

Abstract
Although the occurrence of intracellular glasses in seeds and pollen has been established, physical properties such as rotational correlation times and viscosity have not been studied extensively. Using electron paramagnetic resonance spectroscopy, we examined changes in the molecular mobility of the hydrophilic nitroxide spin probe 3-carboxy-proxyl during melting of intracellular glasses in axes of pea (Pisum sativum L.) seeds and cattail (Typha latifolia L.) pollen. The rotational correlation time of the spin probe in intracellular glasses of both organisms was approximately 10−3 s. Using the distance between the outer extrema of the electron paramagnetic resonance spectrum (2Azz) as a measure of molecular mobility, we found a sharp increase in mobility at a definite temperature during heating. This temperature increased with decreasing water content of the samples. Differential scanning calorimetry data on these samples indicated that this sharp increase corresponded to melting of the glassy matrix. Molecular mobility was found to be inversely correlated with storage stability. With decreasing water content, the molecular mobility reached a minimum, and increased again at very low water content. Minimum mobility and maximum storage stability occurred at a similar water content. This correlation suggests that storage stability might be at least partially controlled by molecular mobility. At low temperatures, when storage longevity cannot be determined on a realistic time scale, 2Azz measurements can provide an estimate of the optimum storage conditions.
Deterioration of seeds and pollen during storage involves many physical and chemical changes, such as disrupted intracellular integrity, decreased activities of enzymes, lipid peroxidation and deesterification, and Maillard reactions (Priestley, 1986; Wilson and McDonald, 1986; Wettlaufer and Leopold, 1991; Van Bilsen and Hoekstra, 1993; Van Bilsen et al., 1994). Since the formation of glasses in dehydrating biological tissues has been established, this physical phenomenon has been put forward as a prominent factor in the control of deterioration rates during storage (Burke, 1986; Williams and Leopold, 1989; Leopold et al., 1994; Leprince and Walters-Vertucci, 1995; Buitink et al., 1996). A glass is a thermodynamically unstable solid-state with an extremely high viscosity (Franks et al., 1991). Its formation is promoted by low water content of tissues and by low temperatures. Both factors are also known to extend the longevity of seeds and pollen (Roberts, 1972; Roberts and Ellis, 1989; Vertucci and Roos, 1993; Buitink et al., 1998), and improved storage stability was observed when glasses were present (Sun and Leopold, 1994; Sun, 1997; Buitink et al., 1998). It is assumed that the high viscosity of intracellular glasses decreases molecular mobility and impedes diffusion within the cytoplasm, thus slowing down deleterious reactions and changes in structure and chemical composition during aging (Sun and Leopold, 1993; Sun, 1997). However, the molecular mobility and viscosity in biological glasses has received little attention.
Molecular mobility has been studied using EPR spectroscopy by labeling polymers and food materials with a suitable, stable spin probe (Steffen et al., 1992; Blackburn et al., 1996; Dzuba, 1996; Hemminga and Van den Dries, 1998). From the EPR spectra of the spin probe, τR can be assessed (Kumler and Boyer, 1976; Kovarskii et al., 1978; Spielberg and Gelerinter, 1982; Ohta and Kuwata, 1985; Roozen and Hemminga, 1990; Roozen et al., 1991; Dzuba et al., 1993). Whereas τR values of 10−12 to 10−9 s can be calculated from conventional EPR spectra (Knowles et al., 1976), ST-EPR spectroscopy further expands this range to very slow (10−6 – 10−3 s; Hemminga, 1983) and ultra slow (10−3 – 102 s; Van den Dries et al., 1998) molecular motions. ST-EPR spectroscopy has been successfully applied to determine τR values of spin probes in sugar glasses (Roozen et al., 1991; Van den Dries et al., 1998) and organic liquids at low temperatures (Ito, 1983). In glassy Suc-water and malto-oligosaccharide mixtures, τR decreases by several orders of magnitude upon approaching Tg (Roozen et al., 1991).
The present paper is aimed at gaining insight into changes in the molecular mobility that accompany glass formation in anhydrobiotes. We used EPR and ST-EPR spectroscopy to characterize the molecular motion of CP, the polar nitroxide spin probe that we incorporated into axes of pea (Pisum sativum L.) seeds and cattail (Typha latifolia L.) pollen. We show that the distance between the outer extrema in powder spectra produced by conventional EPR spectroscopy can be used to detect changes in molecular motion, and that ST-EPR spectroscopy allows an estimation of τR. We discuss the possible relationship between glasses, molecular mobility, and storage stability in these organisms.
MATERIALS AND METHODS
Plant Material and Treatments
Mature male inflorescences of cattail (Typha latifolia L.) were collected from field populations near Wageningen, The Netherlands, in 1996, and allowed to shed their pollen in the laboratory. Pollen (94% germination) was cleaned by sieving through a fine copper mesh, dried in dry air to 0.05 to 0.08 g water g−1 dry weight, and stored at −20°C until use. Pea (Pisum sativum cv Karina) seeds (99% germination) were obtained from Nunhems Zaden (Haelen, The Netherlands) and stored at 5°C until use.
The polar nitroxide spin probe CP (Sigma) was used for spin labeling of pea seeds and cattail pollen. Pollen (3 g) was prehydrated in water vapor for 16 h at 5°C to about 0.7 g water g−1 dry weight, and then mixed at 25°C with 6 mL of liquid germination medium containing 2.5 mm CP. The germination medium consisted of 1.6 mm H3BO3, 1.3 mm Ca(NO3)2·4H2O, 0.8 mm MgSO4·7H2O, 1.0 mm KNO3, and 0.2 m Suc in 2 mm sodium-phosphate-citrate buffer, pH 5.9. After a few minutes, an additional 20 mL of the germination medium was added, and the pollen was recovered by filtration. The pollen was then mixed with 20 mL of a solution of 1 mm CP and 120 mm of the broadening agent potassium ferricyanide. Ferricyanide broadens spin-probe signals in the solution surrounding the cells to invisibility (Golovina and Tikhonov, 1994; Golovina et al., 1997). After 5 more min, the pollen was recovered by filtration, spread out in a large Petri dish, and rapidly dried in a flow of dry air (3% RH) in a drying box to a water content of less than 0.05 g water g−1 dry weight.
The pollen retained high viability after labeling and drying. To determine the τR at high water content (1.5–0.5 g water g−1 dry weight), pollen samples were taken regularly during drying for EPR measurements. After drying, the pollen was stored over various saturated salt solutions or P2O5 (Winston and Bates, 1960) at 25°C for at least 3 d to obtain the various water contents.
Pea seeds were rolled in germination paper and soaked in tap water overnight at 15°C, and then axes were isolated. Subsequently, the axes were incubated in a solution containing 1 mm CP and 200 mm ferricyanide at room temperature. After 45 min, the axes were rinsed with distilled water and dried at 35% RH for 3 d. To determine the τR at high water content (1.5–0.5 g water g−1 dry weight), pea axes were regularly sampled during drying for EPR measurements. After drying, the axes were stored over the saturated salt solutions or P2O5 at 25°C to obtain the various water contents.
With every sample taken for EPR measurements, a sample treated similarly was taken for determination of water content. Water contents were analyzed by weighing the samples before and after heating at 96°C for 36 to 48 h (Buitink et al., 1996) and calculating the water loss on a dry-weight basis.
EPR and ST-EPR Measurements
EPR spectra were recorded with an X-band EPR spectrometer (model 300E, Bruker Analytik, Rheinstetten, Germany). Microwave power was kept low (200 μW or 2 mW) to avoid saturation. Modulation amplitude was 0.4 G for pea axes and 1 G for cattail pollen.
Samples with various water contents were loaded into a 3-mm-diameter EPR capillary. For each measurement, the capillary was filled for a length of 5 cm with pollen or with two isolated pea axes. To prevent water loss during the measurements, the capillaries were sealed at both sides. Temperature was controlled using a controller with liquid nitrogen vapor as the coolant. Samples were rapidly cooled to −150°C and allowed to equilibrate for 30 min, after which scans were recorded at 10°C increments with equilibration for 5 min after each increment.
Conventional EPR spectroscopy can detect changes in τR of spin probes ranging from 10−12 to 10−9 s, which corresponds to the lifetime of the probe in a given orientation. In this motional range, the EPR spectrum of nitroxides consists of three lines (Fig. 1, top spectrum), and τR can be calculated according to the method of Knowles et al. (1976):
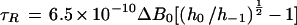 |
1 |
where h−1 and h0 are the heights of the high-field and central lines in the EPR spectra, respectively, and ΔB0 is the line width of the central line in Gauss. The rotational motion of the spin probe was assumed to be isotropic. The τR of partially hydrated (above approximately 0.25 g water g−1 dry weight) pollen and pea axes was determined using Equation 1.
Figure 1.

EPR spectra of CP in cattail pollen at various water contents (g/g, g water g−1 dry weight). Spectra were recorded at room temperature. The h+1, h0, and h−1 peaks are shown in the top spectrum. The distance between the two outer extrema, 2Azz, is indicated in the bottom spectrum.
The characteristic τR for spin probes in organic glasses near and above Tg is approximately 10−5 s or higher (Dzuba et al., 1984; Roozen et al., 1991; Van den Dries et al., 1998). These τR values cannot be determined using Equation 1, because the shapes of the lines in the spectra change due to the appearance of a powder spectrum (Fig. 1, below 0.25 g water g−1 dry weight). In these powder spectra the distance between the two outer extrema (2Azz) is temperature dependent for a number of glass-forming substances at and above their Tg (Kumler and Boyer, 1976; Kovarskii et al., 1978; Spielberg and Gelerinter, 1982; Ohta and Kuwata, 1985; Dzuba et al., 1993) (Fig. 1). We made use of this parameter to obtain a qualitative measurement of the molecular mobility of CP.
From the τR one can derive the viscosity of the matrix in which the spin probe is rotating, according to the modified Stokes-Einstein equation (Roozen et al., 1991):
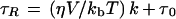 |
2 |
where η is the solvent viscosity, kb is Boltzmann's constant, V is the volume of the rotating molecule, T is the absolute temperature, τ0 is the zero viscosity τR, and k is a dimensionless slip parameter.
ST-EPR spectroscopy was used in the motional region for τR > 10−7 s. This method is based on the diffusion and recovery of saturation between different portions of the powder spectrum in competition with field modulation (Hemminga, 1983). For ST-EPR spectroscopic measurements, the second harmonic quadrature absorption signal was detected under the following conditions: a field-modulation amplitude of 5 G, microwave power of 100 mW, and field-modulation frequency of 50 kHz (Hemminga et al., 1984). The phase was set with the self-null method (Thomas et al., 1976).
In ST-EPR spectroscopy, τR values are usually obtained empirically using reference material with known viscosity. Here, spectra of CP in anhydrous glycerol were used to construct a calibration curve according to the method of Van den Dries et al. (1998). Because the viscosity for anhydrous glycerol is known over a broad temperature range, τR of CP in glycerol can be obtained from Equation 2. Spectra of CP in anhydrous glycerol were recorded every 3°C, and the values of the line-shape parameters L"/L′ and C′/C (explained in Fig. 5) were calculated for each temperature (data not shown). From the curves representing the line-shape parameters of CP in glycerol against τR, the τR values of CP in the axes and pollen were obtained by interpolation of the corresponding line-shape parameters.
Figure 5.

ST-EPR spectra of CP in pea axes at 0.08 g water g−1 dry weight recorded at various temperatures. Scans were recorded at 100 mW, a modulation frequency of 50 kHz, and a modulation amplitude of 5 G.
Differential Scanning Calorimetry
Pollen and pea axes with different water contents were hermetically sealed in aluminum pans for differential scanning calorimetry. Second-order transitions of the samples were determined using a differential scanning calorimeter (Pyris 1, Perkin-Elmer) calibrated for temperature with indium (156.6°C) and methylene chloride (−95°C) standards and for energy with indium (28.54 J g−1). Baselines were determined using an empty pan, and all thermograms were baseline corrected. Scans were taken from −100°C to 120°C at a rate of 10°C min−1. The Tg values were determined by the onset and midpoint of the temperature range over which the change in specific heat occurred. All analyses were performed with Perkin-Elmer software.
RESULTS
Molecular Motion in Pollen and Seeds
Figure 1 shows representative EPR spectra of CP in cattail pollen at different water contents recorded at room temperature. When the water content was decreased from 0.53 to 0.25 g water g−1 dry weight (top two spectra), the relative amplitudes of the outer spectral lines decreased and the width of the central peak increased. Using Equation 1, the τR of CP in hydrated pollen and pea axes (approximately 1.2 g water g−1 dry weight) was calculated to be approximately 10−11 s (data not shown). In the partially dehydrated state (0.53 g water g−1 dry weight), τR of CP in cattail pollen was calculated as 1.6 × 10−10 s (Fig. 1, top spectrum). At 0.25 g water g−1 dry weight (Fig. 1, second spectrum from the top), τR of CP in pollen was 7.9 × 10−10 s. The τR of CP in pea axes at 0.5 g water g−1 dry weight was 3.6 × 10−10 s (data not shown). At a water content below 0.25 g water g−1 dry weight, a powder spectrum (characterized by the two broad peaks at the extremes) overlapped the mobile spectrum (three sharp lines separated by a distance of 15 G). The bottom spectrum shown in Figure 1 has the characteristic shape of a powder spectrum, indicative of slow molecular mobility of the spin probe with τR >10−8 s. With the appearance of this powder spectrum below 0.25 g water g−1 dry weight, the τR of CP cannot be calculated using Equation 1, because the line shapes are distorted. The distance between the two broad peaks at the extremes is referred to as 2Azz (Fig. 1, bottom spectrum). Similar spectra were obtained for pea axes in relation to water content (data not shown).
Figure 2 shows EPR spectra of CP in dry pea axes and cattail pollen (both having 0.07 g water g−1 dry weight) at a range of temperatures. At least two overlapping spectra contributed to the total spectrum observed: a powder spectrum at all temperatures and a mobile spectrum above 20°C, the contribution of which increased with increasing temperature. Note that the contribution of the mobile component to the total spectrum is considerably larger for pollen at 70°C than for pea axes at 90°C.
Figure 2.

EPR spectra of CP in pea axes and cattail pollen recorded at various temperatures. Both contained approximately 0.07 g water g−1 dry weight.
At −150°C, we assumed that the motion of the probe was completely immobilized and therefore we took the corresponding 2Azz values as the maximum values. At this low temperature, the value of 2Azz gives information about the polarity of the spin probe's environment in the tissue (Knowles et al., 1976). In pea axes the maximum 2Azz decreased with decreasing water content from 74 to 70 G, whereas in cattail pollen, it changed from 72.5 to 71.5 G (Fig. 3).
Figure 3.

Change in the distance between the outer extrema of the EPR spectra (2Azz) of CP in pea axes (○) and cattail pollen (•) at −150°C as a function of water content. dw, Dry weight.
Because we observed a powder spectrum of CP in pea axes and cattail pollen at low water contents (<0.2 g water g−1 dry weight), τR cannot be directly calculated from the EPR spectra using Equation 1. However, the change in 2Azz with temperature can be used as an estimate of molecular motion (Van et al., 1974; Dzuba, 1996). A decrease in 2Azz is indicative of an increase in molecular mobility. Figure 4 shows these changes in 2Azz with temperature. When the temperature of pea axes and cattail pollen increased, the 2Azz slowly decreased, then abruptly decreased above a definite temperature. With increasing water contents, this abrupt decrease in 2Azz, denoting an abrupt increase in molecular mobility, commenced at lower temperatures. At a water content of 0.002 g water g−1 dry weight in pea axes the decrease in 2Azz was less clear.
Figure 4.

Comparison of the distance between the outer extrema of the EPR spectra (2Azz) of CP in pea axes and cattail pollen at different water contents against temperature.
We used ST-EPR to obtain an indication of the magnitude of change in τR (reflecting the lifetime of the probe in a given orientation) that corresponds to the change in 2Azz with temperature. This technique is based on recording spectra under saturation conditions, which yields line shapes that are sensitive to τR >10−7 s (Hyde and Dalton, 1979). The ST-EPR spectra can be well characterized by independent line-shape parameters such as the line-height ratios L"/L′ and C′/C (Fig. 5). These ratios are dependent on the τR of the spin probe. To determine the τR of CP corresponding to a certain line-height ratio, anhydrous glycerol was used as a reference solvent (this solvent was previously used as a reference in sugar glasses in Roozen et al. [1991] and Van den Dries et al. [1998]). Although the composition of the cytoplasm in seeds and pollen is not comparable to glycerol, at present the use of glycerol will give the best approximation of the relationship between the line-height ratios and τR. Extrapolation of the τR of CP in seeds and pollen to the corresponding viscosity is not valid, because the Stokes-Einstein law fails at temperatures below approximately 1.3 Tg (Liu and Oppenheim, 1996).
As the viscosity for glycerol is known over a broad temperature range, the corresponding τR could be calculated according to Equation 2. Subsequently, the two line-height ratios were obtained from ST-EPR scans of anhydrous glycerol for a range of temperatures and plotted against the corresponding τR. Those calibration curves were used to obtain τR from line-height ratios calculated from ST-EPR scans of CP in pea axes (Fig. 5). The τR values for pea axes (0.08 g water g−1 dry weight) calculated according to both line-height ratios and plotted against the temperature are shown in Figure 6. τR values of CP in pea axes were in the range of 10−2 to 10−6 s. The τR values of CP in the pollen were in the same range (data not shown). From Figure 6 it can be seen that there is a difference in the τR derived from the L"/L′ ratio and the C′/C ratio. Furthermore, it can be observed that the τR obtained from the C′/C ratio shows an increase in mobility with increasing temperature comparable to the 2Azz curves (Fig. 4).
Figure 6.

τR of CP in pea axes at 0.08 g water g−1 dry weight as a function of temperature. The τR values were obtained by comparing the L"/L′ ratio and the C′/C ratio with those of CP in anhydrous glycerol.
One can speculate on what causes the difference in τR obtained from both line-height ratios. The τR obtained from the L"/L′ ratio probably reflects overall isotropic rotational motion, whereas the τR derived from C′/C also reflects some anisotropic motion. According to Dzuba (1996), the change in 2Azz can be described by a librational model. Unlike rotational motion, in which the spin probe rotates entirely randomly, librational motion assumes that the spin probe rotates within a cone given by a small angle (α). Therefore, it could be that the anisotropic motion seen in the τR obtained from the C′/C ratio arises from librational motion. Whether the change in 2Azz with temperature is due to librational motion is currently under investigation. As ST-EPR is a laborious technique, more investigations are needed to make full use of this technique as a method to determine molecular mobility. Meanwhile, we will consider the measurements of 2Azz as an estimate of molecular motion.
Molecular Motion and Intracellular Glasses
As shown in Figure 4, a sharp increase in the molecular motion of CP was noticeable when the temperature of the sample was increased. Because the temperature corresponding to this sharp increase depended on water content, an attempt was made to explain this behavior according to the glass theory. From the plot of 2Azz against temperature we derived two characteristic temperature points (see inset in Fig. 7 for details): at the intercept (midpoint Tg) and at the point of deviation from a straight line (onset Tg). Figure 7 shows plots of the temperature at which the breaks occurred compared with the water content in pea axes. For cattail pollen a similar type of plot was obtained (data not shown). At low water contents (<0.002 g water g−1 dry weight) the characteristic temperature points were difficult to determine exactly, as the legs below and above the glass transition did not show a sharp drop in 2Azz. The curves in Figure 7 are remarkably similar to state diagrams of intracellular glasses in seeds (Leopold et al., 1994; Leprince and Walters-Vertucci, 1995) and pollen (Buitink et al., 1996).
Figure 7.

Effects of water content on the onset and midpoint of abrupt changes in the distance between the outer extrema of EPR spectra of CP in pea axes. The inset shows how the onset and midpoint temperatures were determined. Onset Tg was taken as the point of deviation from a straight line, and midpoint Tg as the intercept of the two lines (the line drawn through the data points is meant as an aid to visualization). dw, Dry weight.
To ascertain whether the sharp changes in molecular motion were due to the melting of intracellular glasses, the onset Tg was measured by differential scanning calorimetry and compared with the onset Tg as determined by EPR (Fig. 8). Both curves closely matched one another, the EPR data being slightly lower than the differential scanning calorimetry data. The Tg measured by differential scanning calorimetry has been found to occur above the Tg measured at a molecular level (Kalichevsky et al., 1992). The midpoint of Tg from EPR measurements was situated slightly above the midpoint of the Tg measured by differential scanning calorimetry (data not shown). The Tg for dry pollen (62°C) was lower than that for dry pea axes (92°C), which may be related to the high level of oligosaccharides in the latter (Amuti and Pollard, 1977; Saleki-Gerhardt and Zografi, 1994). The constant value of Tg measured by differential scanning calorimetry when the last 2% of water was removed may indicate that the first small amount of water does not contribute to plasticization of the glass. It is possible that this water is not present in the glass but, rather, is located in some other part of the tissue (e.g. cell walls).
Figure 8.

The relationship between Tg and water content in pea axes and cattail pollen. The open symbols show the Tg determined as the onset of the second-order-like transitions measured by differential scanning calorimetry during heating with a scanning rate of 10°C min−1. The closed symbols show the onset of the sudden decrease in 2Azz (derived from Fig. 4). dw, Dry weight.
The change in molecular mobility during melting of intracellular glasses can be measured as a function of temperature or water content, the relationship being reflected by the state diagram (Fig. 8). To determine the change in molecular mobility as a function of water content instead of temperature, it is necessary to correct for the polarity change of the environment in which CP is present for each water content (Fig. 3). Therefore, using curves similar to those shown in Figure 4, the mobility at a certain water content was expressed as the difference between the maximum 2Azz (at −150°C, where the spin probe is assumed to be immobilized) and the 2Azz measured at the desired temperature. We refer to this parameter as ΔAzz. Thus, an increase in ΔAzz represents a relative increase in molecular mobility compared with the completely immobilized situation at −150°C (i.e. the more the value departs from zero, the higher the molecular mobility). Figures 9 and 10 show the dependence of the molecular mobility (ΔAzz) on water content in pea axes and pollen, respectively. Between approximately 0.2 and 0.1 g water g−1 dry weight, the mobility decreased with decreasing water content at all temperatures analyzed. When the tissues reached approximately 0.1 to 0.05 g water g−1 dry weight, the ΔAzz reached a constant value, indicating that the mobility had reached a minimum. When water contents were decreased further, below approximately 0.05 g water g−1 dry weight, mobility increased again for pollen (Fig. 10). For pea axes, the mobility slightly increased again or reached a constant level at very low water content (Fig. 9). The water content corresponding to the minimum mobility (lowest ΔAzz) shifted to higher values with decreasing temperatures.
Figure 9.

Change in ΔAzz, calculated as the difference between the maximum 2Azz (−150°C) and 2Azz at the indicated temperatures, with water content for spectra of CP in pea axes. Curves were fitted with a third-order polynomial. dw, Dry weight.
Figure 10.

Change in ΔAzz, calculated as the difference between the maximum 2Azz (at −150°C) and 2Azz at the indicated temperatures, with water content spectra of CP in cattail pollen. Curves were fitted with a third-order polynomial. dw, Dry weight.
DISCUSSION
Behavior of Nitroxide Spin Probes in Dehydrating Organisms
To characterize molecular motion using EPR, a spin probe must be introduced into the material. Depending on the polarity of the spin probe, it will partition into the apolar oil phase, polar aqueous cytoplasm, or both. Several amphipathic spin probes, such as TEMPO and 4-oxo-2,2,6,6-tetramethyl-1-piperidinyloxy, completely partition into the lipid phase during drying of cattail pollen (Hoekstra and Golovina, 1998). We found that the more polar 4-hydroxy-2,2,6,6-tetramethylpiperidinyloxy partially partitioned into the lipid phase of cattail pollen and pea axes during drying (data not shown). Because we were interested in the molecular mobility in the cytoplasm rather than in the lipid phase, we avoided the use of these spin labels and instead used the polar nitroxide spin probe CP.
There are several reasons why we believe that CP was not present in the lipid phase in the dry organisms. Upon melting of the oil, which occurred in cattail pollen at approximately −20°C and in pea axes at −40°C and −20°C, as determined by differential scanning calorimetry (data not shown), we never observed an abrupt appearance of a mobile EPR spectrum. Furthermore, during drying of pollen at 25°C, when the oil is liquid (Fig. 1), we did not observe the typical lipid signal with a hyperfine splitting constant of 14 G (Knowles et al., 1976). The resolution of the spectrum was high enough to distinguish a possible oil contribution from the aqueous contribution. When samples of elevated water content were heated above 80°C, two peaks were observed in the high-field part of the spectrum, one attributable to the water signal and the other to the lipid signal (data not shown).
During drying the hyperfine splitting constant of CP decreased from 16.5 to 15.5 G (Fig. 1). A decrease in hyperfine splitting constant is associated with a decrease in the polarity of the spin probe environment (Knowles et al., 1976). During drying the apparent polarity of the cytoplasmic environment decreased, possibly because of the decrease of the dielectric constant of the environment associated with the loss of water. This is also substantiated by the observation of the decrease in 2Azz with water loss at −150°C (Fig. 3). It is interesting to note that cattail pollen and pea axes differ in the extent of their 2Azz decrease. Therefore, it seems evident that changes in water content are not solely responsible for this difference and that some intrinsic factors (e.g. salts or proteins) also contribute to the decrease in 2Azz during drying.
Molecular Mobility in Biological Glasses
Although the presence of intracellular glasses in seeds and pollen has been established, little is known about their viscosity and molecular mobility upon melting. We investigated the molecular motion of a spin probe in the cytoplasm of dry tissues to establish the relationship between glasses, molecular mobility, and storage stability.
During drying of cattail pollen and pea axes, the τR of CP increases from 10−11 s in the hydrated state to 10−2 s in the dry state. These values are consistent with values obtained in organic and inorganic glasses (Ito, 1983; Dzuba et al., 1984; Roozen et al., 1991), but contradict a previous study on molecular mobility in dry soybean axes, in which the τR of TEMPO was found to be approximately 10−10 s (Bruni and Leopold, 1990). Referring to the tendency of this spin probe to partition into oil during drying, we attribute this contradiction to the fact that these authors might have observed TEMPO in oilbodies. They nevertheless observed an abrupt change in molecular mobility around Tg. It is interesting that TEMPO in the lipid phase seems to be able to “sense” the glass transition of the cytoplasmic surroundings.
Molecular mobility was assessed in relation to water content and temperature. The change in distance between the two outer extrema of the powder spectrum (2Azz) with increasing temperature revealed a sharp increase in molecular mobility at a certain temperature that depended on the sample water content. This sharp increase was closely associated with the Tg as measured by differential scanning calorimetry. When τR was monitored during melting of the glass, there was a decrease of four orders of magnitude, from 10−2 to 10−6 s.
We found a close correlation between changes in τR, determined by the C′/C ratio of ST-EPR spectra and changes in 2Azz derived from conventional EPR spectra during melting of intracellular glasses (compare Figs. 4 and 6). This correlation indicates that a change in 2Azz represents a change in molecular mobility. One can speculate on the type of molecular mobility that gives rise to the change in 2Azz. Echo-detected EPR spectroscopy of nitroxide spin probes dissolved in organic glasses has revealed that the nitroxides undergo librational motions (Dzuba, 1996). This type of motion is described by a model that assumes that the spin probe rotates within a cone given by a small angle, α (Dzuba et al., 1992; Dzuba, 1996). Since we measured a τR for CP in intracellular glasses of 10−2 to 10−4 s, it seems unlikely that the change in 2Azz was due to a change in overall rotational motion; most likely it represents a change in librational motion, as has been established in wheat embryos (Dzuba et al., 1996). This is further supported by our ST-EPR study, in which some indication for anisotropic motion arising from libration comes from the different values of τR deduced from the line-height ratios L"/L′ and C′/C (Fig. 6). For the interpretation of our results, however, a motional model that describes the change of 2Azz is not needed.
Storage Stability in Relation to Molecular Mobility and Intracellular Glasses
It has been shown that upon formation of glasses, the storage stability of seeds and pollen improves (Sun and Leopold, 1994; Sun, 1997; Buitink et al., 1998). The impact of intracellular glasses on the storage behavior of seeds and pollen has been ascribed to the high viscosity in the glass. Indeed, glasses are known to slow down detrimental reactions such as the rate of browning reactions (Karmas et al., 1992) and to increase the stability of enzymes (Chang et al., 1996). Although the presence of glasses has been associated with increased storage stability of seeds and pollen, there is not much known about the relationship between molecular mobility and storage stability. Our data on molecular motion in pea axes and cattail pollen in relation to water content and temperature enable a comparison with storage behavior, which is also known to depend on water content and temperature (Vertucci et al., 1994; Buitink et al., 1998).
We found a close relationship between the molecular mobility of CP and storage behavior in both pollen and seed axes. With decreasing water content molecular mobility, expressed as a change in the outer extrema of the EPR spectra (ΔAzz), decreased, whereas storage stability increased. Figure 11 clearly demonstrates that molecular mobility and storage stability are linked; the water content for optimum storage at various temperatures corresponds closely to the water content at which minimum mobility is observed (data from Vertucci et al., 1994; Buitink et al., 1998). Although the curves do not converge exactly with the 1:1 line, the slight deviation might be explained from errors in the determination of water content or the lack of an exact determination of the water content for optimum storage stability. Furthermore, the water content of minimum mobility was determined from the minima of the third-order polynomial equations. Especially at lower temperatures, the exact water content of minimum mobility is difficult to assess; there seems to be a plateau of minimum mobility present. To present the data clearly, we calculated a single value of minimum mobility, which should be considered with caution.
Figure 11.

Correlation between water content (wc) of optimum storage longevity for pea seeds and cattail pollen and the water content of minimum mobility (lowest value of ΔAzz). Water contents for optimum storage longevity for cattail were determined by Buitink et al. (1998). The water content of optimum storage longevity was found to increase with decreasing temperatures of storage (45°C, 35°C, 25°C, 15°C, and 5°C). Water contents of optimum storage longevity for pea seeds stored in the light were derived from Vertucci et al. (1994). With decreasing storage temperature (45°C, 35°C, 25°C, 15°C, 5°C, and −5°C), the water contents of optimum storage increased. The water content at which minimum mobility was observed was determined from the minima of the third-order polynomial equations derived from curves similar to Figures 9 and 10 at temperatures comparable to the storage temperatures (45°C, 35°C, 25°C, 15°C, 5°C, and −5°C). dw, Dry weight.
At low water content, molecular mobility seems to increase again. A similar observation has been made by Seitz et al. (1981), who observed that in Artemia cysts at lower hydration levels, the water self-diffusion coefficients increased slightly. Clegg et al. (1982) suggested from NMR studies on Artemia cysts that the increased mobility of water at a low water content might be due to a displacement of the water from polar-binding sites in the cell by sugars. Another explanation is that at these low water contents the spin probe partitions into a more mobile environment. However, it should be noted that this environment cannot be the lipid phase. CP in these phases will rotate faster than 10−8 s at room temperature. Therefore, the spectrum of CP in the oil phase will show the characteristic three sharp lines of a mobile spectrum and does not contribute to the 2Azz of the powder spectrum.
Optimum Storage Conditions Predicted by Molecular Mobility
Recent studies report a water-content limit below which seed longevity did not increase further (Ellis et al., 1989, 1990), and which had an adverse effect on seed viability and seed vigor (Vertucci and Roos, 1993; Vertucci et al., 1994; Buitink et al., 1998). It is thought that the removal of the last remaining water molecules may destabilize biological structures (Sun, 1997; Buitink et al., 1998) or enhance the lifetime of free radicals due to the loss of water as a quencher (Karel, 1975). However, we observed an increase in molecular mobility for cattail pollen (which also occurred to a lesser extent in pea axes) when it was dried to very low water contents. This increase in molecular mobility also might be responsible for the decreased storage stability observed at these low water contents.
In some cases, state diagrams can be used to predict the optimal storage conditions (Sun, 1997). For pea axes the optimum water contents of storage at a certain temperature were found to coincide with Tg (Fig. 8; Vertucci et al., 1994). However, other seed species show a divergence of the optimal storage stability from the Tg curve (Sun, 1997). This is also true for cattail pollen (Buitink et al., 1998), in which we found that the optimum storage conditions coincided with the water content at which molecular mobility was minimum (Fig. 11). This minimum mobility occurred below Tg. Therefore, we propose that measurements of molecular mobility rather than state diagrams be used to predict optimum storage conditions.
Based on the relationship between the minimum molecular mobility and the water content of optimal storage, we made an attempt to predict the optimum storage conditions at subzero temperatures. This was found to be −20°C and −60°C for pea axes (Fig. 9, E and F) and −20°C and −40°C for cattail pollen (Fig. 10, E and F). In practice it will not be possible to analyze storage behavior at these low temperatures on a realistic time scale. At −20°C the minimum molecular motion of pea can be estimated at approximately 0.10 g water g−1 dry weight, and at −60°C between 0.14 and 0.2 g water g−1 dry weight. A similarly elevated optimum water content for low-temperature storage was predicted by Vertucci and Roos (1993) on the basis of thermodynamic considerations. At −20°C the minimal molecular motion of pollen was approximated at 0.1 g water g−1 dry weight, and at −40°C between 0.15 and 0.2 g water g−1 dry weight. With lower temperatures, an increase in the water content at which minimum mobility was observed became evident. This implies that too much drying increases mobility and reduces longevity and should be avoided, particularly when cryogenic storage is considered. Where determinations of optimal storage conditions of seeds and pollen by germination assays take too long to perform, measurements of 2Azz might be considered instead.
ACKNOWLEDGMENTS
The authors thank Drs. Olivier Leprince and Elena Golovina for critically reading the manuscript and Mark Alberda for excellent technical assistance.
Abbreviations:
- CP
3-carboxy-proxyl
- EPR
electron paramagnetic resonance
- ST-EPR
saturation-transfer EPR
- τR
rotational correlation time
- TEMPO
2,2,6,6-tetramethyl-1-piperidinyloxy
- Tg
glass-to-liquid transition temperature
Footnotes
This research was financially supported by the Netherlands Technology Foundation (STW) and was coordinated by the Life Sciences Foundation.
LITERATURE CITED
- Amuti KS, Pollard CJ. Soluble carbohydrates of dry and developing seeds. Phytochemistry. 1977;16:529–532. [Google Scholar]
- Blackburn FR, Wang C-Y, Ediger MD. Translational and rotational motion of probes in supercooled 1,3,5-tris(naphthyl)benzene. J Phys Chem. 1996;100:18249–18257. [Google Scholar]
- Bruni F, Leopold AC. Glass transitions in soybean seed. Plant Physiol. 1990;96:660–663. doi: 10.1104/pp.96.2.660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buitink J, Walters C, Hoekstra FA, Crane J. Storage behavior of Typha latifolia pollen at low water contents: interpretation on the basis of water activity and glass concepts. Physiol Plant. 1998;103:145–153. [Google Scholar]
- Buitink J, Walters-Vertucci C, Hoekstra FA, Leprince O. Calorimetric properties of dehydrating pollen: analysis of a desiccation-tolerant and an -intolerant species. Plant Physiol. 1996;111:235–242. doi: 10.1104/pp.111.1.235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burke MJ (1986) The glassy state and survival of anhydrous biological systems. In AC Leopold, ed, Membranes, Metabolism and Dry Organisms. Cornell University Press, Ithaca, NY, pp 358–363
- Chang BS, Beauvais RM, Dong A, Carpenter JF. Physical factors affecting the storage stability of freeze-dried interleukin-1 receptor antagonist: glass transition and protein conformation. Arch Biochem Biophys. 1996;331:249–258. doi: 10.1006/abbi.1996.0305. [DOI] [PubMed] [Google Scholar]
- Clegg JS, Seitz P, Seitz W, Hazlewood CF. Cellular responses to extreme water loss: the water-replacement hypothesis. Cryobiology. 1982;19:306–316. doi: 10.1016/0011-2240(82)90159-6. [DOI] [PubMed] [Google Scholar]
- Dzuba SA. Librational motion of guest spin probe molecules in glassy media. Phys Lett A. 1996;213:77–84. [Google Scholar]
- Dzuba SA, Golovina EA, Tsvetkov YD. Spin-probe EPR study of some sugars in connection with desiccation tolerance of biological objects. Appl Magn Reson. 1993;4:31–37. [Google Scholar]
- Dzuba SA, Golovina EA, Tsvetkov YD. Spin-probe EPR study of intracellular glass transition in embryos of wheat seeds. Dokl Akad Nauk. 1996;347:119–122. [Google Scholar]
- Dzuba SA, Maryasov AG, Salikhov KM, Tsvetkov YD. Superslow rotations of nitroxide radicals studied by pulse EPR spectroscopy. J Magn Res. 1984;58:95–117. [Google Scholar]
- Dzuba SA, Tsvetkov YD, Maryasov AG. Echo-induced EPR spectra of nitroxides in organic glasses: model of orientational molecular motions near equilibrium position. Chem Phys Lett. 1992;188:217–222. [Google Scholar]
- Ellis RH, Hong TD, Roberts EH. A comparison of the low-moisture-content limit to the logarithmic relation between seed moisture and longevity in twelve species. Ann Bot. 1989;63:601–611. [Google Scholar]
- Ellis RH, Hong TD, Roberts EH, Tao K-L. Low moisture content limits to relations between seed longevity and moisture. Ann Bot. 1990;65:493–504. [Google Scholar]
- Franks F, Hatley RHM, Mathias S. Materials science and the production of shelf-stable biologicals. BioPharm. 1991;4:38–42. , 55. [Google Scholar]
- Golovina EA, Tikhonov AN. The structural differences between the embryos of viable and nonviable wheat seeds as studied with the EPR spectroscopy of lipid-soluble spin labels. Biochim Biophys Acta. 1994;1190:385–392. doi: 10.1016/0005-2736(94)90098-1. [DOI] [PubMed] [Google Scholar]
- Golovina EA, Tikhonov AN, Hoekstra FA. An electron paramagnetic resonance spin probe study of membrane permeability changes with seed aging. Plant Physiol. 1997;114:383–389. doi: 10.1104/pp.114.1.383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hemminga MA. Interpretation of ESR and saturation transfer ESR spectra of spin labeled lipids and membranes. Chem Phys Lipids. 1983;32:323–383. [Google Scholar]
- Hemminga MA, de Jager PA, Marsh D, Fajer P. Standard conditions for the measurement of saturation-transfer ESR spectra. J Magn Reson. 1984;59:160–163. [Google Scholar]
- Hemminga MA, Van den Dries IJ (1998) Spin label applications to food science. In LJ Berliner, ed, Biological Magnetic Resonance, Vol 14. Spin Labeling: The Next Millennium. Plenum Publishing Corp., New York, pp 1–26
- Hoekstra FA, Golovina EA (1998) Role of amphipaths in desiccation tolerance. In AG Taylor, X-L Huang, eds, Proceedings of the Second International Conference on Seed Science and Technology, Guangzhou, China. New York State Agricultural Experiment Station, Geneva, NY (in press)
- Hyde JS, Dalton LR (1979) Saturation transfer ESR. In LJ Berliner, ed, Spin Labeling II. Theory and Applications, Academic Press, New York, pp 1–70
- Ito Y. Rotational correlation times of a nitroxide spin probe in low temperature matrices determined by saturation transfer ESR: their correlation to dynamic behavior of radiation products. J Chem Phys. 1983;79:2650–2658. [Google Scholar]
- Kalichevsky MT, Jaroszkiewicz EM, Ablett S, Blanshard JMV, Lillford PJ. The glass transition of amylopectin measured by differential scanning calorimetry, DMTA and NMR. Carbohydr Polym. 1992;18:77–88. [Google Scholar]
- Karel M (1975) Free radicals in low moisture systems. In RB Duckworth, ed, Water Relations of Foods. Academic Press, New York, pp 435–453
- Karmas R, Buera MP, Karel M. Effect of glass transition on rates of nonenzymatic browning in food systems. J Agric Food Chem. 1992;40:873–879. [Google Scholar]
- Knowles PF, Marsh D, Rattle HWE. Magnetic Resonance of Biomolecules: an Introduction to the Theory and Practice of NMR and ESR in Biological Systems. London: John Wiley & Sons; 1976. [Google Scholar]
- Kovarskii AL, Placek J, Szocs F. Study of rotational mobility of stable nitroxide radicals in solid polymers. Polymer. 1978;19:1137–1141. [Google Scholar]
- Kumler PL, Boyer RF. ESR studies of polymer transitions. Macromolecules. 1976;9:903–910. [Google Scholar]
- Leopold AC, Sun WQ, Bernal-Lugo I. The glassy state in seeds: analysis and function. Seed Sci Res. 1994;4:267–274. [Google Scholar]
- Leprince O, Walters-Vertucci C. A calorimetric study of glass transition behaviors in axes of bean with relevance to storage stability. Plant Physiol. 1995;109:1471–1481. doi: 10.1104/pp.109.4.1471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu CZ-W, Oppenheim I. Enhanced diffusion upon approaching the kinetic glass transition. Phys Rev E. 1996;53:799–802. doi: 10.1103/physreve.53.799. [DOI] [PubMed] [Google Scholar]
- Ohta N, Kuwata K. ESR study on rotational motions of some free radicals in the supercooled and glassy organic solvents. J Chem Phys. 1985;82:3420–3429. [Google Scholar]
- Priestley DA (1986) Seed Aging: Implications for Seed Storage and Persistence in Soil. Comstock Publishing Association, Ithaca, NY, pp 1–304
- Roberts EH (1972) Storage environment and the control of viability. In EH Roberts, ed, Viability of Seeds. Chapman and Hall, Ltd., London, pp 14–58
- Roberts EH, Ellis RH. Water and seed survival. Ann Bot. 1989;63:39–52. [Google Scholar]
- Roozen MJGW, Hemminga MA. Molecular motion in sucrose-water mixtures in the liquid and glassy state as studied by spin probe ESR. J Phys Chem. 1990;94:7326–7329. [Google Scholar]
- Roozen MJGW, Hemminga MA, Walstra P. Molecular motion in glassy water-malto-oligosaccharide (matrodextrin) mixtures as studied by conventional and saturation-transfer spin-probe e.s.r. spectroscopy. Carbohydr Res. 1991;215:229–237. [Google Scholar]
- Saleki-Gerhardt A, Zografi G. Non-isothermal and isothermal crystallization of sucrose from the amorphous state. Pharm Res. 1994;11:1166–1173. doi: 10.1023/a:1018945117471. [DOI] [PubMed] [Google Scholar]
- Seitz PK, Chang DC, Hazlewood CF, Rorschach HE, Clegg JS. The self-diffusion of water in Artemia cysts. Arch Biochem Biophys. 1981;210:517–524. doi: 10.1016/0003-9861(81)90216-2. [DOI] [PubMed] [Google Scholar]
- Spielberg JI, Gelerinter E. An EPR spin label study of the glass forming liquid dibutyl phthalate. J Chem Phys. 1982;77:2159–2165. [Google Scholar]
- Steffen W, Patkowski A, Meier G, Fischer EW. Depolarized dynamic light scattering studies of ortho-terphenyl dynamics above Tg. J Chem Phys. 1992;96:4171–4179. [Google Scholar]
- Sun WQ. Glassy state and seed storage stability: the WLF kinetics of seed viability loss at T-Tg and the plasticization effect of water on storage stability. Ann Bot. 1997;79:291–297. [Google Scholar]
- Sun WQ, Leopold AC. The glassy state and accelerated aging of soybeans. Physiol Plant. 1993;89:767–774. [Google Scholar]
- Sun WQ, Leopold AC. Glassy state and seed storage stability: a viability equation analysis. Ann Bot. 1994;74:601–604. [Google Scholar]
- Thomas DD, Dalton LR, Hyde JS. Rotational diffusion studied by passage saturation transfer electron paramagnetic resonance. J Chem Phys. 1976;65:3006–3024. [Google Scholar]
- Van SP, Birrell GB, Griffith OH. Rapid anisotropic motion of spin labels: models for motion averaging of the ESR parameters. J Magn Res. 1974;15:444–459. [Google Scholar]
- Van Bilsen DGJL, Hoekstra FA. Decreased membrane integrity in aging Typha latifolia L. pollen: accumulation of lysolipids and free fatty acids. Plant Physiol. 1993;101:675–682. doi: 10.1104/pp.101.2.675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Bilsen DGJL, Hoekstra FA, Crowe LM, Crowe JH. Altered phase behavior in membranes of aging dry pollen may cause imbibitional leakage. Plant Physiol. 1994;104:1193–1199. doi: 10.1104/pp.104.4.1193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van den Dries IJ, de Jager PA, Hemminga MA. Sensitivity of saturation transfer electron spin resonance extended to extremely slow mobility in glassy materials. J Magn Reson. 1998;131:241–247. doi: 10.1006/jmre.1997.1354. [DOI] [PubMed] [Google Scholar]
- Vertucci CW, Roos EE. Theoretical basis of protocols for seed storage II: the influence of temperature on optimal moisture levels. Seed Sci Res. 1993;3:201–213. [Google Scholar]
- Vertucci CW, Roos EE, Crane J. Theoretical basis of protocols for seed storage III: optimum moisture contents for pea seeds stored at different temperatures. Ann Bot. 1994;74:531–540. [Google Scholar]
- Wettlaufer SH, Leopold AC. Relevance of Amadori andMaillard products to seed deterioration. Plant Physiol. 1991;97:165–169. doi: 10.1104/pp.97.1.165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williams RJ, Leopold AC. The glassy state in corn embryos. Plant Physiol. 1989;89:977–981. doi: 10.1104/pp.89.3.977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilson DO, Jr, McDonald MB., Jr The lipid peroxidation model of seed aging. Seed Sci Technol. 1986;14:269–300. [Google Scholar]
- Winston PW, Bates DH. Saturated solutions for the control of humidity in biological research. Ecology. 1960;41:232–236. [Google Scholar]