

Abstract
The hallmarks of carcinogenesis are aberrations in gene expression and protein function caused by both genetic and epigenetic modifications. Epigenetics refers to the changes in gene expression programming that alter the phenotype in the absence of a change in DNA sequence. Epigenetic modifications, which include amongst others DNA methylation, covalent modifications of histone tails and regulation by non-coding RNAs, play a significant role in normal development and genome stability. The changes are dynamic and serve as an adaptation mechanism to a wide variety of environmental and social factors including diet. A number of studies have provided evidence that some natural bioactive compounds found in food and herbs can modulate gene expression by targeting different elements of the epigenetic machinery. Nutrients that are components of one-carbon metabolism, such as folate, riboflavin, pyridoxine, cobalamin, choline, betaine and methionine, affect DNA methylation by regulating the levels of S-adenosyl-L-methionine, a methyl group donor, and S-adenosyl-L-homocysteine, which is an inhibitor of enzymes catalyzing the DNA methylation reaction. Other natural compounds target histone modifications and levels of non-coding RNAs such as vitamin D, which recruits histone acetylases, or resveratrol, which activates the deacetylase sirtuin and regulates oncogenic and tumour suppressor micro-RNAs. As epigenetic abnormalities have been shown to be both causative and contributing factors in different health conditions including cancer, natural compounds that are direct or indirect regulators of the epigenome constitute an excellent approach in cancer prevention and potentially in anti-cancer therapy.
Keywords: natural compounds, epigenetics, anti-cancer action, chemoprevention
Introduction
Cancer initiation and progression are driven by the concurrent changes in the expression of multiple genes that occur via genetic and epigenetic alterations, leading to either activation of oncogenes and prometastatic genes, or silencing of tumour suppressor genes and to genome rearrangements and instability (Baylin et al., 2001; Widschwendter and Jones, 2002; Szyf, 2005; Stefanska et al., 2011a). Epigenetic modifications have attracted a significant amount of attention in the prevention and treatment of different illnesses, with cancer at the forefront, mainly due to their reversibility. Epigenetics refers to layers of information in addition to genetics and comprises several components such as DNA methylation, covalent histone modifications, particularly histone acetylation and methylation, and non-coding RNA-related mechanisms (Razin and Riggs, 1980; Strahl and Allis, 2000; Jenuwein and Allis, 2001; Bergmann and Lane, 2003). Recent findings have reported additional DNA modifications taking place on the methyl group such as hydroxy-methylcytosine and formyl- and carboxy-methylcytosine (Ito et al., 2010), whose biological role remains to be determined. Although catalyzed by different enzymes and controlled by different protein complexes, all the elements of the epigenome influence each other at the level of the chromatin structure (Razin and Riggs, 1980; Strahl and Allis, 2000; Bergmann and Lane, 2003). DNA hypomethylation, histone acetylation and histone H3K4 methylation have been associated with active chromatin, whereas DNA hypermethylation, histone deacetylation, and histone H3K9 and K27 di- or trimethylation have been found in inactive chromatin regions (Strahl and Allis, 2000). Studies have shown that methylation of DNA results in the recruitment of histone deacetylases (HDACs), which changes chromatin configuration (Nan et al., 1997; Eden et al., 1998). However, alterations in histone marks can also trigger changes in DNA methylation patterns. For example, histone H3K27 methylation and EZH2 histone methyltranferase were required for methylation of EZH2 target genes (Vire et al., 2006), whereas histone acetylation was followed by DNA demethylation (Selker, 1998; Cervoni and Szyf, 2001; Fahrner et al., 2002). Furthermore, changes in chromatin modifications in patients with mutations in the ATRX gene encoding SWI/SNF-like chromatin modifying protein resulted in diverse alterations in DNA methylation patterns (Gibbons et al., 2000). In addition, the third component of the epigenetic machinery, non-coding RNAs were shown to be required for de novo methylation of imprinted loci (Watanabe et al., 2011) and target mRNA of DNA methyltransferases (DNMTs) and HDACs (Rajewsky, 2006; Tuddenham et al., 2006; Zhou et al., 2010). The trilateral relationship between epigenetic modifications (D'Alessio and Szyf, 2006; Iorio et al., 2010) and their dynamic aspect, confirmed by a long list of studies (Weaver et al., 2004; Levenson et al., 2006; Miller and Sweatt, 2007; Feng et al., 2010), can have important implications for human diseases, including cancer where epigenetic factors are known to have a causal and/or contributing role (Baylin et al., 2001; Fahrner et al., 2002; Widschwendter and Jones, 2002; Szyf et al., 2004; Szyf, 2005; Kanwal and Gupta, 2011; Stefanska et al., 2011a). More interestingly, it has been shown that these epigenetic modifications are regulated by a wide range of bioactive food components (Table 1). Compounds from one-carbon metabolism, such as folate, cobalamin, riboflavin, pirydoxine or methionine, are involved in the regulation of the DNA methylation reaction (Scott and Weir, 1998; Slattery et al., 1999; Selhub, 2002). Their deficiency can lead to hepatocellular carcinoma (HCC) (Newberne and Rogers, 1986; Kanduc et al., 1988; Singh et al., 2003; Asada et al., 2006; Ghoshal et al., 2006; Calvisi et al., 2007) and other types of cancer (Kraunz et al., 2006; Bistulfi et al., 2010; Duthie, 2011). Constituents of tea and soybean such as epigallocatechin and genistein, respectively, reverse hypermethylation and the silencing of tumour suppressor genes and inhibit cancer growth (Fang et al., 2005; Lu et al., 2006). Anthocyanins from black raspberries have been shown to suppress DNMT1 and reactivate tumour suppressor genes by demethylating their promoters (Wang et al., 2011). A vegetarian diet has been found to be associated with promoter hypomethylation and the overexpression of a gene encoding the antioxidative enzyme mitochondrial superoxide dismutase, when compared with aged-matched omnivores (Thaler et al., 2009). A lower incidence of sporadic colorectal cancer in the population of southern Italy in comparison with the rest of the Western world has been linked to the presence of Annurca apple in the diet (Fini et al., 2011). Annurca apple contains polyphenols and is especially rich in chlorogenic acid, caffeic acid, catechin, epicatechin, rutin and phloridizin. Treatment of RKO, SW48 and SW480 colorectal cancer cell lines with an extract of Annurca apple polyphenols was shown to reduce DNA methylation in the promoters of hMLH1, p14 and p16 tumour suppressor genes and restore their expression with a concomitant decrease in DNMT1 and DNMT3B protein levels (Fini et al., 2007). Using natural compounds to modulate the epigenome opens an emerging field of nutritional epigenetics and offers a new approach to cancer prevention and treatment. In this review, we summarize recent data suggesting that many bioactive dietary compounds exert anti-cancer effects through epigenetic mechanisms. Furthermore, we highlight the close relationship between all the components of the epigenome and its importance in the evaluation of the effects of a tested agent. We finally discuss the difficulty in assessing the efficacy of natural compounds due to poor bioavailability and their dose-dependent effects.
Table 1.
Source, pharmacological effects and epigenetic mechanisms of action of promising natural compounds
| Natural compound | Natural sources | Pharmacological effects | Epigenetic mechanisms of action |
|---|---|---|---|
| Folate, cobalamin, riboflavin, pyridoxine, methionine | Folate and riboflavin: spinach, asparagus, beans, peas, lentils, sunflower seeds, almonds | Anti-cancer, anti-proliferative, chemoprevention of malignant transformation | Regulation of one-carbon metabolism, SAM/SAH ratio, DNMT and MBD expression; regulation of miRNAs (tumour suppressor miR-122, miR-34a, miR-127, and oncogenic miR-21, miR-222) |
| Cobalamin: fish, shellfish, poultry, milk, eggs | |||
| Pyridoxine and methionine: grains, nuts, dragon fruit, sesame seeds | |||
| Retinoic acid | Vietnamese gac, crude palm oil, yellow and orange fruits (mango, papaya), orange root vegetables (carrots), spinach, sweet potatoes | Anti-cancer, anti-proliferative, differentiating, pro-apoptotic | Regulation of DNMTs expression and enzyme activity by affecting p21, AP-1, PTEN and ERs; regulation of miRNAs targeting DNMTs; regulation of tumour suppressor miRNAs (miR-15, miR-16, let-7a, let-7c, miR-34a, miR-342) and oncogenic miRNAs (miR-10a); GNMT regulation; histone acetylation |
| Vitamin D3 | Sun exposure, fish, fish liver oils | Anti-cancer, anti-proliferative, differentiating, pro-apoptotic | Regulation of DNMTs expression and enzyme activity by affecting p21, AP-1, PTEN and ERs; regulation of histone acetylation; regulation of oncogenic miRNAs (miR-181a, miR-181b) |
| Resveratrol | Roots of hellebore, grapes, mulberries, apricots, pineapples, peanuts | Anti-cancer, antioxidant, anti-proliferative, anti-angiogenesis, anti-inflammatory, pro-apoptotic, cardioprotective | Regulation of DNMTs expression and enzyme activity by affecting p21, AP-1 and PTEN; activation of deacetylase SIRT1 and p300 HAT; down-regulation of UHRF1; regulation of miRNAs |
| Genistein and daidzein | Soybeans, lupin, kudzu, psoralea, fava beans, coffee | Anti-cancer, antioxidant, antihelminthic, anti-metastatic, cancer protective | Regulation of DNMTs expression and enzyme activity by affecting p21, AP-1 and PTEN; increase in HAT activity; regulation of miRNAs (tumour suppressor miR-1296, miR-16, and oncogenic miR-27a) |
| EGCG | Green tea | Anti-cancer, antioxidant, anti-proliferative, anti-angiogenesis, anti-inflammatory, pro-apoptotic, cancer protective | Regulation of SAM/SAH ratio by COMT-mediated reactions; direct inhibition of DNMTs by binding to catalytic domain of the enzyme; regulation of tumour suppressor miRNAs (miR-16) |
| Curcumin | Spice turmeric | Anti-cancer, antioxidant, protects against heart failure | Direct inhibition of DNMTs by binding to catalytic domain of the enzyme; inhibition of HDACs and p300 HAT; regulation of miRNAs (tumour suppressor miR-22, miR-15a, miR-16, and oncogenic miR-21, miR-199a) |
The role of epigenetic modifications in the regulation of gene transcription – relevance to carcinogenesis
DNA methylation
DNA methylation is a covalent modification of DNA that in mammalian cells takes place mainly at the fifth position of the cytosine pyrimidine ring located predominantly within CpG sequences (Hotchkiss, 1948; Wyatt, 1950; Gruenbaum et al., 1981). CG-rich regions, called CpG islands, are mostly unmethylated in normal cells and located in regulatory regions of housekeeping genes, tissue-specific genes and tumour suppressors (Hermann et al., 2004). CpG islands are also present in promoters of some oncogenes, where their methylated state is involved in the formation of an inactive chromatin structure leading to transcriptional silencing (Szyf et al., 2004). Recent data indicate that methylation can also occur in cytosines within dinucleotide sequences other than CpGs, although the role of non-CpG methylation remains to be elucidated (Lister et al., 2009).
DNA methylation in normal cells is implicated in oncogene repression, the control of expression of genes crucial for cell proliferation, differentiation and normal development as well as in parental imprinting, X chromosome inactivation, and preservation of chromosomal integrity by the silencing of transposons and repetitive elements (Hermann et al., 2004; Szyf et al., 2004). An inverse correlation between the extent of DNA methylation and gene expression was reported in many studies (Rauch et al., 2009; Stefanska et al., 2011a) and several mechanisms were proposed for DNA methylation-mediated transcriptional silencing (Stein et al., 1982). Firstly, some transcription factors, such as CREB (cAMP response element binding protein), E2F (elongation 2 factor), NF-κB and AP-2 (activator protein-2), are unable to recognize specific sequences when they are methylated or methylation occurs in their proximity (Comb and Goodman, 1990; Inamdar et al., 1991; Paluszczak and Baer-Dubowska, 2005). Secondly, the binding of transcription factors to regulatory elements within promoters and enhancers can be hindered by methyl-CpG-binding domain proteins (MBDs) that bind with high affinity to methylated DNA and cover recognition elements (Nan et al., 1997; Reik and Dean, 2001). The third mechanism is associated with the MBD-mediated recruitment of HDACs and histone methyltransferases that set up a compacted inactive chromatin state around the gene (Nan et al., 1997; Eden et al., 1998; Das and Singal, 2004). Numerous data show that the hallmark of cancer is global hypomethylation and hypermethylation of specific regions, mainly within promoters of tumour suppressor genes. The increase in promoter DNA methylation was reported as a common mechanism of tumour suppressor gene silencing observed in many types of cancer (Baylin et al., 2001; Szyf et al., 2004; Hatziapostolou and Iliopoulos, 2011), and the reversal of the aberrant DNA hypermethylation reactivated gene transcription (Szyf, 2005; Stefanska et al., 2010; 2011b). Recent whole-genome approaches demonstrate that loss of DNA methylation in gene promoters can have a detrimental effect in cancer as hypomethylation was linked to the activation of genes implicated in metastasis, invasion and other biological functions typical for cellular transformation (Stefanska et al., 2011a).
DNA methylation is carried out by DNMTs, which catalyze the transfer of a methyl group from the ubiquitous methyl donor, S-adenosyl-L-methionine (SAM), to the fifth position of a cytosine pyrimidine ring (Gruenbaum et al., 1981). The main enzyme responsible for the maintenance of DNA methylation patterns is DNMT1 (Chen et al., 2007a), whereas de novo methylation activity is attributed to DNMT3A and DNMT3B (Okano et al., 1999; Robertson et al., 1999). Both DNMT3s play a role in genomic imprinting and silencing of transposons and repetitive elements during embryonic development (Okano et al., 1998; 1999) and their substrate recognition is regulated by another member of DNMTs, DNMT3L (Bourc'his et al., 2001; Bourc'his and Bestor, 2004).
DNA methylation has been considered as a non-reversible reaction for decades. It was assumed that demethylation can only occur in a passive way by blocking the activity of DNMTs in dividing cells. However, evidence has been obtained showing that active demethylation does take place, such as demethylation of paternal genome, during embryogenesis or demethylation in post-mitotic neurons (Weaver et al., 2004; Mastronardi et al., 2007; Feng et al., 2010). Szyf et al. (Bhattacharya et al., 1999; Ramchandani et al., 1999; Detich et al., 2002) showed that one of the MBDs, MBD2, has demethylase activity and demethylates both fully methylated and hemimethylated DNA in vitro. One of the mechanisms of the active enzymatic removal of a methyl group from DNA can involve oxidation of 5-methylcytosine to 5-hydroxymethylcytosine, followed by the release of a methyl group in formaldehyde (Hamm et al., 2008). This concept becomes more probable in the light of recent findings demonstrating the presence of 5-hydroxymethylcytosine in mammalian DNA (Kriaucionis and Heintz, 2009; Ndlovu et al., 2011). Although the biological function of this modification still remains poorly understood, recent reports providing insights into 5-hydroxymethylcytosine genomic distribution suggest it is potentially highly relevant in gene regulation (Ndlovu et al., 2011). The ten-eleven translocation (TET) family of enzymes has been demonstrated to catalyze the oxidation of 5-methylcytosine to 5-hydroxymethylcytosine. In addition, TET1 was required for the maintainance of demethylation of nanog in embryonic stem cells (Ito et al., 2010). This raises the possibility that this reaction might be an intermediate form of active DNA demethylation (Ndlovu et al., 2011). An alternative mechanism suggested for DNA demethylation is based on the DNA repair system, where 5-methylcytosine is replaced with non-methylated cytosine (Razin et al., 1986; Barreto et al., 2007; Rai et al., 2008). Deamination of 5-methylcytosine to thymine carried out by the activation-induced (cytosine) deaminase (Rai et al., 2008) creates a G : T mismatch that is recognized by a specific thymine glycosylase, Mbd4, and the repair is mediated by the growth arrest and DNA-damage inducible (Gadd45) protein (Barreto et al., 2007).
Histone modifications and non-coding RNAs
Chromatin consists of DNA, histones and non-histone proteins, and plays a role in compacting DNA, preventing DNA damage, controlling gene expression and DNA replication (Kouzarides, 2007; Cedar and Bergman, 2009). DNA is wrapped around a histone octamer that comprises two subunits of each of the core histones, H2A, H2B, H3 and H4 (Kouzarides, 2007; Kanwal and Gupta, 2011), containing amino acid residues with an amino group (Kouzarides, 2007; Bannister and Kouzarides, 2011; Fullgrabe et al., 2011). The positively charged amino group interacts with the negatively charged phosphate groups of DNA, which results in the compaction of chromatin and the formation of transcriptionally inactive heterochromatin. It is well established that post-translational modifications of the histone tails regulate gene expression by determining chromatin structure; these modifications include methylation, acetylation, ubiquitination, phosphorylation, sumoylation, ADP ribosylation, biotinylation and proline isomerization (Jenuwein and Allis, 2001; Kouzarides, 2007; Bannister and Kouzarides, 2011). These modifications are dynamic and controlled by a variety of enzymes that add and remove specific groups. The most studied are histone acetylation mediated by histone acetyltransferases (HATs) and HDACs, and histone methylation controlled by methyltransferases (HMTs) and demethylases (HDMs) (Kouzarides, 2007; Yang and Seto, 2007; Bannister and Kouzarides, 2011). Actively transcribed genes are enriched with active histone marks such as acetylation of H3K9, H3K14 and H3K4, trimethylation of H3K4, dimethylation of H3K79 and monomethylation of H3K4, H3K9, H3K27, H3K79 and H2BK5, whereas regions transcriptionally inactive are characterized by dimethylation of H3K9 and H3K27, and trimethylation of H3K9, H3K27 and H2BK5 (Barski et al., 2007; Koch et al., 2007; Li et al., 2007; Steger et al., 2008; Rosenfeld et al., 2009; Karlic et al., 2010b). Recent studies with genome-wide approaches revealed that histone modifications have different functional outcomes on gene transcription and chromatin structure, depending on their positional context as well as the interplay among various histone modifications (Li et al., 2007; Batta et al., 2011).
Although still controversial, non-coding RNAs have been included as a component of the epigenome (Choudhuri, 2011). Non-coding RNAs are encoded by a major part of our genome and regulate gene expression by controlling translation of mRNA into proteins. Without changing the DNA sequence, non-coding RNAs and more specifically micro-RNAs (miRNAs) are involved in heritable changes in gene expression. More interestingly, they are implicated in reciprocal interconnections between all the components of the epigenome. Aberrant miRNA regulation, especially that associated with cancer, can be influenced by DNA methylation and histone covalent modifications (Lu et al., 2005; Iorio et al., 2010). On the other hand, miRNA affects the epigenome by targeting enzymes of the epigenetic machinery (Iorio et al., 2010; Zhou et al., 2010). DNMT1 and DNMT3B are targets of miR-148a in cholangiocarcinoma and cervical cancer, respectively. HDAC1 is targeted by miR-449a and b in prostate cancer, and HDAC4 by miR-1 in HCC. miRNA genes are transcribed in the nucleus and the miRNA transcript is processed in the cytoplasm where it is cut by the endoribonuclease Dicer to approximately 22 nucleotide fragments. This short, double-stranded RNA is part of the RNA-induced silencing complex where one strand of miRNA is eliminated and the other strand is orientated by Argonaute protein, Ago2, for interaction with a target mRNA. Complementary binding to mRNA is followed by its cleavage, degradation and inhibition of translation (Fire et al., 1998; Zhou et al., 2010; Choudhuri, 2011).
Aberrations in both histone modifications and miRNA regulation have been shown to be associated with several diseases, including cancer (Kanwal and Gupta, 2010; Lovat et al., 2011). Deregulation of miRNAs is linked to tumour initiation and progression in many human cancers (Lovat et al., 2011).
The impact of natural compounds on DNA methylation
Accumulating evidence shows that natural compounds can alter DNA methylation patterns by directly interacting with enzymes responsible for epigenetic marks or indirect regulation of genes that encode proteins implicated in the epigenetic machinery.
Folate and other nutrients of one-carbon metabolism
A group of nutrients involved in one-carbon metabolism such as folate, cobalamin, riboflavin, pirydoxine and methionine have a direct effect on the level of a methyl donor, SAM (Figure 1). Folate is a water-soluble form of vitamin B9, which is important for DNA synthesis, repair and methylation. After dietary intake, folate is converted to tetrahydrofolate, which is involved in the remethylation of homocysteine to methionine (Scott and Weir, 1998; Selhub, 2002). Methionine is a precursor of SAM, which is the primary methyl group donor for most methylation reactions (Waterland, 2006). After transferring a methyl group, SAM is converted to S-adenosyl-L-homocysteine (SAH), an inhibitor of methylation reactions (Figure 1). Numerous studies have shown a causal role of a folate deficiency in cancer development (Duthie et al., 2004), including amongst others colon (Kim, 2004; Duthie, 2011), prostate (Bistulfi et al., 2010) and liver cancer (James et al., 2003), as well as head and neck squamous cell carcinoma (Kraunz et al., 2006). Malignant transformation was accompanied by SAM depletion, global DNA hypomethylation, gene-specific hypomethylation and activation of oncogenes, and paradoxically in hypermethylation and silencing of tumour suppressor genes (James et al., 2003; Davis and Uthus, 2004; Duthie et al., 2004). Folate deficiency has been shown to alter components of the DNA methylation machinery with the increase in DNMT1, DNMT3B, MBD2 and MBD4 mRNA and protein levels during the early stages of hepatocarcinogenesis (Ghoshal et al., 2006). The increase in DNMTs may explain the hypermethylation observed, whereas the stimulation of MBD2 and MBD4 may be responsible for hypomethylation of oncogenes and prometastatic genes, as both proteins have been implicated in active DNA demethylation (Bhattacharya et al., 1999; Ramchandani et al., 1999; Detich et al., 2002; Rai et al., 2008; Popp et al., 2010). Hence, folic acid supplementation may have a chemopreventive effect; this has been demonstrated in a few cancer models, including hepatocarcinogenesis. In rats fed a folic acid-enriched diet followed by the chemical initiation of liver cancer, cell growth and the number of preneoplastic lesions as well as c-myc oncogene expression were diminished compared with a control group (Chagas et al., 2011). Although no changes in global DNA methylation were detected in this study, the SAM/SAH ratio significantly increased, which has the potential to induce gene-specific alterations. Furthermore, folic acid supplementation has been found to reverse cervical dysplasia in patients using oral contraceptives and prevent cervical cancer (Butterworth et al., 1982; Whitehead et al., 1989). Recent prospective and epidemiological studies of 718 Chinese breast cancer cases, Shanghai Women's Health Study (1997–2008), indicated that high folate intake reduces breast cancer risk in premenopausal women and that low intakes of methionine, pyridoxine, cobalamin, niacin and riboflavin are associated with breast carcinogenesis (Shrubsole et al., 2011). A protective role of folate in breast cancer, particularly in oestrogen receptor (ER)-negative cancer, was also shown in another prospective study (Maruti et al., 2009).
Figure 1.
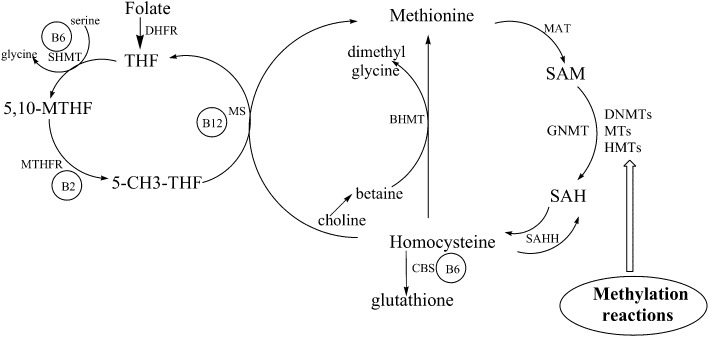
One-carbon metabolism. This network of biochemical reactions is essential for synthesis of S-adenosyl-L-methionine (SAM) that is the unique methyl donor for various methylation reactions including DNA and histone methylation. The crucial co-enzymes and intermediary compounds involved in the one-carbon metabolism comprise bioactive food components such as folate, pyridoxine (vitamin B6), riboflavin (vitamin B2) and cobalamin (vitamin B12), as well as methionine and choline. Abbreviations: 5,10-MTHF, 5,10-methylenetetrahydrofolate; 5-CH3-THF, 5-methyltetrahydrofolate; BHMT, betaine homocysteine methyltransferase; CBS, cystathionine-β-synthase; DHFR, dihydrofolate reductase; DNMTs, DNA methyltransferases; GNMT, glycine N-methyltransferase; HMTs, histone methyltransferases; MAT, methionine adenosyltransferase; MS, methionine synthase; MTHFR, methylenetetrahydrofolate reductase; MTs, methyltransferases; SAH, S-adenosyl-L-homocysteine; SAHH, S-adenosyl-L-homocysteine hydrolase; SHMT, serine hydroxymethyltransferase; THF, tetrahydrofolate.
Other B vitamins implicated in one-carbon metabolism include riboflavin (B2), pyridoxine (B6) and cobalamin (B12) (Figure 1). Riboflavin is a component of FAD (flavin adenine dinucleotide) that acts as a cofactor for methylenetetrahydrofolate (MTHF) reductase catalyzing the conversion of 5,10-MTHF to 5-MTHF, a substrate for homocysteine remethylation to methionine (Slattery et al., 1999). As shown in Figure 1, blocking this enzyme activity will diminish methionine synthesis and DNA methylation reactions. It was reported that a reduced riboflavin intake can lead to changes in the methyl supply, followed by alterations in the DNA methylation state (Singh et al., 2003). Pyridoxine acts not only as a cofactor for serine hydroxymethyltransferase in the synthesis of 5,10-MTHF from THF (Selhub, 2002) but also participates in the trans-sulfuration reactions, where glutathione is formed from homocysteine (Figure 1) (Maruti et al., 2009). A low pyridoxine plasma concentration has been linked to an increased risk of colorectal cancer in a prospective nested case–control study of female participants (The Nurses' Health Study; Wei et al., 2005). As a consequence of pyridoxine depletion, the SAH level increases, resulting in DNA hypomethylation through inhibition of methyltranferases (Maruti et al., 2009). Cobalamin is another vitamin B that, by affecting one-carbon metabolism, is involved in the regulation of DNA methylation. As a cofactor for methionine synthase, cobalamin is important for the methylation of homocysteine to methionine (Figure 1) (Selhub, 2002). Indeed, in many studies, a diet deficient in major dietary sources of the methyl group, including folate and cobalamin, has been linked to the development of HCC (Newberne and Rogers, 1986; Kanduc et al., 1988; Asada et al., 2006; Calvisi et al., 2007). Promoter hypomethylation and increased expression of oncogenes were detected in livers of rats fed methyl-deficient diets (Zapisek et al., 1992). More interestingly, the effects of folate, pyridoxine and cobalamin supplementation on DNA methylation and cancer risk were dependent on MTHFR gene polymorphisms. Colon cancer risk was reduced by 30–40% in individuals with variant TT genotype of the MTHFR gene who consumed adequate amounts of the nutrients compared to those with CC genotype whose diet was low in nutrients (Slattery et al., 1999; Davis and Uthus, 2004).
Retinoic acid and vitamin D3
Contrary to nutrients and vitamins such as folic acid or cobalamin, which act directly on the synthesis of a methyl group donor, the epigenetic activity of retinoic acid (RA) and vitamin D3 (Vit.D3) is mainly mediated by an interaction with their receptors. RA, which is generated via oxidative conversion from β-carotene absorbed from fruits and vegetables, binds to nuclear retinoic acid receptors (RARs) α, β and γ that heterodimerize with retinoid X receptors (RXRs). This complex further modulates transcription through cognate response elements in the promoters of target genes including p21 and AP-1 (Liu et al., 1996; Wang et al., 2001; Benkoussa et al., 2002; Yu et al., 2008). RA activated p21 transcription in cervical squamous carcinoma (Wang et al., 2001; Arany et al., 2003), and inhibited AP-1 transcriptional activity in gastric cancer cells (Wu et al., 2002). As p21 competes with DNMT1 for the same binding site on PCNA that is crucial for DNMT1 activity (Chuang et al., 1997; Milutinovic et al., 2000; Iida et al., 2002), RA-mediated p21 induction decreases DNA methylation efficiency (Figure 2B). On the other hand, AP-1 stimulates DNMT1 transcription through binding sites located within the DNMT1 regulatory region (Bigey et al., 2000). Therefore, inhibiton of AP-1 activity by RA will down-regulate AP-1 responsive genes, including DNMT1 (Wu et al., 2002) and decrease DNA methylation (Figure 2C). As changes in DNA methylation affect gene function and signalling pathway networks (Maradeo and Cairns, 2011), a modulation of DNA methylation patterns can contribute to the anti-proliferative, differentiating and pro-apoptotic actions of RA, which have been observed in a variety of cancers, such as leukaemia (Degos et al., 1995), breast (Fujii et al., 2008), ovarian (Zhang et al., 2000), and head and neck cancer (Youssef et al., 2004). Treatment of non-invasive MCF-7 cells with RA caused a reduction in promoter methylation and an increase in the expression of RARβ2 and PTEN tumour suppressor genes, which was associated with inhibition of breast cancer cell growth (Stefanska et al., 2010; 2011b). Although a decrease in DNMT1 mRNA level was not observed in these studies, genome-wide analysis of all-trans RA on gene methylation and expression in neuroblastoma revealed the down-regulation of methyltransferases DNMT1 and DNMT3B, along with the up-regulation of endogenous miRNAs targeting these DNMTs (Das et al., 2010). These changes in DNMT expression were accompanied by the demethylation and overexpression of 82 genes, including NOS1 that is required for neural cell differentiation. MiRNA-mediated attenuation of DNMTs after RA treatment constitutes another mechanism for the demethylating activity of this compound. Furthermore, a decrease in DNA promoter methylation has been linked to the ability of RA to up-regulate the activity of glycine N-methyltransferase (GNMT) that catalyzes the conversion of SAM to SAH (Rowling et al., 2002). Activation of GNMT leads to the loss of methyl groups crucial for SAM-dependent methylation reactions (Figure 1).
Figure 2.
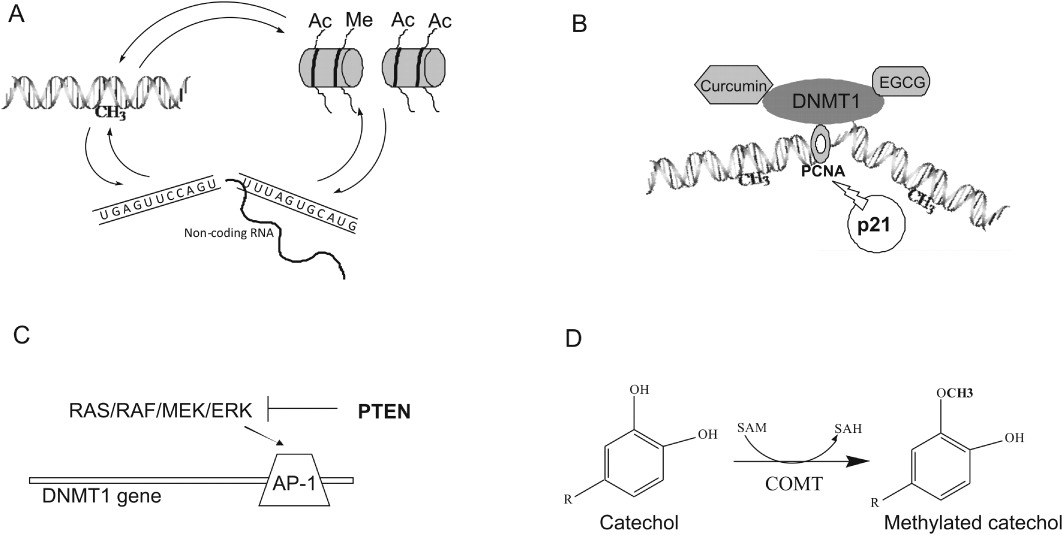
Scheme demonstrating mechanisms used by natural compounds to drive changes in the epigenome. (A) Reciprocal interconnections between all the components of the epigenome: DNA methylation, histone modifications and non-coding RNAs. Compounds driving changes in DNA methylation patterns may have indirect effects on other epigenetic components and vice versa. (B) PCNA is crucial for DNMT1 activity during replication when DNA methylation pattern is copied from a parental to a daughter DNA strand. p21 competes with DNMT1 for the same binding site on PCNA, which impairs DNMT1 activity. Compounds that lead to an increase in p21 expression can affect DNA methylation. Several natural agents such as curcumin and EGCG bind to the DNMT1 catalytic domain and impair its enzymatic activity. (C) AP-1 binding sites have been identified within the DNMT1 regulatory region. Compounds that inhibit the MAPK signalling pathway by, for example, up-regulating PTEN can attenuate DNMT1 transcription. (D) Compounds with a catechol group are excellent substrates for catechol-O-methyltransferase (COMT) that catalyzes their methylation. The COMT-mediated methylation reaction results in depletion of the methyl donor SAM and formation of SAH, which is a potent feedback inhibitor of DNA methylation.
Vit.D3, synthesized upon sun exposure and from diet, acts in a similar manner to RA by binding to vitamin D receptors (VDR) that can homodimerize or heterodimerize with RXR or RAR and affect gene transcription through VDR responsive elements (VDRE) in target genes including p21 (Saramaki et al., 2009) and PTEN (Pan et al., 2010). Epigenetic effects of Vit.D3 that can be partially linked to its activity as a chemopreventive and anti-cancer agent (Dace et al., 1997; Gurlek et al., 2002; Lin et al., 2002; Palmer et al., 2003) were primarily observed as histone modifications, which will be discussed in the next chapter. However, demethylating effects were also reported, for instance in osteocalcin promoter in MG63 osteosarcoma cells (Haslberger et al., 2006) and promoters of tumour suppressor genes in breast cancer cells (Stefanska et al., 2010; 2011b). Exposure to Vit.D3 caused hypomethylation and induction of PTEN and RARβ2 tumour suppressor genes in breast cancer cells (Stefanska et al., 2010; 2011b). Vit.D3-mediated promoter demethylation was also observed in CDH1 gene encoding for E-cadherin, which resulted in the activation of this gene and differentiation of breast cancer cells (Lopes et al., 2012). Similar to RA, changes in DNA methylation patterns induced by Vit.D3 treatment may be attributed to the negative regulation of DNMT1 activity via stimulation of p21 expression and inhibiton of AP-1 activity (Figure 2). In breast cancer, Vit.D3 increases p21 expression (Wang et al., 2001; Stefanska et al., 2011b), whereas in breast cancer and leukaemic cells, Vit.D3 and its derivatives induce PTEN transcription (Hisatake et al., 2001; Stefanska et al., 2011b). Activation of PTEN that is another negative regulator of AP-1 (Chung et al., 2006) may down-regulate AP-1 responsive genes, including DNMT1 (Figure 2C) (Bigey et al., 2000). More interestingly, the effects of Vit.D3 as well as RA seem to depend on cell invasiveness and ER status. Previous studies in human breast cancer cells indicated that activation of ERs stimulates the activity of c-Ha-ras, which leads to induction of the Ras/Raf/MAPK/AP-1 signalling pathway (Pethe and Shekhar, 1999). As the AP-1 transcription complex activates the DNMT1 promoter (Bigey et al., 2000), inactivation of ERs, by causing inhibition of AP-1, can consequently reduce DNMT1 expression. Vit.D3 as well as RA were shown to down-regulate ER abundance and function in MCF-7 cells (Pratt et al., 1996; Swami et al., 2000), which can explain their stronger epigenetic effects in this cell line compared with ER-negative MDA-MB-231 cells (Stefanska et al., 2010; 2011b).
Phytoestrogens
Several polyphenols called phytoestrogens such as resveratrol (RES) and genistein interact with ERs and regulate oestrogen-responsive genes such as p21 (Bowers et al., 2000; Mandal and Davie, 2010). They can act as oestrogen agonists or antagonists depending on their concentrations, tissue type and genes tested on, as well as the presence of oestradiol (Pozo-Guisado et al., 2002; 2004). RES (3,4′,5-trihydroxystilbene) is a dietary antioxidant found in a wide variety of plant species, including mulberries, peanuts, grapes, apricots and pineapples (Le Corre et al., 2004). This polyphenol was shown to modulate the risk of cardiovascular disease (atherosclerosis) and breast cancer, inhibit chemical carcinogenesis in rodents and growth promotion of preneoplastic lesions by effects on multiple signalling pathways, inhibit cell proliferation, block cell cycle progression and induce apoptosis in numerous types of human cancer cell lines (Bowers et al., 2000; Cornwell et al., 2004; Whitsett and Lamartiniere, 2006). Genistein (4′,5,7-trihydroxyisoflavone) is a major isoflavone from soybean. This polyphenol has been shown to prevent carcinogenesis in animal models (Dixon and Ferreira, 2002), reduce mammary carcinogenesis in rats exposed to carcinogens (Cabanes et al., 2004), inhibit growth of transplanted human prostate cancer cells in mice (Zhou et al., 1999) and inhibit carcinogenesis and metastasis in various types of cancer (Arai et al., 2000; Fang et al., 2005). Moreover, it has been reported that genistein inhibits cancer cell proliferation and angiogenesis as well as induces apoptosis and cell cycle arrest (Li et al., 1999; Fang et al., 2005). Dietary intake of genistein decreases mammary tumour incidence and increases tumour latency in rats exposed to chemical carcinogens. These breast cancer protective effects were found to be associated with the down-regulation of the oncogenic Wnt-signalling pathway and the up-regulation of PTEN tumour suppressor gene in the mammary gland, which was coincident with increased apoptosis and enhanced differentiation (Dave et al., 2005; Su et al., 2007). Changes in the epigenome upon treatment with these phytoestrogens have been reported in numerous studies. In non-invasive MCF-7 breast cancer cells, RES exposure led to hypomethylation and reactivation of RARβ2 and PTEN tumour suppressor genes with a concomitant decrease in DNMT1 expression and up-regulation of p21 (Stefanska et al., 2011b). RES prevented BRCA1 promoter hypermethylation and silencing induced by an aromatic hydrocarbon receptor ligand in MCF-7 cells (Papoutsis et al., 2011). Treatment of KYSE510 oesophageal squamous cell carcinoma cells with genistein resulted in a partial reversal of p16, RARβ and MGMT (O6-methylguanine methyltransferase) promoter hypermethylation and in reactivation of these genes, which was associated with the inhibition of DNMT activity (Fang et al., 2005). Similarly, methylation-silenced RARβ was hypomethylated and reactivated in LNCaP and PC3 prostate cancer cells exposed to genistein (Fang et al., 2007). Genistein significantly decreased promoter methylation and reactivated BTG3 tumour suppressor gene in A498, ACHN and HEK-293 renal cell carcinoma cell lines, as well as LNCaP and PC3 prostate carcinoma cell lines (Majid et al., 2009). Its effect was comparable with that of 5-aza-cytydine, which is a potent demethylating agent. Changes in DNA methylation were accompanied by increased histone acetylation and RNA polymerase II binding at the BTG3 promoter, suggesting active chromatin formation (Majid et al., 2009). Daidzein is another phytoestrogen found in soy (Barnes et al., 1996). Multiple studies have indicated that daidzein may act as a growth modulator in various types of cancer, such as colon (Bielecki et al., 2011), prostate (Vardi et al., 2010), breast (Choi and Kim, 2008) and pancreatic cancer (Guo et al., 2004). Treatment of human prostate cancer cells with daidzein led to demethylation of glutathione S-transferase P1 (GSTP1) and ephrin B2 (EPHB2) promoters and to an increase in their protein levels (Vardi et al., 2010). Demethylation and activation of tumour suppressor gene p16 was also detected in RKO colon cancer cells exposed to nordihydroguaiaretic acid found in the creosote bush (Cui et al., 2008) and to quercetin, which is another flavonoid found in a wide variety of vegetables, fruits, leaves and grains (Tan et al., 2009).
The epigenetic activity of phytoestrogens can be attributed to their ability to stimulate p21 via ERE elements in its promoter or other indirect mechanisms (Le Corre et al., 2004; Matsumura et al., 2008; Mandal and Davie, 2010) as well as to suppress AP-1 transcriptional activity via antagonism of ERs (Pethe and Shekhar, 1999) or inducing PTEN expression (Chung et al., 2006). As mentioned earlier, both p21 and AP-1 are involved in the regulation of DNMT1 expression/activity (Figure 2) (Chuang et al., 1997; Bigey et al., 2000; Iida et al., 2002). RES-mediated p21 up-regulation has been reported in many types of cancer, including prostate, bladder, colorectal and breast cancer (Tessitore et al., 2000; Bai et al., 2010; Stefanska et al., 2010; 2011b; Hsieh et al., 2011). Genistein treatment has been shown to induce an up-regulation of p21 in mouse B16-F1 melanoma cells, mouse fibroblasts (Kuzumaki et al., 1998) and breast cancer cells (Privat et al., 2010). Also, exposure to both RES and genistein is associated with an up-regulation of PTEN tumour suppressor gene in breast and prostate cancer (Waite et al., 2005; Kikuno et al., 2008; Rahal and Simmen, 2010; Stefanska et al., 2011b).
Polyphenols with catechol group
Certain polyphenols such as bioflavonoids (quercetin, fisetin and myricetin), tea catechins [catechin, epicatechin, (−)-epigallocatechin-3-gallate (EGCG) ] and coffee polyphenols (caffeic acid, chlorogenic acid) exert a profound inhibitory effect on DNA methylation and DNMT activities (Yang et al., 2008). Treatment of MCF-7 and MDA-MB-231 breast cancer cells with caffeic acid or chlorogenic acid decreased RARβ promoter methylation (Lee and Zhu, 2006). Moreover, consumers of high doses of chlorogenic acid had an increased plasma concentration of total homocysteine, which is an inhibitor of methylation reactions (Olthof et al., 2001). EGCG, a major polyphenol from green tea, inhibited UVB-induced global hypomethylation and protected mice from photocarcinogenesis and transformation of papillomas to carcinomas (Mittal et al., 2003). Exposure of human oesophageal cancer KYSE510 cells to EGCG resulted in demethylation and reactivation of p16, RARβ, MGMT and hMLH1 tumour suppressor genes (Lu et al., 2006). Promoter methylation-mediated silencing of p16 and RARβ was also found to be reversed by EGCG in colon cancer HT-29, prostate cancer PC3, breast cancer MCF-7 and MDA-MB-231 cell lines (Lee et al., 2005; Fang et al., 2007).
The tea catechins and bioflavonoids have also been shown to inhibit the prokaryotic SssI methyltransferase-mediated DNA methylation as well as the human DNMT1-mediated DNA methylation reaction in a concentration-dependent manner with diverse potencies (Lee et al., 2005). The presence of a catechol group in the structure of all these compounds can play a key role in inhibiting methyltransferase activity as compounds with a catechol group are excellent substrates for the methylation mediated by catechol-O-methyltransferase (COMT) (Figure 2D). COMT-mediated methylation results in the depletion of the methyl donor SAM and the formation of SAH, which is a potent feedback inhibitor of DNA methylation (Lee and Zhu, 2006). EGCG can exert a dual inhibitory effect on DNMT-mediated DNA methylation, which explains its high efficacy at inhibiting DNA methylation catalyzed by SssI methyltransferase and human DNMT1 compared with other compounds tested (Lee et al., 2005). Like other catechol-containing polyphenols, EGCG inhibits methyltransferase activity by increase the level of SAH in the reaction catalyzed by COMT. In addition, this tea polyphenol was shown to interact directly with the catalytic site of human DNMT1. Molecular modelling studies indicate that EGCG is well accommodated in a hydrophilic pocket of DNMT1 and effectively tethered within the DNMT1 binding site by at least four hydrogen bonds (Fang et al., 2007) and stabilized by magnesium ions (Figure 2B) (Lee et al., 2005). EGCG analogues without the gallic acid moiety were found to be poor inhibitors of DNMTs, therefore, the galloyl moiety is assumed to be crucial for EGCG binding to DNMTs. Furthermore, myricetin, which has a pyrogallic acid moiety similar to the galloyl group of EGCG, is a more potent, direct inhibitor of DNMT activity than other bioflavonoids, such as quercetin or fisetin, in the presence of magnesium ions (Lee et al., 2005).
Parthenolide and curcumin
Parthenolide was initially shown to have anti-cancer properties by inhibiting NF-κB activation through alkylation of Cys38 of p65 (Wiedhopf et al., 1973; Woynarowski and Konopa, 1981; Garcia-Pineres et al., 2004; Guzman et al., 2005). This compound, with a sesquiterpene lactone structure, is naturally present in the plant feverfew. Recent findings indicate that parthenolide inhibits DNMT1 activity and decreases DNMT1 expression; in K562, Kasumi-1, MV4-11 leukaemia cell lines and MCF-7 breast cancer cells, it decreased the global methylation and led to hypomethylation and activation of the HIN-1 tumour suppressor gene (Liu et al., 2009a). This inhibition of DNMT1 is probably linked to alkylation of the proximal thiolate of Cys1226 of the DNMT1 catalytic domain by parthenolide's γ-methylene lactone, as well as to interference with Sp1 binding to DNMT1 promoter and down-regulation of DNMT1 owing to the induction of subG1 cell cycle arrest (Liu et al., 2009a). Another natural compound directly regulating activity of DNMTs is curcumin, which is present in the spice turmeric. This agent was found to induce global DNA methylation in MV4-11 leukaemia cells (Liu et al., 2009b) and to hypomethylate and reactivate RARβ2 and WIF-1 tumour suppressor genes in human cervical and non-small cell lung cancer cells, respectively (Jha et al., 2010; Liu et al., 2011). Furthermore, exposure to curcumin led to promoter demethylation and re-expression of NEUROG1 and NRF1 in human prostate cancer cells (Khor et al., 2011; Shu et al., 2011). Curcumin can directly interact with SssI methyltransferase, a homologue of DNMT1, and covalently bind to the catalytic thiol group of Cys1226 present in DNMT1, which causes the inhibition of DNMT1 catalytic activity (Figure 2B) (Liu et al., 2009b).
Compounds targeting UHRF1
Down-regulation of UHRF1 (ubiquitin-like containing PHD ring finger 1) may be one of the mechanisms by which natural compounds exert their effect on DNA methylation. Thymoquinone from black cumin oil (Alhosin et al., 2011) and polyphenols from red wine such as RES, anthocyanins and procyanidins (Sharif et al., 2010) have been shown to suppress UHRF. UHRF1 is a putative oncogene that was found to be overexpressed in multiple cancer types such as prostate, lung, cervical, pancreatic, breast, bladder and kidney (Bronner et al., 2007; Unoki et al., 2009). Through its SRA (set and ring associated) domain, UHRF1 recognizes hemimethylated DNA and interacts with DNMT1 in order to form a complex responsible for copying the DNA methylation pattern from a maternal to a daughter strand (Bostick et al., 2007; Sharif et al., 2007). The same domain plays an important role in the cooperation of UHRF1 with HDAC1 (Unoki et al., 2004). UHRF1 up-regulation in cancer is associated with hypermethylation and inhibition of tumour suppressor genes (Daskalos et al., 2011). DNA demethylation caused by natural compounds may be a consequence of UHRF1 inhibition and subsequent attenuation of DNMT activity. UHRF1 suppression may also result from up-regulation of negative regulators such as p53 and p73 that are activated by many natural compounds, leading to cell cycle arrest and apoptosis (Arima et al., 2004; Alhosin et al., 2011).
The impact of natural compounds on histone modifications
Numerous findings demonstrate interactions and feedback regulations between all the components of the epigenome, which should be considered when evaluating epigenetic effects of natural and synthetic agents (Figure 2A) (D'Alessio and Szyf, 2006; Iorio et al., 2010). In addition, many compounds exert a direct effect on both DNA methylation and histone modifications, as well as on miRNAs. For instance, the complex of RA with receptors and coactivators from the p160 and p300 family contains HAT activity which changes chromatin structure, making it more easily accessible for proteins of the transcription complex (Chen et al., 1997; Lamond and Earnshaw, 1998). Genistein-induced changes in DNA methylation are also associated with a concomitant increase in histone acetylation and HAT activity (Majid et al., 2009). Curcumin inhibits the activity of several HDACs, suppressing proliferation and inducing apoptosis of cancer cells (Liu et al., 2005; Chen et al., 2007b). More interestingly, other reports have revealed that curcumin binds p300 HAT, leading to a conformational change in the enzyme and its decreased affinity to histones H3 and H4 and acetyl CoA, which has therapeutic potential for cancer and heart failure (Reuter et al., 2011). Curcumin-mediated hypoacetylation appears to be dependent on reactive oxygen species as antioxidant enzymes diminish its effect (Kang et al., 2005). RES and quercetin activate the protein deacetylase SIRT1 (Borra et al., 2005; Chung et al., 2010), which targets different proteins including histones (Zhang and Kraus, 2010). After binding to SIRT1, RES induces a conformational change in the enzyme, which allows tighter fluorophore and peptide substrate binding to the enzyme (Borra et al., 2005). SIRT1 activation leads to the formation of inactive chromatin and changes in gene transcription (Vaquero et al., 2004). After the other hand, RES activates p300 HAT that participates in the formation of an active chromatin structure (Narayanan et al., 2003). Further, recent studies have shown that RES also inhibits oncogenic miRNAs and induces tumour suppressor miRNAs (Tili et al., 2010). The dual activity of RES on the regulation of histone modifications and targeting non-coding RNAs can partially explain the activation of tumour suppressor genes such as PTEN and PDCD4 and attenuation of expression of prometastatic genes such as TGFβ (Tili et al., 2010). This dual activity was also reported for Vit.D3. The VDR/RXR dimer interacts with HATs such as SRC-1 and CBP/p300 and participates in the activation of transcription (Chakravarti et al., 1996; Voegel et al., 1996). On the other hand, the VDR/RXR complex also binds to the so-called negative VDREs present, amongst others, within CYP24 that recruits transcriptional co-repressors like NCoR and SMRT, leading to histone deacetylation and transcriptional silencing (Sanchez-Martinez et al., 2008). More interestingly, the effects of Vit.D3 are modulated by epigenetic mechanisms that affect VDRs; polymorphic variations of VDRs have been associated variously with cancer development, aggressiveness and metastasis (Ingles et al., 1997; Guy et al., 2004; Khanim et al., 2004; Karlic and Varga, 2011). Pan et al. (2010) found that the HDAC inhibitors, trichostatin A (TSA) and sodium butyrate, and the methylation inhibitor, 5-aza-2'-deoxycytidine (5-azaCdR), reactivate VDR expression and restore the sensitivity of cancer cells to Vit.D3-mediated apoptosis. Targeted co-treatments with Vit.D3 and HDAC inhibitors resulted in re-expression of antiproliferative target genes such as GADD45α and CDKN1A, and synergistic inhibition of proliferation. Microarray studies demonstrated that VDR-reactivation induced by TSA and followed by Vit.D3 treatment uniquely up-regulated a group of target genes associated with the control of proliferation and induction of apoptosis in prostate cancer cells (Khanim et al., 2004). Silencing of VDR by promoter hypermethylation was reported in breast cancer cell lines and primary tumours that were resistant to Vit.D3 (Marik et al., 2010). Treatment with 5-azaCdR restored VDR expression with concurrent reactivation of VDR target genes. Hence, it is increasingly clear that epigenetic mechanisms attenuate and selectively distort the transcriptional responsiveness of VDR, raising the possibility of reversal of Vit.D3 insensitivity by epigenetically active drugs (Karlic et al., 2010a).
A set of natural compounds has been identified as new generation agents that mediate changes in histone, mainly through affecting the activities of histone modifying enzymes. Garlic organosulfur compounds, such as diallyl disulfide, as well as sulforaphane inhibit HDAC activity and affect gene transcription in vitro and in vivo coincident with tumour suppression (Nian et al., 2009; Druesne-Pecollo and Latino-Martel, 2011). Diallyl disulfide can be metabolized to allyl mercaptan that is a competitive HDAC inhibitor. Its anti-proliferative activity was linked to the inhibition of HDAC, histone hyperacetylation and increase in p21 expression in human colon cancer cells (Myzak and Dashwood, 2006; Druesne-Pecollo and Latino-Martel, 2011). Sulforaphane, found in broccoli and broccoli sprouts, induced global histone acetylation and increased histone acetylation associated with p21 and Bax gene promoters in Apcmin mice and human colon and prostate cancer cells (Myzak and Dashwood, 2006; Dashwood and Ho, 2007). Another natural compound tha inhibits HDACs and increases histone acetylation within the p21 promoter is sodium butyrate, a derivative of a short-chain fatty acid present in cheese and butter and produced from dietary fibre in the large intestine (Davie, 2003). Luteolin, a flavonoid found in high concentrations in parsley, thyme, peppermint, basil herb, celery and artichoke, acts as a HDAC inhibitor and is associated with the inhibition of cancer cell growth, survival and invasion, and also potentiated the cytotoxicity of cisplatin in lung LNM35 cells (Attoub et al., 2011).
The impact of natural compounds on the regulation of miRNAs
Aberrant expression of miRNAs, driven by both genetic and epigenetic factors and involved in cancer initiation and progression, has been shown to be reversed by various dietary components (Davis and Ross, 2008; Saini et al., 2010; Parasramka et al., 2012). A recent study by Starlard-Davenport et al. (2010) demonstrated alterations in the expression of miRNAs, induced by methyl deficiency, as a prominent event during the early stages of liver carcinogenesis and suggests that the pattern of miRNAs expression may predict susceptibility to liver carcinogenesis. As mentioned before, rats fed a methyl-deficient diet deprived of folate, methionine and choline developed HCC (Asada et al., 2006). More interestingly, this effect was associated not only with DNA hypomethylation but also with changes in miRNA expression (Davis and Ross, 2008; Saini et al., 2010; Parasramka et al., 2012). MiR-122, that acts as a tumour suppressor in liver, was down-regulated and the levels of miR-34a and miR-127 were also attenuated with concomitant alterations in the expression of E2F3 and Bcl-6 that are their predicted targets, respectively. Several miRNAs were increased including miR-21, which mediates tumour growth and is overexpressed in various types of cancer including breast, colon, lung and leukaemia (Davis and Ross, 2008; Parasramka et al., 2012). Furthermore, changes in miRNAs detected upon culture of human lymphoblastoid cells in media containing low folate levels were reversed after replacement with a complete media. One of the up-regulated oncogenic miRNAs in this culture, miR-222, was found in the blood of individuals with low folate intake (Davis and Ross, 2008; Parasramka et al., 2012), which supports the use of miRNAs as potential cancer biomarkers.
The treatment of acute promyelocytic cells with all-trans RA resulted in alterations in cellular levels of miRNAs implicated in haematopoietic differentiation and apoptosis (Davis and Ross, 2008; Saini et al., 2010; Parasramka et al., 2012). For instance, miR-15/miR-16 and let-7a were up-regulated, which correlated with decreased expression of their targets, Bcl-2 and Ras respectively. Other non-coding RNAs regulated by RA include miR-10a, which promotes metastasis of pancreatic cancer cells, and miR-34a, which acts as a tumour suppressor in neuroblastoma cell lines. Studies have shown that the fusion protein PML/RARα binds to the regulatory regions of certain miRNAs, such as miR-342 and let-7c. Upon RA treatment, the fusion protein is released, leading to transcriptional activation (Saini et al., 2010). This has been proposed to be one of the mechanisms of RA-mediated regulation of miRNAs. Another vitamin implicated in the regulation of miRNAs is Vit.D3. Vit.D3 induces differentiation of human myeloid leukaemia cells, which was associated with p27 up-regulation, and a detailed analysis revealed the concomitant suppression of miR-181a and miR-181b that target p27. More interestingly, this effect of Vit.D3 appears to be interrupted by miR-125b that negatively regulates VDR expression through a potential miR-125b recognition element identified within 3′-UTR of human VDR mRNA (Parasramka et al., 2012).
Exposure of BxPC-3 pancreatic cancer cells to curcumin changes the expression profiles of miRNAs, with a robust increase in the tumour suppressor miR-22 and a decrease in the oncogenic miR-199a (Davis and Ross, 2008; Saini et al., 2010; Parasramka et al., 2012). An up-regulation of miR-22 suppresses the expression of its target genes, Sp1 and ERα. In contrast, miR-21 expression was attenuated by curcumin, which results in the induction of the tumour suppressor PTEN (Parasramka et al., 2012). In another study, curcumin was found to increase the levels of pro-apoptotic miR-15a and miR-16 in MCF-7 breast cancer cells, which was accompanied by the inhibition of their target gene, Bcl-2 (Saini et al., 2010).
Aberrations in the expression of miRNAs and their targets were also observed after exposure to genistein and EGCG. Genistein caused a down-regulation of oncogenic miR-27a in unveal melanoma cells in vitro and in vivo, inhibiting their ability to proliferate (Saini et al., 2010; Parasramka et al., 2012). Furthermore, this isoflavone up-regulated the tumour suppressor miR-1296 in prostate cancer cells, which was accompanied by a decreased expression of its target gene, MCM2, involved in DNA replication and tumorigenesis (Parasramka et al., 2012). Genistein and EGCG augment miR-16 levels in murine chronic lymphocytic leukaemia cells and HCC HepG2 cells, respectively, leading to Bcl-2 inhibition and an increase in apoptosis (Saini et al., 2010; Parasramka et al., 2012).
Given that the expression of miRNAs is dysregulated in numerous types of cancer, a possibility of reversal of the changes by natural compounds may have tremendous potential for cancer prevention.
Bioavailability and dose-dependent effects of natural compounds
Numerous bioactive food components have been shown to possess diverse pharmacological effects, including anti-proliferative, anti-inflammatory, antioxidant, anti-angiogenic and anti-cancer. Clinical trials of natural compounds such as curcumin, RES, EGCG, lycopene, genistein, selenium and Vit.D3 as chemopreventive and therapeutic agents for cancer show poor bioavailability, which raises issues with extrapolation of in vitro results to physiological effects (Bemis et al., 2006; Amin et al., 2009). The main reasons for reduced bioavailability are high rates of metabolism and rapid elimination and clearance from the body. For example, rapid sulfate and glucuronide conjugation by the intestine and liver limits RES and curcumin bioavailability (Walle et al., 2004; Anand et al., 2007). Recent data indicate that administration of RES or curcumin with piperine, an alkaloid derived from black pepper, significantly improves their bioavailability through inhibition of glucuronidation, increasing the degree of exposure and the maximum plasma concentration (Shoba et al., 1998; Johnson et al., 2011). The absorption is even higher when the compounds are entrapped in nanoparticles (Shaikh et al., 2009; Peng et al., 2010). The development of different approaches aimed at increasing the bioavailability of natural agents is of great importance for their efficacy. For instance, stability and metabolism determine the effects of certain polyphenols, including EGCG on SIRT1 stimulation, as the metabolites are less active (de Boer et al., 2006).
As the data from the literature indicate, the effects of several natural compounds depend on their concentrations. Folic acid is an excellent example; it was shown to reduce cancer cell growth, to demethylate and activate methylation-silenced tumour suppressor genes at 0.88 mg·L−1 of media in Caco-2 colon adenocarcinoma cells (Berner et al., 2010). However, a 10-fold higher concentration led to enhanced cancer cell proliferation, with concomitant increase in methylation of tumour suppressor genes such as ERα, p16 and p15 (Berner et al., 2010). Furthermore, oral treatment with folic acid and cobalamin at doses of 0.8 and 0.4 mg·day−1, respectively, was associated with an increased risk of cancer incidence in patients with ischaemic heart disease (Ebbing et al., 2009). Increasing concerns about food fortified with folic acid has resulted in large randomized, placebo-controlled trials, which have not confirmed a statistically significant effect of folic acid supplementation on increased cancer risk (Clarke et al., 2010). Studies in healthy premenopausal women have demonstrated that the action of genistein depends on its circulating levels. Genistein decreases the methylation of RARβ2 and CCND2 at low concentrations, whereas high concentrations increase the methylation of these tumour suppressor genes (Qin et al., 2009). These results should be taken seriously as RARβ2 and CCDN2 hypermethylation is observed at early stages of breast cancer development and is linked to poor prognosis. Other findings demonstrate that RES has no effect on promoter methylation of several tumour suppressor genes such as RASSF1A, GSTP1 and HIN-1 in MCF-7 cells, despite the fact that it attenuates DNMT activity (Paluszczak et al., 2010). The possibility that alterations in DNA methylation are located in regions other than those tested, or that different doses of the compound and/or chronic exposure are needed to observe the epigenetic effect cannot be ruled out.
In the last decade, the anti-cancer activity of multiple bioactive food components with a diverse range of molecular targets has been proven by intensive research but a major concern is their poor bioavailability, which remains a challenge. Attempts at improving their bioavailability are focused on the modulation of the route of administration, structural modifications, blocking metabolic pathways and combinations with other agents.
Future prospects
Numerous data have shown that alterations in epigenetic modifications produce various effects on the phenotype. Compounds that directly affect DNA methylation can induce changes in other components of the epigenome due to a trilateral relationship that exists between DNA methylation, histone covalent modifications and non-coding RNAs. Furthermore, cancer, like other multiple conditions such as diabetes, allergy, rheumatoid arthritis and cardiovascular disease, has been linked to chronic inflammation and metabolic stress. Studies have shown that epigenetics constitute the crossroads of cancer, inflammation and metabolic stress (vel Szic et al., 2010). Signalling pathways responsive to chronic inflammation activate downstream mediators such as transcription factors that drive changes in the expression of target genes. However, binding of the transcription factors to DNA depends on the chromatin structure, which is regulated by epigenetic machinery. Thus, inflammatory gene expression is dependent on epigenetic mechanisms. Reciprocally, inflammatory genes, such as IL-6, have been reported to trigger epigenetic alterations, for instance via the regulation of DNMTs (vel Szic et al., 2010). Dietary phytochemicals that modulate epigenetic components can thereby suppress the expression of inflammatory genes and inflammation-related disorders. Recent studies have added additional layers of complexity, showing that changes in DNA methylation can be transgenerationally transmitted to future generations (Szyf, 2012). Nutritional restriction during pregnancy (Heijmans et al., 2008; Unterberger et al., 2009) and low folic acid content before conception (Sinclair et al., 2007) have been shown to permanently alter DNA methylation patterns, which are linked to chronic diseases in adulthood, such as obesity and cancer (vel Szic et al., 2010). A methyl donor-rich diet or with a high genistein content during pregnancy in Agouti mice increased DNA methylation within a transposable element in the agouti gene, causing changes in fur colour and protecting the offspring from obesity (Waterland and Jirtle, 2003; Dolinoy et al., 2006). Natural compounds are able to modulate our epigenome and probably the epigenome of future generations, which raises questions on phenotypic consequences and pharmacological applications. These important issues should be addressed in future nutritional research. From the experimental evidence discussed in the present review, epigenetic modifications are one of the mechanisms by which natural compounds inhibit cancer cell growth, invasive capacities and metastasis. Epigenetic aberrations lead to dysregulation of gene expression, with the activation of oncogenes and inhibition of tumour suppressor genes at initial and advanced stages of carcinogenesis (Chik et al., 2011). Thus, natural chemicals can be efficacious in both cancer therapy and cancer prevention and are becoming potential targets of drug development. The main challenges in the investigation of phytochemicals as epigenetic agents are new strategies of increasing their bioavailability, assessing the efficacy of the metabolites and determining the role of natural compounds alone or in combination with existing drugs in improving cancer treatment.
Acknowledgments
This work was supported by grants from the Medical University of Lodz, Poland (No. 503/6-099-01/503-01, to K. F.-M.), the Ministry of Science and Higher Education, Poland (No. 2 P05A 036 30, to K. F.-M.) and a fund of the Mayor of the City of Vienna (Medizinisch-Wissenschaftlicher Fonds des Bürgermeisters der Bundeshauptstadt Wien, to H. K.).
Glossary
- DNMTs
DNA methyltransferases
- EGCG
(−)-epigallocatechin-3-gallate
- HATs
histone acetylases
- HCC
hepatocellular carcinoma
- HDACs
histone deacetylases
- MBDs
methyl-CpG-binding domain proteins
- miRNAs
micro-RNAs
- MTHF
methylenetetrahydrofolate
- RA
retinoic acid
- RES
resveratrol
- SAH
S-adenosyl-L-homocysteine
- SAM
S-adenosyl-L-methionine
- UHRF1
ubiquitin-like containing PHD ring finger 1
- Vit.D3
vitamin D3
Conflict of interest
The authors have no conflicts of interest to declare.
References
- Alhosin M, Sharif T, Mousli M, Etienne-Selloum N, Fuhrmann G, Schini-Kerth VB, et al. Down-regulation of UHRF1, associated with re-expression of tumor suppressor genes, is a common feature of natural compounds exhibiting anti-cancer properties. J Exp Clin Cancer Res. 2011;30:41. doi: 10.1186/1756-9966-30-41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amin AR, Kucuk O, Khuri FR, Shin DM. Perspectives for cancer prevention with natural compounds. J Clin Oncol. 2009;27:2712–2725. doi: 10.1200/JCO.2008.20.6235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anand P, Kunnumakkara AB, Newman RA, Aggarwal BB. Bioavailability of curcumin: problems and promises. Mol Pharm. 2007;4:807–818. doi: 10.1021/mp700113r. [DOI] [PubMed] [Google Scholar]
- Arai N, Strom A, Rafter JJ, Gustafsson JA. Estrogen receptor beta mRNA in colon cancer cells: growth effects of estrogen and genistein. Biochem Biophys Res Commun. 2000;270:425–431. doi: 10.1006/bbrc.2000.2444. [DOI] [PubMed] [Google Scholar]
- Arany I, Whitehead WE, Ember IA, Tyring SK. Dose-dependent activation of p21WAF1 transcription by all-trans-acid in cervical squamous carcinoma cells. Anticancer Res. 2003;23:495–497. [PubMed] [Google Scholar]
- Arima Y, Hirota T, Bronner C, Mousli M, Fujiwara T, Niwa S, et al. Down-regulation of nuclear protein ICBP90 by p53/p21Cip1/WAF1-dependent DNA-damage checkpoint signals contributes to cell cycle arrest at G1/S transition. Genes Cells. 2004;9:131–142. doi: 10.1111/j.1356-9597.2004.00710.x. [DOI] [PubMed] [Google Scholar]
- Asada K, Kotake Y, Asada R, Saunders D, Broyles RH, Towner RA, et al. LINE-1 hypomethylation in a choline-deficiency-induced liver cancer in rats: dependence on feeding period. J Biomed Biotechnol. 2006;2006:17142. doi: 10.1155/JBB/2006/17142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Attoub S, Hassan AH, Vanhoecke B, Iratni R, Takahashi T, Gaben AM, et al. Inhibition of cell survival, invasion, tumor growth and histone deacetylase activity by the dietary flavonoid luteolin in human epithelioid cancer cells. Eur J Pharmacol. 2011;651:18–25. doi: 10.1016/j.ejphar.2010.10.063. [DOI] [PubMed] [Google Scholar]
- Bai Y, Mao QQ, Qin J, Zheng XY, Wang YB, Yang K, et al. Resveratrol induces apoptosis and cell cycle arrest of human T24 bladder cancer cells in vitro and inhibits tumor growth in vivo. Cancer Sci. 2010;101:488–493. doi: 10.1111/j.1349-7006.2009.01415.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bannister AJ, Kouzarides T. Regulation of chromatin by histone modifications. Cell Res. 2011;21:381–395. doi: 10.1038/cr.2011.22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barnes S, Sfakianos J, Coward L, Kirk M. Soy isoflavonoids and cancer prevention. Underlying biochemical and pharmacological issues. Adv Exp Med Biol. 1996;401:87–100. doi: 10.1007/978-1-4613-0399-2_7. [DOI] [PubMed] [Google Scholar]
- Barreto G, Schafer A, Marhold J, Stach D, Swaminathan SK, Handa V, et al. Gadd45a promotes epigenetic gene activation by repair-mediated DNA demethylation. Nature. 2007;445:671–675. doi: 10.1038/nature05515. [DOI] [PubMed] [Google Scholar]
- Barski A, Cuddapah S, Cui K, Roh TY, Schones DE, Wang Z, et al. High-resolution profiling of histone methylations in the human genome. Cell. 2007;129:823–837. doi: 10.1016/j.cell.2007.05.009. [DOI] [PubMed] [Google Scholar]
- Batta K, Zhang Z, Yen K, Goffman DB, Pugh BF. Genome-wide function of H2B ubiquitylation in promoter and genic regions. Genes Dev. 2011;25:2254–2265. doi: 10.1101/gad.177238.111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baylin SB, Esteller M, Rountree MR, Bachman KE, Schuebel K, Herman JG. Aberrant patterns of DNA methylation, chromatin formation and gene expression in cancer. Hum Mol Genet. 2001;10:687–692. doi: 10.1093/hmg/10.7.687. [DOI] [PubMed] [Google Scholar]
- Bemis DL, Katz AE, Buttyan R. Clinical trials of natural products as chemopreventive agents for prostate cancer. Expert Opin Investig Drugs. 2006;15:1191–1200. doi: 10.1517/13543784.15.10.1191. [DOI] [PubMed] [Google Scholar]
- Benkoussa M, Brand C, Delmotte MH, Formstecher P, Lefebvre P. Retinoic acid receptors inhibit AP1 activation by regulating extracellular signal-regulated kinase and CBP recruitment to an AP1-responsive promoter. Mol Cell Biol. 2002;22:4522–4534. doi: 10.1128/MCB.22.13.4522-4534.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bergmann A, Lane ME. HIDden targets of microRNAs for growth control. Trends Biochem Sci. 2003;28:461–463. doi: 10.1016/S0968-0004(03)00175-0. [DOI] [PubMed] [Google Scholar]
- Berner C, Aumuller E, Gnauck A, Nestelberger M, Just A, Haslberger AG. Epigenetic control of estrogen receptor expression and tumor suppressor genes is modulated by bioactive food compounds. Ann Nutr Metab. 2010;57:183–189. doi: 10.1159/000321514. [DOI] [PubMed] [Google Scholar]
- Bhattacharya SK, Ramchandani S, Cervoni N, Szyf M. A mammalian protein with specific demethylase activity for mCpG DNA. Nature. 1999;397:579–583. doi: 10.1038/17533. [DOI] [PubMed] [Google Scholar]
- Bielecki A, Roberts J, Mehta R, Raju J. Estrogen receptor-beta mediates the inhibition of DLD-1 human colon adenocarcinoma cells by soy isoflavones. Nutr Cancer. 2011;63:139–150. doi: 10.1080/01635581.2010.516867. [DOI] [PubMed] [Google Scholar]
- Bigey P, Ramchandani S, Theberge J, Araujo FD, Szyf M. Transcriptional regulation of the human DNA methyltransferase (dnmt1) gene. Gene. 2000;242:407–418. doi: 10.1016/s0378-1119(99)00501-6. [DOI] [PubMed] [Google Scholar]
- Bistulfi G, Vandette E, Matsui S, Smiraglia DJ. Mild folate deficiency induces genetic and epigenetic instability and phenotype changes in prostate cancer cells. BMC Biol. 2010;8:6. doi: 10.1186/1741-7007-8-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Boer VC, de Goffau MC, Arts IC, Hollman PC, Keijer J. SIRT1 stimulation by polyphenols is affected by their stability and metabolism. Mech Ageing Dev. 2006;127:618–627. doi: 10.1016/j.mad.2006.02.007. [DOI] [PubMed] [Google Scholar]
- Borra MT, Smith BC, Denu JM. Mechanism of human SIRT1 activation by resveratrol. J Biol Chem. 2005;280:17187–17195. doi: 10.1074/jbc.M501250200. [DOI] [PubMed] [Google Scholar]
- Bostick M, Kim JK, Esteve PO, Clark A, Pradhan S, Jacobsen SE. UHRF1 plays a role in maintaining DNA methylation in mammalian cells. Science. 2007;317:1760–1764. doi: 10.1126/science.1147939. [DOI] [PubMed] [Google Scholar]
- Bourc'his D, Bestor TH. Meiotic catastrophe and retrotransposon reactivation in male germ cells lacking Dnmt3L. Nature. 2004;431:96–99. doi: 10.1038/nature02886. [DOI] [PubMed] [Google Scholar]
- Bourc'his D, Xu GL, Lin CS, Bollman B, Bestor TH. Dnmt3L and the establishment of maternal genomic imprints. Science. 2001;294:2536–2539. doi: 10.1126/science.1065848. [DOI] [PubMed] [Google Scholar]
- Bowers JL, Tyulmenkov VV, Jernigan SC, Klinge CM. Resveratrol acts as a mixed agonist/antagonist for estrogen receptors alpha and beta. Endocrinology. 2000;141:3657–3667. doi: 10.1210/endo.141.10.7721. [DOI] [PubMed] [Google Scholar]
- Bronner C, Achour M, Arima Y, Chataigneau T, Saya H, Schini-Kerth VB. The UHRF family: oncogenes that are drugable targets for cancer therapy in the near future? Pharmacol Ther. 2007;115:419–434. doi: 10.1016/j.pharmthera.2007.06.003. [DOI] [PubMed] [Google Scholar]
- Butterworth CE, Jr, Hatch KD, Gore H, Mueller H, Krumdieck CL. Improvement in cervical dysplasia associated with folic acid therapy in users of oral contraceptives. Am J Clin Nutr. 1982;35:73–82. doi: 10.1093/ajcn/35.1.73. [DOI] [PubMed] [Google Scholar]
- Cabanes A, Wang M, Olivo S, DeAssis S, Gustafsson JA, Khan G, et al. Prepubertal estradiol and genistein exposures up-regulate BRCA1 mRNA and reduce mammary tumorigenesis. Carcinogenesis. 2004;25:741–748. doi: 10.1093/carcin/bgh065. [DOI] [PubMed] [Google Scholar]
- Calvisi DF, Simile MM, Ladu S, Pellegrino R, De Murtas V, Pinna F, et al. Altered methionine metabolism and global DNA methylation in liver cancer: relationship with genomic instability and prognosis. Int J Cancer. 2007;121:2410–2420. doi: 10.1002/ijc.22940. [DOI] [PubMed] [Google Scholar]
- Cedar H, Bergman Y. Linking DNA methylation and histone modification: patterns and paradigms. Nat Rev Genet. 2009;10:295–304. doi: 10.1038/nrg2540. [DOI] [PubMed] [Google Scholar]
- Cervoni N, Szyf M. Demethylase activity is directed by histone acetylation. J Biol Chem. 2001;276:40778–40787. doi: 10.1074/jbc.M103921200. [DOI] [PubMed] [Google Scholar]
- Chagas CE, Bassoli BK, de Souza CA, Deminice R, Junior AA, Paiva SA, et al. Folic acid supplementation during early hepatocarcinogenesis: cellular and molecular effects. Int J Cancer. 2011 doi: 10.1002/ijc.25886. doi: 10.1002/ijc.25886 [Epub ahead of print] [DOI] [PubMed] [Google Scholar]
- Chakravarti D, LaMorte VJ, Nelson MC, Nakajima T, Schulman IG, Juguilon H, et al. Role of CBP/P300 in nuclear receptor signalling. Nature. 1996;383:99–103. doi: 10.1038/383099a0. [DOI] [PubMed] [Google Scholar]
- Chen H, Lin RJ, Schiltz RL, Chakravarti D, Nash A, Nagy L, et al. Nuclear receptor coactivator ACTR is a novel histone acetyltransferase and forms a multimeric activation complex with P/CAF and CBP/p300. Cell. 1997;90:569–580. doi: 10.1016/s0092-8674(00)80516-4. [DOI] [PubMed] [Google Scholar]
- Chen T, Hevi S, Gay F, Tsujimoto N, He T, Zhang B, et al. Complete inactivation of DNMT1 leads to mitotic catastrophe in human cancer cells. Nat Genet. 2007a;39:391–396. doi: 10.1038/ng1982. [DOI] [PubMed] [Google Scholar]
- Chen Y, Shu W, Chen W, Wu Q, Liu H, Cui G. Curcumin, both histone deacetylase and p300/CBP-specific inhibitor, represses the activity of nuclear factor kappa B and Notch 1 in Raji cells. Basic Clin Pharmacol Toxicol. 2007b;101:427–433. doi: 10.1111/j.1742-7843.2007.00142.x. [DOI] [PubMed] [Google Scholar]
- Chik F, Szyf M, Rabbani SA. Role of epigenetics in cancer initiation and progression. Adv Exp Med Biol. 2011;720:91–104. doi: 10.1007/978-1-4614-0254-1_8. [DOI] [PubMed] [Google Scholar]
- Choi EJ, Kim GH. Daidzein causes cell cycle arrest at the G1 and G2/M phases in human breast cancer MCF-7 and MDA-MB-453 cells. Phytomedicine. 2008;15:683–690. doi: 10.1016/j.phymed.2008.04.006. [DOI] [PubMed] [Google Scholar]
- Choudhuri S. From Waddington's epigenetic landscape to small noncoding RNA: some important milestones in the history of epigenetics research. Toxicol Mech Methods. 2011;21:252–274. doi: 10.3109/15376516.2011.559695. [DOI] [PubMed] [Google Scholar]
- Chuang LS, Ian HI, Koh TW, Ng HH, Xu G, Li BF. Human DNA-(cytosine-5) methyltransferase-PCNA complex as a target for p21WAF1. Science. 1997;277:1996–2000. doi: 10.1126/science.277.5334.1996. [DOI] [PubMed] [Google Scholar]
- Chung JH, Ostrowski MC, Romigh T, Minaguchi T, Waite KA, Eng C. The ERK1/2 pathway modulates nuclear PTEN-mediated cell cycle arrest by cyclin D1 transcriptional regulation. Hum Mol Genet. 2006;15:2553–2559. doi: 10.1093/hmg/ddl177. [DOI] [PubMed] [Google Scholar]
- Chung S, Yao H, Caito S, Hwang JW, Arunachalam G, Rahman I. Regulation of SIRT1 in cellular functions: role of polyphenols. Arch Biochem Biophys. 2010;501:79–90. doi: 10.1016/j.abb.2010.05.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clarke R, Halsey J, Lewington S, Lonn E, Armitage J, Manson JE, et al. Effects of lowering homocysteine levels with B vitamins on cardiovascular disease, cancer, and cause-specific mortality: meta-analysis of 8 randomized trials involving 37 485 individuals. Arch Intern Med. 2010;170:1622–1631. doi: 10.1001/archinternmed.2010.348. [DOI] [PubMed] [Google Scholar]
- Comb M, Goodman HM. CpG methylation inhibits proenkephalin gene expression and binding of the transcription factor AP-2. Nucleic Acids Res. 1990;18:3975–3982. doi: 10.1093/nar/18.13.3975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cornwell T, Cohick W, Raskin I. Dietary phytoestrogens and health. Phytochemistry. 2004;65:995–1016. doi: 10.1016/j.phytochem.2004.03.005. [DOI] [PubMed] [Google Scholar]
- Cui Y, Lu C, Liu L, Sun D, Yao N, Tan S, et al. Reactivation of methylation-silenced tumor suppressor gene p16INK4a by nordihydroguaiaretic acid and its implication in G1 cell cycle arrest. Life Sci. 2008;82:247–255. doi: 10.1016/j.lfs.2007.11.013. [DOI] [PubMed] [Google Scholar]
- Dace A, Martin-el Yazidi C, Bonne J, Planells R, Torresani J. Calcitriol is a positive effector of adipose differentiation in the OB 17 cell line: relationship with the adipogenic action of triiodothyronine. Biochem Biophys Res Commun. 1997;232:771–776. doi: 10.1006/bbrc.1997.6372. [DOI] [PubMed] [Google Scholar]
- D'Alessio AC, Szyf M. Epigenetic tete-a-tete: the bilateral relationship between chromatin modifications and DNA methylation. Biochem Cell Biol. 2006;84:463–476. doi: 10.1139/o06-090. [DOI] [PubMed] [Google Scholar]
- Das PM, Singal R. DNA methylation and cancer. J Clin Oncol. 2004;22:4632–4642. doi: 10.1200/JCO.2004.07.151. [DOI] [PubMed] [Google Scholar]
- Das S, Foley N, Bryan K, Watters KM, Bray I, Murphy DM, et al. MicroRNA mediates DNA demethylation events triggered by retinoic acid during neuroblastoma cell differentiation. Cancer Res. 2010;70:7874–7881. doi: 10.1158/0008-5472.CAN-10-1534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dashwood RH, Ho E. Dietary histone deacetylase inhibitors: from cells to mice to man. Semin Cancer Biol. 2007;17:363–369. doi: 10.1016/j.semcancer.2007.04.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Daskalos A, Oleksiewicz U, Filia A, Nikolaidis G, Xinarianos G, Gosney JR, et al. UHRF1-mediated tumor suppressor gene inactivation in nonsmall cell lung cancer. Cancer. 2011;117:1027–1037. doi: 10.1002/cncr.25531. [DOI] [PubMed] [Google Scholar]
- Dave B, Eason RR, Till SR, Geng Y, Velarde MC, Badger TM, et al. The soy isoflavone genistein promotes apoptosis in mammary epithelial cells by inducing the tumor suppressor PTEN. Carcinogenesis. 2005;26:1793–1803. doi: 10.1093/carcin/bgi131. [DOI] [PubMed] [Google Scholar]
- Davie JR. Inhibition of histone deacetylase activity by butyrate. J Nutr. 2003;133:2485S–2493S. doi: 10.1093/jn/133.7.2485S. [DOI] [PubMed] [Google Scholar]
- Davis CD, Ross SA. Evidence for dietary regulation of microRNA expression in cancer cells. Nutr Rev. 2008;66:477–482. doi: 10.1111/j.1753-4887.2008.00080.x. [DOI] [PubMed] [Google Scholar]
- Davis CD, Uthus EO. DNA methylation, cancer susceptibility, and nutrient interactions. Exp Biol Med (Maywood) 2004;229:988–995. doi: 10.1177/153537020422901002. [DOI] [PubMed] [Google Scholar]
- Degos L, Dombret H, Chomienne C, Daniel MT, Miclea JM, Chastang C, et al. All-trans-retinoic acid as a differentiating agent in the treatment of acute promyelocytic leukemia. Blood. 1995;85:2643–2653. [PubMed] [Google Scholar]
- Detich N, Theberge J, Szyf M. Promoter-specific activation and demethylation by MBD2/demethylase. J Biol Chem. 2002;277:35791–35794. doi: 10.1074/jbc.C200408200. [DOI] [PubMed] [Google Scholar]
- Dixon RA, Ferreira D. Genistein. Phytochemistry. 2002;60:205–211. doi: 10.1016/s0031-9422(02)00116-4. [DOI] [PubMed] [Google Scholar]
- Dolinoy DC, Weidman JR, Waterland RA, Jirtle RL. Maternal genistein alters coat color and protects Avy mouse offspring from obesity by modifying the fetal epigenome. Environ Health Perspect. 2006;114:567–572. doi: 10.1289/ehp.8700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Druesne-Pecollo N, Latino-Martel P. Modulation of histone acetylation by garlic sulfur compounds. Anticancer Agents Med Chem. 2011;11:254–259. doi: 10.2174/187152011795347540. [DOI] [PubMed] [Google Scholar]
- Duthie SJ. Folate and cancer: how DNA damage, repair and methylation impact on colon carcinogenesis. J Inherit Metab Dis. 2011;34:101–109. doi: 10.1007/s10545-010-9128-0. [DOI] [PubMed] [Google Scholar]
- Duthie SJ, Narayanan S, Sharp L, Little J, Basten G, Powers H. Folate, DNA stability and colo-rectal neoplasia. Proc Nutr Soc. 2004;63:571–578. doi: 10.1079/pns2004. [DOI] [PubMed] [Google Scholar]
- Ebbing M, Bonaa KH, Nygard O, Arnesen E, Ueland PM, Nordrehaug JE, et al. Cancer incidence and mortality after treatment with folic acid and vitamin B12. JAMA. 2009;302:2119–2126. doi: 10.1001/jama.2009.1622. [DOI] [PubMed] [Google Scholar]
- Eden S, Hashimshony T, Keshet I, Cedar H, Thorne AW. DNA methylation models histone acetylation. Nature. 1998;394:842. doi: 10.1038/29680. [DOI] [PubMed] [Google Scholar]
- Fahrner JA, Eguchi S, Herman JG, Baylin SB. Dependence of histone modifications and gene expression on DNA hypermethylation in cancer. Cancer Res. 2002;62:7213–7218. [PubMed] [Google Scholar]
- Fang M, Chen D, Yang CS. Dietary polyphenols may affect DNA methylation. J Nutr. 2007;137:223S–228S. doi: 10.1093/jn/137.1.223S. [DOI] [PubMed] [Google Scholar]
- Fang MZ, Chen D, Sun Y, Jin Z, Christman JK, Yang CS. Reversal of hypermethylation and reactivation of p16INK4a, RARbeta, and MGMT genes by genistein and other isoflavones from soy. Clin Cancer Res. 2005;11:7033–7041. doi: 10.1158/1078-0432.CCR-05-0406. [DOI] [PubMed] [Google Scholar]
- Feng J, Zhou Y, Campbell SL, Le T, Li E, Sweatt JD, et al. Dnmt1 and Dnmt3a maintain DNA methylation and regulate synaptic function in adult forebrain neurons. Nat Neurosci. 2010;13:423–430. doi: 10.1038/nn.2514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fini L, Selgrad M, Fogliano V, Graziani G, Romano M, Hotchkiss E, et al. Annurca apple polyphenols have potent demethylating activity and can reactivate silenced tumor suppressor genes in colorectal cancer cells. J Nutr. 2007;137:2622–2628. doi: 10.1093/jn/137.12.2622. [DOI] [PubMed] [Google Scholar]
- Fini L, Piazzi G, Daoud Y, Selgrad M, Maegawa S, Garcia M, et al. Chemoprevention of intestinal polyps in ApcMin/+ mice fed with western or balanced diets by drinking annurca apple polyphenol extract. Cancer Prev Res (Phila) 2011;4:907–915. doi: 10.1158/1940-6207.CAPR-10-0359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fire A, Xu S, Montgomery MK, Kostas SA, Driver SE, Mello CC. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature. 1998;391:806–811. doi: 10.1038/35888. [DOI] [PubMed] [Google Scholar]
- Fujii T, Yokoyama G, Takahashi H, Namoto R, Nakagawa S, Toh U, et al. Preclinical studies of molecular-targeting diagnostic and therapeutic strategies against breast cancer. Breast Cancer. 2008;15:73–78. doi: 10.1007/s12282-007-0015-y. [DOI] [PubMed] [Google Scholar]
- Fullgrabe J, Kavanagh E, Joseph B. Histone onco-modifications. Oncogene. 2011;30:3391–3403. doi: 10.1038/onc.2011.121. [DOI] [PubMed] [Google Scholar]
- Garcia-Pineres AJ, Lindenmeyer MT, Merfort I. Role of cysteine residues of p65/NF-kappaB on the inhibition by the sesquiterpene lactone parthenolide and N-ethyl maleimide, and on its transactivating potential. Life Sci. 2004;75:841–856. doi: 10.1016/j.lfs.2004.01.024. [DOI] [PubMed] [Google Scholar]
- Ghoshal K, Li X, Datta J, Bai S, Pogribny I, Pogribny M, et al. A folate- and methyl-deficient diet alters the expression of DNA methyltransferases and methyl CpG binding proteins involved in epigenetic gene silencing in livers of F344 rats. J Nutr. 2006;136:1522–1527. doi: 10.1093/jn/136.6.1522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gibbons RJ, McDowell TL, Raman S, O'Rourke DM, Garrick D, Ayyub H, et al. Mutations in ATRX, encoding a SWI/SNF-like protein, cause diverse changes in the pattern of DNA methylation. Nat Genet. 2000;24:368–371. doi: 10.1038/74191. [DOI] [PubMed] [Google Scholar]
- Gruenbaum Y, Naveh-Many T, Cedar H, Razin A. Sequence specificity of methylation in higher plant DNA. Nature. 1981;292:860–862. doi: 10.1038/292860a0. [DOI] [PubMed] [Google Scholar]
- Guo JM, Xiao BX, Dai DJ, Liu Q, Ma HH. Effects of daidzein on estrogen-receptor-positive and negative pancreatic cancer cells in vitro. World J Gastroenterol. 2004;10:860–863. doi: 10.3748/wjg.v10.i6.860. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gurlek A, Pittelkow MR, Kumar R. Modulation of growth factor/cytokine synthesis and signaling by 1alpha,25-dihydroxyvitamin D(3): implications in cell growth and differentiation. Endocr Rev. 2002;23:763–786. doi: 10.1210/er.2001-0044. [DOI] [PubMed] [Google Scholar]
- Guy M, Lowe LC, Bretherton-Watt D, Mansi JL, Peckitt C, Bliss J, et al. Vitamin D receptor gene polymorphisms and breast cancer risk. Clin Cancer Res. 2004;10:5472–5481. doi: 10.1158/1078-0432.CCR-04-0206. [DOI] [PubMed] [Google Scholar]
- Guzman ML, Rossi RM, Karnischky L, Li X, Peterson DR, Howard DS, et al. The sesquiterpene lactone parthenolide induces apoptosis of human acute myelogenous leukemia stem and progenitor cells. Blood. 2005;105:4163–4169. doi: 10.1182/blood-2004-10-4135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hamm S, Just G, Lacoste N, Moitessier N, Szyf M, Mamer O. On the mechanism of demethylation of 5-methylcytosine in DNA. Bioorg Med Chem Lett. 2008;18:1046–1049. doi: 10.1016/j.bmcl.2007.12.027. [DOI] [PubMed] [Google Scholar]
- Haslberger A, Varga F, Karlic H. Recursive causality in evolution: a model for epigenetic mechanisms in cancer development. Med Hypotheses. 2006;67:1448–1454. doi: 10.1016/j.mehy.2006.05.047. [DOI] [PubMed] [Google Scholar]
- Hatziapostolou M, Iliopoulos D. Epigenetic aberrations during oncogenesis. Cell Mol Life Sci. 2011;68:1681–1702. doi: 10.1007/s00018-010-0624-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heijmans BT, Tobi EW, Stein AD, Putter H, Blauw GJ, Susser ES, et al. Persistent epigenetic differences associated with prenatal exposure to famine in humans. Proc Natl Acad Sci U S A. 2008;105:17046–17049. doi: 10.1073/pnas.0806560105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hermann A, Gowher H, Jeltsch A. Biochemistry and biology of mammalian DNA methyltransferases. Cell Mol Life Sci. 2004;61:2571–2587. doi: 10.1007/s00018-004-4201-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hisatake J, O'Kelly J, Uskokovic MR, Tomoyasu S, Koeffler HP. Novel vitamin D(3) analog, 21-(3-methyl-3-hydroxy-butyl)-19-nor D(3), that modulates cell growth, differentiation, apoptosis, cell cycle, and induction of PTEN in leukemic cells. Blood. 2001;97:2427–2433. doi: 10.1182/blood.v97.8.2427. [DOI] [PubMed] [Google Scholar]
- Hotchkiss RD. The quantitative separation of purines, pyrimidines, and nucleosides by paper chromatography. J Biol Chem. 1948;175:315–332. [PubMed] [Google Scholar]
- Hsieh TC, Huang YC, Wu JM. Control of prostate cell growth, DNA damage and repair and gene expression by resveratrol analogues, in vitro. Carcinogenesis. 2011;32:93–101. doi: 10.1093/carcin/bgq230. [DOI] [PubMed] [Google Scholar]
- Iida T, Suetake I, Tajima S, Morioka H, Ohta S, Obuse C, et al. PCNA clamp facilitates action of DNA cytosine methyltransferase 1 on hemimethylated DNA. Genes Cells. 2002;7:997–1007. doi: 10.1046/j.1365-2443.2002.00584.x. [DOI] [PubMed] [Google Scholar]
- Inamdar NM, Ehrlich KC, Ehrlich M. CpG methylation inhibits binding of several sequence-specific DNA-binding proteins from pea, wheat, soybean and cauliflower. Plant Mol Biol. 1991;17:111–123. doi: 10.1007/BF00036811. [DOI] [PubMed] [Google Scholar]
- Ingles SA, Ross RK, Yu MC, Irvine RA, La Pera G, Haile RW, et al. Association of prostate cancer risk with genetic polymorphisms in vitamin D receptor and androgen receptor. J Natl Cancer Inst. 1997;89:166–170. doi: 10.1093/jnci/89.2.166. [DOI] [PubMed] [Google Scholar]
- Iorio MV, Piovan C, Croce CM. Interplay between microRNAs and the epigenetic machinery: an intricate network. Biochim Biophys Acta. 2010;1799:694–701. doi: 10.1016/j.bbagrm.2010.05.005. [DOI] [PubMed] [Google Scholar]
- Ito S, D'Alessio AC, Taranova OV, Hong K, Sowers LC, Zhang Y. Role of Tet proteins in 5mC to 5hmC conversion, ES-cell self-renewal and inner cell mass specification. Nature. 2010;466:1129–1133. doi: 10.1038/nature09303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- James SJ, Pogribny IP, Pogribna M, Miller BJ, Jernigan S, Melnyk S. Mechanisms of DNA damage, DNA hypomethylation, and tumor progression in the folate/methyl-deficient rat model of hepatocarcinogenesis. J Nutr. 2003;133:3740S–3747S. doi: 10.1093/jn/133.11.3740S. [DOI] [PubMed] [Google Scholar]
- Jenuwein T, Allis CD. Translating the histone code. Science. 2001;293:1074–1080. doi: 10.1126/science.1063127. [DOI] [PubMed] [Google Scholar]
- Jha AK, Nikbakht M, Parashar G, Shrivastava A, Capalash N, Kaur J. Reversal of hypermethylation and reactivation of the RARbeta2 gene by natural compounds in cervical cancer cell lines. Folia Biol (Praha) 2010;56:195–200. doi: 10.14712/fb2010056050195. [DOI] [PubMed] [Google Scholar]
- Johnson JJ, Nihal M, Siddiqui IA, Scarlett CO, Bailey HH, Mukhtar H, et al. Enhancing the bioavailability of resveratrol by combining it with piperine. Mol Nutr Food Res. 2011;55:1169–1176. doi: 10.1002/mnfr.201100117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kanduc D, Ghoshal A, Quagliariello E, Farber E. DNA hypomethylation in ethionine-induced rat preneoplastic hepatocyte nodules. Biochem Biophys Res Commun. 1988;150:739–744. doi: 10.1016/0006-291x(88)90453-6. [DOI] [PubMed] [Google Scholar]
- Kang J, Chen J, Shi Y, Jia J, Zhang Y. Curcumin-induced histone hypoacetylation: the role of reactive oxygen species. Biochem Pharmacol. 2005;69:1205–1213. doi: 10.1016/j.bcp.2005.01.014. [DOI] [PubMed] [Google Scholar]
- Kanwal R, Gupta S. Epigenetics and cancer. J Appl Physiol. 2010;109:598–605. doi: 10.1152/japplphysiol.00066.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kanwal R, Gupta S. Epigenetic modifications in cancer. Clin Genet. 2011;81:303–311. doi: 10.1111/j.1399-0004.2011.01809.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karlic H, Varga F. Impact of vitamin D metabolism on clinical epigenetics. Clin Epigenet. 2011;2:55–61. doi: 10.1007/s13148-011-0021-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karlic H, Thaler R, Varga J, Berger C, Spitzer S, Pfeilstöcker M, et al. Effects of epigenetic drugs (vorinostat, decitabine) on metabolism-related pathway factors in leukemic cells. Open Leuk J. 2010a;3:34–42. [Google Scholar]
- Karlic R, Chung HR, Lasserre J, Vlahovicek K, Vingron M. Histone modification levels are predictive for gene expression. Proc Natl Acad Sci U S A. 2010b;107:2926–2931. doi: 10.1073/pnas.0909344107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khanim FL, Gommersall LM, Wood VH, Smith KL, Montalvo L, O'Neill LP, et al. Altered SMRT levels disrupt vitamin D3 receptor signalling in prostate cancer cells. Oncogene. 2004;23:6712–6725. doi: 10.1038/sj.onc.1207772. [DOI] [PubMed] [Google Scholar]
- Khor TO, Huang Y, Wu TY, Shu L, Lee J, Kong AN. Pharmacodynamics of curcumin as DNA hypomethylation agent in restoring the expression of Nrf2 via promoter CpGs demethylation. Biochem Pharmacol. 2011;82:1073–1078. doi: 10.1016/j.bcp.2011.07.065. [DOI] [PubMed] [Google Scholar]
- Kikuno N, Shiina H, Urakami S, Kawamoto K, Hirata H, Tanaka Y, et al. Genistein mediated histone acetylation and demethylation activates tumor suppressor genes in prostate cancer cells. Int J Cancer. 2008;123:552–560. doi: 10.1002/ijc.23590. [DOI] [PubMed] [Google Scholar]
- Kim YI. Folate and DNA methylation: a mechanistic link between folate deficiency and colorectal cancer? Cancer Epidemiol Biomarkers Prev. 2004;13:511–519. [PubMed] [Google Scholar]
- Koch CM, Andrews RM, Flicek P, Dillon SC, Karaoz U, Clelland GK, et al. The landscape of histone modifications across 1% of the human genome in five human cell lines. Genome Res. 2007;17:691–707. doi: 10.1101/gr.5704207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kouzarides T. Chromatin modifications and their function. Cell. 2007;128:693–705. doi: 10.1016/j.cell.2007.02.005. [DOI] [PubMed] [Google Scholar]
- Kraunz KS, Nelson HH, Lemos M, Godleski JJ, Wiencke JK, Kelsey KT. Homozygous deletion of p16INK4a and tobacco carcinogen exposure in nonsmall cell lung cancer. Int J Cancer. 2006;118:1364–1369. doi: 10.1002/ijc.21522. [DOI] [PubMed] [Google Scholar]
- Kriaucionis S, Heintz N. The nuclear DNA base 5-hydroxymethylcytosine is present in Purkinje neurons and the brain. Science. 2009;324:929–930. doi: 10.1126/science.1169786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuzumaki T, Kobayashi T, Ishikawa K. Genistein induces p21(Cip1/WAF1) expression and blocks the G1 to S phase transition in mouse fibroblast and melanoma cells. Biochem Biophys Res Commun. 1998;251:291–295. doi: 10.1006/bbrc.1998.9462. [DOI] [PubMed] [Google Scholar]
- Lamond AI, Earnshaw WC. Structure and function in the nucleus. Science. 1998;280:547–553. doi: 10.1126/science.280.5363.547. [DOI] [PubMed] [Google Scholar]
- Le Corre L, Fustier P, Chalabi N, Bignon YJ, Bernard-Gallon D. Effects of resveratrol on the expression of a panel of genes interacting with the BRCA1 oncosuppressor in human breast cell lines. Clin Chim Acta. 2004;344:115–121. doi: 10.1016/j.cccn.2004.02.024. [DOI] [PubMed] [Google Scholar]
- Lee WJ, Zhu BT. Inhibition of DNA methylation by caffeic acid and chlorogenic acid, two common catechol-containing coffee polyphenols. Carcinogenesis. 2006;27:269–277. doi: 10.1093/carcin/bgi206. [DOI] [PubMed] [Google Scholar]
- Lee WJ, Shim JY, Zhu BT. Mechanisms for the inhibition of DNA methyltransferases by tea catechins and bioflavonoids. Mol Pharmacol. 2005;68:1018–1030. doi: 10.1124/mol.104.008367. [DOI] [PubMed] [Google Scholar]
- Levenson JM, Roth TL, Lubin FD, Miller CA, Huang IC, Desai P, et al. Evidence that DNA (cytosine-5) methyltransferase regulates synaptic plasticity in the hippocampus. J Biol Chem. 2006;281:15763–15773. doi: 10.1074/jbc.M511767200. [DOI] [PubMed] [Google Scholar]
- Li B, Carey M, Workman JL. The role of chromatin during transcription. Cell. 2007;128:707–719. doi: 10.1016/j.cell.2007.01.015. [DOI] [PubMed] [Google Scholar]
- Li Y, Upadhyay S, Bhuiyan M, Sarkar FH. Induction of apoptosis in breast cancer cells MDA-MB-231 by genistein. Oncogene. 1999;18:3166–3172. doi: 10.1038/sj.onc.1202650. [DOI] [PubMed] [Google Scholar]
- Lin R, Nagai Y, Sladek R, Bastien Y, Ho J, Petrecca K, et al. Expression profiling in squamous carcinoma cells reveals pleiotropic effects of vitamin D3 analog EB1089 signaling on cell proliferation, differentiation, and immune system regulation. Mol Endocrinol. 2002;16:1243–1256. doi: 10.1210/mend.16.6.0874. [DOI] [PubMed] [Google Scholar]
- Lister R, Pelizzola M, Dowen RH, Hawkins RD, Hon G, Tonti-Filippini J, et al. Human DNA methylomes at base resolution show widespread epigenomic differences. Nature. 2009;462:315–322. doi: 10.1038/nature08514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu HL, Chen Y, Cui GH, Zhou JF. Curcumin, a potent anti-tumor reagent, is a novel histone deacetylase inhibitor regulating B-NHL cell line Raji proliferation. Acta Pharmacol Sin. 2005;26:603–609. doi: 10.1111/j.1745-7254.2005.00081.x. [DOI] [PubMed] [Google Scholar]
- Liu M, Iavarone A, Freedman LP. Transcriptional activation of the human p21(WAF1/CIP1) gene by retinoic acid receptor. Correlation with retinoid induction of U937 cell differentiation. J Biol Chem. 1996;271:31723–31728. doi: 10.1074/jbc.271.49.31723. [DOI] [PubMed] [Google Scholar]
- Liu YL, Yang HP, Gong L, Tang CL, Wang HJ. Hypomethylation effects of curcumin, demethoxycurcumin and bisdemethoxycurcumin on WIF-1 promoter in non-small cell lung cancer cell lines. Mol Med Report. 2011;4:675–679. doi: 10.3892/mmr.2011.473. [DOI] [PubMed] [Google Scholar]
- Liu Z, Liu S, Xie Z, Pavlovicz RE, Wu J, Chen P, et al. Modulation of DNA methylation by a sesquiterpene lactone parthenolide. J Pharmacol Exp Ther. 2009a;329:505–514. doi: 10.1124/jpet.108.147934. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu Z, Xie Z, Jones W, Pavlovicz RE, Liu S, Yu J, et al. Curcumin is a potent DNA hypomethylation agent. Bioorg Med Chem Lett. 2009b;19:706–709. doi: 10.1016/j.bmcl.2008.12.041. [DOI] [PubMed] [Google Scholar]
- Lopes N, Carvalho J, Duraes C, Sousa B, Gomes M, Costa JL, et al. 1Alpha,25-dihydroxyvitamin D3 Induces de novo E-cadherin Expression in Triple-negative Breast Cancer Cells by CDH1-promoter Demethylation. Anticancer Res. 2012;32:249–257. [PubMed] [Google Scholar]
- Lovat F, Valeri N, Croce CM. MicroRNAs in the pathogenesis of cancer. Semin Oncol. 2011;38:724–733. doi: 10.1053/j.seminoncol.2011.08.006. [DOI] [PubMed] [Google Scholar]
- Lu J, Getz G, Miska EA, Alvarez-Saavedra E, Lamb J, Peck D, et al. MicroRNA expression profiles classify human cancers. Nature. 2005;435:834–838. doi: 10.1038/nature03702. [DOI] [PubMed] [Google Scholar]
- Lu Q, Qiu X, Hu N, Wen H, Su Y, Richardson BC. Epigenetics, disease, and therapeutic interventions. Ageing Res Rev. 2006;5:449–467. doi: 10.1016/j.arr.2006.07.001. [DOI] [PubMed] [Google Scholar]
- Majid S, Dar AA, Ahmad AE, Hirata H, Kawakami K, Shahryari V, et al. BTG3 tumor suppressor gene promoter demethylation, histone modification and cell cycle arrest by genistein in renal cancer. Carcinogenesis. 2009;30:662–670. doi: 10.1093/carcin/bgp042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mandal S, Davie JR. Estrogen regulated expression of the p21 Waf1/Cip1 gene in estrogen receptor positive human breast cancer cells. J Cell Physiol. 2010;224:28–32. doi: 10.1002/jcp.22078. [DOI] [PubMed] [Google Scholar]
- Maradeo ME, Cairns P. Translational application of epigenetic alterations: ovarian cancer as a model. FEBS Lett. 2011;585:2112–2120. doi: 10.1016/j.febslet.2011.03.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marik R, Fackler M, Gabrielson E, Zeiger MA, Sukumar S, Stearns V, et al. DNA methylation-related vitamin D receptor insensitivity in breast cancer. Cancer Biol Ther. 2010;10:44–53. doi: 10.4161/cbt.10.1.11994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maruti SS, Ulrich CM, White E. Folate and one-carbon metabolism nutrients from supplements and diet in relation to breast cancer risk. Am J Clin Nutr. 2009;89:624–633. doi: 10.3945/ajcn.2008.26568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mastronardi FG, Noor A, Wood DD, Paton T, Moscarello MA. Peptidyl argininedeiminase 2 CpG island in multiple sclerosis white matter is hypomethylated. J Neurosci Res. 2007;85:2006–2016. doi: 10.1002/jnr.21329. [DOI] [PubMed] [Google Scholar]
- Matsumura K, Tanaka T, Kawashima H, Nakatani T. Involvement of the estrogen receptor beta in genistein-induced expression of p21(waf1/cip1) in PC-3 prostate cancer cells. Anticancer Res. 2008;28:709–714. [PubMed] [Google Scholar]
- Miller CA, Sweatt JD. Covalent modification of DNA regulates memory formation. Neuron. 2007;53:857–869. doi: 10.1016/j.neuron.2007.02.022. [DOI] [PubMed] [Google Scholar]
- Milutinovic S, Knox JD, Szyf M. DNA methyltransferase inhibition induces the transcription of the tumor suppressor p21(WAF1/CIP1/sdi1) J Biol Chem. 2000;275:6353–6359. doi: 10.1074/jbc.275.9.6353. [DOI] [PubMed] [Google Scholar]
- Mittal A, Piyathilake C, Hara Y, Katiyar SK. Exceptionally high protection of photocarcinogenesis by topical application of (−)-epigallocatechin-3-gallate in hydrophilic cream in SKH-1 hairless mouse model: relationship to inhibition of UVB-induced global DNA hypomethylation. Neoplasia. 2003;5:555–565. doi: 10.1016/s1476-5586(03)80039-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Myzak MC, Dashwood RH. Histone deacetylases as targets for dietary cancer preventive agents: lessons learned with butyrate, diallyl disulfide, and sulforaphane. Curr Drug Targets. 2006;7:443–452. doi: 10.2174/138945006776359467. [DOI] [PubMed] [Google Scholar]
- Nan X, Campoy FJ, Bird A. MeCP2 is a transcriptional repressor with abundant binding sites in genomic chromatin. Cell. 1997;88:471–481. doi: 10.1016/s0092-8674(00)81887-5. [DOI] [PubMed] [Google Scholar]
- Narayanan BA, Narayanan NK, Re GG, Nixon DW. Differential expression of genes induced by resveratrol in LNCaP cells: P53-mediated molecular targets. Int J Cancer. 2003;104:204–212. doi: 10.1002/ijc.10932. [DOI] [PubMed] [Google Scholar]
- Ndlovu MN, Denis H, Fuks F. Exposing the DNA methylome iceberg. Trends Biochem Sci. 2011;36:381–387. doi: 10.1016/j.tibs.2011.03.002. [DOI] [PubMed] [Google Scholar]
- Newberne PM, Rogers AE. Labile methyl groups and the promotion of cancer. Annu Rev Nutr. 1986;6:407–432. doi: 10.1146/annurev.nu.06.070186.002203. [DOI] [PubMed] [Google Scholar]
- Nian H, Delage B, Ho E, Dashwood RH. Modulation of histone deacetylase activity by dietary isothiocyanates and allyl sulfides: studies with sulforaphane and garlic organosulfur compounds. Environ Mol Mutagen. 2009;50:213–221. doi: 10.1002/em.20454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Okano M, Xie S, Li E. Cloning and characterization of a family of novel mammalian DNA (cytosine-5) methyltransferases. Nat Genet. 1998;19:219–220. doi: 10.1038/890. [DOI] [PubMed] [Google Scholar]
- Okano M, Bell DW, Haber DA, Li E. DNA methyltransferases Dnmt3a and Dnmt3b are essential for de novo methylation and mammalian development. Cell. 1999;99:247–257. doi: 10.1016/s0092-8674(00)81656-6. [DOI] [PubMed] [Google Scholar]
- Olthof MR, Hollman PC, Zock PL, Katan MB. Consumption of high doses of chlorogenic acid, present in coffee, or of black tea increases plasma total homocysteine concentrations in humans. Am J Clin Nutr. 2001;73:532–538. doi: 10.1093/ajcn/73.3.532. [DOI] [PubMed] [Google Scholar]
- Palmer HG, Sanchez-Carbayo M, Ordonez-Moran P, Larriba MJ, Cordon-Cardo C, Munoz A. Genetic signatures of differentiation induced by 1alpha,25-dihydroxyvitamin D3 in human colon cancer cells. Cancer Res. 2003;63:7799–7806. [PubMed] [Google Scholar]
- Paluszczak J, Baer-Dubowska W. Epigenome and cancer: new possibilities of cancer prevention and therapy? Postepy Biochem. 2005;51:244–250. [PubMed] [Google Scholar]
- Paluszczak J, Krajka-Kuzniak V, Baer-Dubowska W. The effect of dietary polyphenols on the epigenetic regulation of gene expression in MCF7 breast cancer cells. Toxicol Lett. 2010;192:119–125. doi: 10.1016/j.toxlet.2009.10.010. [DOI] [PubMed] [Google Scholar]
- Pan L, Matloob AF, Du J, Pan H, Dong Z, Zhao J, et al. Vitamin D stimulates apoptosis in gastric cancer cells in synergy with trichostatin A/sodium butyrate-induced and 5-aza-2′-deoxycytidine-induced PTEN upregulation. FEBS J. 2010;277:989–999. doi: 10.1111/j.1742-4658.2009.07542.x. [DOI] [PubMed] [Google Scholar]
- Papoutsis AJ, Borg JL, Selmin OI, Romagnolo DF. BRCA-1 promoter hypermethylation and silencing induced by the aromatic hydrocarbon receptor-ligand TCDD are prevented by resveratrol in MCF-7 Cells. J Nutr Biochem. 2011 doi: 10.1016/j.jnutbio.2011.08.001. doi: 10.1016/j.jnutbio.2011.08.001 [Epub ahead of print] [DOI] [PubMed] [Google Scholar]
- Parasramka MA, Ho E, Williams DE, Dashwood RH. MicroRNAs, diet, and cancer: new mechanistic insights on the epigenetic actions of phytochemicals. Mol Carcinog. 2012;51:213–230. doi: 10.1002/mc.20822. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peng H, Xiong H, Li J, Xie M, Liu Y, Bai C, et al. Vanilin cross-linked chitosan microspheres for controlled release of resveratrol. Food Chem. 2010;121:23–28. [Google Scholar]
- Pethe V, Shekhar PV. Estrogen inducibility of c-Ha-ras transcription in breast cancer cells. Identification of functional estrogen-responsive transcriptional regulatory elements in exon 1/intron 1 of the c-Ha-ras gene. J Biol Chem. 1999;274:30969–30978. doi: 10.1074/jbc.274.43.30969. [DOI] [PubMed] [Google Scholar]
- Popp C, Dean W, Feng S, Cokus SJ, Andrews S, Pellegrini M, et al. Genome-wide erasure of DNA methylation in mouse primordial germ cells is affected by AID deficiency. Nature. 2010;463:1101–1105. doi: 10.1038/nature08829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pozo-Guisado E, Alvarez-Barrientos A, Mulero-Navarro S, Santiago-Josefat B, Fernandez-Salguero PM. The antiproliferative activity of resveratrol results in apoptosis in MCF-7 but not in MDA-MB-231 human breast cancer cells: cell-specific alteration of the cell cycle. Biochem Pharmacol. 2002;64:1375–1386. doi: 10.1016/s0006-2952(02)01296-0. [DOI] [PubMed] [Google Scholar]
- Pozo-Guisado E, Lorenzo-Benayas MJ, Fernandez-Salguero PM. Resveratrol modulates the phosphoinositide 3-kinase pathway through an estrogen receptor alpha-dependent mechanism: relevance in cell proliferation. Int J Cancer. 2004;109:167–173. doi: 10.1002/ijc.11720. [DOI] [PubMed] [Google Scholar]
- Pratt MA, Deonarine D, Teixeira C, Novosad D, Tate BF, Grippo JF. The AF-2 region of the retinoic acid receptor alpha mediates retinoic acid inhibition of estrogen receptor function in breast cancer cells. J Biol Chem. 1996;271:20346–20352. doi: 10.1074/jbc.271.34.20346. [DOI] [PubMed] [Google Scholar]
- Privat M, Aubel C, Arnould S, Communal Y, Ferrara M, Bignon YJ. AKT and p21 WAF1/CIP1 as potential genistein targets in BRCA1-mutant human breast cancer cell lines. Anticancer Res. 2010;30:2049–2054. [PubMed] [Google Scholar]
- Qin W, Zhu W, Shi H, Hewett JE, Ruhlen RL, MacDonald RS, et al. Soy isoflavones have an antiestrogenic effect and alter mammary promoter hypermethylation in healthy premenopausal women. Nutr Cancer. 2009;61:238–244. doi: 10.1080/01635580802404196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rahal OM, Simmen RC. PTEN and p53 cross-regulation induced by soy isoflavone genistein promotes mammary epithelial cell cycle arrest and lobuloalveolar differentiation. Carcinogenesis. 2010;31:1491–1500. doi: 10.1093/carcin/bgq123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rai K, Huggins IJ, James SR, Karpf AR, Jones DA, Cairns BR. DNA demethylation in zebrafish involves the coupling of a deaminase, a glycosylase, and gadd45. Cell. 2008;135:1201–1212. doi: 10.1016/j.cell.2008.11.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rajewsky N. microRNA target predictions in animals. Nat Genet. 2006;38(Suppl.):S8–S13. doi: 10.1038/ng1798. [DOI] [PubMed] [Google Scholar]
- Ramchandani S, Bhattacharya SK, Cervoni N, Szyf M. DNA methylation is a reversible biological signal. Proc Natl Acad Sci U S A. 1999;96:6107–6112. doi: 10.1073/pnas.96.11.6107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rauch TA, Wu X, Zhong X, Riggs AD, Pfeifer GP. A human B cell methylome at 100-base pair resolution. Proc Natl Acad Sci U S A. 2009;106:671–678. doi: 10.1073/pnas.0812399106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Razin A, Riggs AD. DNA methylation and gene function. Science. 1980;210:604–610. doi: 10.1126/science.6254144. [DOI] [PubMed] [Google Scholar]
- Razin A, Szyf M, Kafri T, Roll M, Giloh H, Scarpa S, et al. Replacement of 5-methylcytosine by cytosine: a possible mechanism for transient DNA demethylation during differentiation. Proc Natl Acad Sci U S A. 1986;83:2827–2831. doi: 10.1073/pnas.83.9.2827. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reik W, Dean W. DNA methylation and mammalian epigenetics. Electrophoresis. 2001;22:2838–2843. doi: 10.1002/1522-2683(200108)22:14<2838::AID-ELPS2838>3.0.CO;2-M. [DOI] [PubMed] [Google Scholar]
- Reuter S, Gupta SC, Park B, Goel A, Aggarwal BB. Epigenetic changes induced by curcumin and other natural compounds. Genes Nutr. 2011;6:93–108. doi: 10.1007/s12263-011-0222-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robertson KD, Uzvolgyi E, Liang G, Talmadge C, Sumegi J, Gonzales FA, et al. The human DNA methyltransferases (DNMTs) 1, 3a and 3b: coordinate mRNA expression in normal tissues and overexpression in tumors. Nucleic Acids Res. 1999;27:2291–2298. doi: 10.1093/nar/27.11.2291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosenfeld JA, Wang Z, Schones DE, Zhao K, DeSalle R, Zhang MQ. Determination of enriched histone modifications in non-genic portions of the human genome. BMC Genomics. 2009;10:143. doi: 10.1186/1471-2164-10-143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rowling MJ, McMullen MH, Schalinske KL. Vitamin A and its derivatives induce hepatic glycine N-methyltransferase and hypomethylation of DNA in rats. J Nutr. 2002;132:365–369. doi: 10.1093/jn/132.3.365. [DOI] [PubMed] [Google Scholar]
- Saini S, Majid S, Dahiya R. Diet, microRNAs and prostate cancer. Pharm Res. 2010;27:1014–1026. doi: 10.1007/s11095-010-0086-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanchez-Martinez R, Zambrano A, Castillo AI, Aranda A. Vitamin D-dependent recruitment of corepressors to vitamin D/retinoid X receptor heterodimers. Mol Cell Biol. 2008;28:3817–3829. doi: 10.1128/MCB.01909-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saramaki A, Diermeier S, Kellner R, Laitinen H, Vaisanen S, Carlberg C. Cyclical chromatin looping and transcription factor association on the regulatory regions of the p21 (CDKN1A) gene in response to 1alpha,25-dihydroxyvitamin D3. J Biol Chem. 2009;284:8073–8082. doi: 10.1074/jbc.M808090200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scott JM, Weir DG. Folic acid, homocysteine and one-carbon metabolism: a review of the essential biochemistry. J Cardiovasc Risk. 1998;5:223–227. [PubMed] [Google Scholar]
- Selhub J. Folate, vitamin B12 and vitamin B6 and one carbon metabolism. J Nutr Health Aging. 2002;6:39–42. [PubMed] [Google Scholar]
- Selker EU. Trichostatin A causes selective loss of DNA methylation in Neurospora. Proc Natl Acad Sci U S A. 1998;95:9430–9435. doi: 10.1073/pnas.95.16.9430. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shaikh J, Ankola DD, Beniwal V, Singh D, Kumar MN. Nanoparticle encapsulation improves oral bioavailability of curcumin by at least 9-fold when compared to curcumin administered with piperine as absorption enhancer. Eur J Pharm Sci. 2009;37:223–230. doi: 10.1016/j.ejps.2009.02.019. [DOI] [PubMed] [Google Scholar]
- Sharif J, Muto M, Takebayashi S, Suetake I, Iwamatsu A, Endo TA, et al. The SRA protein Np95 mediates epigenetic inheritance by recruiting Dnmt1 to methylated DNA. Nature. 2007;450:908–912. doi: 10.1038/nature06397. [DOI] [PubMed] [Google Scholar]
- Sharif T, Auger C, Alhosin M, Ebel C, Achour M, Etienne-Selloum N, et al. Red wine polyphenols cause growth inhibition and apoptosis in acute lymphoblastic leukaemia cells by inducing a redox-sensitive up-regulation of p73 and down-regulation of UHRF1. Eur J Cancer. 2010;46:983–994. doi: 10.1016/j.ejca.2009.12.029. [DOI] [PubMed] [Google Scholar]
- Shoba G, Joy D, Joseph T, Majeed M, Rajendran R, Srinivas PS. Influence of piperine on the pharmacokinetics of curcumin in animals and human volunteers. Planta Med. 1998;64:353–356. doi: 10.1055/s-2006-957450. [DOI] [PubMed] [Google Scholar]
- Shrubsole MJ, Shu XO, Li HL, Cai H, Yang G, Gao YT, et al. Dietary B vitamin and methionine intakes and breast cancer risk among Chinese women. Am J Epidemiol. 2011;173:1171–1182. doi: 10.1093/aje/kwq491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shu L, Khor TO, Lee JH, Boyanapalli SS, Huang Y, Wu TY, et al. Epigenetic CpG demethylation of the promoter and reactivation of the expression of Neurog1 by curcumin in prostate LNCaP cells. AAPS J. 2011;13:606–614. doi: 10.1208/s12248-011-9300-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sinclair KD, Allegrucci C, Singh R, Gardner DS, Sebastian S, Bispham J, et al. DNA methylation, insulin resistance, and blood pressure in offspring determined by maternal periconceptional B vitamin and methionine status. Proc Natl Acad Sci U S A. 2007;104:19351–19356. doi: 10.1073/pnas.0707258104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singh SM, Murphy B, O'Reilly RL. Involvement of gene-diet/drug interaction in DNA methylation and its contribution to complex diseases: from cancer to schizophrenia. Clin Genet. 2003;64:451–460. doi: 10.1046/j.1399-0004.2003.00190.x. [DOI] [PubMed] [Google Scholar]
- Slattery ML, Potter JD, Samowitz W, Schaffer D, Leppert M. Methylenetetrahydrofolate reductase, diet, and risk of colon cancer. Cancer Epidemiol Biomarkers Prev. 1999;8:513–518. [PubMed] [Google Scholar]
- Starlard-Davenport A, Tryndyak V, Kosyk O, Ross SR, Rusyn I, Beland FA, et al. Dietary methyl deficiency, microRNA expression and susceptibility to liver carcinogenesis. World Rev Nutr Diet. 2010;101:123–130. doi: 10.1159/000314517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stefanska B, Rudnicka K, Bednarek A, Fabianowska-Majewska K. Hypomethylation and induction of retinoic acid receptor beta 2 by concurrent action of adenosine analogues and natural compounds in breast cancer cells. Eur J Pharmacol. 2010;638:47–53. doi: 10.1016/j.ejphar.2010.04.032. [DOI] [PubMed] [Google Scholar]
- Stefanska B, Huang J, Bhattacharyya B, Suderman M, Hallett M, Han ZG, et al. Definition of the landscape of promoter DNA hypomethylation in liver cancer. Cancer Res. 2011a;71:5891–5903. doi: 10.1158/0008-5472.CAN-10-3823. [DOI] [PubMed] [Google Scholar]
- Stefanska B, Salame P, Bednarek A, Fabianowska-Majewska K. Comparative effects of retinoic acid, vitamin D and resveratrol alone and in combination with adenosine analogues on methylation and expression of phosphatase and tensin homologue tumour suppressor gene in breast cancer cells. Br J Nutr. 2011b;107:781–790. doi: 10.1017/S0007114511003631. [DOI] [PubMed] [Google Scholar]
- Steger DJ, Lefterova MI, Ying L, Stonestrom AJ, Schupp M, Zhuo D, et al. DOT1L/KMT4 recruitment and H3K79 methylation are ubiquitously coupled with gene transcription in mammalian cells. Mol Cell Biol. 2008;28:2825–2839. doi: 10.1128/MCB.02076-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stein R, Razin A, Cedar H. In vitro methylation of the hamster adenine phosphoribosyltransferase gene inhibits its expression in mouse L cells. Proc Natl Acad Sci U S A. 1982;79:3418–3422. doi: 10.1073/pnas.79.11.3418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strahl BD, Allis CD. The language of covalent histone modifications. Nature. 2000;403:41–45. doi: 10.1038/47412. [DOI] [PubMed] [Google Scholar]
- Su Y, Simmen FA, Xiao R, Simmen RC. Expression profiling of rat mammary epithelial cells reveals candidate signaling pathways in dietary protection from mammary tumors. Physiol Genomics. 2007;30:8–16. doi: 10.1152/physiolgenomics.00023.2007. [DOI] [PubMed] [Google Scholar]
- Swami S, Krishnan AV, Feldman D. 1alpha,25-Dihydroxyvitamin D3 down-regulates estrogen receptor abundance and suppresses estrogen actions in MCF-7 human breast cancer cells. Clin Cancer Res. 2000;6:3371–3379. [PubMed] [Google Scholar]
- vel Szic KS, Ndlovu MN, Haegeman G, Vanden Berghe W. Nature or nurture: let food be your epigenetic medicine in chronic inflammatory disorders. Biochem Pharmacol. 2010;80:1816–1832. doi: 10.1016/j.bcp.2010.07.029. [DOI] [PubMed] [Google Scholar]
- Szyf M. DNA methylation and demethylation as targets for anticancer therapy. Biochemistry (Mosc) 2005;70:533–549. doi: 10.1007/s10541-005-0147-7. [DOI] [PubMed] [Google Scholar]
- Szyf M. The early life social environment and DNA methylation. Clin Genet. 2012;81:341–349. doi: 10.1111/j.1399-0004.2012.01843.x. [DOI] [PubMed] [Google Scholar]
- Szyf M, Pakneshan P, Rabbani SA. DNA methylation and breast cancer. Biochem Pharmacol. 2004;68:1187–1197. doi: 10.1016/j.bcp.2004.04.030. [DOI] [PubMed] [Google Scholar]
- Tan S, Wang C, Lu C, Zhao B, Cui Y, Shi X, et al. Quercetin is able to demethylate the p16INK4a gene promoter. Chemotherapy. 2009;55:6–10. doi: 10.1159/000166383. [DOI] [PubMed] [Google Scholar]
- Tessitore L, Davit A, Sarotto I, Caderni G. Resveratrol depresses the growth of colorectal aberrant crypt foci by affecting bax and p21(CIP) expression. Carcinogenesis. 2000;21:1619–1622. [PubMed] [Google Scholar]
- Thaler R, Karlic H, Rust P, Haslberger AG. Epigenetic regulation of human buccal mucosa mitochondrial superoxide dismutase gene expression by diet. Br J Nutr. 2009;101:743–749. doi: 10.1017/S0007114508047685. [DOI] [PubMed] [Google Scholar]
- Tili E, Michaille JJ, Alder H, Volinia S, Delmas D, Latruffe N, et al. Resveratrol modulates the levels of microRNAs targeting genes encoding tumor-suppressors and effectors of TGFbeta signaling pathway in SW480 cells. Biochem Pharmacol. 2010;80:2057–2065. doi: 10.1016/j.bcp.2010.07.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tuddenham L, Wheeler G, Ntounia-Fousara S, Waters J, Hajihosseini MK, Clark I, et al. The cartilage specific microRNA-140 targets histone deacetylase 4 in mouse cells. FEBS Lett. 2006;580:4214–4217. doi: 10.1016/j.febslet.2006.06.080. [DOI] [PubMed] [Google Scholar]
- Unoki M, Nishidate T, Nakamura Y. ICBP90, an E2F-1 target, recruits HDAC1 and binds to methyl-CpG through its SRA domain. Oncogene. 2004;23:7601–7610. doi: 10.1038/sj.onc.1208053. [DOI] [PubMed] [Google Scholar]
- Unoki M, Brunet J, Mousli M. Drug discovery targeting epigenetic codes: the great potential of UHRF1, which links DNA methylation and histone modifications, as a drug target in cancers and toxoplasmosis. Biochem Pharmacol. 2009;78:1279–1288. doi: 10.1016/j.bcp.2009.05.035. [DOI] [PubMed] [Google Scholar]
- Unterberger A, Szyf M, Nathanielsz PW, Cox LA. Organ and gestational age effects of maternal nutrient restriction on global methylation in fetal baboons. J Med Primatol. 2009;38:219–227. doi: 10.1111/j.1600-0684.2008.00320.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vaquero A, Scher M, Lee D, Erdjument-Bromage H, Tempst P, Reinberg D. Human SirT1 interacts with histone H1 and promotes formation of facultative heterochromatin. Mol Cell. 2004;16:93–105. doi: 10.1016/j.molcel.2004.08.031. [DOI] [PubMed] [Google Scholar]
- Vardi A, Bosviel R, Rabiau N, Adjakly M, Satih S, Dechelotte P, et al. Soy phytoestrogens modify DNA methylation of GSTP1, RASSF1A, EPH2 and BRCA1 promoter in prostate cancer cells. In Vivo. 2010;24:393–400. [PubMed] [Google Scholar]
- Vire E, Brenner C, Deplus R, Blanchon L, Fraga M, Didelot C, et al. The Polycomb group protein EZH2 directly controls DNA methylation. Nature. 2006;439:871–874. doi: 10.1038/nature04431. [DOI] [PubMed] [Google Scholar]
- Voegel JJ, Heine MJ, Zechel C, Chambon P, Gronemeyer H. TIF2, a 160 kDa transcriptional mediator for the ligand-dependent activation function AF-2 of nuclear receptors. EMBO J. 1996;15:3667–3675. [PMC free article] [PubMed] [Google Scholar]
- Waite KA, Sinden MR, Eng C. Phytoestrogen exposure elevates PTEN levels. Hum Mol Genet. 2005;14:1457–1463. doi: 10.1093/hmg/ddi155. [DOI] [PubMed] [Google Scholar]
- Walle T, Hsieh F, DeLegge MH, Oatis JE, Jr, Walle UK. High absorption but very low bioavailability of oral resveratrol in humans. Drug Metab Dispos. 2004;32:1377–1382. doi: 10.1124/dmd.104.000885. [DOI] [PubMed] [Google Scholar]
- Wang LS, Arnold M, Huang YW, Sardo C, Seguin C, Martin E, et al. Modulation of genetic and epigenetic biomarkers of colorectal cancer in humans by black raspberries: a phase I pilot study. Clin Cancer Res. 2011;17:598–610. doi: 10.1158/1078-0432.CCR-10-1260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Q, Lee D, Sysounthone V, Chandraratna RAS, Christakos S, Korah R, et al. 1,25-dihydroxyvitamin D3 and retonic acid analogues induce differentiation in breast cancer cells with function- and cell-specific additive effects. Breast Cancer Res Treat. 2001;67:157–168. doi: 10.1023/a:1010643323268. [DOI] [PubMed] [Google Scholar]
- Watanabe T, Tomizawa S, Mitsuya K, Totoki Y, Yamamoto Y, Kuramochi-Miyagawa S, et al. Role for piRNAs and noncoding RNA in de novo DNA methylation of the imprinted mouse Rasgrf1 locus. Science. 2011;332:848–852. doi: 10.1126/science.1203919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Waterland RA. Assessing the effects of high methionine intake on DNA methylation. J Nutr. 2006;136:1706S–1710S. doi: 10.1093/jn/136.6.1706S. [DOI] [PubMed] [Google Scholar]
- Waterland RA, Jirtle RL. Transposable elements: targets for early nutritional effects on epigenetic gene regulation. Mol Cell Biol. 2003;23:5293–5300. doi: 10.1128/MCB.23.15.5293-5300.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weaver IC, Cervoni N, Champagne FA, D'Alessio AC, Sharma S, Seckl JR, et al. Epigenetic programming by maternal behavior. Nat Neurosci. 2004;7:847–854. doi: 10.1038/nn1276. [DOI] [PubMed] [Google Scholar]
- Wei EK, Giovannucci E, Selhub J, Fuchs CS, Hankinson SE, Ma J. Plasma vitamin B6 and the risk of colorectal cancer and adenoma in women. J Natl Cancer Inst. 2005;97:684–692. doi: 10.1093/jnci/dji116. [DOI] [PubMed] [Google Scholar]
- Whitehead N, Reyner F, Lindenbaum J. The Journal of the American Medical Association, Volume 226, 1973: megaloblastic changes in cervical epithelium. Association with oral contraceptive therapy and reversal with folic acid. Nutr Rev. 1989;47:318–321. doi: 10.1111/j.1753-4887.1989.tb02756.x. [DOI] [PubMed] [Google Scholar]
- Whitsett TG, Jr, Lamartiniere CA. Genistein and resveratrol: mammary cancer chemoprevention and mechanisms of action in the rat. Expert Rev Anticancer Ther. 2006;6:1699–1706. doi: 10.1586/14737140.6.12.1699. [DOI] [PubMed] [Google Scholar]
- Widschwendter M, Jones PA. DNA methylation and breast carcinogenesis. Oncogene. 2002;21:5462–5482. doi: 10.1038/sj.onc.1205606. [DOI] [PubMed] [Google Scholar]
- Wiedhopf RM, Young M, Bianchi E, Cole JR. Tumor inhibitory agent from Magnolia grandiflora (Magnoliaceae). I. Parthenolide. J Pharm Sci. 1973;62:345. doi: 10.1002/jps.2600620244. [DOI] [PubMed] [Google Scholar]
- Woynarowski JM, Konopa J. Inhibition of DNA biosynthesis in HeLa cells by cytotoxic and antitumor sesquiterpene lactones. Mol Pharmacol. 1981;19:97–102. [PubMed] [Google Scholar]
- Wu Q, Zhang M, Liu S, Chen Y, Su W. Retinoic acid receptor beta is required for anti-activator protein-1 activity by retinoic acid in gastric cancer cells. Chin Med J (Engl) 2002;115:810–814. [PubMed] [Google Scholar]
- Wyatt GR. Occurrence of 5-methylcytosine in nucleic acids. Nature. 1950;166:237–238. doi: 10.1038/166237b0. [DOI] [PubMed] [Google Scholar]
- Yang CS, Fang M, Lambert JD, Yan P, Huang TH. Reversal of hypermethylation and reactivation of genes by dietary polyphenolic compounds. Nutr Rev. 2008;66(Suppl. 1):S18–S20. doi: 10.1111/j.1753-4887.2008.00059.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang XJ, Seto E. HATs and HDACs: from structure, function and regulation to novel strategies for therapy and prevention. Oncogene. 2007;26:5310–5318. doi: 10.1038/sj.onc.1210599. [DOI] [PubMed] [Google Scholar]
- Youssef EM, Lotan D, Issa JP, Wakasa K, Fan YH, Mao L, et al. Hypermethylation of the retinoic acid receptor-beta(2) gene in head and neck carcinogenesis. Clin Cancer Res. 2004;10:1733–1742. doi: 10.1158/1078-0432.ccr-0989-3. [DOI] [PubMed] [Google Scholar]
- Yu Z, Li W, Lu Q, Wang L, Zhang X, Han P, et al. p21 is required for atRA-mediated growth inhibition of MEPM cells, which involves RAR. J Cell Biochem. 2008;104:2185–2192. doi: 10.1002/jcb.21773. [DOI] [PubMed] [Google Scholar]
- Zapisek WF, Cronin GM, Lyn-Cook BD, Poirier LA. The onset of oncogene hypomethylation in the livers of rats fed methyl-deficient, amino acid-defined diets. Carcinogenesis. 1992;13:1869–1872. doi: 10.1093/carcin/13.10.1869. [DOI] [PubMed] [Google Scholar]
- Zhang D, Holmes WF, Wu S, Soprano DR, Soprano KJ. Retinoids and ovarian cancer. J Cell Physiol. 2000;185:1–20. doi: 10.1002/1097-4652(200010)185:1<1::AID-JCP1>3.0.CO;2-O. [DOI] [PubMed] [Google Scholar]
- Zhang T, Kraus WL. SIRT1-dependent regulation of chromatin and transcription: linking NAD(+) metabolism and signaling to the control of cellular functions. Biochim Biophys Acta. 2010;1804:1666–1675. doi: 10.1016/j.bbapap.2009.10.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou H, Hu H, Lai M. Non-coding RNAs and their epigenetic regulatory mechanisms. Biol Cell. 2010;102:645–655. doi: 10.1042/BC20100029. [DOI] [PubMed] [Google Scholar]
- Zhou JR, Gugger ET, Tanaka T, Guo Y, Blackburn GL, Clinton SK. Soybean phytochemicals inhibit the growth of transplantable human prostate carcinoma and tumor angiogenesis in mice. J Nutr. 1999;129:1628–1635. doi: 10.1093/jn/129.9.1628. [DOI] [PubMed] [Google Scholar]