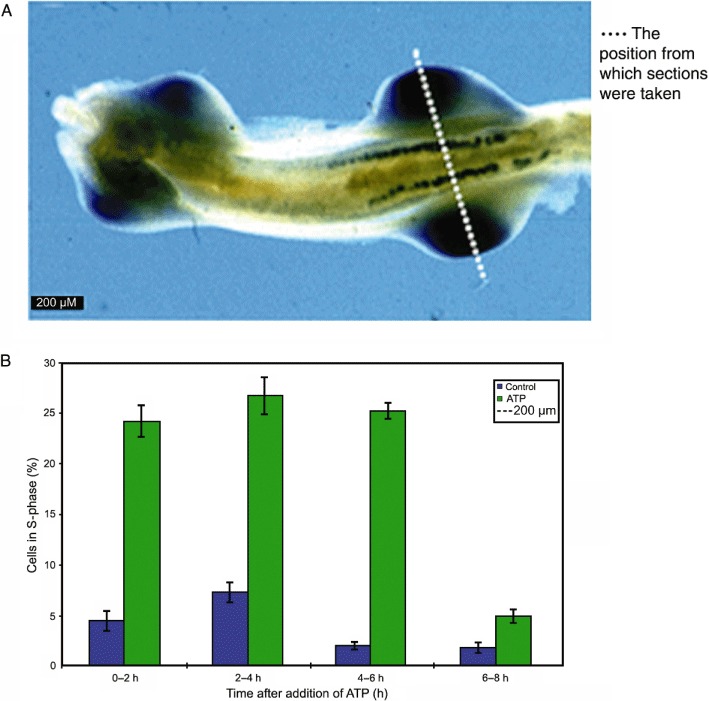
(A) Expression of cP2Y1 in the mesonephros. Ventral view of stage 20 embryo showing expression in mesonephros and limb buds. [Reproduced from Meyer et al. (1999), with permission from Wiley-Liss, Inc.] (B) Extracellular ATP is a potent mitogen for chick limb bud mesenchyme cells. Single dose of 100 µM ATP elicits a sustained increase in proliferation of bromo-2′-deoxyuridine. (Unpublished data, courtesy of Dr. M.P. Meyer.)
There appears to be an increase in the ATP component of co-transmission in pathological conditions, particularly during inflammation and stress. For example, while the purinergic component in parasympathetic nerves supplying the healthy bladder is minimal, in interstitial cystitis, outflow obstruction and neurogenic bladder, the purinergic component increases to up to 40% (see Burnstock, 2011). Also in spontaneously hypertensive rats, there are reports of a significantly greater co-transmitter role for ATP in sympathetic nerves supplying blood vessels (Erlinge and Burnstock, 2008).
Platelet aggregation is mediated by ADP acting via P2Y1 and P2Y12 receptors. A major clinical advance was made when clopidogrel, which is currently widely used against stroke and thrombosis, was found to block P2Y12 receptors (Gachet, 2006; Figure 12).
Figure 12.
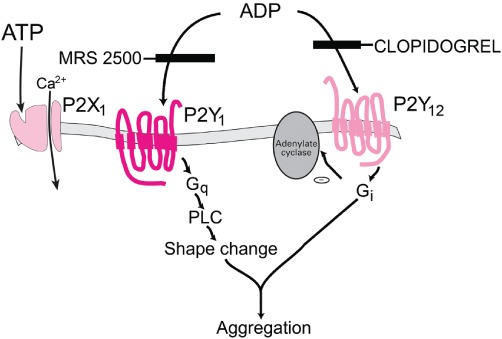
Functional role of three P2 receptor subtypes in ADP-induced platelet activation. [Modified from Kunapuli and Daniel (1998), with permission from Elsevier.]
A hypothesis proposing the involvement of purinergic signalling in the initiation of pain was presented in the Lancet (Burnstock, 1996). It was suggested that P2X3 receptors, expressed on nociceptive nerve endings, were stimulated by ATP released as a co-transmitter from sympathetic nerves during causalgia and reflex sympathetic dystrophy. It was also proposed that ATP was released from endothelial cells in the microvasculature supplying heart, skeletal muscle and cerebral vessels during angina, ischaemia and migraine, and from tumour cells during cancer pain. Later, purinergic mechanosensory transduction was identified (Burnstock, 1999) as well as its involvement in the initiation of visceral pain (Figure 13A, B; Burnstock, 2009b; 2012a). In a seminal paper, Kazu Inoue and colleagues showed that in neuropathic pain there was increased expression of P2X4 receptors on microglia. P2X4 receptors were involved, as neuropathic pain was reduced by antagonists to this receptor or after their removal (Tsuda et al., 2004). Subsequently, antagonists to P2X7 and P2Y receptors expressed on microglia were also shown to reduce neuropathic pain (Burnstock, 2009b).
Figure 13.
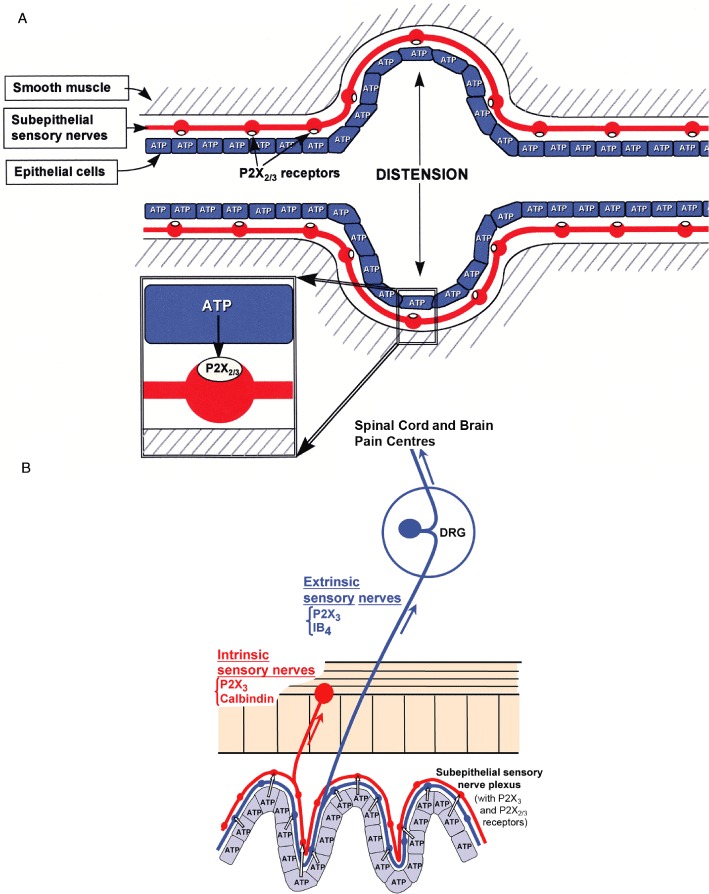
Purinergic mechanosensory transduction. (A) Schematic representation of hypothesis for purinergic mechanosensory transduction in tubes (e.g. ureter, vagina, salivary and bile ducts, gut) and sacs (e.g. urinary and gall bladders, and lung). It is proposed that distension leads to release of ATP from epithelium lining the tube or sac, which then acts on P2X3 and/or P2X2/3 receptors on subepithelial sensory nerves to convey sensory/nociceptive information to the CNS. [From Burnstock (1999), reproduced with permission from Blackwell Publishing.] (B) Schematic of a novel hypothesis about purinergic mechanosensory transduction in the gut. It is proposed that ATP released from mucosal epithelial cells during moderate distension acts preferentially on P2X3 and/or P2X2/3 receptors on low-threshold subepithelial intrinsic sensory nerve fibres (labelled with calbindin) to modulate peristaltic reflexes. ATP released during extreme (colic) distension also acts on P2X3 and/or P2X2/3 receptors on high-threshold extrinsic sensory nerve fibres [labelled with isolectin B4 (IB4) ] that send messages via the dorsal root ganglia (DRG) to pain centres in the central nervous system. [From Burnstock (1999), reproduced with permission from Wiley.]
Purinergic signalling is also involved in bone development, and regeneration and therapeutic strategies are being explored for osteoporosis (Orriss et al., 2010) and for kidney disease (Bailey et al., 2007; Taylor et al., 2009). There is growing interest in the role of purines and pyrimidines in normal behaviour including learning and memory, sleep and arousal, locomotion and feeding. There are also investigations of the roles of purinergic signalling in disorders of the brain, including trauma following accidents, surgery, stroke and ischaemia, neurodegenerative diseases such as Alzheimer's, Parkinson's and Huntington's, as well as multiple sclerosis, epilepsy and neuropsychiatric disorders, including depression anxiety and schizophrenia (see Burnstock, 2007; 2008b; Burnstock et al., 2011a, b). Finally, it was recognized early that ATP was effective against cancer (Rapaport, 1983). There are now many studies extending these findings, showing that P2Y1 and P2Y2 receptors mediate proliferation in most tumours, that P2X5 receptors mediate differentiation and therefore are anti-proliferative, while P2X7 receptors lead to apoptotic death of tumour cells (Figure 14; White and Burnstock, 2006; Shabbir and Burnstock, 2009; G. Burnstock and F. Di Virgilio, unpubl. data).
Figure 14.

(A) Left-hand panel: Effect of ATP on the growth of implanted DU145 tumour cells in vivo after 60 days initial growth; the lower mouse received ATP treatment versus no treatment in the upper mouse. Right-hand panel: Effect of ATP on the fractional growth of hormone refractory prostate cancer DU145 tumour cells in vivo after 60 days initial growth. [Reproduced from Shabbir et al. (2008), with permission from Blackwell Publishing.] (B) Different mechanisms by which P2 receptor subtypes might alter cancer cell function. P2Y1 and P2Y2 receptors might affect the rate of cell proliferation by modulating AC and altering the intracellular levels of cAMP, or by increasing the intracellular level of Ca2+ through the phospholipase C (PLC) pathway. P2X5 and P2Y11 receptor activation might switch the cell cycle from proliferation into a state of differentiation. The P2X7 receptor activates the apoptotic caspase enzyme system. Abbreviations: DAG, diacylglycerol; Ins(1,4,5)P3, inositol (1,4,5)-trisphosphate; PtdIns(4,5)P2, phosphatidylinositol (4,5)-bisphosphate. [Reproduced from White and Burnstock (2006), with permission from Elsevier.]
A recent hypothesis proposes that purinergic signalling is a major factor in the physiological mechanism responsible for the effects of acupuncture (see Burnstock, 2009a). It is suggested that the mechanical stimulation by twisting needles in the skin and tongue or heat or electrical currents leads to the release of ATP from keratinocytes. The ATP then initiates activity in sensory nerves in the skin via P2X receptors that relay through inter-neurones to the brain stem where they modulate the activity of motor neurones that control autonomic function. They also interrupt pain pathways, leading to the cortex.
Summary
Figure 15 shows the remarkable growth of papers published about purinergic signalling via ATP since 1972. Therapeutic approaches to pathological disorders include the development of selective P1 and P2 receptor subtype agonists and antagonists, as well as of inhibitors of extracellular ATP breakdown and of ATP transport enhancers and inhibitors. Medicinal chemists are starting to develop small molecule purinergic drugs that are orally bioavailable and stable in vivo (see Baqi et al., 2010; Gever et al., 2010; Burnstock, 2011; Burnstock and Kennedy, 2011).
Figure 15.
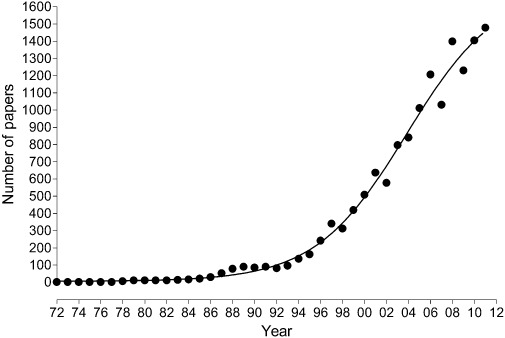
Graph showing the number of papers published on P2 purinergic signalling between 1972 and the end of 2011.
Supporting information
Additional Supporting Information may be found in the online version of this article:
Video S1 The birth and postnatal development of purinergic signalling. Professor Geoffrey Burnstock delivers the Gaddum Prize Lecture at the BPS Winter Meeting, London 2011.
Please note: Wiley-Blackwell are not responsible for the content or functionality of any supporting materials supplied by the authors. Any queries (other than missing material) should be directed to the corresponding author for the article.
References
- Abbracchio MP, Burnstock G. Purinoceptors: are there families of P2X and P2Y purinoceptors? Pharmacol Ther. 1994;64:445–475. doi: 10.1016/0163-7258(94)00048-4. [DOI] [PubMed] [Google Scholar]
- Abbracchio MP, Burnstock G. Purinergic signalling: pathophysiological roles. Jpn J Pharmacol. 1998;78:113–145. doi: 10.1254/jjp.78.113. [DOI] [PubMed] [Google Scholar]
- Abbracchio MP, Burnstock G, Verkhratsky A, Zimmermann H. Purinergic signalling in the nervous system: an overview. Trends Neurosci. 2009;32:19–29. doi: 10.1016/j.tins.2008.10.001. [DOI] [PubMed] [Google Scholar]
- Agboh KC, Webb TE, Evans RJ, Ennion SJ. Functional characterization of a P2X receptor from Schistosoma mansoni. J Biol Chem. 2004;279:41650–41657. doi: 10.1074/jbc.M408203200. [DOI] [PubMed] [Google Scholar]
- Bailey MA, Shirley DG, King BF, Burnstock G, Unwin RJ. Extracellular nucleotides and renal function. In: Alpern RJ, Herbert SCG, editors. The Kidney: Physiology and Pathophysiology. San Diego: Elsevier; 2007. pp. 425–443. [Google Scholar]
- Baqi Y, Lee SY, Iqbal J, Ripphausen P, Lehr A, Scheiff AB, et al. Development of potent and selective inhibitors of ecto-5′-nucleotidase based on an anthraquinone scaffold. J Med Chem. 2010;53:2076–2086. doi: 10.1021/jm901851t. [DOI] [PubMed] [Google Scholar]
- Barnard EA, Burnstock G, Webb TE. G protein-coupled receptors for ATP and other nucleotides: a new receptor family. Trends Pharmacol Sci. 1994;15:67–70. doi: 10.1016/0165-6147(94)90280-1. [DOI] [PubMed] [Google Scholar]
- Belai A, Burnstock G. Evidence for coexistence of ATP and nitric oxide in non-adrenergic, non-cholinergic (NANC) inhibitory neurones in the rat ileum, colon and anococcygeus muscle. Cell Tissue Res. 1994;278:197–200. doi: 10.1007/BF00305792. [DOI] [PubMed] [Google Scholar]
- Bodin P, Burnstock G. Purinergic signalling: ATP release. Neurochem Res. 2001;26:959–969. doi: 10.1023/a:1012388618693. [DOI] [PubMed] [Google Scholar]
- Bogdanov YD, Dale L, King BF, Whittock N, Burnstock G. Early expression of a novel nucleotide receptor in the neural plate of Xenopus embryos. J Biol Chem. 1997;272:12583–12590. doi: 10.1074/jbc.272.19.12583. [DOI] [PubMed] [Google Scholar]
- Brake AJ, Wagenbach MJ, Julius D. New structural motif for ligand-gated ion channels defined by an ionotropic ATP receptor. Nature. 1994;371:519–523. doi: 10.1038/371519a0. [DOI] [PubMed] [Google Scholar]
- Burnstock G. The effects of acetylcholine on membrane potential, spike frequency, conduction velocity and excitability in the taenia coli of the guinea-pig. J Physiol. 1958a;143:165–182. doi: 10.1113/jphysiol.1958.sp006051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burnstock G. The action of adrenaline on excitability and membrane potential in the taenia coli of the guinea-pig and the effect of DNP on this action and on the action of acetylcholine. J Physiol. 1958b;143:183–194. doi: 10.1113/jphysiol.1958.sp006052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burnstock G. Purinergic nerves. Pharmacol Rev. 1972;24:509–581. [PubMed] [Google Scholar]
- Burnstock G. Do some nerve cells release more than one transmitter? Neuroscience. 1976;1:239–248. doi: 10.1016/0306-4522(76)90054-3. [DOI] [PubMed] [Google Scholar]
- Burnstock G. A basis for distinguishing two types of purinergic receptor. In: Straub RW, Bolis L, editors. Cell Membrane Receptors for Drugs and Hormones: A Multidisciplinary Approach. New York: Raven Press; 1978. pp. 107–118. [Google Scholar]
- Burnstock G. The changing face of autonomic neurotransmission. (The First von Euler Lecture in Physiology) Acta Physiol Scand. 1986;126:67–91. doi: 10.1111/j.1748-1716.1986.tb07790.x. [DOI] [PubMed] [Google Scholar]
- Burnstock G. Local control of blood pressure by purines. Blood Vessels. 1987;24:156–160. doi: 10.1159/000158691. [DOI] [PubMed] [Google Scholar]
- Burnstock G. Noradrenaline and ATP as cotransmitters in sympathetic nerves. Neurochem Int. 1990;17:357–368. doi: 10.1016/0197-0186(90)90158-p. [DOI] [PubMed] [Google Scholar]
- Burnstock G. Introduction: changing face of autonomic and sensory nerves in the circulation. In: Edvinsson L, Uddman R, editors. Vascular Innervation and Receptor Mechanisms: New Perspectives. San Diego: Academic Press Inc, USA; 1993. pp. 1–22. [Google Scholar]
- Burnstock G. A unifying purinergic hypothesis for the initiation of pain. Lancet. 1996;347:1604–1605. doi: 10.1016/s0140-6736(96)91082-x. [DOI] [PubMed] [Google Scholar]
- Burnstock G. Release of vasoactive substances from endothelial cells by shear stress and purinergic mechanosensory transduction. J Anat. 1999;194:335–342. doi: 10.1046/j.1469-7580.1999.19430335.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burnstock G. Purinergic signalling in gut. In: Abbracchio MP, Williams M, editors. Handbook of Experimental Pharmacology, Volume 151/II. Purinergic and Pyrimidinergic Signalling II – Cardiovascular, Respiratory, Immune, Metabolic and Gastrointestinal Tract Function. Berlin: Springer-Verlag; 2001. pp. 141–238. [Google Scholar]
- Burnstock G. Purinergic signalling and vascular cell proliferation and death. Arterioscler Thromb Vasc Biol. 2002;22:364–373. doi: 10.1161/hq0302.105360. [DOI] [PubMed] [Google Scholar]
- Burnstock G. Introduction: ATP and its metabolites as potent extracellular agonists. In: Schwiebert EM, editor. Current Topics in Membranes, Vol. 54. Purinergic Receptors and Signalling. San Diego: Academic Press; 2003. pp. 1–27. [Google Scholar]
- Burnstock G. A moment of excitement. Living history series. The discovery of non-adrenergic, non-cholinergic neurotransmission. Physiol News. 2004;56:7–9. [Google Scholar]
- Burnstock G. Physiology and pathophysiology of purinergic neurotransmission. Physiol Rev. 2007;87:659–797. doi: 10.1152/physrev.00043.2006. [DOI] [PubMed] [Google Scholar]
- Burnstock G. Purinergic signalling in atherosclerosis and restenosis. 2008a. IAS website http://www.athero.org/commentaries/comm770.pdf.
- Burnstock G. Purinergic signalling and disorders of the central nervous system. Nat Rev Drug Discov. 2008b;7:575–590. doi: 10.1038/nrd2605. [DOI] [PubMed] [Google Scholar]
- Burnstock G. Acupuncture: a novel hypothesis for the involvement of purinergic signalling. Med Hypotheses. 2009a;73:470–472. doi: 10.1016/j.mehy.2009.05.031. [DOI] [PubMed] [Google Scholar]
- Burnstock G. Purinergic mechanosensory transduction and visceral pain. Mol Pain. 2009b;5:69. doi: 10.1186/1744-8069-5-69. doi: 10.1186/1744-8069-5-69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burnstock G. Therapeutic potential of purinergic signalling for diseases of the urinary tract. BJU Int. 2011;107:192–204. doi: 10.1111/j.1464-410X.2010.09926.x. [DOI] [PubMed] [Google Scholar]
- Burnstock G. Targeting the visceral purinergic system for pain control. Curr Opin Pharmacol. 2012a;12:80–86. doi: 10.1016/j.coph.2011.10.008. [DOI] [PubMed] [Google Scholar]
- Burnstock G. Cotransmission. In: Robertson D, Biaggioni I, Burnstock G, Low P, Paton JF, editors. Primer of the Autonomic Nervous System. 3rd edn. Oxford: Academic Press; 2012b. pp. 27–34. [Google Scholar]
- Burnstock G. Purinergic Signalling: its unpopular beginning, its acceptance and its exciting future. Bio Essays. 2012c;34:218–225. doi: 10.1002/bies.201100130. [DOI] [PubMed] [Google Scholar]
- Burnstock G, Costa M. Adrenergic Neurones: Their Organisation, Function and Development in the Peripheral Nervous System. London: Chapman and Hall; 1975. pp. 1–225. [Google Scholar]
- Burnstock G, Holman ME. Autonomic nerve-smooth muscle transmission. Nature. 1960;187:951–952. doi: 10.1038/187951a0. [DOI] [PubMed] [Google Scholar]
- Burnstock G, Holman ME. The transmission of excitation from autonomic nerve to smooth muscle. J Physiol. 1961;155:115–133. doi: 10.1113/jphysiol.1961.sp006617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burnstock G, Kennedy C. Is there a basis for distinguishing two types of P2-purinoceptor? Gen Pharmacol. 1985;16:433–440. doi: 10.1016/0306-3623(85)90001-1. [DOI] [PubMed] [Google Scholar]
- Burnstock G, Kennedy C. P2X receptors in health and disease. In: Jacobson KA, Linden J, editors. Purine and Pyrimidine Receptor Pharmacology. Burlington: Academic Press; 2011. pp. 333–372. [DOI] [PubMed] [Google Scholar]
- Burnstock G, Straub RW. A method for studying the effects of ions and drugs on the resting and action potentials in smooth muscle with external electrodes. J Physiol. 1958;140:156–167. doi: 10.1113/jphysiol.1958.sp005924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burnstock G, Ulrich H. Purinergic signalling in embryonic and stem cell development. Cell Mol Life Sci. 2011;68:1369–1394. doi: 10.1007/s00018-010-0614-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burnstock G, Verkhratsky A. Evolutionary origins of the purinergic signalling system. Acta Physiol. 2009;195:415–447. doi: 10.1111/j.1748-1716.2009.01957.x. [DOI] [PubMed] [Google Scholar]
- Burnstock G, Verkhratsky A. Long-term (trophic) purinergic signalling: purinoceptors control cell proliferation, differentiation and death. Cell Death Dis. 2010a;1:e9. doi: 10.1038/cddis.2009.11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burnstock G, Verkhratsky A. Vas deferens – a model used to establish sympathetic cotransmission. Trends Pharmacol Sci. 2010b;31:131–139. doi: 10.1016/j.tips.2009.12.002. [DOI] [PubMed] [Google Scholar]
- Burnstock G, Verkhratsky A. Purinergic Signalling and the Nervous System. Heidelberg, Berlin: Springer-Verlag; 2012. pp. 1–715. [Google Scholar]
- Burnstock G, Wong H. Comparison of the effects of ultraviolet light and purinergic nerve stimulation on the guinea-pig taenia coli. Br J Pharmacol. 1978;62:293–302. doi: 10.1111/j.1476-5381.1978.tb08459.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burnstock G, Campbell G, Bennett M, Holman ME. Innervation of the guinea-pig taenia coli: are there intrinsic inhibitory nerves which are distinct from sympathetic nerves? Int J Neuropharmacol. 1964;3:163–166. doi: 10.1016/0028-3908(64)90003-6. [DOI] [PubMed] [Google Scholar]
- Burnstock G, Campbell G, Rand MJ. The inhibitory innervation of the taenia of the guinea-pig caecum. J Physiol. 1966;182:504–526. doi: 10.1113/jphysiol.1966.sp007834. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burnstock G, Campbell G, Satchell D, Smythe A. Evidence that adenosine triphosphate or a related nucleotide is the transmitter substance released by non-adrenergic inhibitory nerves in the gut. Br J Pharmacol. 1970;40:668–688. doi: 10.1111/j.1476-5381.1970.tb10646.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burnstock G, Cocks T, Crowe R, Kasakov L. Purinergic innervation of the guinea-pig urinary bladder. Br J Pharmacol. 1978;63:125–138. doi: 10.1111/j.1476-5381.1978.tb07782.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burnstock G, Fredholm BB, North RA, Verkhratsky A. The birth and postnatal development of purinergic signalling. Acta Physiol. 2010;199:93–147. doi: 10.1111/j.1748-1716.2010.02114.x. [DOI] [PubMed] [Google Scholar]
- Burnstock G, Krügel U, Abbracchio MP, Illes P. Purinergic signalling: from normal behaviour to pathological brain function. Prog Neurobiol. 2011b;95:229–274. doi: 10.1016/j.pneurobio.2011.08.006. [DOI] [PubMed] [Google Scholar]
- Burnstock G, Verkhratsky A, Fredholm B. Adenosine and ATP receptors in the brain. Curr Top Med Chem. 2011a;11:973–1011. doi: 10.2174/156802611795347627. [DOI] [PubMed] [Google Scholar]
- Cheung K-K, Chan WY, Burnstock G. Expression of P2X receptors during rat brain development and their inhibitory role on motor axon outgrowth in neural tube explant cultures. Neuroscience. 2005;133:937–945. doi: 10.1016/j.neuroscience.2005.03.032. [DOI] [PubMed] [Google Scholar]
- Clark G, Roux SJ. Extracellular nucleotides: ancient signaling molecules. Plant Sci. 2009;177:239–244. [Google Scholar]
- Daly JW. Adenosine receptors. Adv Cyclic Nucleotide Protein Phosphorylation Res. 1985;19:29–46. [PubMed] [Google Scholar]
- Demidchik V, Nichols C, Oliynyk M, Dark A, Glover BJ, Davies JM. Is ATP a signaling agent in plants? Plant Physiol. 2003;133:456–461. doi: 10.1104/pp.103.024091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Demidchik V, Shang Z, Shin R, Colaco R, Laohavisit A, Shabala S, et al. Receptor-like activity evoked by extracellular ADP in Arabidopsis root epidermal plasma membrane. Plant Physiol. 2011;156:1375–1385. doi: 10.1104/pp.111.174722. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drury AN, Szent-Györgyi A. The physiological activity of adenine compounds with special reference to their action upon the mammalian heart. J Physiol. 1929;68:213–237. doi: 10.1113/jphysiol.1929.sp002608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Edwards FA, Gibb AJ, Colquhoun D. ATP receptor-mediated synaptic currents in the central nervous system. Nature. 1992;359:144–147. doi: 10.1038/359144a0. [DOI] [PubMed] [Google Scholar]
- Erlinge D, Burnstock G. P2 receptors in cardiovascular physiology and disease. Purinergic Signal. 2008;4:1–20. doi: 10.1007/s11302-007-9078-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Evans RJ, Derkach V, Surprenant A. ATP mediates fast synaptic transmission in mammalian neurons. Nature. 1992;357:503–505. doi: 10.1038/357503a0. [DOI] [PubMed] [Google Scholar]
- Fountain SJ, Burnstock G. An evolutionary history of P2X receptors. Purinergic Signal. 2009;5:269–272. doi: 10.1007/s11302-008-9127-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fountain SJ, Parkinson K, Young MT, Cao L, Thompson CR, North RA. An intracellular P2X receptor required for osmoregulation in Dictyostelium discoideum. Nature. 2007;448:200–203. doi: 10.1038/nature05926. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fountain SJ, Cao L, Young MT, North RA. Permeation properties of a P2X receptor in the green algae Ostreococcus tauri. J Biol Chem. 2008;283:15122–15126. doi: 10.1074/jbc.M801512200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fredholm BB, IJzerman AP, Jacobson KA, Klotz KN, Linden J. International Union of Pharmacology. XXV. Nomenclature and classification of adenosine receptors. Pharmacol Rev. 2001;53:527–552. [PMC free article] [PubMed] [Google Scholar]
- Fujii S, Sasaki H, Mikoshiba K, Kuroda Y, Yamazaki Y, Mostafa TA, et al. A chemical LTP induced by co-activation of metabotropic and N-methyl-D-aspartate glutamate receptors in hippocampal CA1 neurons. Brain Res. 2004;999:20–28. doi: 10.1016/j.brainres.2003.11.058. [DOI] [PubMed] [Google Scholar]
- Gachet C. Regulation of platelet functions by P2 receptors. Annu Rev Pharmacol Toxicol. 2006;46:277–300. doi: 10.1146/annurev.pharmtox.46.120604.141207. [DOI] [PubMed] [Google Scholar]
- Gever JR, Rothschild S, Henningsen R, Martin R, Hackos D, Panicker S, et al. AF-353, a novel, potent orally bioavailable P2X3/P2X2/3 receptor antagonist. Br J Pharmacol. 2010;160:1387–1398. doi: 10.1111/j.1476-5381.2010.00796.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guarnieri S, Fanò G, Rathbone MP, Mariggiò MA. Cooperation in signal transduction of extracellular guanosine 5′ triphosphate and nerve growth factor in neuronal differentiation of PC12 cells. Neuroscience. 2004;128:697–712. doi: 10.1016/j.neuroscience.2004.06.073. [DOI] [PubMed] [Google Scholar]
- Henning RH. Purinoceptors in neuromuscular transmission. Pharmacol Ther. 1997;74:115–128. doi: 10.1016/s0163-7258(97)00015-6. [DOI] [PubMed] [Google Scholar]
- Holton P. The liberation of adenosine triphosphate on antidromic stimulation of sensory nerves. J Physiol. 1959;145:494–504. doi: 10.1113/jphysiol.1959.sp006157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Höpker VH, Saffrey MJ, Burnstock G. Neurite outgrowth of striatal neurons in vitro: involvement of purines in the growth promoting effect of myenteric plexus explants. Int J Dev Neurosci. 1996;14:439–451. [PubMed] [Google Scholar]
- Hoyle CHV. Purinergic cotransmission: parasympathetic and enteric nerves. Semin Neurosci. 1996;8:207–215. [Google Scholar]
- Jo YH, Role LW. Cholinergic modulation of purinergic and GABAergic co-transmission at in vitro hypothalamic synapses. J Neurophysiol. 2002;88:2501–2508. doi: 10.1152/jn.00352.2002. [DOI] [PubMed] [Google Scholar]
- Kasakov L, Burnstock G. The use of the slowly degradable analog, α,β-methylene ATP, to produce desensitisation of the P2-purinoceptor: effect on non-adrenergic, non-cholinergic responses of the guinea-pig urinary bladder. Eur J Pharmacol. 1983;86:291–294. doi: 10.1016/0014-2999(82)90330-2. [DOI] [PubMed] [Google Scholar]
- Kawate T, Michel JC, Birdsong WT, Gouaux E. Crystal structure of the ATP-gated P2X4 ion channel in the closed state. Nature. 2009;460:592–598. doi: 10.1038/nature08198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim SY, Sivaguru M, Stacey G. Extracellular ATP in plants. Visualization, localization, and analysis of physiological significance in growth and signaling. Plant Physiol. 2006;142:984–992. doi: 10.1104/pp.106.085670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krügel U, Schraft T, Kittner H, Kiess W, Illes P. Basal and feeding-evoked dopamine release in the rat nucleus accumbens is depressed by leptin. Eur J Pharmacol. 2003;482:185–187. doi: 10.1016/j.ejphar.2003.09.047. [DOI] [PubMed] [Google Scholar]
- Kunapuli SP, Daniel JL. P2 receptor subtypes in the cardiovascular system. Biochem J. 1998;336:513–523. doi: 10.1042/bj3360513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lazarowski ER. Vesicular and conductive mechanisms of nucleotide release. Purinergic Signal. 2012;8:359–373. doi: 10.1007/s11302-012-9304-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lazarowski ER, Sesma JI, Seminario-Vidal L, Kreda SM. Molecular mechanisms of purine and pyrimidine nucleotide release. Adv Pharmacol. 2011;61:221–261. doi: 10.1016/B978-0-12-385526-8.00008-4. [DOI] [PubMed] [Google Scholar]
- Lustig KD, Shiau AK, Brake AJ, Julius D. Expression cloning of an ATP receptor from mouse neuroblastoma cells. Proc Natl Acad Sci U S A. 1993;90:5113–5117. doi: 10.1073/pnas.90.11.5113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meyer MP, Clarke JDW, Patel K, Townsend-Nicholson A, Burnstock G. Selective expression of purinoceptor cP2Y1 suggests a role for nucleotide signalling in development of the chick embryo. Dev Dyn. 1999;214:152–158. doi: 10.1002/(SICI)1097-0177(199902)214:2<152::AID-AJA5>3.0.CO;2-L. [DOI] [PubMed] [Google Scholar]
- Mori M, Heuss C, Gahwiler BH, Gerber U. Fast synaptic transmission mediated by P2X receptors in CA3 pyramidal cells of rat hippocampal slice cultures. J Physiol. 2001;535:115–123. doi: 10.1111/j.1469-7793.2001.t01-1-00115.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neary JT, Zimmermann H. Trophic functions of nucleotides in the central nervous system. Trends Neurosci. 2009;32:189–198. doi: 10.1016/j.tins.2009.01.002. [DOI] [PubMed] [Google Scholar]
- Orriss IR, Burnstock G, Arnett TR. Purinergic signalling and bone remodelling. Curr Opin Pharmacol. 2010;10:322–330. doi: 10.1016/j.coph.2010.01.003. [DOI] [PubMed] [Google Scholar]
- Poelchen W, Sieler D, Wirkner K, Illes P. Co-transmitter function of ATP in central catecholaminergic neurons of the rat. Neuroscience. 2001;102:593–602. doi: 10.1016/s0306-4522(00)00529-7. [DOI] [PubMed] [Google Scholar]
- Ralevic V, Burnstock G. Receptors for purines and pyrimidines. Pharmacol Rev. 1998;50:413–492. [PubMed] [Google Scholar]
- Rapaport E. Treatment of human tumor cells with ADP or ATP yields arrest of growth in the S phase of the cell cycle. J Cell Physiol. 1983;114:279–283. doi: 10.1002/jcp.1041140305. [DOI] [PubMed] [Google Scholar]
- Richardson PJ, Brown SJ. ATP release from affinity-purified rat cholinergic nerve terminals. J Neurochem. 1987;48:622–630. doi: 10.1111/j.1471-4159.1987.tb04138.x. [DOI] [PubMed] [Google Scholar]
- Shabbir M, Burnstock G. Purinergic receptor-mediated effects of adenosine 5′-triphosphate in urological malignant diseases. Int J Urol. 2009;16:143–150. doi: 10.1111/j.1442-2042.2008.02207.x. [DOI] [PubMed] [Google Scholar]
- Shabbir M, Thompson CS, Jarmulowicz M, Mikhailidis DP, Burnstock G. Effect of extracellular ATP on the growth of hormone refractory prostate cancer in vivo. BJU Int. 2008;102:108–112. doi: 10.1111/j.1464-410X.2008.07578.x. [DOI] [PubMed] [Google Scholar]
- Silinsky EM, Hubbard JI. Release of ATP from rat motor nerve terminals. Nature. 1973;243:404–405. doi: 10.1038/243404a0. [DOI] [PubMed] [Google Scholar]
- Silinsky EM, Gerzanich V, Vanner SM. ATP mediates excitatory synaptic transmission in mammalian neurones. Br J Pharmacol. 1992;106:762–763. doi: 10.1111/j.1476-5381.1992.tb14408.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sneddon P, Burnstock G. Inhibition of excitatory junction potentials in guinea-pig vas deferens by α,β-methylene-ATP: further evidence for ATP and noradrenaline as cotransmitters. Eur J Pharmacol. 1984a;100:85–90. doi: 10.1016/0014-2999(84)90318-2. [DOI] [PubMed] [Google Scholar]
- Sneddon P, Burnstock G. ATP as a co-transmitter in rat tail artery. Eur J Pharmacol. 1984b;106:149–152. doi: 10.1016/0014-2999(84)90688-5. [DOI] [PubMed] [Google Scholar]
- Sperlágh B, Sershen H, Lajtha A, Vizi ES. Co-release of endogenous ATP and [3H]noradrenaline from rat hypothalamic slices: origin and modulation by α2-adrenoceptors. Neuroscience. 1998;82:511–520. doi: 10.1016/s0306-4522(97)00306-0. [DOI] [PubMed] [Google Scholar]
- Su C, Bevan JA, Burnstock G. [3H]adenosine triphosphate: release during stimulation of enteric nerves. Science. 1971;173:337–339. doi: 10.1126/science.173.3994.336. [DOI] [PubMed] [Google Scholar]
- Taylor SRJ, Turner CM, Elliott JI, Hewitt R, Pickering M, Cook HT, et al. P2X7-deficiency ameliorates accelerated nephrotoxic nephritis in mice. J Am Soc Nephrol. 2009;20:1275–1281. doi: 10.1681/ASN.2008060559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsuda M, Mizokoshi A, Shigemoto-Mogami Y, Koizumi S, Inoue K. Activation of p38 mitogen-activated protein kinase in spinal hyperactive microglia contributes to pain hypersensitivity following peripheral nerve injury. Glia. 2004;45:89–95. doi: 10.1002/glia.10308. [DOI] [PubMed] [Google Scholar]
- Valera S, Hussy N, Evans RJ, Adani N, North RA, Surprenant A, et al. A new class of ligand-gated ion channel defined by P2X receptor for extra-cellular ATP. Nature. 1994;371:516–519. doi: 10.1038/371516a0. [DOI] [PubMed] [Google Scholar]
- Webb TE, Simon J, Krishek BJ, Bateson AN, Smart TG, King BF, et al. Cloning and functional expression of a brain G-protein-coupled ATP receptor. FEBS Lett. 1993;324:219–225. doi: 10.1016/0014-5793(93)81397-i. [DOI] [PubMed] [Google Scholar]
- Westfall DP, Stitzel RE, Rowe JN. The postjunctional effects and neural release of purine compounds in the guinea-pig vas deferens. Eur J Pharmacol. 1978;50:27–38. doi: 10.1016/0014-2999(78)90250-9. [DOI] [PubMed] [Google Scholar]
- White N, Burnstock G. P2 receptors and cancer. Trends Pharmacol Sci. 2006;27:211–217. doi: 10.1016/j.tips.2006.02.004. [DOI] [PubMed] [Google Scholar]
- Yegutkin GG. Nucleotide- and nucleoside-converting ectoenzymes: important modulators of purinergic signalling cascade. Biochim Biophys Acta. 2008;1783:673–694. doi: 10.1016/j.bbamcr.2008.01.024. [DOI] [PubMed] [Google Scholar]
- Zimmermann H. Nucleotide signaling in nervous system development. Pflugers Arch. 2006;452:573–588. doi: 10.1007/s00424-006-0067-4. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.