

The role of natural killer cells in dendritic cell–based cancer vaccination strategies is comprehensively reviewed. Different strategies to improve dendritic cell vaccine preparations through engagement of natural killer cells are discussed and a summary of relevant natural killer cell parameters for immune monitoring is provided.
Keywords: Natural killer cells, Dendritic cell–based vaccination, Cancer immunotherapy, NK cell immune monitoring, NK–DC crosstalk
Learning Objectives
After completing this course, the reader will be able to:
Describe the current in vivo experimental and clinical dendritic cell (DC) vaccination studies encompassing the monitoring of natural killer (NK) cells.
Discuss the evaluation of NK cell stimulating potency in the design of DC-based cancer vaccines in the preclinical phase and in clinical trials.
Explain the added value of immune monitoring of NK cells in cancer vaccination trials.
This article is available for continuing medical education credit at CME.TheOncologist.com
Abstract
The cytotoxic and regulatory antitumor functions of natural killer (NK) cells have become attractive targets for immunotherapy. Manipulation of specific NK cell functions and their reciprocal interactions with dendritic cells (DCs) might hold therapeutic promise. In this review, we focus on the engagement of NK cells in DC-based cancer vaccination strategies, providing a comprehensive overview of current in vivo experimental and clinical DC vaccination studies encompassing the monitoring of NK cells. From these studies, it is clear that NK cells play a key regulatory role in the generation of DC-induced antitumor immunity, favoring the concept that targeting both innate and adaptive immune mechanisms may synergistically promote clinical outcome. However, to date, DC vaccination trials are only infrequently accompanied by NK cell monitoring. Here, we discuss different strategies to improve DC vaccine preparations via exploitation of NK cells and provide a summary of relevant NK cell parameters for immune monitoring. We underscore that the design of DC-based cancer vaccines should include the evaluation of their NK cell stimulating potency both in the preclinical phase and in clinical trials.
Natural Killer Cells in Cancer
In the early 1980s, the role of natural killer (NK) cells in defense against cancer was described in seminal reviews [1, 2]. A myriad of reports rapidly followed, supporting the involvement and therapeutic potential of NK cells in cancer immunity [3, 4]. A range of solid tumors [5–12] and hematological malignancies [13–19] were shown to be associated with significantly impaired NK cell functions. Importantly, NK cell abnormalities have been shown to be, at least in part, responsible for the failure of antitumor immunity. Deficiencies can reside in all NK cell populations, located in peripheral blood, in (lymphoid) organs, and in the tumor itself [16]. Functional impairment can originate from (a) primary NK cell dysfunction (e.g., imbalanced NK cell receptor expression, impaired cytolytic capacity, reduced cytokine secretion potency), (b) insufficient interaction with other immune cells (e.g., impaired killing of dendritic cells [DCs]) [14], (c) active immune suppression (e.g., regulatory T cell [Treg]-mediated suppression) [20, 21], and (d) NK cell resistance mechanisms by tumor cells (e.g., shedding of decoy molecules for activating receptors) [22]. In this regard, multiple cancer studies point toward a prognostic value for NK cells. Table 1 summarizes valuable NK cell parameters used for prognosis of disease progression and patient survival as well as for prediction of therapy efficacy.
Table 1.
Prognostic and predictive NK cell parameters
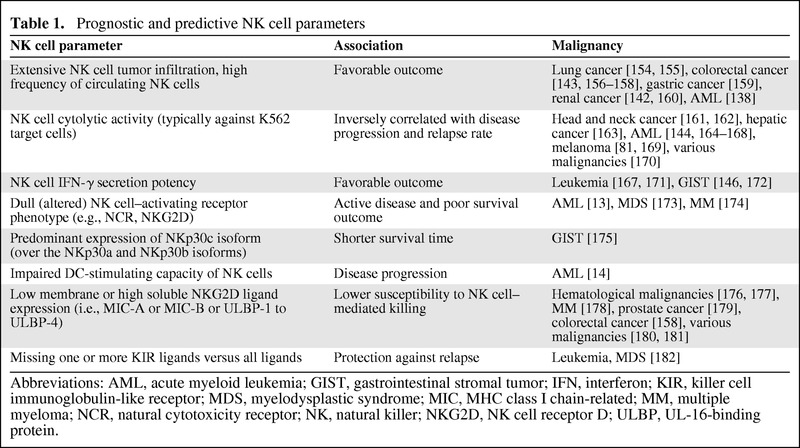
Abbreviations: AML, acute myeloid leukemia; GIST, gastrointestinal stromal tumor; IFN, interferon; KIR, killer cell immunoglobulin-like receptor; MDS, myelodysplastic syndrome; MIC, MHC class I chain-related; MM, multiple myeloma; NCR, natural cytotoxicity receptor; NK, natural killer; NKG2D, NK cell receptor D; ULBP, UL-16-binding protein.
In humans, NK cells are characterized by a CD56+CD3−NKp46+ phenotype. Based on their CD56 cell-surface density, they can be divided into two subsets with distinct phenotypic properties and key effector functions [23]. The majority (∼90%) of peripheral blood NK cells have a CD56dim CD16bright phenotype and were originally regarded as the more naturally cytotoxic subset, characterized by high cytotoxic granule and perforin expression and lower cytokine-secreting capacity. The smaller CD56brightCD16dim/− NK cell fraction (∼10%) constitutively expresses a higher number of cytokine and chemokine receptors and a lower amount of cytotoxic granules, generally showing a poorer cytotoxic capacity but a superior ability to produce abundant immunoregulatory cytokines following activation, in particular the prototypic cytokine interferon (IFN)-γ. Remarkably, these seemingly subtype-specific effector functions appear to be not as restricted as previously thought. Several research groups recently demonstrated that CD56dimCD16bright NK cells can be more prominent cytokine and chemokine producers than CD56brightCD16dim/− NK cells, depending on the stimuli received [24–26].
Altogether, it is well accepted that NK cells possess potent antitumor functions that could be targeted for immune-based therapy [27–30]. Their direct antitumor effects can be attributed to their cellular cytotoxicity and cytokine-producing capacities. NK cells can also indirectly contribute to tumor control by communicating with other immune cells (e.g., DCs, NKT cells, and T cells), leading to an efficient adaptive antitumor response [31–33]. In this review, we focus on the role of NK cells in DC-based cancer vaccination strategies, providing a comprehensive overview of current in vivo preclinical and clinical DC vaccination studies. We discuss different strategies to improve DC vaccine preparations through engagement of NK cells and we provide a summary of relevant NK cell parameters for immune monitoring.
DC- and NK Cell–Based Immunotherapy of Cancer
DCs are widely recognized as key antigen-presenting cells, critical for the induction of cellular immunity and central orchestrators of the immune system, bridging innate and adaptive immunity [34]. In this perspective, it has become clear that NK cells and DCs are in close communication, supporting the development of an efficient antitumor immune response. DC-mediated activation of NK cells contributes to the development of potent innate immunity, whereas, in turn, activated NK cells provide signals for DC activation, maturation, and cytokine production, promoting adaptive immunity (Fig. 1) (extensively reviewed elsewhere [31, 34–37]). Current understanding supports the idea of exploiting NK–DC immune interactions in cancer immunotherapy (Fig. 1). A number of (combined) therapies, for example, cytokine [38], monoclonal antibody [39], and drug [40] therapy, qualify as acting on both immune cell types. Also, tumor cell vaccines that target both the hosts' NK cells and DCs may be successful [41–44]. Overall, improvement in tumor cell recognition by NK cells (cytotoxicity) and DCs (phagocytosis and antigen presentation), including stimulation of NK–DC interaction to advance sustained T cell immunity, should be considered in the design of novel immunotherapeutic protocols.
Figure 1.
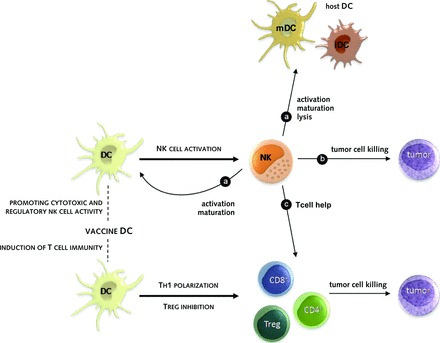
How NK cells can contribute to the antitumor efficacy of DC-based vaccination. Vaccine DCs can activate NK cells to (A) further stimulate vaccine and host DCs to advance sustained antitumor T-cell immunity, to (B) directly kill tumor cells, reducing the tumor burden and providing tumor cell material for further processing, and to (C) facilitate robust T-cell activation.
Abbreviations: iDC, immature dendritic cell; mDC, mature dendritic cell; NK, natural killer cell; Th1, T helper 1; Treg, regulatory T cell.
Several NK cell–based immunotherapeutic strategies have been developed, comprising both direct and indirect targeting of NK cell functions. Two main approaches can be distinguished, that is, adoptive transfer of NK cells [45–47] and in vivo modulation of endogenous NK cells [28, 29, 35, 48, 49], to bring about antitumor effects. Focusing on the latter, the most powerful immunoregulatory approach would be to attract NK cells to the site of interest, to target both their cytotoxic and their immunoregulatory functions, and to break tumor-mediated immune suppression or escape (Fig. 1). Following the expanding knowledge on NK–DC crosstalk, several groups have advocated the need to harness these immune interactions in the design of active specific immunotherapy [48, 50, 51]. In view of this, DC-based vaccination would be an obvious strategy to bring about NK cell activation (Fig. 1).
DC-based therapy originates from the unique capacity of DCs to present captured antigens to T cells, which is essential for generating specific T cell immunity [51–54]. Loading of DCs with tumor-associated antigens (TAAs) has therefore become an attractive therapeutic tool in order to generate tumor-specific T cells. The immunological outcome of DC vaccination strategies is predominantly evaluated based on induction of specific T cell responses. Indeed, most (pre)clinical trials provide evidence of tumor-specific CD8+ and/or CD4+ T cell responses. However, a discrepancy between these immunological responses and clinical outcome remains [55]. Studies that could demonstrate clinical tumor regression without detecting specific T cell generation prompted the investigation of other mechanisms that could be attributed to the observed effect [56, 57]. Given the importance of NK–DC crosstalk in the development of an immune response, specific monitoring of NK cells and their responses is emerging as a valuable avenue in DC vaccine protocols. Vaccine DCs can stimulate the cytotoxic activity, cytokine secretion (IFN-γ, tumor necrosis factor [TNF]), and migration of NK cells by soluble (e.g., interleukin [IL]-12, IL-15, IL-18, type I IFN, CXC chemokine receptor ligand 9 [CXCL9], CXCL10, CXCL11, C-C chemokine ligand 5 [CCL5]) and contact-dependent (e.g., NKp30, NKG2D) signals, contributing to the development of potent innate immunity (Fig. 1). In turn, activated NK cells can further stimulate vaccine and host DCs, contributing to recruitment, activation or maturation, and lysis of DCs to advance sustained antitumor T cell immunity and limit inappropriate T cell tolerization (Fig. 1A). Additionally, enhanced killing of antigen-expressing target cells by NK cells can directly reduce tumor burden and provide (extra) tumor cell material for uptake by DCs and antigen presentation to T cells (Fig. 1B). Furthermore, both NK cell regulatory and lytic activities can facilitate robust T cell activation in direct (e.g., IFN-γ, killing) and indirect (e.g., T helper 1 [Th1] DC polarization, lysis of immature DCs [iDCs] to prevent inappropriate T cell tolerization) ways, whereas production of Th1 cytokines by T cells, in turn, can further promote NK cell activation (Fig. 1C).
NK Cells in Preclinical DC-Based Vaccination
Fernandez and colleagues were the first to address NK–DC crosstalk in cancer, uncovering that DCs support the antitumor activity of NK cells [58]. They demonstrated, in mice, significant NK cell–mediated tumor growth delay upon adoptive transfer of DCs and confirmed the NK–DC interaction in vitro by showing DC-induced improvement in NK cell–mediated cytotoxic and IFN-γ–producing capacity. Ensuing from these data, they emphasized that DC-based immunotherapy may not only promote T cell–dependent antitumor immunity, but it may also directly trigger NK cells in vivo. At the same time, Cayeux et al. [59] demonstrated that DCs, either peptide pulsed, antigen loaded, or gene modified, generated specific cytotoxic T lymphocytes (CTLs) and rejected in vivo tumor challenge, together with an increase in splenic NK cells and lysis of DCs by IL-2–activated NK cells, inferring that antigens released from lysed DC vaccines could in turn be taken up by host antigen-presenting cells to further enhance immunity [59].
A plethora of research publications on this novel topic of NK–DC interaction rapidly followed, although up until now the effectiveness of DC-based vaccines in clinical settings has rarely been accompanied by NK cell data. Some groups have put forward the potential of DC vaccination to stimulate NK cell functions and supported the idea that DC vaccine efficacy is likely to be dependent on NK cell stimulatory capacity [31, 60–62]. Although, in general, DC-based vaccines are aimed to elicit antitumor T cell responses, in fact, a range of mouse in vivo studies demonstrated that immunization with activated DCs leads to significant antitumor immunity in an NK cell–dependent manner, regulated via direct NK cell effects and/or a critical interplay among NK cells, T cells, and DCs. Table 2 provides a comprehensive overview of preclinical DC vaccination studies exploring the involvement of NK cells. Routinely, bone marrow–derived DCs were loaded with tumor antigens (peptide, protein, lysate) via pulsing or viral transduction and administered through various routes. In some cases, a surrogate tumor antigen, such as ovalbumin (Ova) or β-galactosidase (β-gal), was used to verify antigen specificity. The involvement of NK cells was principally evidenced by in vivo antibody-mediated NK cell depletion or the use of beige NK cell–deficient mice (Table 2).
Table 2.
Preclinical DC-based cancer vaccination models evaluating involvement of NK cells
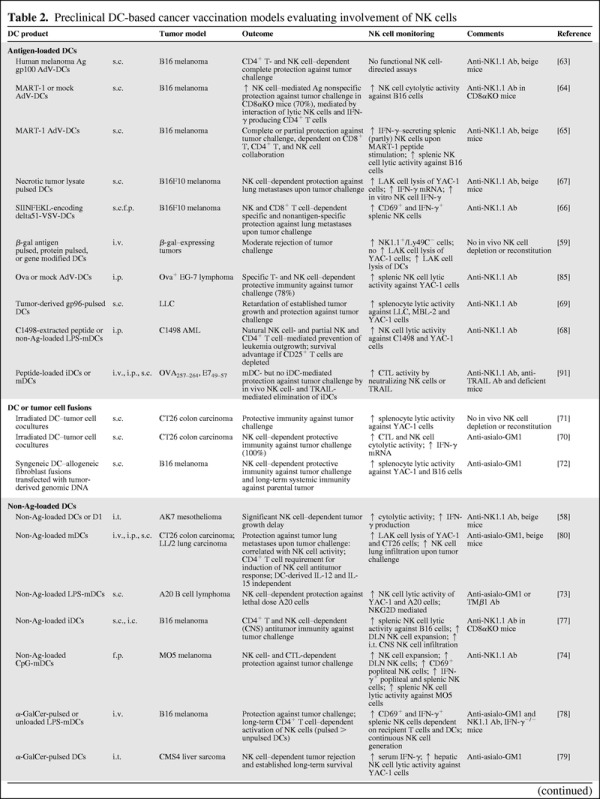
Table 2.
(Continued)
Abbreviations: 5-FU, 5-fluorouracil; α-GalCer, α-galactosylceramide; Ab, antibody; AdV, adenovirus-transduced; Ag, antigen; AML, acute myeloid leukemia; β-gal, β-galactosidase; beige mice, lack NK cell function; CNS, central nervous system; CpG, DNA sequence TLR9 ligand; CTL, cytotoxic T lymphocyte; D1, splenic immature dendritic cell line; DC, bone marrow-derived dendritic cell; Dex, DC-derived exosomes; DLN, draining lymph nodes; f.p., footpad; gp, glycoprotein; i.c., intracranial; i.d., intradermal; iDC, immature DC; IL, interleukin; i.p., intraperitoneal; i.t., intratumoral; KO, knockout; LAK, lymphokine-activated killer; LLC, Lewis lung cancer; LPS, lipopolysaccharide TLR4 ligand; MART-1, melanoma antigen recognized by T cells; MBL-2, murine lymphoma cell line; mDC, mature DC; MO5, B16F10 melanoma derivative; NK cell, natural killer cell; NOG mice, nonobese diabetic severe combined immunodeficient IL2Rγnull mice lacking lymphocytes; pDC, plasmacytoid DC; rSeV, recombinant Sendai virus; SV40, simian virus 40; TAA, tumor-associated antigen; TLR, Toll-like receptor; TRAIL, tumor necrosis factor-related apoptosis-inducing ligand; VSV, vesicular stomatitis virus-transduced; YAC-1, NK-sensitive mouse T cell lymphoma.
TAA-loaded DC vaccines were most frequently studied in the B16 murine melanoma model (Table 2). Immunization of mice with melanoma antigen–transduced DCs [63–66] or tumor lysate–pulsed DCs [67] resulted in an NK cell– and T cell–mediated complete or partial protection against tumor challenge. The outcome was typically measured by the absence or significant control of pulmonary metastases. TAA-loaded DC immunization has also led to efficient NK cell– and T cell–dependent antitumor immunity in other tumor models, resulting in protection against tumor challenge and prevention or retardation of established tumor growth [68, 69]. Additionally, it was demonstrated that, in combination with preventive anti-CD25 depletion to block Treg cells, the DC-mediated antitumor effect was more beneficial, allowing rejection of a much higher tumor burden and the development of long-lasting tumor protective immunity [68]. This is in agreement with the work of Ghiringhelli and colleagues, demonstrating that Treg cells inhibit NK cell functions [20]. The approach of DC–tumor cell fusions exploiting the whole tumor cell antigenic profile has also shown potent NK cell stimulatory capacities, resulting in NK cell–dependent protective immunity against tumor challenge [70–72] and long-term systemic immunity [72] (Table 2). Interestingly, several groups documented the ability of nonantigen-loaded DCs (lacking TAAs), alone or with an adjuvant, to exert protection against tumor challenge, to prevent the development of tumor metastases, to establish long-term survival, and to clear established tumor in an NK cell–dependent manner [73–84] (Table 2). Also, in control experiments of DC vaccination studies, the nonantigen-loaded DC control was able to induce NK cell–mediated immunity [64, 66, 68, 78, 85, 86].
In an alternative (nonantigen-loaded) strategy, DCs are modified to express high amounts of defined proinflammatory cytokines (e.g., IL-12, IL-15, IFN-γ, IFN-α, IL-18, IL-2) [53]. Cytokines play a pivotal role in the polarization and skewing of adaptive immunity and hence in DC-mediated T cell activation. Importantly, these cytokines could also play a key role in the activation of NK cells. Monitoring of NK cells following cytokine-secreting DC vaccination was performed in only four mouse studies, all evaluating DCs modified to secrete IL-12 (Th1-skewing cytokine and important NK cell activation signal) [86–88], including one additionally testing IL-15–secreting DCs (T cell– and NK cell–activating signal) [89]. Vaccination with IL-12– and IL-15–secreting DCs resulted in combinatorial NK cell– and T cell–dependent tumor eradication in all studies [86–89] and induced long-term protection against rechallenge with tumor cells [86] (Table 2). Rodriguez-Calvillo et al. [88] showed that tumor rejection was completely reliant on IFN-γ and IL-12. In an equivalent in vitro study with human DCs, the cytolytic activity and cytokine-secreting capacity of autologous NK cells was improved only if DCs were transfected to express human IL-12. Later on, that study was followed by a phase I clinical study in patients with metastatic gastrointestinal carcinomas, resulting in activated NK cells in five of 17 patients [90] (Table 3).
Table 3.
Clinical DC-based cancer vaccination trials with implementation of NK cell monitoring
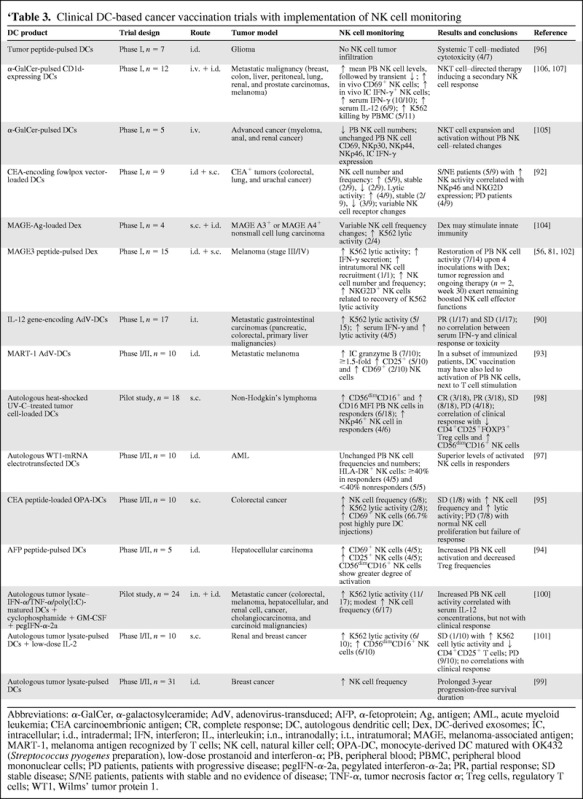
Abbreviations: α-GalCer, α-galactosylceramide; AdV, adenovirus-transduced; AFP, α-fetoprotein; Ag, antigen; AML, acute myeloid leukemia; CEA carcinoembrionic antigen; CR, complete response; DC, autologous dendritic cell; Dex, DC-derived exosomes; IC, intracellular; i.d., intradermal; IFN, interferon; IL, interleukin; i.n., intranodally; i.t., intratumoral; MAGE, melanoma-associated antigen; MART-1, melanoma antigen recognized by T cells; NK cell, natural killer cell; OPA-DC, monocyte-derived DC matured with OK432 (Streptococcus pyogenes preparation), low-dose prostanoid and interferon-α; PB, peripheral blood; PBMC, peripheral blood mononuclear cells; PD patients, patients with progressive disease; pegIFN-α-2a, pegylated interferon-α-2a; PR, partial response; SD stable disease; S/NE patients, patients with stable and no evidence of disease; TNF-α, tumor necrosis factor α; Treg cells, regulatory T cells; WT1, Wilms' tumor protein 1.
Several of the in vivo animal studies analyzed potential mechanisms of DC vaccine–induced antitumor effects in more detail, demonstrating the requirement of either the lytic capacity, cytokine-secreting properties, or both of NK cells and/or T cells (Table 2). For example, Ribas and colleagues demonstrated that effective DC-induced antitumor immunity was mediated by lytic NK cells and IFN-γ–producing CD4+ T cells [64]. Mechanistically, these preclinical studies support the complete or partial CD4+ T cell [64, 68, 77, 78, 82, 83], CD8+ T cell [66, 69, 74, 75, 89], or CD4+CD8+ T cell [65, 85, 86] requirements for DC vaccine–induced NK cell–mediated antitumor effects (Table 2). To a further extent, some immunization protocols could induce long-term activation of NK cells via continuous generation of NK cells, rather than a true memory response [78, 83], whereas transfer of activated splenocytes from DC-immunized mice to naïve mice was shown to exert a similar NK cell–mediated protective antitumor immune response [64]. In the illustrated studies, mostly matured DCs were used, described to be more resistant to NK cell–mediated killing. In this context, Hayakawa et al. [91] demonstrated in vivo that immature peptide-pulsed DCs were eliminated by NK cells via a TNF-related apoptosis-inducing ligand (TRAIL)-dependent pathway. Moreover, NK cell depletion or neutralizing TRAIL led to enhanced CTL activity and a higher tumor rejection rate upon live tumor challenge. Concomitantly, immunization with mature peptide-pulsed DCs showed high protection and, in contrast to iDCs this effect was not improved when NK cells were depleted or when TRAIL was neutralized [91].
NK Cells in Clinical DC-Based Cancer Vaccination
In general, the preclinical studies described above demonstrate that NK cells are necessary but not sufficient to induce adaptive T cell responses following DC vaccination, suggesting critical interplay among NK cells, T lymphocytes, and DCs. These findings provide sound evidence that DC-based strategies designed to engage both NK cell and T cell immunity could markedly improve current DC vaccine efficacy for the treatment of tumors (Fig. 1). In practice, though, only a few groups have implemented evaluation of NK cells in their clinical DC trials. Table 3 provides an exhaustive overview of DC-based tumor vaccination trials implementing NK cell monitoring, either planned or post hoc.
Monitoring of NK cells was most frequently studied in tumor antigen–loaded DC immunization trials (Table 3). Evaluating viral vector–transduced DC vaccination, Osada et al. [92] demonstrated, in patients with a carcinoembryonic antigen–positive tumor, that NKp46 and NKG2D expression correlated with patients' NK cell cytolytic activity and that NK cell cytolytic activity correlated with clinical outcome. Moreover, they postulated that NK cell responses may predict clinical outcome more closely than T cell responses. Butterfield et al. [93] evaluated a melanoma antigen recognized by T cells (MART)-1–transduced DC vaccine that was designed to activate MART-1–specific CD8+ and CD4+ T cells. Based on preclinical work from the same lab [65], they additionally performed NK cell analysis and could demonstrate, 2 weeks postvaccination, increased intracellular granzyme B content in NK cells in seven of 10 patients. These exploratory studies indicated that, in a subset of patients, the DC vaccine may have led to activation of circulating NK cells. Peptide-pulsed DC vaccination was shown to induce NK cell activation in some patients in two studies, demonstrated by increased peripheral blood NK cell frequency, activation status (CD69, CD25), and lytic activity [94, 95]. Bray et al. [94] also demonstrated a parallel decrease in Treg frequencies. A third study did not detect NK cell changes as assessed by NK cell tumor infiltration post-DC therapy [96]. In a recently published phase I trial by our own group [97], evaluating a therapeutic Wilms' tumor I antigen–targeted DC vaccine in patients with acute myeloid leukemia, we could demonstrate the induction of complete and molecular remission in 50% of patients. Clinical responses were correlated with both innate and adaptive immune responses postvaccination. Four of five patients who responded to the DC vaccine had a highly activated NK cell phenotype, in comparison with none of the nonresponders. Extensive NK cell monitoring in a subsequent phase II trial is ongoing.
All studies with tumor lysate–pulsed DC vaccination, alone [98, 99] or as a part of combination therapy [100, 101], led to NK cell changes (Table 3). Di Nicola and colleagues found a positive correlation between an increased frequency of CD56dimCD16+ NK cells and clinical response post-DC vaccination [98]. Strengthening the clinical observations of others [94, 101], they also found a positive correlation between a decreased frequency of CD4+CD25+FOXP3+ Treg cells and clinical response. The other studies also demonstrated favorable NK cell changes (increased NK cell frequency and lytic activity) in some patients, but failed to make NK cell–related correlations with clinical outcome [99–101].
Two groups investigated a cell-free DC-based vaccine preparation consisting of melanoma-associated antigen–pulsed autologous DC-derived exosomes (Dex; exosomes are nanometer-sized membrane vesicles invaginating from multivesicular bodies and secreted from different cell types [102, 103]) (Table 3). Both could demonstrate an increase in NK cell lytic activity in half of the patients receiving Dex [56, 104]. Escudier and coworkers also showed that NK cells were capable of recognizing autologous tumor cells following Dex therapy and, for one patient, recruitment of NK cells inside tumor areas could be documented. Based on these results, they pursued exploring the NK cell–stimulating capacity of Dex vaccination with preclinical and clinical studies, including additional ex vivo experiments [81] of patient samples taken during their phase I study [56]. In line with the murine data, they confirmed in vitro NKG2D-mediated NK cell activation by human Dex. From the phase I study, they additionally showed that Dex boosted recirculation of NK cells and restored NKG2D expression and the cytotoxic capacity of NK cells in half of the immunized patients. In two patients with tumor regression and continuing treatment, NK cell effector functions remained boosted at later time points.
Secondary NK cell responses were monitored in two phase I studies following NKT cell–directed therapy with α-galactosylceramide (α-GalCer)-pulsed DCs in a mixed patient group with metastatic malignancies (Table 3) [105–107]. The glycolipid antigen antitumor compound α-GalCer is a specific NKT cell ligand with the potential to sufficiently activate NKT cells and induce secondary immune effects, including T cell and NK cell activation [107]. Both groups could demonstrate a transient decrease in peripheral blood NK cells. However, Nieda and colleagues demonstrated that α-GalCer–pulsed DCs induced a significant secondary NK cell response [106, 107], whereas Chang et al. [105] could not detect significant differences for any of the NK cell parameters measured.
From the clinical trials discussed above, we can conclude that DC-based cancer vaccination can have a significant impact on primary and secondary changes in the NK cell compartment. The results strongly indicate that increased DC vaccine–induced NK cell activity, in terms of phenotype, lytic, and/or regulatory functions, points to a more beneficial outcome [56, 81, 95, 97] and show that some DC-mediated NK cell changes are correlated with clinical response [92, 98] (Table 3). However, the limited number of (uniform) trials (e.g., mixed patient groups) and the diversity of DC vaccines (preparation, routes of administration, vaccine doses, and intervals) and of NK cell immunomonitoring assays do not allow the drawing of solid conclusions. For example, some studies demonstrated an increase [81, 95, 98–100] in peripheral blood NK cell frequency after DC vaccination, whereas others saw no [97] or variable [92, 104] changes or a decrease [105–107]. In this context, scrutiny of changes in NK cell frequency and function in blood, lymphoid, and tumor tissues is needed to fully judge the impact of DC vaccines on NK cells in vivo in humans. In accordance with the preclinical data of Martin-Fontecha and colleagues [108], DC-mediated transient NK cell recruitment to lymphoid tissue, where NK–DC interactions are likely to take place, could account for the peripheral decrease in NK cells observed in a number of clinical trials [105–107]. Further exploration of NK cell engagement and consistent monitoring in DC-based trials is therefore of utmost importance to learn more about the relevance of NK cells in diverse DC-based cancer vaccination approaches.
Improvement of DC Vaccines via NK Cell Engagement
Given the limited clinical benefit so far, increasing attention is being paid to the amelioration of the therapeutic efficacy of DC vaccines. The list of approaches is growing and several have been identified as highly innovative [53, 54, 62, 109]. As highlighted in the present review, a promising strategy to improve DC-based vaccination is through ex vivo instruction of DCs to preferentially activate NK cells. Based on the current knowledge on NK–DC crosstalk, DC preparations could be optimized for a variety of parameters to harness DCs for NK cell activation during immunotherapy for cancer. Activated NK cells can, in turn, lead to further stimulation of vaccine and host DCs (Fig. 1).
According to the DC subset and differentiation or maturation protocol used, DCs can have different effects on NK cells [110, 111]. It has been demonstrated that DC generation with inclusion of IFN-α [112–116] or IL-15 [117–119] in differentiation cocktails and the use of Toll-like receptor (TLR) agonists in maturation cocktails [115, 117, 120–124] hold promise in enhancing the NK cell–activating capacity of DC vaccines and their reciprocal crosstalk and T cell activation. As illustrated in a human in vitro study [116], IFN-α–treated monocyte-derived DCs (IFN-DCs) induced specific CD8+ T cell responses against melanoma peptides only in the presence of NK cells. IFN-DCs were also shown to recruit NK cells [113] and to directly activate NK cells, as measured by CD69 expression and IFN-γ production [112]. Depending on their maturation status, DCs are sensitive to NK cell–mediated killing [59, 91, 125], which can affect the survival and immunogenicity of the DC vaccine.
The mode of transfection or transduction and the antigen preparation are critical for the DC activation status and for communication with different immune cells. Numerous antigen preparations (peptide, protein, cell lysate, apoptotic tumor cells, nucleic acids) and loading methods (pulsing, viral transduction, electroporation, lipofection, gene transduction) have been investigated to optimize T cell–stimulating capacity, but some were also examined for their NK cell–activating competence (Table 2). For example, the use of apoptotic tumor cell–loaded DCs was demonstrated to induce an NK cell–dependent T cell response [126, 127]. Considering the mode of delivery, Vujanovic and colleagues [124] demonstrated, in a recent human trial, that adenoviral transduction, like TLR4 ligand/IFN-γ maturation, increased the expression of transmembrane TNF and IL-15 on DCs, resulting in enhanced NK cell activation, IFN-γ secretion, proliferation, and antitumor activities [93, 124]. They showed, in vitro, that adenovirus-transduced DCs, as compared with protein- or peptide-loaded DCs, are the most successful at inducing NK cell activation [94]. It was confirmed, in mouse models, that adenovirus-transduced DCs induced significant NK cell antitumor activity in vivo [85, 124]. Analogous results were found for prophylactic and therapeutic vesicular stomatitis virus–transduced DC vaccination in mice [66, 84] (Table 2).
Recruiting immune cells to the sites of interest (tumor and immune cell–enriched sites) and bringing them in proximity of each other is believed to be fruitful for immune activation [48, 128]. In this regard, the route of vaccine administration is an important determinant (Tables 2 and 3) [52]. A number of preclinical and clinical studies demonstrated that defined DC preparations were able to elicit the recruitment of NK cells through specific mechanisms. A first observation by Martin-Fontecha et al. [108] showed, in mice, that s.c. injection of mature DCs led to rapid recruitment of NK cells into the lymph nodes, providing IFN-γ for Th1 priming. In line with these findings, others have demonstrated the induction of NK cell infiltration in lymph nodes [83, 129] as well as in tumor [75] following immunization of mice with TLR ligand–activated DCs, resulting in NK cell and T cell activation as well as tumor regression. The NK cell recruiting capacities of diverse human DC vaccines have also been examined in multiple in vitro studies. Mature IFN-DCs [112, 113], DCs loaded with reovirus-infected melanoma cells [130], and DCs matured with bacterial fragments [123] were shown to secrete distinct NK cell–attracting chemokines (i.e., the CXCR3 ligands CXCL9, CXCL10, CXCL11 and the CCR5 ligand CCL5), mediating in vitro migration and stimulating NK cell activation, NK cell–mediated cytotoxicity, and IFN-γ production. In parallel, the chemokine receptor profile of NK cells (CCR5, CCR7, CXCR1, CXCR3, CX3CR1) can be differentially regulated by DC vaccines to promote NK cell trafficking [131].
A more direct approach is the transfection or transduction of DCs with NK cell stimulatory molecules, such as genes encoding IL-12, IL-15, or other NK cell–stimulating factors (so-called “designer” DCs). This strategy has already been demonstrated to be effective in activating NK cells and T cells, for example, by vaccinating metastatic gastrointestinal cancer patients with nonantigen-loaded IL-12–secreting DCs [90] (Table 3). Also, cotransfection (by electroporation) of TAAs with IL-12–encoding genes into mature human DCs induced an increase in activated NK cells and TAA-specific CTLs in vitro [131].
Finally, NK cells themselves can be used to improve the therapeutic efficacy of the DC vaccine [31, 50]. The generation of DC preparations in the presence of NK cells or NK cell–related factors was carried out by several groups in the preclinical setting [126, 132–135]. Using this strategy, ex vivo–generated DCs are brought into contact with NK cell–related signals with or without adjuvant, already bringing DCs to an activated status and priming the DCs for Th1 polarization.
In conclusion, these data underscore why future research efforts should also focus on optimizing the NK cell–activating properties of DC vaccines, in addition to improving their T cell stimulatory and Treg inhibitory capacities (Fig. 1). The NK cell–activating character of DC vaccine preparations can be imprinted at multiple levels: (a) DC differentiation and maturation, (b) source of antigen and delivery mode to DCs, (c) NK cell–trafficking potential, (d) direct DC modulation, and (e) with support of other immune cells such as NK cells themselves. DC vaccination trials should carefully evaluate how NK cell–mediated surveillance against tumor cells can be incorporated into DC-based therapy.
NK Cell Immunomonitoring
From a variety of cancer immunotherapeutic approaches (e.g., hematopoietic cell transplantation [136–139], cytokine biotherapy [38, 140, 141]), it could be perceived that NK cell activation can contribute to the overall antitumor response. An array of both common and experimental NK cell parameters that have been implemented at the clinical and preclinical level, respectively, are listed in Table 4. Classical NK cell immunomonitoring includes evaluation, prior to and after therapy, of NK cell frequencies and numbers, regularly complemented by multiparametric phenotypic analysis (Table 4). In general, the activity of peripheral blood NK cells is tested, although NK cells residing in lymphoid tissue or infiltrated inside [7, 8, 12, 142] or surrounding [6, 143] the tumor have also been assessed for monitoring. Functional assays are routinely focusing on NK cell–mediated cytotoxic activity against NK-sensitive target cells (K562; standard assay for the majority of studies) and NK-resistant cells (Daudi, Raji) in the presence or absence of the cytotoxicity-promoting cytokine IL-2. NK cell cytolytic activity was demonstrated to be a valuable prognostic marker for several malignancies (Table 1). Importantly, some studies performed killing assays against autologous primary tumor cells to evaluate therapy efficacy [102, 144], which is likely to be of high relevance in the setting of eliminating residual tumor cells to prevent relapse. Commonly relying on the established chromium release killing assay, some clinical studies sought to provide more detailed information on the potential mechanisms of cytotoxicity using single-cell assays (Table 4). Another functional characteristic that has also been assigned as a predictive parameter is NK cell IFN-γ production (Table 1). Higher IFN-γ production is an indicator of immunostimulatory resident NK cells, which is especially interesting in view of DC-based approaches. Other NK cell–related cytokines (e.g., TNF-α, GM-CSF, IL-5, IL-10, IL-13, IL-15) may be considered as interesting for therapy-related effects. For example, GM-CSF is a known DC differentiation factor and IL-10 is an acknowledged immune-inhibitory cytokine, whereas IL-15 favors the proliferation and survival of NK cells and T cells and is involved in NK–DC crosstalk [124, 145]. Of particular interest for DC-based therapy is the examination of the ability of NK cells to activate, maturate, attract, and promote phagocytosis of DCs (Table 4). Only a few studies have evaluated such functionalities in an experimental setting [11, 42, 112, 113, 123, 135, 146]. NK cell–mediated killing of iDCs is also an attractive parameter to follow [14]. Ideally, it could provide useful information on which patient subsets are more likely to respond to DC-based therapy [147]. Finally, indirect NK cell–related parameters are the subject of research, because a range of transformed cancer cells has been shown to downregulate or secrete soluble molecules (e.g., ligands for the activating receptor NKG2D) in order to evade NK cell immunosurveillance [16]. In line with this, evaluation of tumor cell sensitivity to NK cell–mediated killing would be a significant predictive marker for therapy effectiveness.
Table 4.
Human NK cell parameters for immunomonitoring
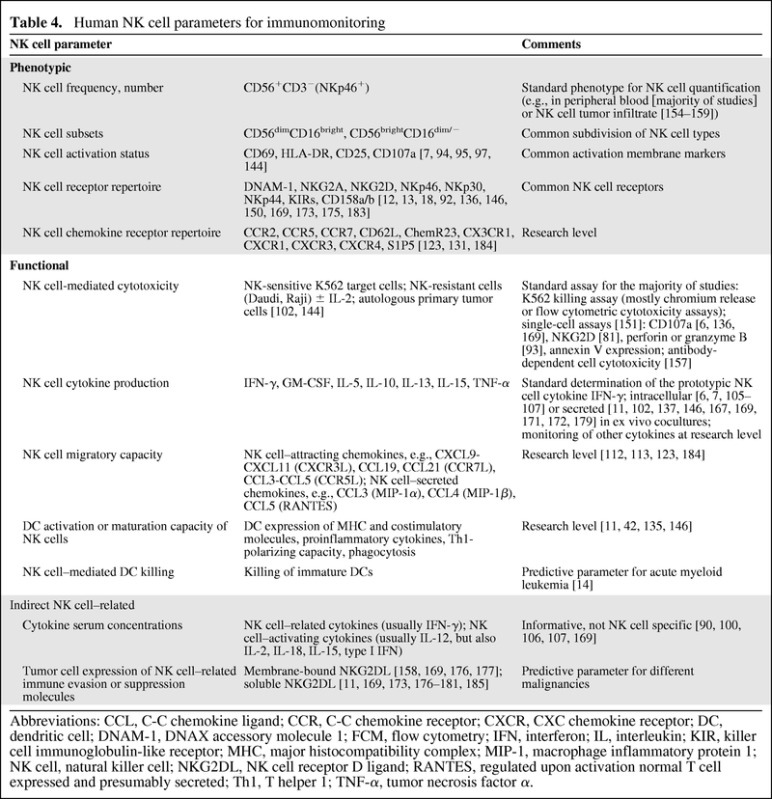
Abbreviations: CCL, C-C chemokine ligand; CCR, C-C chemokine receptor; CXCR, CXC chemokine receptor; DC, dendritic cell; DNAM-1, DNAX accessory molecule 1; FCM, flow cytometry; IFN, interferon; IL, interleukin; KIR, killer cell immunoglobulin-like receptor; MHC, major histocompatibility complex; MIP-1, macrophage inflammatory protein 1; NK cell, natural killer cell; NKG2DL, NK cell receptor D ligand; RANTES, regulated upon activation normal T cell expressed and presumably secreted; Th1, T helper 1; TNF-α, tumor necrosis factor α.
Since the perception of the clinical significance of NK cell activity, there has been a need for accurate data generation and analysis [148, 149]. Today it is clear that NK cell biology is more complex than initially considered. Consequently, NK cell activity cannot merely be monitored by plain cytotoxicity assays with NK-sensitive target cells or simple phenotypic analyses [48]. As depicted in Table 4, an assortment of supplementary phenotypic and functional NK cell–related parameters have already been applied. Notwithstanding, guiding principles for significant parameters should be developed and standardized, ultimately providing useful information on outcomes and therapies. Just recently, Björkström et al. [150] and Bryceson et al. [151] provided precise, up-to-date protocols for NK cell receptor repertoire analysis and functional analysis of human NK cells with flow cytometry. Importantly, similar to the assessment of T cell responses [152], monitoring NK cell parameters must be done with prudence. Functionally diverse NK cell subsets may act in a distinctive manner per malignancy and per therapy, and the use of various technical protocols may result in significant variable outcomes. For example, we demonstrated variation in the quantification of IFN-γ secreted by human NK cells following tumor cell stimulation using different methods [153].
Future Perspectives
DC-based vaccines are highly potent NK cell–stimulating candidates. Unlike the growing body of evidence acquired through experimental research (Table 2), there is a relative scarcity of data reporting NK cell function in DC trials (Table 3) [48, 50]. Herein, we provide an overview of mouse and human in vivo studies that investigated the influence of NK cell participation in antitumor immunity following DC-based vaccination strategies. In summary, these studies underscore a substantial role for both the cytotoxic and the regulatory functions of NK cells in the development of DC-mediated adaptive antitumor immunity, advocating for clear implementation of NK cell monitoring in cancer immunotherapy, in particular for DC-based therapy. Careful consideration of defined NK cell parameters and protocols is needed. Ultimately, this will contribute to a more complete understanding of therapy efficacy, generating valuable information that could be exploited in the development of novel adjuvants to improve anticancer immunotherapies. With the need for surrogate markers that could predict clinical outcome, research on NK cell activation as a prognostic factor warrants further intensification. In conclusion, a clear understanding of NK cell functions and mechanisms is essential to envision therapeutic breakthroughs in DC vaccine design. Hence, the development of DC-based vaccination strategies should implement NK cell–stimulating potency both in the preclinical phase and in clinical trials.
Acknowledgments
This work was supported in part by research grants of the Research Foundation – Flanders (FWO-Vlaanderen, G.0082.08 and G.0218.12), the Foundation Against Cancer (Stichting tegen Kanker), the Methusalem financing program of the Flemish Government to the Antwerp University and the Interuniversity Attraction Poles financing program (IAP #P6/41) of the Belgian Government. E.L. was funded by a Stichting Emmanuel van der Schueren research grant of the Vlaamse Liga tegen Kanker (VLK). E.S. is postdoctoral researcher of the Research Foundation – Flanders (FWO-Vlaanderen). The authors wish to thank Dr. Sébastien Anguille (Belgium) for assistance in the design of Figure 1 and for granting us the right to reproduce this artwork in the current publication.
Footnotes
- (C/A)
- Consulting/advisory relationship
- (RF)
- Research funding
- (E)
- Employment
- (H)
- Honoraria received
- (OI)
- Ownership interests
- (IP)
- Intellectual property rights/inventor/patent holder
- (SAB)
- Scientific advisory board
Author Contributions
Conception/Design: Eva Lion
Manuscript writing: Eva Lion
Final approval of manuscript: Evelien L.J.M. Smits, Zwi N. Berneman, Viggo F.I. Van Tendeloo
References
- 1.Herberman RB, Ortaldo JR. Natural killer cells: Their roles in defenses against disease. Science. 1981;214:24–30. doi: 10.1126/science.7025208. [DOI] [PubMed] [Google Scholar]
- 2.Herberman RB. Multiple functions of natural killer cells, including immunoregulation as well as resistance to tumor growth. Concepts Immunopathol. 1985;1:96–132. [PubMed] [Google Scholar]
- 3.Whiteside TL, Herberman RB. The role of natural killer cells in immune surveillance of cancer. Curr Opin Immunol. 1995;7:704–710. doi: 10.1016/0952-7915(95)80080-8. [DOI] [PubMed] [Google Scholar]
- 4.Brittenden J, Heys SD, Ross J, et al. Natural killer cells and cancer. Cancer. 1996;77:1226–1243. doi: 10.1002/(sici)1097-0142(19960401)77:7<1226::aid-cncr2>3.0.co;2-g. [DOI] [PubMed] [Google Scholar]
- 5.Bobek V, Boubelik M, Fiserová A, et al. Anticoagulant drugs increase natural killer cell activity in lung cancer. Lung Cancer. 2005;47:215–223. doi: 10.1016/j.lungcan.2004.06.012. [DOI] [PubMed] [Google Scholar]
- 6.Cai L, Zhang Z, Zhou L, et al. Functional impairment in circulating and intrahepatic NK cells and relative mechanism in hepatocellular carcinoma patients. Clin Immunol. 2008;129:428–437. doi: 10.1016/j.clim.2008.08.012. [DOI] [PubMed] [Google Scholar]
- 7.Carrega P, Morandi B, Costa R, et al. Natural killer cells infiltrating human nonsmall-cell lung cancer are enriched in CD56 bright CD16(-) cells and display an impaired capability to kill tumor cells. Cancer. 2008;112:863–875. doi: 10.1002/cncr.23239. [DOI] [PubMed] [Google Scholar]
- 8.Esendagli G, Bruderek K, Goldmann T, et al. Malignant and non-malignant lung tissue areas are differentially populated by natural killer cells and regulatory T cells in non-small cell lung cancer. Lung Cancer. 2008;59:32–40. doi: 10.1016/j.lungcan.2007.07.022. [DOI] [PubMed] [Google Scholar]
- 9.Gallego-Melcón S, Español Boren T, Sanchez de Toledo J, et al. Natural killer cell function in children with malignant solid neoplasias. Med Pediatr Oncol. 1991;19:175–181. doi: 10.1002/mpo.2950190306. [DOI] [PubMed] [Google Scholar]
- 10.Groh V, Wu J, Yee C, et al. Tumour-derived soluble MIC ligands impair expression of NKG2D and T-cell activation. Nature. 2002;419:734–738. doi: 10.1038/nature01112. [DOI] [PubMed] [Google Scholar]
- 11.Jinushi M, Takehara T, Tatsumi T, et al. Impairment of natural killer cell and dendritic cell functions by the soluble form of MHC class I-related chain A in advanced human hepatocellular carcinomas. J Hepatol. 2005;43:1013–1020. doi: 10.1016/j.jhep.2005.05.026. [DOI] [PubMed] [Google Scholar]
- 12.Le Maux Chansac B, Moretta A, Vergnon I, et al. NK cells infiltrating a MHC class I-deficient lung adenocarcinoma display impaired cytotoxic activity toward autologous tumor cells associated with altered NK cell-triggering receptors. J Immunol. 2005;175:5790–5798. doi: 10.4049/jimmunol.175.9.5790. [DOI] [PubMed] [Google Scholar]
- 13.Fauriat C, Just-Landi S, Mallet F, et al. Deficient expression of NCR in NK cells from acute myeloid leukemia: Evolution during leukemia treatment and impact of leukemia cells in NCRdull phenotype induction. Blood. 2007;109:323–330. doi: 10.1182/blood-2005-08-027979. [DOI] [PubMed] [Google Scholar]
- 14.Fauriat C, Moretta A, Olive D, et al. Defective killing of dendritic cells by autologous natural killer cells from acute myeloid leukemia patients. Blood. 2005;106:2186–2188. doi: 10.1182/blood-2005-03-1270. [DOI] [PubMed] [Google Scholar]
- 15.Konjevic G, Jurisic V, Banicevic B, et al. The difference in NK-cell activity between patients with non-Hodgkin's lymphomas and Hodgkin's disease. Br J Haematol. 1999;104:144–151. doi: 10.1046/j.1365-2141.1999.01129.x. [DOI] [PubMed] [Google Scholar]
- 16.Lion E, Willemen Y, Berneman ZN, et al. Natural killer cell immune escape in acute myeloid leukemia. Leukemia. 2012 Mar 26; doi: 10.1038/leu.2012.87. [Epub ahead of print]. doi: 10.1038/leu.2012.87. [DOI] [PubMed] [Google Scholar]
- 17.Sørskaar D, Lie SO, Førre O. Natural killer cell activity of peripheral blood and bone marrow mononuclear cells from patients with childhood acute lymphoblastic leukemia. Acta Paediatr Scand. 1985;74:433–437. doi: 10.1111/j.1651-2227.1985.tb10998.x. [DOI] [PubMed] [Google Scholar]
- 18.Verheyden S, Bernier M, Demanet C. Identification of natural killer cell receptor phenotypes associated with leukemia. Leukemia. 2004;18:2002–2007. doi: 10.1038/sj.leu.2403525. [DOI] [PubMed] [Google Scholar]
- 19.Kwong YL. Natural killer-cell malignancies: Diagnosis and treatment. Leukemia. 2005;19:2186–2194. doi: 10.1038/sj.leu.2403955. [DOI] [PubMed] [Google Scholar]
- 20.Ghiringhelli F, Ménard C, Martin F, et al. The role of regulatory T cells in the control of natural killer cells: Relevance during tumor progression. Immunol Rev. 2006;214:229–238. doi: 10.1111/j.1600-065X.2006.00445.x. [DOI] [PubMed] [Google Scholar]
- 21.Smyth MJ, Teng MW, Swann J, et al. CD4+-CD25+ T regulatory cells suppress NK cell-mediated immunotherapy of cancer. J Immunol. 2006;176:1582–1587. doi: 10.4049/jimmunol.176.3.1582. [DOI] [PubMed] [Google Scholar]
- 22.Zwirner NW, Fuertes MB, Girart MV, et al. Cytokine-driven regulation of NK cell functions in tumor immunity: Role of the MICA-NKG2D system. Cytokine Growth Factor Rev. 2007;18:159–170. doi: 10.1016/j.cytogfr.2007.01.013. [DOI] [PubMed] [Google Scholar]
- 23.Cooper MA, Fehniger TA, Caligiuri MA. The biology of human natural killer-cell subsets. Trends Immunol. 2001;22:633–640. doi: 10.1016/s1471-4906(01)02060-9. [DOI] [PubMed] [Google Scholar]
- 24.De Maria A, Bozzano F, Cantoni C, et al. Revisiting human natural killer cell subset function revealed cytolytic CD56(dim)CD16+ NK cells as rapid producers of abundant IFN-gamma on activation. Proc Natl Acad Sci U S A. 2011;108:728–732. doi: 10.1073/pnas.1012356108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Fauriat C, Long EO, Ljunggren HG, et al. Regulation of human NK-cell cytokine and chemokine production by target cell recognition. Blood. 2010;115:2167–2176. doi: 10.1182/blood-2009-08-238469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Korbel DS, Norman PJ, Newman KC, et al. Killer Ig-like receptor (KIR) genotype predicts the capacity of human KIR-positive CD56dim NK cells to respond to pathogen-associated signals. J Immunol. 2009;182:6426–6434. doi: 10.4049/jimmunol.0804224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Albertsson PA, Basse PH, Hokland M, et al. NK cells and the tumour microenvironment: Implications for NK-cell function and anti-tumour activity. Trends Immunol. 2003;24:603–609. doi: 10.1016/j.it.2003.09.007. [DOI] [PubMed] [Google Scholar]
- 28.Malmberg KJ, Bryceson YT, Carlsten M, et al. NK cell-mediated targeting of human cancer and possibilities for new means of immunotherapy. Cancer Immunol Immunother. 2008;57:1541–1552. doi: 10.1007/s00262-008-0492-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Sutlu T, Alici E. Natural killer cell-based immunotherapy in cancer: Current insights and future prospects. J Intern Med. 2009;266:154–181. doi: 10.1111/j.1365-2796.2009.02121.x. [DOI] [PubMed] [Google Scholar]
- 30.Zamai L, Ponti C, Mirandola P, et al. NK cells and cancer. J Immunol. 2007;178:4011–4016. doi: 10.4049/jimmunol.178.7.4011. [DOI] [PubMed] [Google Scholar]
- 31.Kalinski P, Giermasz A, Nakamura Y, et al. Helper role of NK cells during the induction of anticancer responses by dendritic cells. Mol Immunol. 2005;42:535–539. doi: 10.1016/j.molimm.2004.07.038. [DOI] [PubMed] [Google Scholar]
- 32.Vivier E, Tomasello E, Baratin M, et al. Functions of natural killer cells. Nat Immunol. 2008;9:503–510. doi: 10.1038/ni1582. [DOI] [PubMed] [Google Scholar]
- 33.Vivier E, Raulet DH, Moretta A, et al. Innate or adaptive immunity? The example of natural killer cells. Science. 2011;331:44–49. doi: 10.1126/science.1198687. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Zitvogel L. Dendritic and natural killer cells cooperate in the control/switch of innate immunity. J Exp Med. 2002;195:F9–F14. doi: 10.1084/jem.20012040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Degli-Esposti MA, Smyth MJ. Close encounters of different kinds: Dendritic cells and NK cells take centre stage. Nat Rev Immunol. 2005;5:112–124. doi: 10.1038/nri1549. [DOI] [PubMed] [Google Scholar]
- 36.Ferlazzo G, Mn̈z C. Dendritic cell interactions with NK cells from different tissues. J Clin Immunol. 2009;29:265–273. doi: 10.1007/s10875-009-9283-y. [DOI] [PubMed] [Google Scholar]
- 37.Strowig T, Brilot F, Mn̈z C. Noncytotoxic functions of NK cells: Direct pathogen restriction and assistance to adaptive immunity. J Immunol. 2008;180:7785–7791. doi: 10.4049/jimmunol.180.12.7785. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Waldmann TA. The biology of interleukin-2 and interleukin-15: Implications for cancer therapy and vaccine design. Nat Rev Immunol. 2006;6:595–601. doi: 10.1038/nri1901. [DOI] [PubMed] [Google Scholar]
- 39.Lee SC, Srivastava RM, López-Albaitero A, et al. Natural killer (NK): dendritic cell (DC) cross talk induced by therapeutic monoclonal antibody triggers tumor antigen-specific T cell immunity. Immunol Res. 2011;50:248–254. doi: 10.1007/s12026-011-8231-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Hussain M, Javeed A, Ashraf M, et al. Non-steroidal anti-inflammatory drugs, tumour immunity and immunotherapy. Pharmacol Res. 2012;66:7–18. doi: 10.1016/j.phrs.2012.02.003. [DOI] [PubMed] [Google Scholar]
- 41.Kb̈ler K, tho Pesch C, Gehrke N, et al. Immunogenic cell death of human ovarian cancer cells induced by cytosolic poly(I:C) leads to myeloid cell maturation and activates NK cells. Eur J Immunol. 2011;41:3028–3039. doi: 10.1002/eji.201141555. [DOI] [PubMed] [Google Scholar]
- 42.Lion E, Anguille S, Berneman ZN, et al. Poly(I:C) enhances the susceptibility of leukemic cells to NK cell cytotoxicity and phagocytosis by DC. PLoS One. 2011;6:e20952. doi: 10.1371/journal.pone.0020952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Lion E, Smits EL, Berneman ZN, et al. Acute myeloid leukemic cell lines loaded with synthetic dsRNA trigger IFN-gamma secretion by human NK cells. Leuk Res. 2009;33:539–546. doi: 10.1016/j.leukres.2008.08.020. [DOI] [PubMed] [Google Scholar]
- 44.Smits EL, Ponsaerts P, Van de Velde AL, et al. Proinflammatory response of human leukemic cells to dsRNA transfection linked to activation of dendritic cells. Leukemia. 2007;21:1691–1699. doi: 10.1038/sj.leu.2404763. [DOI] [PubMed] [Google Scholar]
- 45.Benjamin JE, Gill S, Negrin RS. Biology and clinical effects of natural killer cells in allogeneic transplantation. Curr Opin Oncol. 2010;22:130–137. doi: 10.1097/CCO.0b013e328335a559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Passweg JR, Koehl U, Uharek L, et al. Natural-killer-cell-based treatment in haematopoietic stem-cell transplantation. Best Pract Res Clin Haematol. 2006;19:811–824. doi: 10.1016/j.beha.2006.06.004. [DOI] [PubMed] [Google Scholar]
- 47.Ruggeri L, Aversa F, Martelli MF, et al. Allogeneic hematopoietic transplantation and natural killer cell recognition of missing self. Immunol Rev. 2006;214:202–218. doi: 10.1111/j.1600-065X.2006.00455.x. [DOI] [PubMed] [Google Scholar]
- 48.Terme M, Ullrich E, Delahaye NF, et al. Natural killer cell-directed therapies: Moving from unexpected results to successful strategies. Nat Immunol. 2008;9:486–494. doi: 10.1038/ni1580. [DOI] [PubMed] [Google Scholar]
- 49.Vivier E, Ugolini S, Blaise D, et al. Targeting natural killer cells and natural killer T cells in cancer. Nat Rev Immunol. 2012;12:239–252. doi: 10.1038/nri3174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Woo CY, Clay TM, Lyerly HK, et al. Role of natural killer cell function in dendritic cell-based vaccines. Expert Rev Vaccines. 2006;5:55–65. doi: 10.1586/14760584.5.1.55. [DOI] [PubMed] [Google Scholar]
- 51.Kalinski P, Urban J, Narang R, et al. Dendritic cell-based therapeutic cancer vaccines: What we have and what we need. Future Oncol. 2009;5:379–390. doi: 10.2217/FON.09.6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Figdor CG, de Vries IJ, Lesterhuis WJ, et al. Dendritic cell immunotherapy: Mapping the way. Nat Med. 2004;10:475–480. doi: 10.1038/nm1039. [DOI] [PubMed] [Google Scholar]
- 53.Smits EL, Anguille S, Cools N, et al. Dendritic cell-based cancer gene therapy. Hum Gene Ther. 2009;20:1106–1118. doi: 10.1089/hum.2009.145. [DOI] [PubMed] [Google Scholar]
- 54.Boudreau JE, Bonehill A, Thielemans K, et al. Engineering dendritic cells to enhance cancer immunotherapy. Mol Ther. 2011;19:841–853. doi: 10.1038/mt.2011.57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Rosenberg SA, Yang JC, Restifo NP. Cancer immunotherapy: Moving beyond current vaccines. Nat Med. 2004;10:909–915. doi: 10.1038/nm1100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Escudier B, Dorval T, Chaput N, et al. Vaccination of metastatic melanoma patients with autologous dendritic cell (DC) derived-exosomes: Results of the first phase I clinical trial. J Transl Med. 2005;3:10. doi: 10.1186/1479-5876-3-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Morse MA, Clay TM, Hobeika AC, et al. Phase I study of immunization with dendritic cells modified with fowlpox encoding carcinoembryonic antigen and costimulatory molecules. Clin Cancer Res. 2005;11:3017–3024. doi: 10.1158/1078-0432.CCR-04-2172. [DOI] [PubMed] [Google Scholar]
- 58.Fernandez NC, Lozier A, Flament C, et al. Dendritic cells directly trigger NK cell functions: Cross-talk relevant in innate anti-tumor immune responses in vivo. Nat Med. 1999;5:405–411. doi: 10.1038/7403. [DOI] [PubMed] [Google Scholar]
- 59.Cayeux S, Richter G, Becker C, et al. Direct and indirect T cell priming by dendritic cell vaccines. Eur J Immunol. 1999;29:225–234. doi: 10.1002/(SICI)1521-4141(199901)29:01<225::AID-IMMU225>3.0.CO;2-W. [DOI] [PubMed] [Google Scholar]
- 60.Anguille S, Lion E, Smits E, et al. Dendritic cell vaccine therapy for acute myeloid leukemia: Questions and answers. Hum Vaccin. 2011;7:579–584. doi: 10.4161/hv.7.5.14652. [DOI] [PubMed] [Google Scholar]
- 61.Ferlazzo G, Pack M, Thomas D, et al. Distinct roles of IL-12 and IL-15 in human natural killer cell activation by dendritic cells from secondary lymphoid organs. Proc Natl Acad Sci U S A. 2004;101:16606–16611. doi: 10.1073/pnas.0407522101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Shurin MR, Gregory M, Morris JC, et al. Genetically modified dendritic cells in cancer immunotherapy: A better tomorrow? Expert Opin Biol Ther. 2010;10:1539–1553. doi: 10.1517/14712598.2010.526105. [DOI] [PubMed] [Google Scholar]
- 63.Wan Y, Bramson J, Pilon A, et al. Genetically modified dendritic cells prime autoreactive T cells through a pathway independent of CD40L and interleukin 12: Implications for cancer vaccines. Cancer Res. 2000;60:3247–3253. [PubMed] [Google Scholar]
- 64.Ribas A, Wargo JA, Comin-Anduix B, et al. Enhanced tumor responses to dendritic cells in the absence of CD8-positive cells. J Immunol. 2004;172:4762–4769. doi: 10.4049/jimmunol.172.8.4762. [DOI] [PubMed] [Google Scholar]
- 65.Wargo JA, Schumacher LY, Comin-Anduix B, et al. Natural killer cells play a critical role in the immune response following immunization with melanoma-antigen-engineered dendritic cells. Cancer Gene Ther. 2005;12:516–527. doi: 10.1038/sj.cgt.7700818. [DOI] [PubMed] [Google Scholar]
- 66.Boudreau JE, Bridle BW, Stephenson KB, et al. Recombinant vesicular stomatitis virus transduction of dendritic cells enhances their ability to prime innate and adaptive antitumor immunity. Mol Ther. 2009;17:1465–1472. doi: 10.1038/mt.2009.95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Kim A, Noh YW, Kim KD, et al. Activated natural killer cell-mediated immunity is required for the inhibition of tumor metastasis by dendritic cell vaccination. Exp Mol Med. 2004;36:428–443. doi: 10.1038/emm.2004.55. [DOI] [PubMed] [Google Scholar]
- 68.Delluc S, Hachem P, Rusakiewicz S, et al. Dramatic efficacy improvement of a DC-based vaccine against AML by CD25 T cell depletion allowing the induction of a long-lasting T cell response. Cancer Immunol Immunother. 2009;58:1669–1677. doi: 10.1007/s00262-009-0678-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Shinagawa N, Yamazaki K, Tamura Y, et al. Immunotherapy with dendritic cells pulsed with tumor-derived gp96 against murine lung cancer is effective through immune response of CD8+ cytotoxic T lymphocytes and natural killer cells. Cancer Immunol Immunother. 2008;57:165–174. doi: 10.1007/s00262-007-0359-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Kim KD, Choi SC, Kim A, et al. Dendritic cell-tumor coculturing vaccine can induce antitumor immunity through both NK and CTL interaction. Int Immunopharmacol. 2001;1:2117–2129. doi: 10.1016/s1567-5769(01)00137-0. [DOI] [PubMed] [Google Scholar]
- 71.Kim KD, Kim JK, Kim SJ, et al. Protective antitumor activity through dendritic cell immunization is mediated by NK cell as well as CTL activation. Arch Pharm Res. 1999;22:340–347. doi: 10.1007/BF02979055. [DOI] [PubMed] [Google Scholar]
- 72.Nakamura M, Kikuchi T, Kufe DW, et al. Antitumor effects of fusions composed of dendritic cells and fibroblasts transfected with genomic DNA from tumor cells. Cancer Immunol Immunother. 2004;53:690–696. doi: 10.1007/s00262-004-0511-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Adam C, King S, Allgeier T, et al. DC-NK cell cross talk as a novel CD4+ T-cell-independent pathway for antitumor CTL induction. Blood. 2005;106:338–344. doi: 10.1182/blood-2004-09-3775. [DOI] [PubMed] [Google Scholar]
- 74.Karimi K, Boudreau JE, Fraser K, et al. Enhanced antitumor immunity elicited by dendritic cell vaccines is a result of their ability to engage both CTL and IFN gamma-producing NK cells. Mol Ther. 2008;16:411–418. doi: 10.1038/sj.mt.6300347. [DOI] [PubMed] [Google Scholar]
- 75.Liu C, Lou Y, Lizée G, et al. Plasmacytoid dendritic cells induce NK cell-dependent, tumor antigen-specific T cell cross-priming and tumor regression in mice. J Clin Invest. 2008;118:1165–1175. doi: 10.1172/JCI33583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Nagasaki E, Takahara A, Koido S, et al. Combined treatment with dendritic cells and 5-fluorouracil elicits augmented NK cell-mediated antitumor activity through the tumor necrosis factor-alpha pathway. J Immunother. 2010;33:467–474. doi: 10.1097/CJI.0b013e3181d36726. [DOI] [PubMed] [Google Scholar]
- 77.Prins RM, Vo DD, Khan-Farooqi H, et al. NK and CD4 cells collaborate to protect against melanoma tumor formation in the brain. J Immunol. 2006;177:8448–8455. doi: 10.4049/jimmunol.177.12.8448. [DOI] [PubMed] [Google Scholar]
- 78.Shimizu K, Fujii S. DC therapy induces long-term NK reactivity to tumors via host DC. Eur J Immunol. 2009;39:457–468. doi: 10.1002/eji.200838794. [DOI] [PubMed] [Google Scholar]
- 79.Tatsumi T, Takehara T, Yamaguchi S, et al. Intrahepatic delivery of alpha-galactosylceramide-pulsed dendritic cells suppresses liver tumor. Hepatology. 2007;45:22–30. doi: 10.1002/hep.21447. [DOI] [PubMed] [Google Scholar]
- 80.van den Broeke LT, Daschbach E, Thomas EK, et al. Dendritic cell-induced activation of adaptive and innate antitumor immunity. J Immunol. 2003;171:5842–5852. doi: 10.4049/jimmunol.171.11.5842. [DOI] [PubMed] [Google Scholar]
- 81.Viaud S, Terme M, Flament C, et al. Dendritic cell-derived exosomes promote natural killer cell activation and proliferation: A role for NKG2D ligands and IL-15Rα. PLoS One. 2009;4:e4942. doi: 10.1371/journal.pone.0004942. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Komaru A, Ueda Y, Furuya A, et al. Sustained and NK/CD4+ T cell-dependent efficient prevention of lung metastasis induced by dendritic cells harboring recombinant sendai virus. J Immunol. 2009;183:4211–4219. doi: 10.4049/jimmunol.0803845. [DOI] [PubMed] [Google Scholar]
- 83.Shimizu K, Asakura M, Fujii S. Prolonged antitumor NK cell reactivity elicited by CXCL10-expressing dendritic cells licensed by CD40L+ CD4+ memory T cells. J Immunol. 2011;186:5927–5937. doi: 10.4049/jimmunol.1003351. [DOI] [PubMed] [Google Scholar]
- 84.Boudreau JE, Stephenson KB, Wang F, et al. IL-15 and type I interferon are required for activation of tumoricidal NK cells by virus-infected dendritic cells. Cancer Res. 2011;71:2497–2506. doi: 10.1158/0008-5472.CAN-10-3025. [DOI] [PubMed] [Google Scholar]
- 85.Miller G, Lahrs S, Pillarisetty VG, et al. Adenovirus infection enhances dendritic cell immunostimulatory properties and induces natural killer and T-cell-mediated tumor protection. Cancer Res. 2002;62:5260–5266. [PubMed] [Google Scholar]
- 86.Tatsumi T, Takehara T, Yamaguchi S, et al. Injection of IL-12 gene-transduced dendritic cells into mouse liver tumor lesions activates both innate and acquired immunity. Gene Ther. 2007;14:863–871. doi: 10.1038/sj.gt.3302941. [DOI] [PubMed] [Google Scholar]
- 87.Miller G, Lahrs S, Dematteo RP. Overexpression of interleukin-12 enables dendritic cells to activate NK cells and confer systemic antitumor immunity. FASEB Journal. 2003;17:728–730. doi: 10.1096/fj.02-0900fje. [DOI] [PubMed] [Google Scholar]
- 88.Rodríguez-Calvillo M, Duarte M, Tirapu I, et al. Upregulation of natural killer cells functions underlies the efficacy of intratumorally injected dendritic cells engineered to produce interleukin-12. Exp Hematol. 2002;30:195–204. doi: 10.1016/s0301-472x(01)00792-5. [DOI] [PubMed] [Google Scholar]
- 89.Vera M, Razquin N, Prieto J, et al. Intratumoral injection of dendritic cells transduced by an SV40-based vector expressing interleukin-15 induces curative immunity mediated by CD8+ T lymphocytes and NK cells. Mol Ther. 2005;12:950–959. doi: 10.1016/j.ymthe.2005.03.030. [DOI] [PubMed] [Google Scholar]
- 90.Mazzolini G, Alfaro C, Sangro B, et al. Intratumoral injection of dendritic cells engineered to secrete interleukin-12 by recombinant adenovirus in patients with metastatic gastrointestinal carcinomas. J Clin Oncol. 2005;23:999–1010. doi: 10.1200/JCO.2005.00.463. [DOI] [PubMed] [Google Scholar]
- 91.Hayakawa Y, Screpanti V, Yagita H, et al. NK cell TRAIL eliminates immature dendritic cells in vivo and limits dendritic cell vaccination efficacy. J Immunol. 2004;172:123–129. doi: 10.4049/jimmunol.172.1.123. [DOI] [PubMed] [Google Scholar]
- 92.Osada T, Clay T, Hobeika A, et al. NK cell activation by dendritic cell vaccine: A mechanism of action for clinical activity. Cancer Immunol Immunother. 2006;55:1122–1131. doi: 10.1007/s00262-005-0089-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Butterfield LH, Comin-Anduix B, Vujanovic L, et al. Adenovirus MART-1-engineered autologous dendritic cell vaccine for metastatic melanoma. J Immunother. 2008;31:294–309. doi: 10.1097/CJI.0b013e31816a8910. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Bray SM, Vujanovic L, Butterfield LH. Dendritic cell-based vaccines positively impact natural killer and regulatory T cells in hepatocellular carcinoma patients. Clin Dev Immunol. 2011;2011:249281. doi: 10.1155/2011/249281. doi: 10.1155/2011/249281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Sakakibara M, Kanto T, Hayakawa M, et al. Comprehensive immunological analyses of colorectal cancer patients in the phase I/II study of quickly matured dendritic cell vaccine pulsed with carcinoembryonic antigen peptide. Cancer Immunol Immunother. 2011;60:1565–1575. doi: 10.1007/s00262-011-1051-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Yu JS, Wheeler CJ, Zeltzer PM, et al. Vaccination of malignant glioma patients with peptide-pulsed dendritic cells elicits systemic cytotoxicity and intracranial T-cell infiltration. Cancer Res. 2001;61:842–847. [PubMed] [Google Scholar]
- 97.Van Tendeloo VF, Van de Velde A, Van Driessche A, et al. Induction of complete and molecular remissions in acute myeloid leukemia by Wilms' tumor 1 antigen-targeted dendritic cell vaccination. Proc Natl Acad Sci U S A. 2010;107:13824–13829. doi: 10.1073/pnas.1008051107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Di Nicola M, Zappasodi R, Carlo-Stella C, et al. Vaccination with autologous tumor-loaded dendritic cells induces clinical and immunologic responses in indolent B-cell lymphoma patients with relapsed and measurable disease: A pilot study. Blood. 2009;113:18–27. doi: 10.1182/blood-2008-06-165654. [DOI] [PubMed] [Google Scholar]
- 99.Qi CJ, Ning YL, Han YS, et al. Autologous dendritic cell vaccine for estrogen receptor (ER)/progestin receptor (PR) double-negative breast cancer. Cancer Immunol Immunother. 2012 Jan 31; doi: 10.1007/s00262-011-1192-2. [Epub ahead or print]. doi: 10.1007/s00262-011-1192-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Alfaro C, Perez-Gracia JL, Suarez N, et al. Pilot clinical trial of type 1 dendritic cells loaded with autologous tumor lysates combined with GM-CSF, pegylated IFN, and cyclophosphamide for metastatic cancer patients. J Immunol. 2011;187:6130–6142. doi: 10.4049/jimmunol.1102209. [DOI] [PubMed] [Google Scholar]
- 101.Baek S, Kim CS, Kim SB, et al. Combination therapy of renal cell carcinoma or breast cancer patients with dendritic cell vaccine and IL-2: Results from a phase I/II trial. J Transl Med. 2011;9:178. doi: 10.1186/1479-5876-9-178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Chaput N, Flament C, Viaud S, et al. Dendritic cell derived-exosomes: Biology and clinical implementations. J Leukoc Biol. 2006;80:471–478. doi: 10.1189/jlb.0206094. [DOI] [PubMed] [Google Scholar]
- 103.Chaput N, Taieb J, André F, et al. The potential of exosomes in immunotherapy. Expert Opin Biol Ther. 2005;5:737–747. doi: 10.1517/14712598.5.6.737. [DOI] [PubMed] [Google Scholar]
- 104.Morse MA, Garst J, Osada T, et al. A phase I study of dexosome immunotherapy in patients with advanced non-small cell lung cancer. J Transl Med. 2005;3:9. doi: 10.1186/1479-5876-3-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Chang DH, Osman K, Connolly J, et al. Sustained expansion of NKT cells and antigen-specific T cells after injection of α-galactosyl-ceramide loaded mature dendritic cells in cancer patients. J Exp Med. 2005;201:1503–1517. doi: 10.1084/jem.20042592. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Nicol AJ, Tazbirkova A, Nieda M. Comparison of clinical and immunological effects of intravenous and intradermal administration of α-galactosylceramide (KRN7000)-pulsed dendritic cells. Clin Cancer Res. 2011;17:5140–5151. doi: 10.1158/1078-0432.CCR-10-3105. [DOI] [PubMed] [Google Scholar]
- 107.Nieda M, Okai M, Tazbirkova A, et al. Therapeutic activation of Vα24+Vβ11+ NKT cells in human subjects results in highly coordinated secondary activation of acquired and innate immunity. Blood. 2004;103:383–389. doi: 10.1182/blood-2003-04-1155. [DOI] [PubMed] [Google Scholar]
- 108.Martin-Fontecha A, Thomsen LL, Brett S, et al. Induced recruitment of NK cells to lymph nodes provides IFN-γ for T(H)1 priming. Nat Immunol. 2004;5:1260–1265. doi: 10.1038/ni1138. [DOI] [PubMed] [Google Scholar]
- 109.Kalinski P. Dendritic cells in immunotherapy of established cancer: Roles of signals 1, 2, 3 and 4. Curr Opin Investig Drugs. 2009;10:526–535. [PMC free article] [PubMed] [Google Scholar]
- 110.Gerosa F, Gobbi A, Zorzi P, et al. The reciprocal interaction of NK cells with plasmacytoid or myeloid dendritic cells profoundly affects innate resistance functions. J Immunol. 2005;174:727–734. doi: 10.4049/jimmunol.174.2.727. [DOI] [PubMed] [Google Scholar]
- 111.Münz C, Dao T, Ferlazzo G, et al. Mature myeloid dendritic cell subsets have distinct roles for activation and viability of circulating human natural killer cells. Blood. 2005;105:266–273. doi: 10.1182/blood-2004-06-2492. [DOI] [PubMed] [Google Scholar]
- 112.Gustafsson K, Ingelsten M, Bergqvist L, et al. Recruitment and activation of natural killer cells in vitro by a human dendritic cell vaccine. Cancer Res. 2008;68:5965–5971. doi: 10.1158/0008-5472.CAN-07-6494. [DOI] [PubMed] [Google Scholar]
- 113.Gustafsson K, Junevik K, Werlenius O, et al. Tumour-loaded α-type 1-polarized dendritic cells from patients with chronic lymphocytic leukaemia produce a superior NK-, NKT- and CD8(+) T cell-attracting chemokine profile. Scand J Immunol. 2011;74:318–326. doi: 10.1111/j.1365-3083.2011.02580.x. [DOI] [PubMed] [Google Scholar]
- 114.Jinushi M, Takehara T, Kanto T, et al. Critical role of MHC class I-related chain A and B expression on IFN-α-stimulated dendritic cells in NK cell activation: Impairment in chronic hepatitis C virus infection. J Immunol. 2003;170:1249–1256. doi: 10.4049/jimmunol.170.3.1249. [DOI] [PubMed] [Google Scholar]
- 115.Sakakibara M, Kanto T, Inoue M, et al. Quick generation of fully mature dendritic cells from monocytes with OK432, low-dose prostanoid, and interferon-α as potent immune enhancers. J Immunother. 2006;29:67–77. doi: 10.1097/01.cji.0000183093.77687.46. [DOI] [PubMed] [Google Scholar]
- 116.Tosi D, Valenti R, Cova A, et al. Role of cross-talk between IFN-α-induced monocyte-derived dendritic cells and NK cells in priming CD8+ T cell responses against human tumor antigens. J Immunol. 2004;172:5363–5370. doi: 10.4049/jimmunol.172.9.5363. [DOI] [PubMed] [Google Scholar]
- 117.Anguille S, Smits EL, Cools N, et al. Short-term cultured, interleukin-15 differentiated dendritic cells have potent immunostimulatory properties. J Transl Med. 2009;7:109. doi: 10.1186/1479-5876-7-109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Dubsky P, Saito H, Leogier M, et al. IL-15-induced human DC efficiently prime melanoma-specific naive CD8+ T cells to differentiate into CTL. Eur J Immunol. 2007;37:1678–1690. doi: 10.1002/eji.200636329. [DOI] [PubMed] [Google Scholar]
- 119.Hardy MY, Kassianos AJ, Vulink A, et al. NK cells enhance the induction of CTL responses by IL-15 monocyte-derived dendritic cells. Immunol Cell Biol. 2009;87:606–614. doi: 10.1038/icb.2009.44. [DOI] [PubMed] [Google Scholar]
- 120.Beck B, Dörfel D, Lichtenegger FS, et al. Effects of TLR agonists on maturation and function of 3-day dendritic cells from AML patients in complete remission. J Transl Med. 2011;9:151. doi: 10.1186/1479-5876-9-151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Smits EL, Cools N, Lion E, et al. The toll-like receptor 7/8 agonist resiquimod greatly increases the immunostimulatory capacity of human acute myeloid leukemia cells. Cancer Immunol Immunother. 2010;59:35–46. doi: 10.1007/s00262-009-0721-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Spranger S, Javorovic M, Br̈dek M, et al. Generation of Th1-polarizing dendritic cells using the TLR7/8 agonist CL075. J Immunol. 2010;185:738–747. doi: 10.4049/jimmunol.1000060. [DOI] [PubMed] [Google Scholar]
- 123.Van Elssen CH, Vanderlocht J, Frings PW, et al. Klebsiella pneumoniae-triggered DC recruit human NK cells in a CCR5-dependent manner leading to increased CCL19-responsiveness and activation of NK cells. Eur J Immunol. 2010;40:3138–3149. doi: 10.1002/eji.201040496. [DOI] [PubMed] [Google Scholar]
- 124.Vujanovic L, Szymkowski DE, Alber S, et al. Virally infected and matured human dendritic cells activate natural killer cells via cooperative activity of plasma membrane-bound TNF and IL-15. Blood. 2010;116:575–583. doi: 10.1182/blood-2009-08-240325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Ferlazzo G, Tsang ML, Moretta L, et al. Human dendritic cells activate resting natural killer (NK) cells and are recognized via the NKp30 receptor by activated NK cells. J Exp Med. 2002;195:343–351. doi: 10.1084/jem.20011149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Dao T, Gomez-Nunez M, Antczak C, et al. Natural killer cells license dendritic cell cross-presentation of B lymphoma cell–associated antigens. Clin Cancer Res. 2005;11:8763–8772. doi: 10.1158/1078-0432.CCR-05-0975. [DOI] [PubMed] [Google Scholar]
- 127.Schnurr M, Scholz C, Rothenfusser S, et al. Apoptotic pancreatic tumor cells are superior to cell lysates in promoting cross-priming of cytotoxic T cells and activate NK and γδ T cells. Cancer Res. 2002;62:2347–2352. [PubMed] [Google Scholar]
- 128.Ferlazzo G, Mn̈z C. NK cell compartments and their activation by dendritic cells. J Immunol. 2004;172:1333–1339. doi: 10.4049/jimmunol.172.3.1333. [DOI] [PubMed] [Google Scholar]
- 129.Zanoni I, Foti M, Ricciardi-Castagnoli P, et al. TLR-dependent activation stimuli associated with Th1 responses confer NK cell stimulatory capacity to mouse dendritic cells. J Immunol. 2005;175:286–292. doi: 10.4049/jimmunol.175.1.286. [DOI] [PubMed] [Google Scholar]
- 130.Prestwich RJ, Errington F, Steele LP, et al. Reciprocal human dendritic cell-natural killer cell interactions induce antitumor activity following tumor cell infection by oncolytic reovirus. J Immunol. 2009;183:4312–4321. doi: 10.4049/jimmunol.0901074. [DOI] [PubMed] [Google Scholar]
- 131.Bontkes HJ, Kramer D, Ruizendaal JJ, et al. Tumor associated antigen and interleukin-12 mRNA transfected dendritic cells enhance effector function of natural killer cells and antigen specific T-cells. Clin Immunol. 2008;127:375–384. doi: 10.1016/j.clim.2008.02.001. [DOI] [PubMed] [Google Scholar]
- 132.Pham TN, Hong CY, Min JJ, et al. Enhancement of antitumor effect using dendritic cells activated with natural killer cells in the presence of toll-like receptor agonist. Exp Mol Med. 2010;42:407–419. doi: 10.3858/emm.2010.42.6.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Nguyen-Pham TN, Im CM, Nguyen TA, et al. Induction of myeloma-specific cytotoxic T lymphocytes responses by natural killer cells stimulated-dendritic cells in patients with multiple myeloma. Leuk Res. 2011;35:1241–1247. doi: 10.1016/j.leukres.2011.03.032. [DOI] [PubMed] [Google Scholar]
- 134.Nguyen-Pham TN, Yang DH, Nguyen TA, et al. Optimal culture conditions for the generation of natural killer cell-induced dendritic cells for cancer immunotherapy. Cell Mol Immunol. 2012;9:45–53. doi: 10.1038/cmi.2011.23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Wong JL, Mailliard RB, Moschos SJ, et al. Helper activity of natural killer cells during the dendritic cell-mediated induction of melanoma-specific cytotoxic T cells. J Immunother. 2011;34:270–278. doi: 10.1097/CJI.0b013e31820b370b. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Pittari G, Fregni G, Roguet L, et al. Early evaluation of natural killer activity in post-transplant acute myeloid leukemia patients. Bone Marrow Transplant. 2010;45:862–871. doi: 10.1038/bmt.2009.265. [DOI] [PubMed] [Google Scholar]
- 137.Reittie JE, Gottlieb D, Heslop HE, et al. Endogenously generated activated killer cells circulate after autologous and allogeneic marrow transplantation but not after chemotherapy. Blood. 1989;73:1351–1358. [PubMed] [Google Scholar]
- 138.Savani BN, Mielke S, Adams S, et al. Rapid natural killer cell recovery determines outcome after T-cell-depleted HLA-identical stem cell transplantation in patients with myeloid leukemias but not with acute lymphoblastic leukemia. Leukemia. 2007;21:2145–2152. doi: 10.1038/sj.leu.2404892. [DOI] [PubMed] [Google Scholar]
- 139.Vitale C, Chiossone L, Morreale G, et al. Human natural killer cells undergoing in vivo differentiation after allogeneic bone marrow transplantation: Analysis of the surface expression and function of activating NK receptors. Mol Immunol. 2005;42:405–411. doi: 10.1016/j.molimm.2004.07.019. [DOI] [PubMed] [Google Scholar]
- 140.Brassard DL, Grace MJ, Bordens RW. Interferon-α as an immunotherapeutic protein. J Leukoc Biol. 2002;71:565–581. [PubMed] [Google Scholar]
- 141.Fehniger TA, Cooper MA, Caligiuri MA. Interleukin-2 and interleukin-15: Immunotherapy for cancer. Cytokine Growth Factor Rev. 2002;13:169–183. doi: 10.1016/s1359-6101(01)00021-1. [DOI] [PubMed] [Google Scholar]
- 142.Schleypen JS, Baur N, Kammerer R, et al. Cytotoxic markers and frequency predict functional capacity of natural killer cells infiltrating renal cell carcinoma. Clin Cancer Res. 2006;12:718–725. doi: 10.1158/1078-0432.CCR-05-0857. [DOI] [PubMed] [Google Scholar]
- 143.Menon AG, Janssen-van Rhijn CM, Morreau H, et al. Immune system and prognosis in colorectal cancer: A detailed immunohistochemical analysis. Lab Invest. 2004;84:493–501. doi: 10.1038/labinvest.3700055. [DOI] [PubMed] [Google Scholar]
- 144.Lowdell MW, Craston R, Samuel D, et al. Evidence that continued remission in patients treated for acute leukaemia is dependent upon autologous natural killer cells. Br J Haematol. 2002;117:821–827. doi: 10.1046/j.1365-2141.2002.03495.x. [DOI] [PubMed] [Google Scholar]
- 145.Mortier E, Woo T, Advincula R, et al. IL-15Rα chaperones IL-15 to stable dendritic cell membrane complexes that activate NK cells via trans presentation. J Exp Med. 2008;205:1213–1225. doi: 10.1084/jem.20071913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Ménard C, Blay JY, Borg C, et al. Natural killer cell IFN-γ levels predict long-term survival with imatinib mesylate therapy in gastrointestinal stromal tumor-bearing patients. Cancer Res. 2009;69:3563–3569. doi: 10.1158/0008-5472.CAN-08-3807. [DOI] [PubMed] [Google Scholar]
- 147.Farag SS. DC therapy? Let the natural killer answer. Blood. 2005;106:1892. [Google Scholar]
- 148.Pross HF, Maroun JA. The standardization of NK cell assays for use in studies of biological response modifiers. J Immunol Methods. 1984;68:235–249. doi: 10.1016/0022-1759(84)90154-6. [DOI] [PubMed] [Google Scholar]
- 149.Rusthoven JJ. Clinical significance of natural killer cell cytotoxicity: Need for proper data analysis in the design of clinical studies. Eur J Cancer Clin Oncol. 1985;21:1287–1293. doi: 10.1016/0277-5379(85)90305-0. [DOI] [PubMed] [Google Scholar]
- 150.Björkström NK, Fauriat C, Bryceson YT, et al. Analysis of the KIR repertoire in human NK cells by flow cytometry. Methods Mol Biol. 2010;612:353–364. doi: 10.1007/978-1-60761-362-6_24. [DOI] [PubMed] [Google Scholar]
- 151.Bryceson YT, Fauriat C, Nunes JM, et al. Functional analysis of human NK cells by flow cytometry. Methods Mol Biol. 2010;612:335–352. doi: 10.1007/978-1-60761-362-6_23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Clay TM, Hobeika AC, Mosca PJ, et al. Assays for monitoring cellular immune responses to active immunotherapy of cancer. Clin Cancer Res. 2001;7:1127–1135. [PubMed] [Google Scholar]
- 153.Lion E, Smits EL, Berneman ZN, et al. Quantification of IFN-γ produced by human purified NK cells following tumor cell stimulation: Comparison of three IFN-γ assays. J Immunol Methods. 2009;350:89–96. doi: 10.1016/j.jim.2009.08.014. [DOI] [PubMed] [Google Scholar]
- 154.Takanami I, Takeuchi K, Giga M. The prognostic value of natural killer cell infiltration in resected pulmonary adenocarcinoma. J Thorac Cardiovasc Surg. 2001;121:1058–1063. doi: 10.1067/mtc.2001.113026. [DOI] [PubMed] [Google Scholar]
- 155.Villegas FR, Coca S, Villarrubia VG, et al. Prognostic significance of tumor infiltrating natural killer cells subset CD57 in patients with squamous cell lung cancer. Lung Cancer. 2002;35:23–28. doi: 10.1016/s0169-5002(01)00292-6. [DOI] [PubMed] [Google Scholar]
- 156.Coca S, Perez-Piqueras J, Martinez D, et al. The prognostic significance of intratumoral natural killer cells in patients with colorectal carcinoma. Cancer. 1997;79:2320–2328. doi: 10.1002/(sici)1097-0142(19970615)79:12<2320::aid-cncr5>3.0.co;2-p. [DOI] [PubMed] [Google Scholar]
- 157.Maréchal R, De Schutter J, Nagy N, et al. Putative contribution of CD56 positive cells in cetuximab treatment efficacy in first-line metastatic colorectal cancer patients. BMC Cancer. 2010;10:340. doi: 10.1186/1471-2407-10-340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.McGilvray RW, Eagle RA, Watson NF, et al. NKG2D ligand expression in human colorectal cancer reveals associations with prognosis and evidence for immunoediting. Clin Cancer Res. 2009;15:6993–7002. doi: 10.1158/1078-0432.CCR-09-0991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Ishigami S, Natsugoe S, Tokuda K, et al. Clinical impact of intratumoral natural killer cell and dendritic cell infiltration in gastric cancer. Cancer Lett. 2000;159:103–108. doi: 10.1016/s0304-3835(00)00542-5. [DOI] [PubMed] [Google Scholar]
- 160.Belldegrun A, Tso CL, Kaboo R, et al. Natural immune reactivity-associated therapeutic response in patients with metastatic renal cell carcinoma receiving tumor-infiltrating lymphocytes and interleukin-2-based therapy. J Immunother Emphasis Tumor Immunol. 1996;19:149–161. doi: 10.1097/00002371-199603000-00008. [DOI] [PubMed] [Google Scholar]
- 161.González FM, Vargas JA, López-Cortijo C, et al. Prognostic significance of natural killer cell activity in patients with laryngeal carcinoma. Arch Otolaryngol Head Neck Surg. 1998;124:852–856. doi: 10.1001/archotol.124.8.852. [DOI] [PubMed] [Google Scholar]
- 162.Schantz SP, Ordonez NG. Quantitation of natural killer cell function and risk of metastatic poorly differentiated head and neck cancer. Nat Immun Cell Growth Regul. 1991;10:278–288. [PubMed] [Google Scholar]
- 163.Taketomi A, Shimada M, Shirabe K, et al. Natural killer cell activity in patients with hepatocellular carcinoma: A new prognostic indicator after hepatectomy. Cancer. 1998;83:58–63. doi: 10.1002/(sici)1097-0142(19980701)83:1<58::aid-cncr8>3.0.co;2-a. [DOI] [PubMed] [Google Scholar]
- 164.Lowdell MW, Ray N, Craston R, et al. The in vitro detection of anti-leukaemia-specific cytotoxicity after autologous bone marrow transplantation for acute leukaemia. Bone Marrow Transplant. 1997;19:891–897. doi: 10.1038/sj.bmt.1700756. [DOI] [PubMed] [Google Scholar]
- 165.Pizzolo G, Trentin L, Vinante F, et al. Natural killer cell function and lymphoid subpopulations in acute non-lymphoblastic leukaemia in complete remission. Br J Cancer. 1988;58:368–372. doi: 10.1038/bjc.1988.221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Pross HF, Lotzová E. Role of natural killer cells in cancer. Nat Immun. 1993;12:279–292. [PubMed] [Google Scholar]
- 167.Tajima F, Kawatani T, Endo A, et al. Natural killer cell activity and cytokine production as prognostic factors in adult acute leukemia. Leukemia. 1996;10:478–482. [PubMed] [Google Scholar]
- 168.Tratkiewicz JA, Szer J. Loss of natural killer activity as an indicator of relapse in acute leukaemia. Clin Exp Immunol. 1990;80:241–246. doi: 10.1111/j.1365-2249.1990.tb05241.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Fregni G, Perier A, Pittari G, et al. Unique functional status of natural killer cells in metastatic stage IV melanoma patients and its modulation by chemotherapy. Clin Cancer Res. 2011;17:2628–2637. doi: 10.1158/1078-0432.CCR-10-2084. [DOI] [PubMed] [Google Scholar]
- 170.Imai K, Matsuyama S, Miyake S, et al. Natural cytotoxic activity of peripheral-blood lymphocytes and cancer incidence: An 11-year follow-up study of a general population. Lancet. 2000;356:1795–1799. doi: 10.1016/S0140-6736(00)03231-1. [DOI] [PubMed] [Google Scholar]
- 171.Li Z, Qiao Y, Liu B, et al. Combination of imatinib mesylate with autologous leukocyte-derived heat shock protein and chronic myelogenous leukemia. Clin Cancer Res. 2005;11:4460–4468. doi: 10.1158/1078-0432.CCR-05-0250. [DOI] [PubMed] [Google Scholar]
- 172.Borg C, Terme M, Taieb J, et al. Novel mode of action of c-kit tyrosine kinase inhibitors leading to NK cell-dependent antitumor effects. J Clin Invest. 2004;114:379–388. doi: 10.1172/JCI21102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Epling-Burnette PK, Bai F, Painter JS, et al. Reduced natural killer (NK) function associated with high-risk myelodysplastic syndrome (MDS) and reduced expression of activating NK receptors. Blood. 2007;109:4816–4824. doi: 10.1182/blood-2006-07-035519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.El-Sherbiny YM, Meade JL, Holmes TD, et al. The requirement for DNAM-1, NKG2D, and NKp46 in the natural killer cell-mediated killing of myeloma cells. Cancer Res. 2007;67:8444–8449. doi: 10.1158/0008-5472.CAN-06-4230. [DOI] [PubMed] [Google Scholar]
- 175.Delahaye NF, Rusakiewicz S, Martins I, et al. Alternatively spliced NKp30 isoforms affect the prognosis of gastrointestinal stromal tumors. Nat Med. 2011;17:700–707. doi: 10.1038/nm.2366. [DOI] [PubMed] [Google Scholar]
- 176.Salih HR, Antropius H, Gieseke F, et al. Functional expression and release of ligands for the activating immunoreceptor NKG2D in leukemia. Blood. 2003;102:1389–1396. doi: 10.1182/blood-2003-01-0019. [DOI] [PubMed] [Google Scholar]
- 177.Salih HR, Goehlsdorf D, Steinle A. Release of MICB molecules by tumor cells: Mechanism and soluble MICB in sera of cancer patients. Hum Immunol. 2006;67:188–195. doi: 10.1016/j.humimm.2006.02.008. [DOI] [PubMed] [Google Scholar]
- 178.Rebmann V, Schẗt P, Brandhorst D, et al. Soluble MICA as an independent prognostic factor for the overall survival and progression-free survival of multiple myeloma patients. Clin Immunol. 2007;123:114–120. doi: 10.1016/j.clim.2006.11.007. [DOI] [PubMed] [Google Scholar]
- 179.Wu JD, Higgins LM, Steinle A, et al. Prevalent expression of the immunostimulatory MHC class I chain-related molecule is counteracted by shedding in prostate cancer. J Clin Invest. 2004;114:560–568. doi: 10.1172/JCI22206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Holdenrieder S, Stieber P, Peterfi A, et al. Soluble MICB in malignant diseases: Analysis of diagnostic significance and correlation with soluble MICA. Cancer Immunol Immunother. 2006;55:1584–1589. doi: 10.1007/s00262-006-0167-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Holdenrieder S, Stieber P, Peterfi A, et al. Soluble MICA in malignant diseases. Int J Cancer. 2006;118:684–687. doi: 10.1002/ijc.21382. [DOI] [PubMed] [Google Scholar]
- 182.Miller JS, Cooley S, Parham P, et al. Missing KIR ligands are associated with less relapse and increased graft-versus-host disease (GVHD) following unrelated donor allogeneic HCT. Blood. 2007;109:5058–5061. doi: 10.1182/blood-2007-01-065383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183.Dauguet N, Récher C, Demur C, et al. Pre-eminence and persistence of immature natural killer cells in acute myeloid leukemia patients in first complete remission. Am J Hematol. 2010;86:203–213. doi: 10.1002/ajh.21906. [DOI] [PubMed] [Google Scholar]
- 184.Grégoire C, Chasson L, Luci C, et al. The trafficking of natural killer cells. Immunol Rev. 2007;220:169–182. doi: 10.1111/j.1600-065X.2007.00563.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Waldhauer I, Steinle A. Proteolytic release of soluble UL16-binding protein 2 from tumor cells. Cancer Res. 2006;66:2520–2526. doi: 10.1158/0008-5472.CAN-05-2520. [DOI] [PubMed] [Google Scholar]