

Abstract
Among a myriad of spider web geometries, the orb web presents a fascinating, exquisite example in architecture and evolution. Orb webs can be divided into two categories according to the capture silk used in construction: cribellate orb webs (composed of pseudoflagelliform silk) coated with dry cribellate threads and ecribellate orb webs (composed of flagelliform silk fibres) coated by adhesive glue droplets. Cribellate capture silk is generally stronger but less-extensible than viscid capture silk, and a body of phylogenic evidence suggests that cribellate capture silk is more closely related to the ancestral form of capture spiral silk. Here, we use a coarse-grained web model to investigate how the mechanical properties of spiral capture silk affect the behaviour of the whole web, illustrating that more elastic capture spiral silk yields a decrease in web system energy absorption, suggesting that the function of the capture spiral shifted from prey capture to other structural roles. Additionally, we observe that in webs with more extensible capture silk, the effect of thread strength on web performance is reduced, indicating that thread elasticity is a dominant driving factor in web diversification.
Keywords: spider web mechanics, evolution, viscid capture silk, cribellate capture silk, Deinopoidea, Araneoidea
1. Introduction
The great diversity of spider silks found in nature has fascinated researchers for years [1]. Spider silk's remarkable properties of great strength and extensibility have inspired significant research into its structure and function [2–7]. From the plethora of spider species, orb weavers have not surprisingly captured significant attention, as a result of a highly functional yet beautiful architecture of the orb in combination with superior material properties of its silk.
Today's orb-weaving spiders belong to the Orbicularian clade [8], which is composed of seven different families [9]. Two distinct lineages characterize the Orbicularian clade: the Deinopoidea super family (which includes cribellate orb weavers) and the Araneoidea super family (which contains the ecribellate orb weavers). The appearance of orb webs in the early Triassic period, over 200 Ma, correlated with the evolution of strong major ampullate (MA) silk, which is used to construct radially extending threads in a typical orb web [10–14]. Along with MA silk, cribellate silk first appeared at this time. Cribellate silk was a major innovation and can be found today in all araneomorph spiders. By the time orb webs were abundant, in the late Jurassic period and possibly earlier, ecribellate orb weavers split from cribellate orb weavers, as viscid capture silk replaced cribellate capture silk [8,15,16]. Because out-groups of the orb-weaving spiders use cribellate capture silk, there is strong phylogenic evidence to suggest that cribellate capture silk is more closely related to the ancestral form of capture silk [17,18]. Over time, the Araneoidea super family extended and became much more diverse than the Deinopoidea super family, implying a possible evolutionary advantage for araneoid orb weavers. In fact, today 95 per cent of all orb-weaving spiders use viscid capture silk and belong to the Araneoidea super family [19].
The capture silk used by orb weavers within the two super families varies significantly, and has been empirically shown to have drastically different material properties [20]. Deinopoids use rigid setae (known collectively as the calamistrum), to draw out dry, fuzzy cribellate silk from the cribellum—a flat spinning plate—to construct the spiral, retaining prey by trapping it with sticky cribellate microfibrils that cover the anchoring thread fibres [21–23]. Viscid capture threads, by contrast, are composed of flagelliform silk, drawn out of two flagelliform glands and covered by sticky aqueous glue droplets secreted from two aggregate glands [24–27]. Although the radial threads in both families are made of strong MA silk and exhibit similar mechanical properties [28], the production mechanism, composition and mechanical properties of the capture spirals between the two super families exhibit stark differences.
The subject of spider evolution and the diversification of orb weavers, in particular, has been a central focus in a variety of studies that addressed driving factors ranging from spider silk composition and variation in web architecture, to physical and behavioural features of spiders. Specifically, a number of studies have discussed various advantageous aspects of viscid capture silk over cribellate capture silk. Opell hypothesized that the economical sticky capture silk produced by araneoids maximizes stickiness per capture area and is more economical with regard to material usage, yielding an evolutionary advantage on the basis of efficiency and economy [19,29,30]. Studies comparing trends in spider weight, orb web architecture and capture area concluded that araneoids were better equipped to dissipate the kinetic energy of faster, larger prey [31]. Blackledge et al. [8] compared the spiral silk spinning process, contending that a simpler process in araneoids contributed to their diversity. To produce cribellate threads, a spider spins a core thread, which is followed by a long, arduous process of adding a layer of microfibres, giving cribellate silk its properties [17]. On the other hand, to produce viscid capture silk the spider spins flagelliform threads and covers them with aqueous glue as is the web is spun, speeding up the spinning process and minimizing energy costs. Similarly, Swanson et al. [32] concede that spinning morphology and variation in silk sequence are important evolutionary factors. In addition to a shift in spinning mechanism, Blackledge et al. [8] find a divergence from substrate-bound webs, arguing for behavioural implications to orb web evolution. Similarly, Vollrath et al. [16] suggested that behavioural patterns determining web orientation and construction, as well as prey-sensing and prey-trapping mechanisms served as important evolutionary driving forces.
A particularly striking difference between cribellate capture silk and viscid capture silk is the shift in mechanical properties, which was recently characterized by a study by Swanson et al. [20]—the most extensive study on this topic to date, shown in figure 1. Capture silk has a key operational role in orb webs, particularly for prey interception and prey retention [29,33,34]. It is also the most energetically costly component for spiders in both families [35]. The apparent trade-off in strength and extensibility of capture silks between the two super families [20,36] indicates that increased extensibility and reduced strength may be evolutionary advantageous. Additionally, it has been suggested that the vertical orientation of araneoid orb webs, compared with a more commonly found horizontal orientation of deinopoid orb webs, results in an ability to withstand greater impact forces by faster flying prey [31,33,37]. This predicts effectively that the energy-absorption capacity of webs in the Araneoidea super family must be superior.
Figure 1.
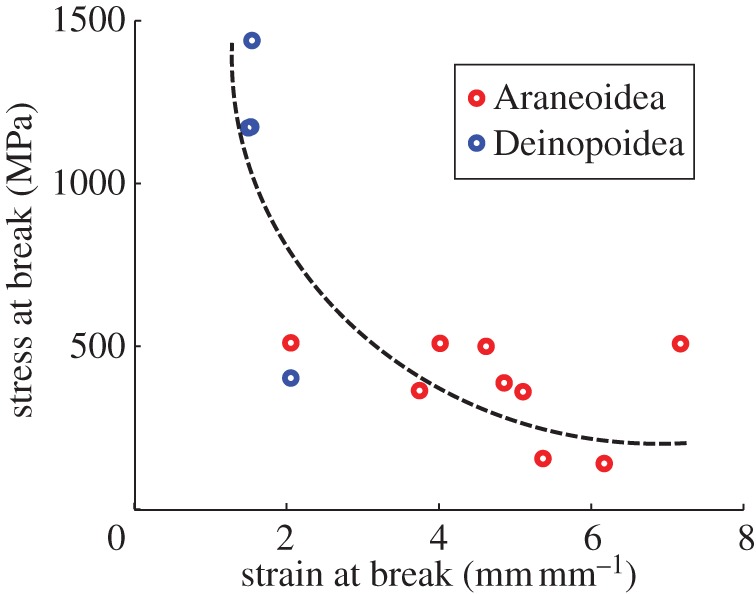
A wide diversity of material properties are exhibited by capture silk threads in orb webs. Stress and strain of spiral capture threads at break were quantified in empirical tensile tests for 13 spider species, adapted from Swanson et al. [20]. Capture silks in the Deinopoidea super family tend to have higher ultimate stresses and lower ultimate strains than capture silks found in the Araneoidea super family. This material ‘trade-off’ correlates with an apparent evolutionary advantage attributed to the more numerous araneoids. (Online version in colour.)
Spinning silk and creating the web is the single most dominant expense in the energy budget of spiders [38]. First of all, there are metabolic costs involved with synthesizing the amino acids in silk. The protein components also require a caloric energy expenditure. As the web is spun, there are costs associated with the spider's movement to create the orb [39]. These energetic expenses need to be counterbalanced by an effective prey-capture strategy. Such a biological trade-off strongly suggests that reducing metabolic, material and behavioural costs of building the web should play a large part in determining the evolution of the orb and its component parts. It is likely, then, that the transition from cribellate capture spiral silk to viscid spiral silk may largely be accounted for by an energetic advantage in using the latter.
Few studies have explicitly considered the relationship between material and structural variation across the spectrum of spiral silk and the connection to energy-absorptive capacity of orb webs. Kohler et al. [40] compared the mechanical properties of the threads, proposing a different mechanistic approach of energy dissipation. It was observed that energy was dissipated by araneoid webs through absorption by extensibility, while deinopoids dissipate energy by the friction of the small cribellate microfibrils. Craig [33] used web models to study the relationship of energy absorption to various architectural features, including radial prestress, the number of radials and spirals, and the angle of the web with respect to loading; however, she did not explicitly consider the effect of silk material properties on energy absorption. Additionally, Craig measured the energy-absorption capabilities of five real araneoid webs, on the basis of artificial loading experiments by calculating the kinetic energy of prey caught by each web. She concluded that web geometry is a key factor for high energy-absorbing webs, while properties of individual threads determine energy dissipation in small, low energy-absorbing webs. She suggested that because the latter was more evolutionary advantageous, the implication is that individual silk properties played an increasingly important role in the development of webs. Lin et al. [41] considered a finite-element model of a web and found that aerodynamic damping plays a crucial role in energy dissipation during prey capture, however only for one species of spider. Sensenig et al. [42] studied 22 species of spiders and focused on the connection between silk quality and web geometry, demonstrating that larger spiders spin higher quality silk, using improved material properties and ‘sparser’ web architectures.
The consequences on web mechanics due to the apparent shift from stronger, less-extensible threads (such as those found in deinopoids) to weaker, more elastic threads in araneoids have never been explicitly studied, though several authors have proposed that the role of a material transition is the key in the evolution of orb-weaving spiders [17,43]. This variability is the product of a complex process, where ecological, behavioural and genetic factors play a significant role. Admittedly, these driving factors are numerous and often subtly interconnected, and conclusions drawn after complex evolutionary mechanisms have taken place may not be definitive. Yet, by focusing on specific patterns, we can address individual factors that may account for particular trends in order to formulate a wholesome understanding of evolutionary processes driving species diversification.
Here, we test the hypothesis that the observed shift in material behaviour of spiral silk is responsible for araneoids' ability to absorb and dissipate energy better than deinopoids [10,14,31,33]. Essentially, we use an inverse approach to understanding evolution, considering products of the evolutionary process within a web model to understand what mechanistic advantages ground the process itself.
2. Methods
We use a computational approach to probe the effect of varying capture silk material behaviour on the performance of the web, by systematically exploring material property trends representative of natural silk. The model permits controlled variation of capture silk properties to evaluate their effect on the response of the web, upon loading. Web structure, size and loading conditions are controlled while a systematic variation of capture thread material properties is applied based on observed empirical data [20]. Rather than predicting patterns and relationships grounded on observations of natural systems, we directly incorporate evolutionary products into the model.
2.1. Web model
We use an orb web model composed of eight radially extending threads responsible for structural support and a spiral thread involved primarily in prey retention (figure 2a). Threads are represented by bead-spring elements with an equilibrium spacing of 0.01 m, mimicking a realistic size. The orb is approximated by an arithmetic spiral supported by radial threads at regular angular intervals. Connection points contain additional links to simulate the properties of the glue-like cement.
Figure 2.
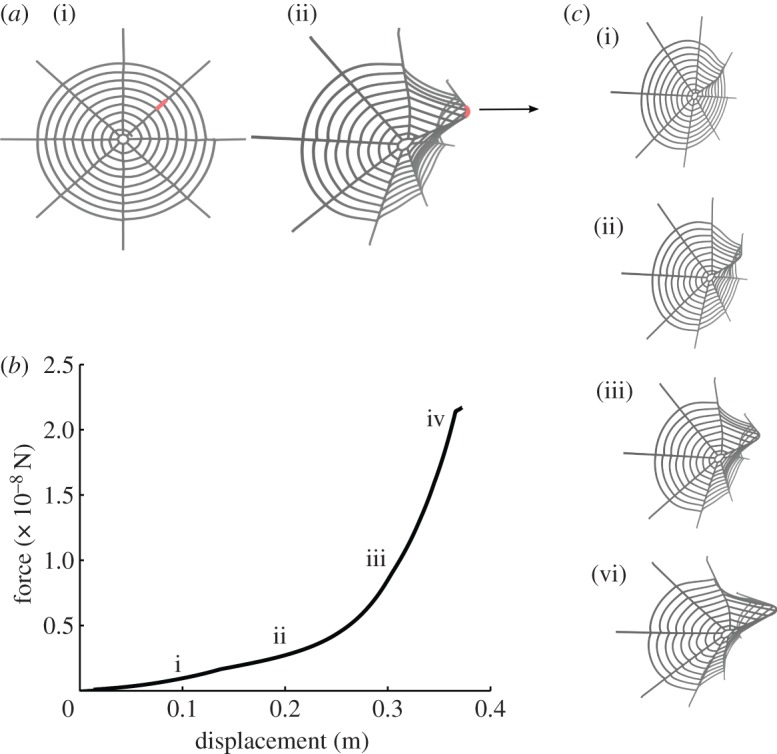
Model development and application. (a) A spider web model exhibiting orb web geometry with eight radially extending threads responsible for structural support and a spiral thread involved primarily in prey retention is developed. This geometry is abundantly found in spiders of both Deinopoidea and Araneoidea super families (i). Constant-velocity force increments are applied to a radial thread segment (shown in red) to study web deformation behaviour for various capture silk material properties (ii). (b) Force–displacement data for a web pulled in the out-of-plane direction until failure. Numerals (i–iv) correspond to web deformation states as shown in (c).
Although many factors contribute to the mechanical behaviour of the web, in this study we are primarily concerned with the effect of varying material behaviour of spiral threads. To measure this effect, we compare a series of material laws describing capture spiral behaviour in the web, modelled after naturally observed spiral threads to reflect both the nonlinear elastic behaviour and ultimate values of strength and extensibility (figure 3a). The approximate material behaviour of the threads as well as the range of maximum strength and extensibility values considered is modelled after Swanson et al. [20]. The total energy of the web system is defined according to a particle dynamics formulation as the sum of the elastic potentials of all threads, where ϕmaterial refers to the constitutive energy expression of the specific silk material:
| 2.1 |
Figure 3.
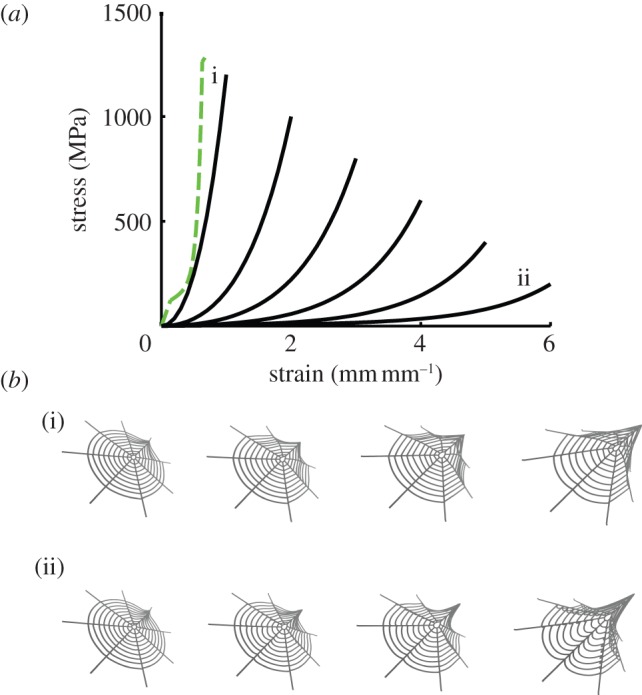
Material law variation and impact on web mechanics. (a) Six material laws were used to model capture spiral threads in the web in order to characterize the effect of extensibility-for-strength exchange observed in spider capture silk (shown in black). Radial silk material behaviour exhibited little variation across species and was consequently kept consistent across the models. The dotted green line represents the atomistically derived material behaviour of dragline silk radial threads [44]. The spread of maximum stress and strain values for capture silk is based on the observed silk material variation across spider species (figure 1). An exponential material law was fit to tensile experiments [20], and kept consistent for all models. The web was parametrized accordingly for the six different spiral thread material laws, and deformation was applied as shown in figure 2a. (b) Snapshots of the web corresponding to two extreme cases (labelled i, ii in (a)).
The material behaviour of the threads is implemented by a combination of atomistically derived and empirically parametrized models. Because radial thread behaviour shows little variation across species when compared with the material variation of spiral threads [20,28], a single material law is used for radial threads across the models. It is parametrized from full atomistic simulations of MA dragline spider silk (figure 3a) [45–49], defined by the constitutive behaviour:
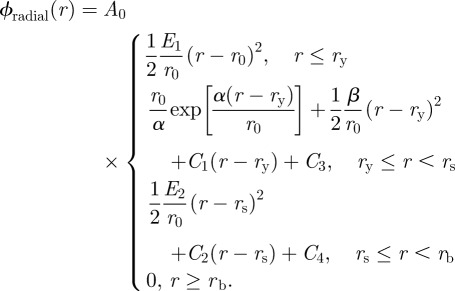 |
2.2 |
All parameters are consistent with our earlier model reported in Cranford et al. [44]. Model parameters and experimentally derived geometric thread parameters are listed in table 1. Six material laws are considered for capture spiral threads in the web model in order to characterize the effect of extensibility-for-strength exchange observed in capture silk across different species of spiders (figure 3a) [20]. An exponential material law is fit to tensile experiments [20], and kept consistent across the models to controllably observe the effect of strength and extensibility variation. The material law for spiral capture silk is described by
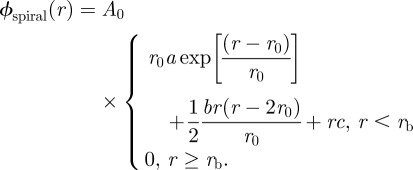 |
2.3 |
Table 1.
Atomistically derived dragline silk potential parameters and physical dimensions of radial and spiral thread diameters, taken from experimental findings [50,51].
| parameter | value |
|---|---|
| initial stiffness, E1 | 875.9 MPa |
| exponential parameter, α | 14.2 |
| tangent stiffness parameter, β | 180 MPa |
| final stiffness, E2 | 491.2 MPa |
| yield strain, ɛy | 0.1356 |
| softening strain, ɛs | 0.6322 |
| ultimate (breaking) strain, ɛb | 0.6725 |
| radial thread diameter (from earlier studies [50,51]) | 3.93 μm |
| spiral thread diameter (from earlier studies [50,51]) | 2.40 μm |
The spread of ultimate strength and strain values, (σf, ɛf), for capture silk is based on observed silk material behaviour variation across a series of spider species (figure 1) [20]. Critical values for the six material laws are listed in table 2. The modulus at small deformation, E1, is assumed to be 3 MPa for capture silk [52]. The ultimate strength and strain values of the silks define the constants in equation (2.3) as
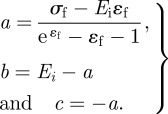 |
2.4 |
Table 2.
Ultimate strength and strain parameters for the material laws depicted for six spiral silk models in figure 3a.
| model | ultimate tensile strength, σf (MPa) | ultimate tensile strain, ɛf (mm mm−1) |
|---|---|---|
| 1 | 1200 | 1 |
| 2 | 1000 | 2 |
| 3 | 800 | 3 |
| 4 | 600 | 4 |
| 5 | 400 | 5 |
| 6 | 200 | 6 |
2.2. Loading conditions
Mechanical loading is applied to web models as illustrated in figure 2a (loaded threads shown in red). A constant-velocity spring load (a spring constant of 0.007 N m−1 and a constant velocity of 0.002 m s−1, resulting in a constant applied force of 0.000014 N s−1) is applied to a segment of thread in the out-of-plane direction, incrementally until failure. This loading scenario is meant to mimic the force experienced by the web upon impact of prey applied primarily at a right angle [1]. Out-of-plane deflection and force are calculated. A sample plot is presented in figure 2b. Corresponding web deformation states are shown in figure 2c, depicting a sample dataset to show how energy absorption is quantified.
Web deformation and failure mechanics are quantified in the models by comparing the energy-absorption capacity of the web. It has been suggested that among various strategies of dissipating energy, specifically among internal energy dissipation of radial and spiral threads and aerodynamic damping, radials play the most significant role in prey capture [53]. Nevertheless, understanding how material behaviour of the spirals affects energy dissipation can elucidate the functional role spirals play in the web. In our study, we define web performance by the amount of energy it can absorb. This energy is quantified as the area under the force–displacement curve of the web, shown in figure 2b, as it is stretched by a spring-load, until failure, defined as the breaking point of the loaded threads. The kinetic energy of the prey can either be stored through elastic deformation of spiral and radial threads or it could be dissipated in the form of heat through friction as the web stretches [41,52]. So, more accurately, the energy we calculate includes both the energy absorbed by elastic deformation and the energy dissipated through friction. It has been shown in sequential loading and unloading experiments of real silk fibres that hysteresis in silk threads is about 65 per cent, indicating that 65 per cent of the kinetic energy is dissipated in the form of heat, upon impact [1,52]. The area ‘lost’ to hysteresis corresponds to the energy dissipated through friction. So, when we refer to energy-absorption capacity, we are in fact discussing the total energy associated with deformation, which is a combination of energy dissipated in the form of heat and the energy absorbed through elastic stretching. Friction is simulated by a damping force introduced in the model while elastic energy dissipated is governed by the formulation of the material behaviour of the threads.
3. Results and discussion
To capture the variation in energy-absorption capabilities of a web due exclusively to fluctuation in capture silk material behaviour, six material laws were considered (shown in figure 3a) representing the observed trend in capture thread material [20]. Each model was identically deformed to track the force–displacement behaviour and the energy absorbed. Four representative loading scenarios were studied, yielding similar results (see the electronic supplementary material, figure S1). Loading regions surrounding both spiral and radial threads were considered, as well as regions close to and far away from the hub. Results for the loading scenario shown in figure 2c are presented in figure 4. Energy absorbed by the web and toughness of individual capture silk fibres is shown in figure 4a. Toughness of thread fibres was calculated as the area under the stress–strain curve used to parameterize individual threads in the web models. A peak in thread toughness is observed at a balance of strength and extensibility, where (σf, ɛf) = (800 MPa, 3 mm mm−1). Contrary to expectation, energy absorption of the web steadily decreases as extensibility replaces strength in the fibres of the capture silk, even as toughness of the individual fibres peaks. The maximum force and displacement of the web prior to breaking is captured in figure 4b,c for the corresponding material law models from figure 3a. Three additional loading scenarios are considered, and the web is found to behave in a similar manner (see the electronic supplementary material, figure S1). The force to break the web decreases with decreased capture thread strength, while maximum extension of the web increases with increased capture thread extensibility, as expected. Energy absorbed by the web is determined both by the maximum force required to break the web and by the ultimate extension that the web can reach before breaking. We attribute the surprising finding that energy-absorption capacity decreases with increasing capture thread extensibility, even as thread toughness peaks, to the fact that the force required to break the web must be the dominating factor determining web performance.
Figure 4.
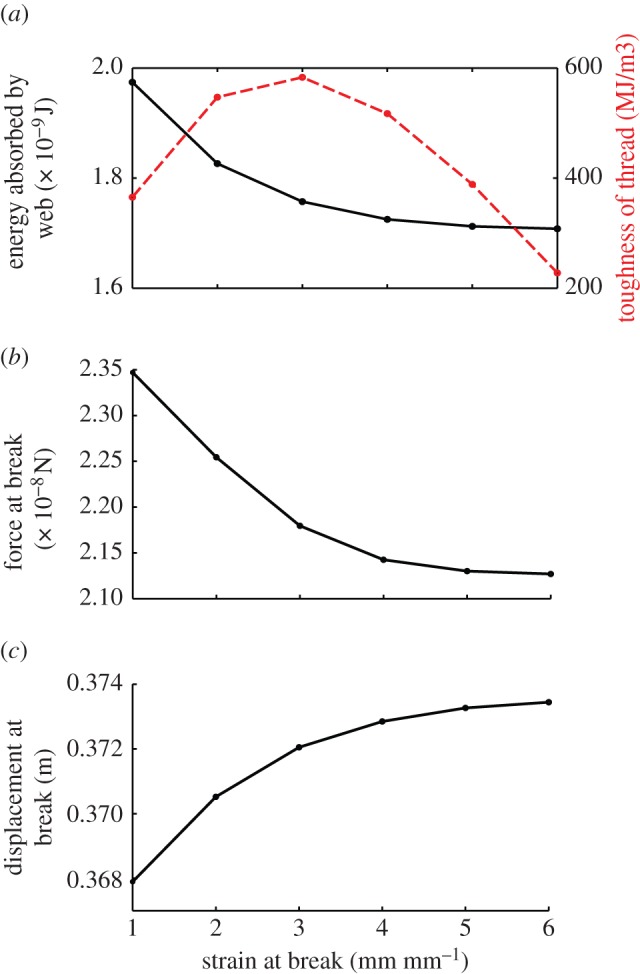
Energy absorption, toughness and other mechanical properties of the web. (a) Energy absorbed by the web (defined as the area under the force–displacement curve) and the corresponding capture thread toughness (defined as the area under the stress–strain curve for a single spiral thread), (b) force of the web at break, and (c) displacement of the web at break compared for six material laws shown in figure 3a. (Online version in colour.)
We conclude that the mechanism of deformation that appears in webs made of viscid capture silk, rather than the overall ability of these webs to absorb energy, is more significant to the observed success of viscid capture silk. The difference in energy-dissipation mechanisms between the two spider super families has been observed previously by Kohler et al. [40]. In this study, we find that the mechanism of web deformation may be more relevant to spider diversification than the ability of the web to absorb more energy upon deformation. Web deformation is captured explicitly for two representative web models in figure 3a, comparing two limiting cases: (i) exceptionally strong spiral threads exhibiting minimal extension and (ii) weak, yet extremely elastic spiral threads. Snapshots of the web under deformation are shown in figure 3b. For the stronger, less-extensible spiral threads (i), the web extends as an umbrella, where spiral threads act cooperatively with radials, causing a more global deformation as a result of a similar material law defining radial and spiral thread mechanics. Weaker, more extensible spirals (ii) have a greater effect on the web's deformation mechanism, as they are more pronouncedly engaged in full-structure deformation, resulting in localization. This achieves greater web extensibility; however, because deformation is governed by the weaker spirals, the total energy-absorption capacity is reduced. The wide diversification of araneoid spiders implies that even with a reduced capability to absorb and dissipate energy, the mechanism of web deformation through extension is advantageous in prey capture.
Our rather surprising observation that viscid capture silk in the web results in lower energy dissipation also implies that capture threads in araneoids play a weaker prey-catching function, as has been suggested in a recent study by Sensenig et al. [53]. We propose that the mechanism of prey capture in the orb is more important from the evolutionary standpoint of capture silk than the web's total dissipative capacity.
In an earlier study, we compared the mechanical response of a loaded web composed of threads with varying material laws [44]. In particular, we found that a web composed of natural silk with a characteristic nonlinear material law was more robust and damage tolerant than a web made of threads with material laws mimicking standard engineering materials with a stiffer initial regime upon deformation. Similarly, the more elastic viscid silk compared favourably against stiffer cribellate silk. One reason for this may be the minimization of damage to the web in the case of mechanical failure. If the web breaks, we observe that damage is much more localized; so it is more economical for the spider to rebuild. Hence, the reduced energy-dissipation capacity observed for weaker capture threads may be counterbalanced by the conservation of metabolic energy used to restore damaged regions. Effectively, the capture threads adopt a new function in the web: they optimize the spider's energy primarily by improving the web's structural mechanics rather than through prey capture.
This observation may also shed light on the role of capture silk in prey retention after it has been intercepted by the web. In webs made of viscid capture silk, prey is retained by sticking to the aqueous glue droplets coating the threads, which cling to the prey as it struggles to escape. Previous studies have discussed the importance of web geometry and close alignment of spiral threads for engulfing the struggling prey as it adheres to adjacent threads [9]. The increased elasticity of capture thread plays an important role in dissipating kinetic energy of the prey upon recoil. We propose that in addition to serving a dissipative function as the prey oscillates in the web, more elastic capture silk keeps the prey in a contained region of the web, forcing it to attach to adjacent threads more efficiently.
To systematically compare the effect of capture silk material variation on web mechanics, 24 material laws were studied, spanning the full spectrum of experimentally observed capture silk material laws (figure 5a,c). Again, four loading scenarios were considered, illustrated in the electronic supplementary material, figure S1. Results for the representative loading scenario in figure 2 is shown in figure 5. The effect of increasing thread extensibility for a constant thread strength (figure 5a) as well as the effect of varying strength at a constant extensibility (figure 5c) was evaluated. The effect of increasing thread extensibility for a given strength value is comparable to the effect of increasing extensibility and simultaneously decreasing strength, as in figure 3a. Here too, energy absorption decreases with increasing thread extensibility, because deformation becomes primarily governed by the weaker spiral threads. A similar loading scenario, but closer to the hub of the web, yielded a similar result (see the electronic supplementary material, figure S2a). Spiral thread loading was considered for comparison, but we found that, as a result of the highly variant mechanical properties between spiral and radial threads, loading of spirals resulted in very localized failure of the loaded thread, and the web's energy absorption was therefore governed by the mechanical properties of the spiral threads alone (see the electronic supplementary material, figure S3).
Figure 5.

The effect on web performance due to variation in capture silk properties. (a) Summary of 24 material laws used for parametrizing capture silk in web models. Material laws are colour-coded, showing a trend of increasing extensibility for a given thread strength. (b) Energy absorbed by the web under loading conditions shown in figure 2a is recorded for each model. (c) The same 24 material laws are shown, coloured to show a trend of increasing strength for a given thread extensibility. (d) The effect on energy absorption by the web when increasing the strength of the material while keeping extensibility constant for six different breaking strains shown in (c). Baseline energy absorption is defined as the energy absorbed for breaking stress of 600 MPa for a set of material laws at a given breaking strain.
When extensibility is held constant for a set of strength values, as in figure 5c, an interesting trend is observed. As threads become more extensible, the ultimate strength of the thread has an increasingly reduced effect on web performance, quantified by the web's energy-absorption capacity. For instance, the energy absorbed in webs with (σf, ɛf) = (600 MPa, 6 mm mm−1) and (1200 MPa, 6 mm mm−1) is approximately equal, while the difference is much more pronounced between webs with (σf, ɛf) = (600 MPa, 1 mm mm−1) and (1200 MPa, 1 mm mm−1; figure 5b). Increase from baseline energy, defined as the energy absorption for breaking strength of 600 MPa, is explicitly shown in figure 5d. The least significant increase is observed for the most extensible threads. Loading closer to the hub results in a similar trend (see the electronic supplementary material, figure S2b). This gives a direct mechanistic insight into the material ‘trade-off’ observed in the evolution of webs: more extensible threads do not ‘need’ strength to provide a particular function in a full web structure, as deformation is governed by extensibility. For a large thread extensibility, strength plays an insignificant role in improving energy-dissipation capacity. Therefore, more elastic, weaker threads can achieve the same effect and thereby reduce associated production costs for the spider. So, to the extent that spiral thread contributes to prey capture, the spider optimizes cost effectiveness by making weaker threads that achieve the same functional role.
These findings suggest that the key for the observed evolutionary trade-off between strength and extensibility of spirals is not primarily a result of improved energy-absorption capacity in more extensible webs. Rather, it is a combination of at least two factors. First of all, more extensible, weaker threads, representative of viscid capture silk, result in increased and highly localized deformation of the web structure. This method of energy dissipation—through elastic deformation—functions in arresting quick-flying prey. It seems that the evolution of viscid capture silk is tied to a different mechanism of dissipating energy rather than to a greater total energy dissipation per se. This may be advantages from a structural point of view (as a source of minimizing damage, should it occur), and from the prey-capture perspective (as web extension localizes prey upon impact, making capture easier). Secondly, as threads become more extensible, less strength is needed to achieve the same effective energy-dissipation response in the web—an additional metabolic advantage with regard to energy conservation.
We note that because our study considers variation in spiral threads only, it may be offset in natural systems by parallel changes in radial thread diversification, although variation in radial threads is generally much less significant than in spirals [28]. However, our goal is to understand the mechanistic effect on the web's response resulting from variations in spiral thread material behaviour alone, as this trend is most striking and largely unexplored across species. We show that the observed trade-off is not only behaviourally and energetically advantageous for the spider, but mechanically optimized. As the thread becomes more extensible, it needs less strength to absorb equivalent amounts of energy. In other words, the ultimate strength of spiral threads has a reduced effect on the behaviour of the web system as threads become more elastic.
Various complementary studies have tried to explain the success of viscid capture silk and its role in the evolution of the orb web [8,16,17,19,29,32,43]. Now, we can add to the list the mechanical benefits of the observed trade-off between strength and extensibility, demonstrative of the shift from cribellate to viscid capture silk: viscid capture silk is beneficial for minimizing energetic costs associated with orb creation, helps to reduce damage under extreme loads and retains prey by localizing it in the web.
4. Conclusions
The two primary findings of this study are as follows. First, by systematically varying the material behaviour of capture silk within the web model, we find that the governing evolutionary principle from the mechanical standpoint is the mechanism of deformation rather than an increased energy-absorption capacity. We attribute the loss in energy-absorption capacity to the effective deformation mechanism dominated by weaker spiral threads. We explain this shift in the light of an increased damage tolerance and better prey-retention capability. The second contribution of this study is the finding that more extensible spiral threads require minimal strength for optimized effect. Extensibility dominates over strength in capture silk, in defining web performance, as the effect of strength is progressively reduced when extensibility is increased. These results suggest that the effective prey-capture mechanism is tied to a ‘less-expensive’ metabolic process for making silk, allowing the spider to conserve energy by producing weaker but equally functional silk.
The cooperative effect of threads in the web plays a major role in determining web function and needs to be considered when evaluating evolutionary trends in material behaviour of individual fibres. This approach may be instrumental in understanding the evolution of other hierarchical structures. The use of modelling may be applied more broadly to understand evolutionary shifts by simulating modifications not only in material behaviour, but also in a variety of other factors, such as environmental conditions or system structure. Furthermore, these findings suggest that the material shift naturally derived through evolutionary mechanisms in spider silk may be used more generally in engineered heterogeneous materials requiring conservation of phases contributing to material strength. We propose that natural structures provide an excellent material template for biomimetic, synthetic materials, and the evolutionary process provides an insight into smarter modification of existing materials.
Acknowledgements
This work was supported by the Office of Naval Research (PECASE, grant no. N000141010562), the National Science Foundation (CAREER, grant no. CMMI-0642545) and by a Robert A. Brown Presidential Fellowship. All simulations have been carried out at MIT's Laboratory for Atomistic and Molecular Mechanics (LAMM). We thank the reviewers for their helpful comments and suggestions.
References
- 1.Denny M. 1976. Physical properties of spiders silk and their role in design of orb-webs. J. Exp. Biol. 65, 483–506 [Google Scholar]
- 2.Agnarsson I., Kunter M., Blackledge T. A. 2010. Bioprospecting finds the toughest biological material: extraordinary silk from a giant riverine orb spider. PLoS ONE 5, e11234 10.1371/journal.pone.0011234 (doi:10.1371/journal.pone.0011234) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Heim M., Romer L., Scheibel T. 2010. Hierarchical structures made of proteins. The complex architecture of spider webs and their constituent silk proteins. Chem. Soc. Rev. 39, 156–164 10.1039/b813273a (doi:10.1039/b813273a) [DOI] [PubMed] [Google Scholar]
- 4.Omenetto F. G., Kaplan D. L. 2010. New opportunities for an ancient material. Science 329, 528–531 10.1126/science.1188936 (doi:10.1126/science.1188936) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Vepari C., Kaplan D. L. 2007. Silk as a biomaterial. Prog. Polym. Sci. 32, 991–1007 10.1016/j.progpolymsci.2007.05.013 (doi:10.1016/j.progpolymsci.2007.05.013) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Vollrath F. 1999. Biology of spider silk. Int. J. Biol. Macromol. 24, 81–88 10.1016/S0141-8130(98)00076-2 (doi:10.1016/S0141-8130(98)00076-2) [DOI] [PubMed] [Google Scholar]
- 7.Tarakanova A., Buehler M. J. 2012. A materiomics approach to spider silk: protein molecules to webs. JOM 64, 214–225 10.1007/s11837-012-0250-3 (doi:10.1007/s11837-012-0250-3) [DOI] [Google Scholar]
- 8.Blackledge T. A., Scharff N., Coddington J. A., Szuts T., Wenzel J. W., Hayashi C. Y., Agnarsson I. 2009. Reconstructing web evolution and spider diversification in the molecular era. Proc. Natl Acad. Sci. USA 106, 5229–5234 10.1073/pnas.0901377106 (doi:10.1073/pnas.0901377106) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Blackledge T. A., Kuntner M., Agnarsson I. 2011. The form and function of spider orb webs: evolution from silk to ecosystems. Adv. Insect Physiol. 41, 175–262 10.1016/B978-0-12-415919-8.00004-5 (doi:10.1016/B978-0-12-415919-8.00004-5) [DOI] [Google Scholar]
- 10.Craig C., Brunetta L. 2010. Spider silk: evolution and 400 million years of spinning, waiting, snagging, and mating, 1st edn. New Haven, CT: Yale University Press [Google Scholar]
- 11.Zschokke S. 2003. Spider-web silk from the Early Cretaceous. Nature 424, 636–637 10.1038/424636a (doi:10.1038/424636a) [DOI] [PubMed] [Google Scholar]
- 12.Penney D., Ortuno V. M. 2006. Oldest true orb-weaving spider (Araneae: Araneidae). Biol. Lett. 2, 447–450 10.1098/rsbl.2006.0506 (doi:10.1098/rsbl.2006.0506) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Selden P. A. 1989. Orb-web weaving spiders in the Early Cretaceous. Nature 340, 711–713 10.1038/340711a0 (doi:10.1038/340711a0) [DOI] [Google Scholar]
- 14.Harmer A. M. T., Blackledge T. A., Madin J. S., Herberstein M. E. 2011. High-performance spider webs: integrating biomechanics, ecology and behaviour. J. R. Soc. Interface 8, 457–471 10.1098/rsif.2010.0454 (doi:10.1098/rsif.2010.0454) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Coddington J. 1982. Monophyletic origin of orb-webs. Am. Zool. 22, 886 [Google Scholar]
- 16.Vollrath F., Selden P. 2007. The role of behavior in the evolution of spiders, silks, and webs. Annu. Rev. Ecol. Evol. Syst. 38, 819–846 10.1146/annurev.ecolsys.37.091305.110221 (doi:10.1146/annurev.ecolsys.37.091305.110221) [DOI] [Google Scholar]
- 17.Blackledge T. A., Hayashi C. Y. 2006. Unraveling the mechanical properties of composite silk threads spun by cribellate orb-weaving spiders. J. Exp. Biol. 209, 3131–3140 10.1242/jeb.02327 (doi:10.1242/jeb.02327) [DOI] [PubMed] [Google Scholar]
- 18.Griswold C. E., Coddington J. A., Platnick N. I., Forster R. R. 1999. Towards a phylogeny of entelegyne spiders (Araneae, Araneomorphae, Entelegynae). J. Arachnol 27, 53–63 [Google Scholar]
- 19.Opell B. D. 1998. Economics of spider orb-webs: the benefits of producing adhesive capture thread and of recycling silk. Funct. Ecol. 12, 613–624 10.1046/j.1365-2435.1998.00222.x (doi:10.1046/j.1365-2435.1998.00222.x) [DOI] [Google Scholar]
- 20.Swanson B. O., Blackledge T. A., Hayashi C. Y. 2007. Spider capture silk: performance implications of variation in an exceptional biomaterial. J. Exp. Zool. A 307, 654–666 10.1002/jez.420 (doi:10.1002/jez.420) [DOI] [PubMed] [Google Scholar]
- 21.Eberhard W., Pereira F. 1993. Ultrastructure of cribellate silk of 9 species in 8 families and possible taxonomic implications (Araneae, Amaurobiidae, Deinopidae, Desidae, Dictynidae, Filistatidae, Hypochilidae, Stiphidiidae, Tengellidae). J. Arachnol. 21, 161–174 [Google Scholar]
- 22.Opell B. D. 1994. Factors governing the stickiness of cribellar prey capture threads in the spider family Uloboridae. J. Morphol. 221, 111–119 10.1002/jmor.1052210109 (doi:10.1002/jmor.1052210109) [DOI] [PubMed] [Google Scholar]
- 23.Dondale C. D. and National Research Council Canada 2003. Agriculture and agri-food Canada. Research Branch. The orb-weaving spiders of Canada and Alaska: Araneae : Uloboridae, Tetragnathidae, Araneidae, Theridiosomatidae. Ottawa, Canada: NRC Research Press [Google Scholar]
- 24.Peters H. M. 1995. Ultrastructure of orb spiders gluey capture threads. Naturwissenschaften 82, 380–382 10.1007/BF01134566 (doi:10.1007/BF01134566) [DOI] [Google Scholar]
- 25.Tillinghast E. K., Townley M. A., Wight T. N., Uhlenbruck G., Janssen E. 1993. The adhesive glycoprotein of the orb web of Argiope aurantia (Araneae, Araneidae). Mater. Res. Soc. Symp. P 292, 9–23 10.1557/PROC-292-9 (doi:10.1557/PROC-292-9) [DOI] [Google Scholar]
- 26.Vollrath F., Tillinghast E. K. 1991. Glycoprotein glue beneath a spider webs aqueous coat. Naturwissenschaften 78, 557–559 10.1007/BF01134447 (doi:10.1007/BF01134447) [DOI] [Google Scholar]
- 27.Vollrath F., Fairbrother W. J., Williams R. J. P., Tillinghast E. K., Bernstein D. T., Gallagher K. S., Townley M. K. 1990. Compounds in the droplets of the orb spiders viscid spiral. Nature 345, 526–528 10.1038/345526a0 (doi:10.1038/345526a0) [DOI] [Google Scholar]
- 28.Swanson B. O., Blackledge T. A., Beltrán J., Hayashi C. Y. 2005. Variation in the material properties of spider dragline silk across species. Appl. Phys. A 82, 213–218 10.1007/s00339-005-3427-6 (doi:10.1007/s00339-005-3427-6) [DOI] [Google Scholar]
- 29.Opell B. D. 1999. Redesigning spider webs: stickiness, capture area and the evolution of modern orb-webs. Evol. Ecol. Res. 1, 503–516 [Google Scholar]
- 30.Opell B. D., Schwend H. S. 2009. Adhesive efficiency of spider prey capture threads. Zoology 112, 16–26 10.1016/j.zool.2008.04.002 (doi:10.1016/j.zool.2008.04.002) [DOI] [PubMed] [Google Scholar]
- 31.Opell B. D. 1997. A comparison of capture thread and architectural features of deinopoid and araneoid orb-webs. J. Arachnol. 25, 295–306 [Google Scholar]
- 32.Swanson B. O., Anderson S. P., Digiovine C., Ross R. N., Dorsey J. P. 2009. The evolution of complex biomaterial performance: the case of spider silk. Integr. Comp. Biol. 49, 21–31 10.1093/icb/icp013 (doi:10.1093/icb/icp013) [DOI] [PubMed] [Google Scholar]
- 33.Craig C. L. 1987. The ecological and evolutionary interdependence between web architecture and web silk spun by orb web weaving spiders. Biol. J. Linn. Soc. 30, 135–62 10.1111/j.1095-8312.1987.tb00294.x (doi:10.1111/j.1095-8312.1987.tb00294.x) [DOI] [Google Scholar]
- 34.Opell B. D. 1996. Functional similarities of spider webs with diverse architectures. Am. Nat. 148, 630–48 10.1086/285944 (doi:10.1086/285944) [DOI] [Google Scholar]
- 35.Sherman P. M. 1994. The orb-web: an energetic and behavioral estimator of a spiders dynamic foraging and reproductive strategies. Anim. Behav. 48, 19–34 10.1006/anbe.1994.1208 (doi:10.1006/anbe.1994.1208) [DOI] [Google Scholar]
- 36.Opell B. D., Bond J. E. 2001. Changes in the mechanical properties of capture threads and the evolution of modern orb-weaving spiders. Evol. Ecol. Res. 3, 567–581 [Google Scholar]
- 37.Eberhard W. G. 1989. Effects of orb-web orientation and spider size on prey retention. Bull. Br. Arachnol. Soc. 8, 45–48 [Google Scholar]
- 38.Tanaka K. 1989. Energetic cost of web construction and its effect on web relocation in the web-building spider Agelena limbata. Oecologia 81, 459–464 10.1007/BF00378952 (doi:10.1007/BF00378952) [DOI] [PubMed] [Google Scholar]
- 39.Peakall D. B., Witt P. N. 1976. Energy budget of an orb web-building spider. Comp. Biochem. Phys. A 54, 187–190 10.1016/S0300-9629(76)80094-1 (doi:10.1016/S0300-9629(76)80094-1) [DOI] [PubMed] [Google Scholar]
- 40.Kohler T., Vollrath F. 1995. Thread biomechanics in the 2 orb-weaving spiders Araneus diadematus (Araneae, Araneidae) and Uloborus walckenaerius (Araneae, Uloboridae). J. Exp. Zool. 271, 1–17 10.1002/jez.1402710102 (doi:10.1002/jez.1402710102) [DOI] [Google Scholar]
- 41.Lin L. H., Edmonds D. T., Vollrath F. 1995. Structural-engineering of an orb-spiders web. Nature 373, 146–148 10.1038/373146a0 (doi:10.1038/373146a0) [DOI] [Google Scholar]
- 42.Sensenig A., Agnarsson I., Blackledge T. A. 2010. Behavioural and biomaterial coevolution in spider orb webs. J. Evol. Biol. 23, 1839–1856 10.1111/j.1420-9101.2010.02048.x (doi:10.1111/j.1420-9101.2010.02048.x) [DOI] [PubMed] [Google Scholar]
- 43.Bond J. E., Opell B. D. 1998. Testing adaptive radiation and key innovation hypotheses in spiders. Evolution 52, 403–414 10.2307/2411077 (doi:10.2307/2411077) [DOI] [PubMed] [Google Scholar]
- 44.Cranford S., Tarakanova A., Pugno N. M., Buehler M. J. 2012. Nonlinear material behaviour of spider silk yields robust webs. Nature 482, 72–76 10.1038/nature10739 (doi:10.1038/nature10739) [DOI] [PubMed] [Google Scholar]
- 45.Keten S., Buehler M. J. 2010. Atomistic model of the spider silk nanostructure. Appl. Phys. Lett. 96, 153701 10.1063/1.3385388 (doi:10.1063/1.3385388) [DOI] [Google Scholar]
- 46.Keten S., Xu Z., Ihle B., Buehler M. J. 2010. Nanoconfinement controls stiffness, strength and mechanical toughness of beta sheet crystals in silk. Nat. Mater. 9, 359–367 10.1038/nmat2704 (doi:10.1038/nmat2704) [DOI] [PubMed] [Google Scholar]
- 47.Keten S., Buehler M. J. 2010. Nanostructure and molecular mechanics of spider dragline silk protein assemblies. J. R. Soc. Interface 7, 1709–1721 10.1098/rsif.2010.0149 (doi:10.1098/rsif.2010.0149) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Nova A., Keten S., Pugno N. M., Redaelli A., Buehler M. J. 2010. Molecular and nanostructural mechanisms of deformation, strength and toughness of spider silk fibrils. Nano Lett. 10, 2626–2634 10.1021/nl101341w (doi:10.1021/nl101341w) [DOI] [PubMed] [Google Scholar]
- 49.van Beek J. D., Hess S., Vollrath F., Meier B. H. 2002. The molecular structure of spider dragline silk: folding and orientation of the protein backbone. Proc. Natl Acad. Sci. USA 99, 10 266–10 271 10.1073/pnas.152162299 (doi:10.1073/pnas.152162299) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Alam M. S., Wahab M. A., Jenkins C. H. 2007. Mechanics in naturally compliant structures. Mech. Mater. 39, 145–160 10.1016/j.mechmat.2006.04.005 (doi:10.1016/j.mechmat.2006.04.005) [DOI] [Google Scholar]
- 51.Lin L. H., Sobek W. 1998. Structural hierarchy in spider webs and spiderweb-type system. Struct. Eng. 76, 59–64 [Google Scholar]
- 52.Gosline J. M., Guerette P. A., Ortlepp C. S., Savage K. N. 1999. The mechanical design of spider silks: from fibroin sequence to mechanical function. J. Exp. Biol. 202, 3295–3303 [DOI] [PubMed] [Google Scholar]
- 53.Sensenig A. T., Lorentz K. A., Kelly S. P., Blackledge T. A. 2012. Spider orb webs rely on radial threads to absorb prey kinetic energy. J. R. Soc. Interface. 10.1098/rsif.2011.0851 (doi:10.1098/rsif.2011.0851) [DOI] [PMC free article] [PubMed] [Google Scholar]