

Abstract
Microsomal ω-3 fatty acid desaturase catalyzes the conversion of 18:2 (linoleic acid) to 18:3 (α-linolenic acid) in phospholipids, which are the main constituents of extrachloroplast membranes. Transgenic tobacco (Nicotiana tabacum) plants with increased 18:3 contents (designated SIIn plants) were produced through the introduction of a construct with the tobacco microsomal ω-3 fatty acid desaturase gene under the control of the highly efficient promoter containing the E12Ω sequence. 18:3 contents in the SIIn plants were increased by about 40% in roots and by about 10% in leaves compared with the control plants. With regard to growth at 15°C and 25°C and the ability to tolerate chilling at 1°C and 5°C, there were no discernible differences between the SIIn and the control plants. Freezing tolerance in leaves and roots, which was assessed by electrolyte leakage, was almost the same between the SIIn and the control plants. The fluidity of plasma membrane from the SIIn plants was almost the same as that of the control plants. These results indicate that an increase in the 18:3 level in phospholipids is not directly involved in compensation for the diminishment in growth or membrane properties observed under low temperatures.
Polyunsaturated fatty acids, especially 18:3 (α-linolenic acid), in membrane lipids have been found to increase in many plant species when the plants are exposed to low temperatures (Smolenska and Kuiper, 1977; Clarkson et al., 1980; Kodama et al., 1995). An increase in polyunsaturated fatty acids in lipids has been thought to increase the membrane fluidity by decreasing the scope for orderly packing of acyl chains within the membrane interior (Chapman, 1975). Because a decrease in membrane fluidity at low temperatures probably causes a loss of membrane permeability (Kuiper, 1974) and diminishment of activities of membrane-associated enzymes (Raison, 1973; Cronan and Gelmann, 1975), maintaining a constant fluidity of membranes has been considered essential for survival at chilling and freezing temperatures (Wolfe, 1978). Although many attempts to find a causal relationship between an increase in polyunsaturated fatty acid content and chilling or freezing acclimation have been made in higher plants, the results have been controversial (Steponkus, 1984). The acclimation to low temperatures in plants is associated with not only an increase in 18:3 content but also with alterations in other biochemical metabolic processes: changes in lipid composition and the synthesis of low-temperature-induced proteins and cryoprotectants (Guy, 1990). Therefore, the precise role of increases in the 18:3 content in membrane lipids is difficult to ascertain from a comparison of low-temperature-acclimated and -nonacclimated plants.
Although it remains unclear how important fatty acid polyunsaturation is in determining membrane fluidity and low-temperature tolerance, studies using Arabidopsis mutants with altered fatty acid composition in glycerolipids have suggested that polyunsaturated fatty acids are required for the growth and tolerance of higher plants at low, nonfreezing temperatures. The Arabidopsis fad5 and fad6 mutants, in which desaturation steps in the prokaryotic pathway are deficient and in which the contents of polyunsaturated fatty acids in the plastidic lipids are decreased, showed leaf chlorosis, reduced growth rates, and impaired chloroplast development at low temperatures (Hugly and Somerville, 1992). On the other hand, the fad2 mutant defective in desaturation at the eukaryotic pathway, which exhibits a preferential decrease in polyunsaturated fatty acids in the microsomal lipids, showed increased chilling injuries such as inhibition of stem elongation at 12°C and loss of viability at 6°C (Miquel et al., 1993).
The ω-3 fatty acid desaturases are membrane-bound enzymes found in microsomes and in plastid envelopes (for review, see Mazliak, 1994) that catalyze the conversion of 18:2 (linoleic acid ) to 18:3 in lipids. Three genes, FAD3, encoding the microsomal desaturase, and FAD7 and FAD8, encoding the plastid desaturases, were isolated from Arabidopsis (Iba et al., 1993; Yadav et al., 1993; Gibson et al., 1994). Isolation of such desaturase genes enabled us to modify the 18:3 content in both the plastidial and extraplastidial membrane lipids. In fact, the transgenic tobacco (Nicotiana tabacum) plants that contained increased trienoic fatty acids, namely 16:3 (hexadecatrienoic acid) and 18:3, in plastid membrane lipids were produced by an overexpression of the FAD7 gene (Kodama et al., 1994). Because such FAD7 transgenic tobacco leaves exhibited increased chilling tolerance, the increase in trienoic fatty acids within the plastid membrane would be one of the prerequisites for normal leaf growth at low, nonfreezing temperatures (Kodama et al., 1995). We also reported the successful production of transgenic tobacco plants that expressed the transcripts of the tobacco microsomal ω-3 fatty acid desaturase gene (NtFAD3) in sense and antisense orientations under the control of the CaMV 35S promoter (Hamada et al., 1996). In this report we describe engineering of the transgenic tobacco plants in which the 18:3 content in plasma membrane lipids increased drastically after the introduction of NtFAD3 cDNA under the control of the El2Ω sequence, a highly efficient promoter for enhanced expression. Such NtFAD3 transformants as those produced here allowed us to determine the effects of an increase in 18:3 within plasma membrane lipids on the function of higher plant cells without regard to other biochemical changes associated with the acclimation to low temperatures.
MATERIALS AND METHODS
Plasmid Construction
The 1.4-kb full-length cDNA of NtFAD3 (Hamada et al., 1994) was subcloned into the BamHI unique site of pUC18 to produce a plasmid, pF1-18S, which contains NtFAD3 cDNA in antisense orientation relative to the lacZ region. Most of the 5′ noncoding region of the NtFAD3 cDNA was deleted by digestion of pF1-18S with BbvII. The resulting fragment was repaired with the Klenow fragment to create blunt ends, and then cut with SacI to produce a truncated fragment. This cDNA fragment was cloned into the SmaI-SacI site of the binary plasmid pBE2113-GUS (Mitsuhara et al., 1996) to replace the GUS gene. The resulting plasmid, pTF1SIIn, contained the NtFAD3 cDNA fragment from nucleotides 52 to 1381 (see the NtFAD3 cDNA sequence, accession no. D26509) in sense orientation relative to the promoter sequences.
Plant Transformation
Tobacco (Nicotiana tabacum cv SR1) was transformed using Agrobacterium tumefaciens LBA4404 containing the plasmid pTF1SIIn, as described previously (Hamada et al., 1996). R1 seeds resulting from self-pollination were aseptically germinated in continuous light (60 μmol m−2 s−1) at 26°C on Murashige and Skoog medium (Murashige and Skoog, 1962) supplemented with 100 μg/mL kanamycin. The kanamycin-resistant R1 and R2 seedlings were subjected to further analyses. The seeds of the SIIn-24 plants hemizygous for the T-DNA allele were produced by backcrossing the homozygous R1 plant with the wild-type plant as a pollen donor.
Lipid and Fatty Acid Analysis
Individual lipids were purified from leaves and roots as described previously (Kodama et al., 1995). The fatty acid compositions of whole tissues and the individual lipids were determined by GC (GC-14B, Shimadzu, Kyoto, Japan) as described by Kodama et al. (1994).
Measurement of Root Growth
The hemizygous SIIn-24 and wild-type seeds were germinated on plates with Murashige and Skoog medium containing 0.8% (w/v) gellan gum at 26°C. The Murashige and Skoog plates were arranged vertically to directly observe root elongation. One week after sowing, halves of these Murashige and Skoog plates with seedlings were transferred to 15°C, and then the positions of the root tips were plotted each day as a means of measuring root elongation.
EL Test
In the leaf EL test, the third leaves (approximately 3-4 cm2) were excised from 4-week-old tobacco plants that had been cultured on soil under continuous light (3000 lux) at 26°C. Each detached leaf was cut in half and the halves were placed in a test tube. Deionized water (100 μL) was added to each test tube and leaf segments were fixed closely to the surface of the test tubes.
In the root EL test, 2-month-old, soil-grown tobacco plants were transplanted into hydroponic culture. In this hydroponic culture, plants were rooted in an aerated solution containing one-tenth-strength Murashige and Skoog medium for about 2 weeks under continuous illumination. Roots of 10 to 15 cm in length were rinsed briefly with deionized water and then cut into about 4- to 5-cm-long segments. The root segments were transferred to test tubes and fixed closely to the surface of the tubes, as in the case of the leaf samples.
Samples were cooled to −0.5°C for 30 min by using a program freezer (model MPF-40, Tokyorika, Tokyo, Japan), and then ice formation was achieved by introducing a small piece of ice into the test tubes. After a 40-min equilibration period at −0.5°C, the bath temperature was reduced automatically by 1°C increments every 30 min. The samples were withdrawn from the bath at specified temperatures and placed on ice. Samples were thawed overnight on ice and then incubated with 5 mL of deionized water at 25°C for 1 h. The EL from the frozen samples (ELfrozen) was measured by using a conductivity meter. The EL from unfrozen samples (ELunfrozen) and from samples frozen in liquid N2 (ELLN) were taken as 0% and 100% EL, respectively. The percentage of EL from frozen leaves and roots at the specified temperatures was calculated by the following equation:
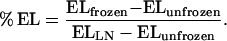 |
Isolation of Root Plasma Membrane
Roots excised from 2.5-month-old tobacco plants were washed with distilled water and prechilled at 4°C before use. The microsomal fraction from the roots was obtained as described by Yoshida et al. (1986). Root plasma membranes were purified from this fraction by aqueous two-phase partitioning and subsequent discontinuous Suc-density gradient (Yoshida et al., 1986). In the alternative method, plasma membranes were isolated from the microsome fraction by the aqueous two-phase partitioning method as described by Larsson et al. (1994). The fraction enriched with plasma membrane was diluted with Suc medium (0.25 m Suc, 5 mm Mops-KOH [pH 7.3], and 1 mm DTT) and the plasma membrane was pelleted by centrifugation at 156,000g for 20 min. The membrane pellets were suspended in a sorbitol buffer containing 0.25 m sorbitol, 5 mm Mops-KOH (pH 7.3), and 1 mm DTT. When the plasma membrane was prepared for the ESR assay, the membrane pellets were suspended in a sorbitol buffer without DTT, frozen in liquid N2, and stored at −80°C until use.
ESR Spectroscopy
The spin-label molecule (5-SLS) was dissolved in dichloromethane to a final concentration of 0.104 mm. The solution containing 3.12 nmol of 5-SLS was placed into a sample tube and then dried under a vacuum for 1 h. The solution containing the plasma membrane equivalent to 600 to 900 nmol of fatty acids was placed into the sample tube and shaken vigorously for 2 min to stimulate the incorporation of 5-SLS into the membrane fragments. The ESR spectra of the samples were measured using a JES-RE-1X spectrometer (Jeol, Tokyo, Japan) equipped with a temperature controller.
ATP Hydrolysis
The ATPase activity was determined by measuring the Pi released from ATP. The reaction mixture contained 3 mm Na-ATP, 3 mm MgSO4, 30 mm Tris-Mes (pH 7.0), and 50 mm KCl in a final volume of 150 μL, and was placed on ice until use. One minute before commencement of the reaction, the temperature of the reaction mixture was equilibrated at specified temperatures (0.5°C–40°C). The reaction was started by adding a plasma membrane solution containing 5 to 20 μg of proteins and was run for 30 min at specified temperatures. The reaction was stopped by adding 30 μL of 50% (v/v) ice-cold TCA, then centrifuged at 14,000g at 0°C for 10 min. The Pi concentration in the supernatant was determined by the method of Taussky and Shorr (1953).
RESULTS
Transgenic Tobacco Plants Harboring the NtFAD Construct
Previously, we showed that modulation of the NtFAD3 mRNA level is effective for modifying the 18:3 content in transgenic tobacco plants (Hamada et al., 1996). To produce plants with extremely high 18:3 content, we introduced the NtFAD3 cDNA under the control of the El2Ω sequence. The El2Ω sequence contains two tandem repeats of a part of the CaMV 35S promoter, which are followed by the Ω sequence (Mitsuhara et al., 1996). The El2Ω sequence can confer about 10-fold higher levels of expression than the CaMV 35S promoter. The Ω sequence has been reported to improve the translation efficiency within plants when it is located within the 5′ untranslated region of the target gene. To ensure the function of the Ω sequence, most of the 5′ untranslated region of the NtFAD3 cDNA was deleted and then the cDNA was linked behind the Ω sequence to create a plasmid, pTF1SIIn (see Methods). The tobacco transformants with pTF1SIIn were designated SIIn lines. Of the 23 SIIn transgenic lines, two lines (SIIn-20 and SIIn-24), which were segregated as a single T-DNA insertion plant, were selected as representatives of those plants with high 18:3 content and used in the following analyses.
Fatty Acid Composition of Leaf and Root Polar Lipids
Both the SIIn-20 and SIIn-24 lines had similar fatty acid compositions, and the 18:3 content in leaf tissues of the SIIn-20 plants increased by about 10%, whereas the 18:2 content decreased correspondingly compared with that of the control plants transformed with pBI121 (Table I). In contrast, the total 16:3 level of the SIIn-20 plants was almost the same as that of the control plants. In root tissues the 18:3 level of the SIIn-20 plants increased by about 40%, and was associated with a corresponding decrease in 18:2.
Table I.
Fatty acid composition of root and leaf tissues of control (Cl) and SIIn plants (SIIn-20)
| Fatty Acid | Leaf
|
Root
|
||
|---|---|---|---|---|
| Cl | SIIn-20 | Cl | SIIn-20 | |
| 16:0 | 10.0 ± 0.7a | 11.0 ± 0.9 | 26.2 ± 0.7 | 11.0 ± 0.9 |
| 16:1 | 2.4 ± 0.2 | 2.1 ± 0.1 | –b | – |
| 16:2 | 2.1 ± 0.2 | 1.2 ± 0.1 | – | – |
| 16:3 | 8.2 ± 1.0 | 8.0 ± 1.2 | – | – |
| 18:0 | 0.8 ± 0.2 | 0.9 ± 0.2 | 4.5 ± 2.32 | 3.7 ± 0.4 |
| 18:1 | 2.2 ± 0.2 | 1.6 ± 0.6 | 2.8 ± 0.8 | 3.4 ± 0.6 |
| 18:2 | 18.1 ± 1.7 | 7.2 ± 1.5 | 51.4 ± 2.2 | 8.4 ± 2.3 |
| 18:3 | 56.1 ± 1.1 | 67.9 ± 1.6 | 15.1 ± 1.9 | 56.8 ± 3.8 |
The kanamycin-resistant R1 seedlings were transferred to soil and cultured under continuous light (60 μmol m−2 s−1) at 26°C. Fatty acid compositions were determined in whole roots and whole third leaves of 30-d-old seedlings.
The values are mol % ± sd (n = 5).
–, Not detected.
To elucidate the effects of overexpression of the NtFAD3 gene on lipid metabolism, total lipids were extracted from leaf and root tissues, and the fatty acid composition of each polar lipid was determined. In both leaves and roots of the SIIn-20 plants, the proportion of the individual glycerolipids was almost the same as that of the control plants (Tables II and III). This indicates that enhancement of the ω-3 desaturation step does not significantly affect the synthesis of each lipid class.
Table II.
Fatty acid composition of individual leaf polar lipids from the control (Cl) and SIIn-20 plants grown at 26°C
| Polar Lipid and Plant | Total Polar Lipids | Fatty Acid Composition
|
|||||||
|---|---|---|---|---|---|---|---|---|---|
| 16:0 | 16:1 | 16:2 | 16:3 | 18:0 | 18:1 | 18:2 | 18:3 | ||
| % | |||||||||
| PC | |||||||||
| Cl | 9.0a | 20.5 | –b | – | – | 4.0 | 4.5 | 52.2 | 18.9 |
| SIIn-20 | 8.5 | 24.2 | – | – | – | 5.7 | 3.9 | 21.7 | 44.5 |
| PE | |||||||||
| Cl | 4.2 | 26.7 | – | – | – | 4.1 | 1.8 | 51.7 | 15.6 |
| SIIn-20 | 4.9 | 31.3 | – | – | – | 5.1 | 2.0 | 10.0 | 51.6 |
| PI | |||||||||
| Cl | 0.9 | 45.8 | 3.4 | – | – | 11.0 | 3.6 | 16.5 | 19.7 |
| SIIn-20 | 0.9 | 41.5 | 4.9 | – | – | 16.2 | 5.4 | 6.6 | 25.2 |
| PG | |||||||||
| Cl | 11.9 | 19.6 | 38.9 | – | – | 3.1 | 6.2 | 12.0 | 20.1 |
| SIIn-20 | 10.9 | 22.3 | 41.3 | – | – | 2.4 | 5.1 | 8.0 | 21.0 |
| DGDc | |||||||||
| Cl | 21.4 | 14.8 | – | – | 1.6 | 1.6 | 1.1 | 5.5 | 75.4 |
| SIIn-20 | 23.0 | 15.0 | – | – | 1.5 | 1.8 | – | 2.9 | 78.8 |
| MGD | |||||||||
| Cl | 53.6 | 2.0 | – | 1.5 | 18.7 | – | – | 3.8 | 74.0 |
| SIIn-20 | 49.0 | 1.8 | – | 1.6 | 19.8 | – | – | 1.8 | 74.9 |
Each value is the mean of two independent experiments.
–, Not detected.
DGD, Digalactosyldiacylglycerol.
Table III.
Fatty acid composition of individual root polar lipids from the control (Cl) and SIIn-20 plants grown at 26°C
| Polar Lipid and Plant | Total Polar Lipids | Fatty Acid
Composition
|
||||
|---|---|---|---|---|---|---|
| 16:0 | 18:0 | 18:1 | 18:2 | 18:3 | ||
| % | ||||||
| PC | ||||||
| Cl | 44.4a | 25.2 | 6.3 | 5.3 | 55.3 | 7.9 |
| SIIn-20 | 48.0 | 24.4 | 5.8 | 8.4 | 17.9 | 43.5 |
| PE | ||||||
| Cl | 26.9 | 22.8 | 4.0 | 1.8 | 63.0 | 8.4 |
| SIIn-20 | 27.6 | 27.5 | 4.6 | 2.4 | 20.3 | 45.2 |
| PI | ||||||
| Cl | 10.3 | 52.6 | 9.2 | 3.5 | 28.3 | 6.5 |
| SIIn-20 | 8.9 | 53.1 | 7.2 | 4.2 | 4.9 | 30.7 |
| PS | ||||||
| Cl | 8.8 | 27.8 | 11.0 | 4.9 | 47.6 | 8.8 |
| SIIn-20 | 9.1 | 30.9 | 11.3 | 5.0 | 12.0 | 40.9 |
| PG | ||||||
| Cl | 2.0 | 62.0 | 17.1 | 1.1 | 15.6 | 4.2 |
| SIIn-20 | 2.7 | 65.7 | 17.4 | 3.3 | 5.0 | 8.6 |
| MGD | ||||||
| Cl | 4.7 | 15.6 | 9.1 | 3.1 | 37.1 | 35.1 |
| SIIn-20 | 3.7 | 23.0 | 9.4 | 4.7 | 10.4 | 52.6 |
Each value is the mean of two independent experiments.
In leaf lipids the most remarkable changes in fatty acid composition through the overexpression of the NtFAD3 gene were observed in PC and PE, which are the major components of extrachloroplastic membranes (Table II). The 18:3 content of these two lipid classes in the SIIn-20 plants increased by more than 25% compared with the 18:3 content of the control plants. MGD, digalactosyldiacylglycerol, and PG are major chloroplast membrane lipids and account for about 80% of the leaf polar lipids. The 16:3 and 18:3 contents of these plastidial lipids in the SIIn-20 plants did not show any significant changes compared with those of the control plants.
In root tissues the major membrane lipids were PC and PE, which together account for more than 70% of root polar lipids (Table III). In the SIIn-20 plants the 18:3 content increased about 5.5-fold in PC and PE, about 4.6-fold in PI and phosphatidylserine, and less than 2-fold in PG and MGD compared with the levels in the control plants. These results indicate that in root tissues, the effects of the overexpression of the NtFAD3 gene were evident both in plastid and extraplastid membrane lipids.
Plant Growth at Low and Normal Temperatures
To determine the effect of a high 18:3 content in membrane lipids on plant growth, the growth of the SIIn (wild-type) and control plants was examined. Four-week-old seedlings grown at 26°C were transferred to 26°C or 15°C and cultured under continuous light. Three to four months after the temperature shift, no visible differences were observed in the aerial parts of the mature plants. In addition, no discernible difference in the flowering time relative to the wild-type plants was found in the SIIn plants at the two temperatures tested.
To directly observe root elongation, seedlings were grown on the Murashige and Skoog plate without being transferred to soil. The root growth of the SIIn-24 plant was the same as that of the wild-type plant when grown at 26°C and 15°C (Fig. 1). Therefore, increased 18:3 content in membrane lipids only slightly affects the growth of vegetative organs in either the aerial or the subterranean parts of tobacco plants when grown at 26°C and 15°C.
Figure 1.

Effect of temperature on root growth. Seedlings of the SIIn-24 (•) and wild-type (○) plants were grown at 26°C (A) and 15°C (B). Vertical lines indicate sd (n = 9).
We also examined chilling tolerance in the SIIn plants at 1°C or 5°C. Ten-day-old SIIn and control seedlings grown on Murashige and Skoog solid medium in plastic boxes at 26°C were exposed to 1°C under continuous light (60 μmol m−2 s−1) for 7 d. Then, these seedlings were transferred to 26°C under continuous light. Two days later, leaf chlorosis was observed in both the SIIn and control seedlings, and the number of seedlings with chlorosis and the leaf growth were almost the same. When 10-d-old SIIn and control seedlings grown at 26°C were transferred to 5°C under continuous light (60 μmol m−2 s−1), the growth of both types of seedlings was inhibited. Two weeks later, the leaves of both types of seedlings turned pale green.
Freezing Tolerance in Vegetative Tissues
EL of leaves and roots of the SIIn plants was examined to determine the effect of high 18:3 contents on freezing tolerance. Figure 2 shows the percentage of EL of frozen tissues at specified temperatures (−0.5°C to −3°C). In the SIIn-20 and control plants, the EL of leaves and roots began to increase at −1.0°C and reached a maximum at −2.5°C. The temperatures at which 50% EL occurred in leaves from the SIIn-20 and control plants were approximately −1.6°C and −1.7°C, respectively, whereas those for roots from these two plants were approximately −1.5°C. Furthermore, in the transgenic plants (T-4 and T-6) overexpressing the plastidial ω-3 desaturase gene (FAD7), the same experiments were carried out. Although these plants had increased levels of 18:3 in plastid membrane lipids, similar results were observed (data not shown). These results indicated that an increase in 18:3 content in both plastidial and extraplastidial membrane lipid was not associated with any alteration to the freezing-tolerance ability in the vegetative tissues of tobacco plants.
Figure 2.

Freezing tolerance of leaves (A) and roots (B) of the SIIn-20 (•) and control (○) plants. Survival was determined by the EL measurement from leaves and roots after a freeze/thaw treatment. Vertical lines indicate sd (n = 4 in leaves, and n = 3 in roots).
H+-ATPase Activity and Fluidity of Plasma Membrane
The root plasma membrane was subjected to the assessment of H+-ATPase activity and membrane fluidity. In the plasma membrane fraction isolated from the SIIn-20 and control roots by the aqueous two-phase partitioning method, the ATPase activity was suppressed significantly by an inhibitor of the plasma membrane H+-ATPase, vanadate, but was hardly affected by inhibitors of vacuolar and mitochondrial membrane H+-ATPases, nitrate and azide, or by an inhibitor of the acidic phosphatase, molybdate (Table IV). This result indicated that the contamination of other membranes was negligible in this plasma membrane fraction.
Table IV.
Effect of inhibitors on the ATPase activity of plasma membranes isolated from the roots of SIIn-20 and control (Cl) plants
| Plant | ATPase Activity
|
|||
|---|---|---|---|---|
| Nitrate | Azide | Molybdate | Vanadate | |
| % of control without inhibitor | ||||
| Cl | 102.2 | 98.6 | 99.3 | 56.1 |
| SIIn-20 | 100.0 | 103.7 | 100.0 | 52.2 |
ATP activity was assayed at 30°C for 30 min in the presence of nitrate (50 mm KNO3), azide (1 mm NaN3), molybdate (100 μm Na2MoO4), or vanadate (100 μm Na3VO4).
Plasma membrane H+-ATPase activity is affected by the phospholipid environment in the membrane (for review, see Kasamo and Sakakibara, 1995). High 18:3 content was evident in the plasma membrane fraction purified from the SIIn-20 plants (data not shown). For elucidation of the effects of such a high 18:3 content on the membrane-bound enzyme activity, the H+-ATPase activity in the plasma membranes, which were isolated from the roots of 26°C-grown SIIn-20 and control plants, was assayed at temperatures ranging from 0.5°C to 40°C (Fig. 3). In Arrhenius plots of the vanadate-sensitive ATPase activity, breaks were observed around 10°C, and observed slopes were indistinguishable in the two membrane fractions prepared from the SIIn-20 and control plants. These results indicate that plasma membrane H+-ATPase activity is only slightly affected by an increase in 18:3 content within the surrounding phospholipids.
Figure 3.

Arrhenius plots of ATPase activity found in the root plasma membrane. Total ATPase activity (• and ○) and vanadate-sensitive ATPase activity (▴ and ▵) were determined in root plasma membranes prepared from SIIn-20 (• and ▴) and control (○ and ▵) plants. Vanadate-sensitive ATPase activity was determined in the presence of 100 μm vanadate. Each value is the mean of two independent experiments.
The spin-label molecule 5-SLS was used as a probe to monitor the dynamic properties of plasma membranes. Since overall splitting (2A) reflects the mobility of the nitroxide group of 5-SLS in the hydrophobic environment of the membrane (Jost et al., 1971; Utsumi et al., 1978), this ESR parameter can be used as an indicator of membrane fluidity. Lower values of overall splitting indicate that the membrane has become more fluid. The overall splitting values of the plasma membrane from SIIn-20 plants were almost the same as those from control plants at temperatures from −3°C to 30°C (Fig. 4). At temperatures from −3°C to 30°C, the plasma membrane with a high 18:3 content exhibited thermotropic properties similar to those of the control plasma membrane.
Figure 4.

Effect of temperature on the overall splitting value of the spin-labeled membrane vesicles. The spin-label molecule 5-SLS was incorporated into the root plasma membranes prepared from the SIIn-20 (•) and control (○) plants. mT, Millitesla.
DISCUSSION
During low-temperature acclimation of higher plants, there is a preferential synthesis of polyunsaturated fatty acids. In particular, an increase in 18:3 is observed in root tissues of herbaceous plants such as wheat and rye when they are exposed to low temperatures. For example, in the roots of rye during acclimation to low temperatures, the level of 18:3 in PC and PE increased from 20% to 40%, whereas the level of 18:2 correspondingly decreased from 50% to 30% (Clarkson et al., 1980). In the root the major membrane lipids are PC and PE, which together account for 70% to 75% of root polar lipids, and the 18:3 level would be regulated by the conversion of 18:2 to 18:3 in these lipids, in which the conversion is catalyzed by the microsomal ω-3 fatty acid desaturase in the eukaryotic pathway (Browse et al., 1993). For investigation of the role played by an increase in 18:3 in membrane lipids, we thus introduced a construct with the NtFAD3 gene under the control of the El2Ω sequence to produce transgenic tobacco plants with an extremely high 18:3 content. Overexpression of the NtFAD3 gene was effective at increasing the 18:3 content in phospholipids in leaf and root tissues of transgenic tobacco (Tables II and III). Because lipid composition and the ratio of fatty acid to protein content in the SIIn plants were almost the same as those in the control plants (data not shown), it seems that the precise role of a high 18:3 phenotype could be evaluated by using the SIIn plants produced here.
Influences of High 18:3 on Plasma Membrane Properties
The degree of unsaturation in fatty acids has long been considered to be one of the major factors that determine membrane fluidity (Chapman, 1975). Hypothetically, a decrease in temperature leads to a decrease in membrane fluidity, although an increase in polyunsaturated fatty acids might compensate for this decrease in membrane fluidity. Because fatty acid desaturation at low temperatures in plants is observed mainly from 18:2 to 18:3, we investigated the effects of increased 18:3 content on membrane fluidity. As shown in Figure 4, a decrease in temperature caused an increase in the viscosity of plasma membranes, although increased 18:3 content was not associated with the discernible changes in the ESR parameter showing membrane fluidity. This observation suggests that the role of desaturation from 18:2 to 18:3 in plasma membranes during acclimation to low temperatures is very limited in its compensation for a decrease in membrane fluidity at low temperatures. By using the Arabidopsis fad7 mutant, thylakoid membranes with a drastically reduced content of trienoic fatty acids (16:3 and 18:3) were assessed for their membrane fluidity, and these membranes were found to have only a slightly rigid nature at the temperatures tested compared with the corresponding membranes prepared from wild-type plants (McCourt et al., 1987). These results show that conversion from 18:2 to 18:3 in both plasma membrane lipids and thylakoid membrane lipids only slightly affects membrane fluidity.
The plasma membrane is implicated in chilling and freezing injury in temperate plant species (Lyons, 1973; Steponkus, 1984). The proportion of polyunsaturated species of PC and PE in the plasma membrane increases naturally during the 1st week of cold acclimation in both rye and oat (Uemura and Steponkus, 1994). Previously, the correlation between an increase in the 18:3 content in wheat seedlings during cold acclimation and the establishment of enhanced freezing tolerance was investigated by use of the inhibitors of 18:3 synthesis (Willemot, 1977; de la Roche, 1979). However, the results obtained were controversial. The freeze-induced lesions can be classified into two different destabilization mechanisms of the plasma membrane (Uemura et al., 1995). Over the range of −2°C to −4°C, the predominant freezing injury is caused by expansion-induced lysis, which is a consequence of the osmotic contraction by endocytotic vesiculation of plasma membranes during a freeze-thaw cycle. Over the range of −4°C to −8°C, the predominant phase of injury is freeze-induced lamellar-to-hexagonal II phase transitions of plasma membranes and other cellular membranes. The data shown in Figure 2 indicate that high 18:3 content in the plasma membrane did not directly affect freezing tolerance, such as avoiding leakage of ions from cytosol at temperatures of −0.5°C to −3°C in tobacco cells. Therefore, the increased 18:3 content was not effective at preventing the probable expansion-induced lysis.
Possible Role of Increased 18:3 Content on Low-Temperature Acclimation
At normal and low temperatures (26°C and 15°C, respectively), where continuous growth can take place, no visible differences in the growth of aerial and subterranean parts were observed between the high-18:3 and the control plants (Fig. 1). At lower temperatures (1°C and 5°C), there were no visible differences regarding chilling injuries in these plants. The Arabidopsis fad3 mutant, in which the 18:3 content in extrachloroplastic lipids was decreased (Browse et al., 1993), did not show any detectable phenotypic alteration in growth at either low or high temperatures (Browse et al., 1995). These results indicate that high or low 18:3 content in microsomal membrane lipids had almost no effect on cell division or expansion.
Because detectable differences were not observed among the membrane physical properties or in the growth response to temperature between the wild-type and high-18:3 plants, the role of 18:3 content in phospholipids observed in acclimated plants remains unclear. Transgenic tobacco plants with elevated levels of trienoic fatty acids (16:3 and 18:3) in chloroplast lipids attributable to an overexpression of the Arabidopsis FAD7 cDNA demonstrated an enhanced chilling tolerance (Kodama et al., 1994, 1995). Such different behavior regarding chilling tolerance may be explained by the fact that the target membrane system is quite different between transgenic tobacco plants introduced with the plastidial and microsomal ω-3 fatty acid desaturases. The chilling tolerance observed in the FAD7 transformants could be mediated by enhancement of the chloroplast functions required at low temperatures. During acclimation the expression of a number of low-temperature-responsive genes is induced. Among them, the Cor15a gene product is reported to interact with chloroplast membranes as a cryoprotectant (Artus et al., 1996). It is possible that during acclimation, molecular mechanisms conferring low-temperature tolerance are developed based on some interaction between cellular membranes modified at low temperatures and newly expressed proteins that are protective against chilling and freezing temperatures.
ACKNOWLEDGMENTS
We thank Dr. Yuko Ohashi (National Institute of Agrobiological Resources, Tsukuba, Japan) for providing the pBE2113-GUS plasmid. We also acknowledge Dr. Shizuo Yoshida (Hokkaido University), Dr. Toshinori Kinoshita (Kyushu University), and Mr. Michiharu Hara for their technical advice.
Abbreviations:
- CaMV
cauliflower mosaic virus
- EL
electrolyte leakage
- ESR
electron spin resonance
- MGD
monogalactosyldiacylglycerol
- PC
phosphatidylcholine
- PE
phosphatidylethanolamine
- PG
phosphatidylglycerol
- PI
phosphatidylinositol
- 5-SLS
5-(4′,4′-dimethyloxazolidine-N-oxy) stearic acid
- X:Y
a fatty acyl group containing X carbon atoms and Y cis double bonds
Footnotes
This work was supported in part by a grant-in-aid (Biotechnology no. 1317) from the Ministry of Agriculture, Forestry, and Fishery, Japan, and by a grant from the Japan Society for the Promotion of Science (no. JSPS-RFTF96L00602).
LITERATURE CITED
- Artus NN, Uemura M, Steponkus PL, Gilmour SJ, Lin C, Thomashow MF. Constitutive expression of the cold-regulated Arabidopsis thaliana Cor15a gene affects both chloroplast and protoplast freezing tolerance. Proc Natl Acad Sci USA. 1996;93:13404–13409. doi: 10.1073/pnas.93.23.13404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Browse J, Lightner J, McConn M. Membrane lipid structure and plant function: what are the relationships? In: Kader J-C, Mazliak P, editors. Plant Lipid Metabolism. Dordrecht, The Netherlands: Kluwer Academic Publishers; 1995. pp. 9–14. [Google Scholar]
- Browse J, McConn M, James D, Miquel M. Mutants of Arabidopsis deficient in the synthesis of α-linolenate. J Biol Chem. 1993;268:16345–16351. [PubMed] [Google Scholar]
- Chapman D. Phase transitions and fluidity characteristics of lipids and cell membranes. Q Rev Biophys. 1975;8:185–235. doi: 10.1017/s0033583500001797. [DOI] [PubMed] [Google Scholar]
- Clarkson DT, Hall KC, Roberts JKM. Phospholipid composition and fatty acid desaturation in the roots of rye during acclimatization of low temperature. Planta. 1980;149:464–471. doi: 10.1007/BF00385749. [DOI] [PubMed] [Google Scholar]
- Cronan JE, Gelmann EP. Physical properties of membrane lipids: biological relevance and regulation. Bacteriol Rev. 1975;39:232–256. doi: 10.1128/br.39.3.232-256.1975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de la Roche IA. Increase in linolenic acid is not a prerequisite for development of freezing tolerance in wheat. Plant Physiol. 1979;63:5–8. doi: 10.1104/pp.63.1.5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gibson S, Arondel V, Iba K, Somerville C. Cloning of a temperature-regulated gene encoding a chloroplast ω-3 desaturase from Arabidopsis thaliana. Plant Physiol. 1994;106:1615–1621. doi: 10.1104/pp.106.4.1615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guy CL. Cold acclimation and freezing stress tolerance: role of protein metabolism. Annu Rev Plant Physiol. 1990;41:187–223. [Google Scholar]
- Hamada T, Kodama H, Nishimura M, Iba K. Cloning of a cDNA encoding tobacco ω-3 fatty acid desaturase. Gene. 1994;147:293–294. doi: 10.1016/0378-1119(94)90084-1. [DOI] [PubMed] [Google Scholar]
- Hamada T, Kodama H, Nishimura M, Iba K. Modification of fatty acid composition by over- and antisense-expressions of a microsomal ω-3 fatty acid desaturase gene in transgenic tobacco. Transgenic Res. 1996;5:115–121. doi: 10.1007/BF01969429. [DOI] [PubMed] [Google Scholar]
- Hugly S, Somerville C. A role for membrane lipid polyunsaturation in chloroplast biogenesis at low temperature. Plant Physiol. 1992;99:197–202. doi: 10.1104/pp.99.1.197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iba K, Gibson S, Nishiuchi T, Fuse T, Nishimura M, Arondel V, Hugly S, Somerville C. A gene encoding a chloroplast ω-3 fatty acid desaturase complements alterations in fatty acid desaturation and chloroplast copy number of the fad7 mutant of Arabidopsis thaliana. J Biol Chem. 1993;268:24099–24105. [PubMed] [Google Scholar]
- Jost P, Libertini LJ, Hebert VC, Griffith OH. Lipid spin labels in lecithin multilayers: a study of motion along fatty acid chains. J Mol Biol. 1971;59:77–98. doi: 10.1016/0022-2836(71)90414-1. [DOI] [PubMed] [Google Scholar]
- Kasamo K, Sakakibara Y. The plasma membrane H+-ATPase from higher plants: functional reconstitution into liposomes and its regulation by phospholipids. Plant Sci. 1995;111:117–131. [Google Scholar]
- Kodama H, Hamada T, Horiguchi G, Nishimura M, Iba K. Genetic enhancement of cold tolerance by expression of a gene for chloroplast ω-3 fatty acid desaturase in transgenic tobacco. Plant Physiol. 1994;105:601–605. doi: 10.1104/pp.105.2.601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kodama H, Horiguchi G, Nishiuchi T, Nishimura M, Iba K. Fatty acid desaturation during chilling acclimation is one of the factors involved in conferring low-temperature tolerance to young tobacco leaves. Plant Physiol. 1995;107:1177–1185. doi: 10.1104/pp.107.4.1177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuiper PJC. Role of lipids in water and ion transport. Proc Phytochem Soc. 1974;12:359–386. [Google Scholar]
- Larsson C, Sommarin M, Widell S. Isolation of highly purified plant plasma membranes and separation of inside-out and right-side-out vesicles. Methods Enzymol. 1994;228:451–469. [Google Scholar]
- Lyons JM. Chilling injury in plants. Annu Rev Plant Physiol. 1973;24:445–466. [Google Scholar]
- Mazliak P. Desaturation processes in fatty acid and acyl lipid biosynthesis. J Plant Physiol. 1994;143:399–406. [Google Scholar]
- McCourt P, Kunst L, Browse J, Somerville CR. The effects of reduced amounts of lipid unsaturation on chloroplast ultrastructure and photosynthesis in a mutant of Arabidopsis. Plant Physiol. 1987;84:353–360. doi: 10.1104/pp.84.2.353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miquel M, James D, Dooner H, Browse J. Arabidopsis requires polyunsaturated lipids for low-temperature survival. Proc Natl Acad Sci USA. 1993;90:6208–6212. doi: 10.1073/pnas.90.13.6208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mitsuhara I, Ugaki M, Hirochika H, Ohshima M, Murakami T, Gotoh Y, Katayose Y, Nakamura S, Honkura R, Nishimiya S and others. Efficient promoter cassettes for enhanced expression of foreign genes in dicotyledonous and monocotyledonous plants. Plant Cell Physiol. 1996;37:49–59. doi: 10.1093/oxfordjournals.pcp.a028913. [DOI] [PubMed] [Google Scholar]
- Murashige T, Skoog F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant. 1962;15:473–497. [Google Scholar]
- Raison JK. Temperature-induced phase changes in membrane lipids and their influence on metabolic regulation. Symp Soc Exp Biol. 1973;27:485–512. [PubMed] [Google Scholar]
- Smolenska G, Kuiper PJC. Effect of low temperature upon lipid and fatty acid composition of roots and leaves of winter rape plants. Physiol Plant. 1977;41:29–35. [Google Scholar]
- Steponkus PL. Role of the plasma membrane in freezing injury and cold acclimation. Annu Rev Plant Physiol. 1984;35:543–584. [Google Scholar]
- Taussky HH, Shorr E. A microcolorimetric method for the determination of inorganic phosphorus. J Biol Chem. 1953;202:675–685. [PubMed] [Google Scholar]
- Uemura M, Joseph RA, Steponkus PL. Cold acclimation of Arabidopsis thaliana. Plant Physiol. 1995;109:15–30. doi: 10.1104/pp.109.1.15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Uemura M, Steponkus PL. A contrast of the plasma membrane lipid composition of oat and rye leaves in relation to freezing tolerance. Plant Physiol. 1994;104:479–496. doi: 10.1104/pp.104.2.479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Utsumi H, Inoue K, Nojima S, Kwan T. Interaction of spin-labeled lysophosphatidylcholine with rabbit erythrocytes. Biochemistry. 1978;17:1990–1996. doi: 10.1021/bi00603a030. [DOI] [PubMed] [Google Scholar]
- Willemot C. Simultaneous inhibition of linolenic acid synthesis in winter wheat roots and frost hardening by BASF 13-338, a derivative of pyridazinone. Plant Physiol. 1977;60:1–4. doi: 10.1104/pp.60.1.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wolfe J. Chilling injury in plants: the role of membrane. Plant Cell Environ. 1978;1:241–247. [Google Scholar]
- Yadav NS, Wierzbicki A, Aegerter M, Caster CS, Pérez-Grau L, Kinney AJ, Hitz WD, Booth JR, Schweiger B, Stecca KL and others. Cloning of higher plant ω-3 fatty acid desaturases. Plant Physiol. 1993;103:467–476. doi: 10.1104/pp.103.2.467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoshida S, Kawata T, Uemura M, Niki T. Properties of plasma membrane isolated from chilling-sensitive etiolated seedlings of Vigna radiata L. Plant Physiol. 1986;80:152–160. doi: 10.1104/pp.80.1.152. [DOI] [PMC free article] [PubMed] [Google Scholar]