

Abstract
Many foods and food components boost the immune system, but little data are available regarding the mechanisms by which they do. Bacterial strains have disparate effects in stimulating the immune system. Indendritic cells, the gram-negative bacteria Escherichia coli upregulates proinflammatory cytokines, whereas gram-positive Lactobacillus acidophilus induces a robust interferon (IFN)-β response. The immune-modulating effects of astragalus root and elderberry fruit extracts were examined in bone marrow-derived murine dendritic cells that were stimulated with L. acidophilus or E. coli. IFN-β and other cytokines were measured by ELISA and RT-PCR. Endocytosis of fluorescence-labeled dextran and L. acidophilus in the presence of elderberry fruit or astragalus root extract was evaluated in dendritic cells. Our results show that both extracts enhanced L. acidophilus-induced IFN-β production and slightly decreased the proinflammatory response to E. coli. The enhanced IFN-β production was associated with upregulation of toll-like receptor 3 and to a varying degree, the cytokines IL-12, IL-6, IL-1β and TNF-α. Both extracts increased endocytosis in immature dendritic cells, and only slightly influenced the viability of the cells. In conclusion, astragalus root and elderberry fruit extracts increase the IFN-β inducing activity of L. acidophilus in dendritic cells, suggesting that they may exert antiviral and immune-enhancing activity.
Introduction
Foods and food components with immune-boosting activity are constantly sought after [1]. Some of the most promising components are probiotic bacteria, which stimulate the immune system on oral administration [2], including Lactobacillus strains. Certain lactobacilli induce interferon (IFN)-β and interleukin (IL)-12 production in dendritic cells, which leads to upregulation of type 1 helper T cells (Th1) [3].IFN-β induces IL-12, TLR3 and other genes that mediate viral defense—culminating in robust stimulation of the adaptive immune system [4]. In addition to its strong antiviral effects and regulation of innate and adaptive immunity [4], IFN-β has anti-inflammatory properties, upregulating IL-1β receptor antagonist [5], which mitigates the proinflammatory effects of IL-1β [6]. Accordingly, the stimulation of the IFN-βproduction in DCs may boost the immune systems while simultaneously leading to reduction in low-grade inflammation in the tissue.
In contrast to L.acidophilus, although the gram-negative Escherichia coli strain Nissle 1917 also leads to IL-12 production, albeit at lower levels, it does not induce significant IFN-β production or upregulation of viral defense genes [3]. Thus, the induction of IL-12 may take place by two distinct pathways, indicating the importance of the system used to assess the immunomodulatory activity of bioactive food or food compounds. Dendritic cells are professional antigen-presenting cells that respond to a microbial signal through pathogen recognition receptors, such as toll-like receptors (TLRs), and orchestrate the responses of other immune cells [7], [8]. Thus, dendritic cells are model cells that can be used to assess the immune-modulating activities of food and food components. The responses that are generated in dendritic cells, particularly with regard to surface marker expression and cytokine production, depend on the cellular events that are induced by specific microorganisms [9].
Many of the claims for the immune-modulating activities of plants and plant components originated in traditional medicine, and only recently have such mechanisms been researched [10]. Reducing chronic inflammation in tissues and enhancing immune responses during viral and bacterial infections using food and food components, including complementary and alternative medicine, has received growing interest [11]. Astragalus, Astragalus propinquus S. syn. Astragalus membranaceus F. (Family Fabaceae), is a flowering plant that exists worldwide. Its membranous milk-vetch roots are used in traditional Chinese medicine as a powder or in a decoction to ameliorate general weakness and increase overall vitality [12]. Astragalus is being investigated with regard to its immune-stimulating properties. In human peripheral blood mononuclear (PBM) cells, astragalus root extract induces monocyte maturation [13].
Chemically, astragalus comprises polysaccharides, saponins, flavonoids (including calycosin and formononetin), amino acids, and trace elements. Saponins are primarily astragalosides [14]. Polysaccharide fractions of Astragalus root extract have also been isolated [14]–[17]. Whereas certain plant polysaccharides induce maturation of dendritic cells [18], they have not been reported to upregulate the Th1-polarizing cytokines IFN-β and IL-12. In general, low-molecular-weight compounds from plants are incapable of stimulating antigen-presenting cells, but they are often potent modulators of microbially induced antigen presentation [19], [20].
Elderberry, Sambucus nigra (Family Adoxaceae), is a shrub that grows in moist soil, bearing stems that reach up to 4 m high, and is found in Europe, west and central Asia, and north Africa. Its fruits are black-purple, edible, berry-like drupes [21] that have been traditionally used for their antiviral, antibacterial, and anti-inflammatory properties [21], [22], the primary constituents of which are flavonoids. Other major secondary metabolites include triterpenes, sterols, phenolic acids and their corresponding glycosides, and essential oil [22]. Several lectins have also been isolated from elderberry [23]–[25].
Although immune-modulating and -stimulating properties of specific probiotic bacteria and herbal extracts have been demonstrated, the effects of herbal extracts hitherto have been investigated by stimulation with LPS or other microbial ligands or plant derived mitogens and, to our knowledge no studies have reported the effects of probiotics and plant extracts in combination. As a result of a screening of the immunomodulatory activity of a large number of plant extracts we identified two extracts from astragalus root and elderberry, respectively, that were especially potent in enhancing the IL-12 production induced by L.acidophilus, while they reduced the IL-12 production induced by E.coli. Accordingly, in the present study we aimed to investigate the immunomodulatory effect of astragalus root and elderberry fruit extracts in dendritic cells that were stimulated with L. acidophilus or E. coli. We hypothesized that the disparate effects of the extracts on the two bacteria were due to an enhancing effect on the IFN-β response, which is almost absent in E.coli stimulated DCs [3].
Materials and Methods
Astragalus root and elderberry fruit extract preparation
Astragalus root and elderberry fruit extracts were produced in an industrial process at Naturex Inc, USA (Astragalus root extract reference: ED161024, Lot number: D184/012/D10 with a content of 50% Gum arabicum. Elderberry fruit extract reference: EA145100, batch number: 231/55/A9 with 65% maltodextrin as carrier material). Both extracts were analyzed at AINIA (Valencia, Spain) per the norms in vigor for microorganism content, including aerobic mesophiles (UNE-EN ISO 4833:2003), yeast, fungi (European pharmacopoeia 6.7, part 2.6.12), enterobacteriaceae (ISO 21528-2:2004), E. coli (ISO 21528-1:2004), Staphylococcus spp. (UNE EN-ISO 6888-1:2000), and Salmonella spp. (UNE EN-ISO 6579:2003); pesticides (European pharmacopoeia); and heavy metals (Cd, Pb, Hg, Ar). The microorganism content in both extracts did not exceed 1×103 CFU per g, and the amounts of pesticides and heavy metals were below the maximum residue limits [26].
For all analyses, 50 mg of the lyophilized samples was added to 5 mL deionized sterile water and incubated for at least 30 min on a shaker and centrifuged at 400 g for 10 min. The supernatants were transferred to new tubes and assayed on the same day.
Bacterial strains, growth conditions, and preparation of UV-killed bacteria
The gram-positive lactic acid bacterium Lactobacillus acidophilus NCFM (Danisco, Copenhagen, Denmark) was grown anaerobically overnight at 37°C in de Man Rogosa Sharpe (MRS) broth (Merck, Darmstadt, Germany) and subcultured twice. Cells were harvested by centrifugation at 2000 g for 15 min, washed twice in phosphate-buffered saline (PBS, Bio Whittaker, East Rutherford, NJ, USA), and resuspended in 1/10 of the growth volume of PBS. Because they affect the same stimulatory patterns as live bacteria, UV-killed bacteria were used to avoid growth during dendritic cell stimulation and ensure reproducibility. The bacteria were killed in a 20-min exposure to UV light.
Escherichia coli Nissle 1917 O6:K5:H1 (Statens Serum Institut, Copenhagen, Denmark), a gram-negative probiotic bacterium, was grown aerobically overnight at 37°C in Luria-Bertani (LB) broth (Merck) and killed by a 45-min exposure to UV light. The bacteria were stored at −80°C, their concentration was calculated as the content of dry matter per mL on lyophilization, and the dry weight was corrected for buffer salt content. The absence of viable cells was verified by plating the UV-exposed bacteria on MRS and LB agar.
Generation of murine dendritic cells
Bone marrow-derived dendritic cells were prepared as described [9]. Briefly, bone marrow from C57BL/6 mice was flushed from the femur and tibia and washed twice in sterile PBS. Bone marrow cells (3×105) were used to seed Petri dishes in 10 mL RPMI 1640 (Sigma-Aldrich, St. Louis, MO), containing 10% (v/v) heat-inactivated fetal calve serum that was supplemented with penicillin (100 U/mL), streptomycin (100 µg/mL), glutamine (4 mM), 50 µM 2-mercaptoethanol (all purchased from Cambrex Bio Whittaker, Charles City, IA), and 15 ng/mL murine granulocyte-macrophage colony-stimulating factor (GM-CSF) added as <1% culture supernatant harvested from the GM-CSF-tranfected Ag8.653 cell line [27]; which was kindly provided by Dr. R. Tisch (University North Carolina, Chapel Hill, NC).
All animals used as source of bone marrow cells were housed under conditions approved by the Danish Animal Experiments Inspectorate (Forsøgdyrstilsynet) and experiments were carried out in accordance with the guidelines of ‘The Council of Europe Convention for the Protection of Vertebrate Animals used for Experimental and other Scientific Purposes’. Since the animals were employed as a source of cells, and that no animals were used for the experiments, no specific approval was required for this study. Hence, the animals used for this study are included in the general facility approval for the faculty of Life Science, University of Copenhagen.
The cells were incubated for 8 days at 37°C in a 5% CO2 humidified atmosphere. On Day 3, 10 mL of complete medium, containing 15 ng/mL GM-CSF, was added. On Day 6, 10 mL of medium was removed and replaced with fresh medium. Nonadherent immature dendritic cells were harvested on Day 8 and examined for purity by CD11c immunostaining and flow cytometry.
Immunostaining and flow cytometry
Dendritic cells were harvested and resuspended in cold PBS that contain 1% (v/v) fetal bovine serum, 0.15% (w/v) sodium azide (PBS-Az), and anti-mouse FcγRII/III (3 µg/mL, BD Biosciences, San Jose, CA, USA) to block the nonspecific binding of antibody reagents. The cells were stained with phycoerythrin (PE)-conjugated anti-mouse MHCII, allophycocyanin (APC)-conjugated anti-mouse CD86, and PE-conjugated anti-mouse CD11c (Southern Biotech, Birmingham, AL) and analyzed on a BD FACS array flow cytometer (BD Biosciences) in 10,000-cell counts. The geometric mean fluorescence intensity (MFI) was determined, reflecting the level of expression. To assess the viability of the cells on stimulation, 7-AAD (Pharmingen, CA, USA) was added to each sample for the last 10 minutes of the incubation with antibodies.
Cell treatments and stimulation with bacteria
Immature dendritic cells (2×106 cells/mL) were resuspended in fresh medium that was supplemented with 10 ng/mL GM-CSF, and 500 µL/well was used to seed 48-well tissue culture plates (Nunc, Roskilde, Denmark). Cells (106) were treated with herbal extracts at final dilutions of the reconstituted extracts of 1∶18, 1∶60, 1∶180, and 1∶600 (corresponding to the concentrations 500, 150, 50 and 15 µg/mL, respectively).
Dendritic cells were treated with the herbal extract and incubated for 30 minutes prior to the addition of L. acidophilus or E. coli, which were suspended in medium and added (100 µL/well) to a final concentration of 10 µg/mL. Optimal bacterial concentrations were determined in a previous study [28]. The cell cultures were incubated at 37°C in 5% CO2 for 20 hr, and supernatants were harvested for cytokine quantification. In the PCR analysis experiments, the cells were harvested 2–10 hours after stimulation. The two carrier materials, gum arabicum and maltodextrin were tested for immunomodulatory effects in the cellular assay per se, but showed no immunomodulatory activity.
RNA extraction
Murine dendritic cells were harvested after stimulation and homogenized using a QIAshredder (Qiagen, Ballerup, Denmark), and RNA was extracted using the MagMAX-96 Total RNA Isolation kit (AM1830, Applied Biosystem) on a MagMAX Express Magnetic Particle Processor (Applied Biosystem). RNA concentration was measured on a Nanodrop (Thermo, Wilmington, USA).
Quantitative Real-Time PCR
One microgram of total RNA was reverse-transcribed using the TaqMan Reverse Transcription Reagent kit (Applied Biosystems, Foster City, USA) with random hexamer primers per the manufacturer's instructions. The resulting cDNA was stored in aliquots at −80°C. To design the primer and probe sequences, we retrieved the regions that encoded for the genes of interest from the GenBank EMBL databases: TLR-3 (NM_126166), IFN-β (NM_010510), IL-12 p40 (NM_008352), IL-10 (NM_010548), and beta actin (NM_007393). Primers and probes were designed using Primer Express 3.0 (Applied Biosystems) and tested for specificity using BLAST. HPLC-purified forward and reverse primers were manufactured by DNA Technology (Aarhus, Denmark).
The probes were labeled with the 5′ reporter 6-carboxy-fluorescein (FAM) and the 3′ quencher NFQ-MGB (Applied Biosystems). The sequences of the primers and probes are listed in Table 1. Primer and probe concentrations were optimized, and standard curves were generated for each set of primers and probe to determine the efficiency of the amplification for each dilution, (data not shown).
Table 1. Real-Time PCR primers and probes.
| Target | Primers and probes | Sequences (5′-3′) |
| IFN-β (NM_010510) | Forward | CCCTATGGAGATGACGGAGAAG |
| Reverse | GAGCATCTCTTGGATGGCAAA | |
| Probe | TGCAGAAGAGTTACACTGC | |
| IL-12 p40 (NM_008352) | Forward | GCCAGTACACCTGCCACAAA |
| Reverse | TGTGGAGCAGCAGATGTGAGT | |
| Probe | AGGCGAGACTCTG | |
| IL-10 (NM_010548) | Forward | AGCATGGCCCAGAAATCAAG |
| Reverse | CGCATCCTGAGGGTCTTCAG | |
| Probe | AGCATTTGAATTCCCTGGGT | |
| β-actin (NM_007393) | Forward | CGATGCCCTGAGGCTCTTT |
| Reverse | TGGATGCCACAGGATTCCA | |
| Probe | CCAGCCTTCCTTCTT |
The amplifications were performed in 20 µL, containing 1×TaqMan Universal PCR Master Mix (Applied Biosystems), forward and reverse primer (concentration 900 nM each), 200 nM TaqMan MGB probe, and purified target cDNA. The cycling program comprised 20 sec at 95°C and 40 cycles of 3 sec at 95°C and 30 sec at 60°C, run on an ABI Prism 7500 (Applied Biosystems). Amplifications were performed in triplicate, and DNA contamination controls were included. The reactions were normalized to beta actin. Relative transcript levels were calculated per Pfaffl [29].
Cytokine ELISA
After 20 hr of stimulation, culture supernatants were collected and stored at −80°C for cytokine analysis. Murine IL-12(p70), IL-10, IL-6, TNF-α, IL-1β, and IFN-β were analyzed using commercial ELISA kits (R&D Systems, Minneapolis, USA).
Endocytotic activity
Fluorescein isothiocyanate (FITC)-conjugated dextran (10 kD) was obtained from Sigma-Aldrich (St. Louis, MO). L. acidophilus was labeled with Alexa Fluor 647 per the supplier's protocol. Briefly, 1 volume ofAlexa Fluor 647, solubilized in DMSO (5 mg/mL), was added to 9 volumes of bacteria in 0.1 M sodium carbonate buffer, pH 9. After 1 hour of incubation, the bacteria was washed 3 times, Dendritic cells were incubated with elderberry fruit or astragalus root extract or stimulated with 1 µg/mL lipopolysaccharide (LPS, E. coli, Sigma-Aldrich) prior to the addition of fluorescence-labeled dextran or L. acidophilus. Endocytosis was measured by flow cytometry and was expressed as the percentage of cells that endocytosed fluorescence-labeled dextran or L. acidophilus or the amount of endocytosed dextran/bacteria per cell, based on mean florescence intensity (MFI).
Statistical analysis
Statistical analyses were performed by Prof. Alberto José Ferrer-Riquelme and Ass. Prof. José Manuel Prats-Montalbán, Multivariate Statistical Engineering Research Group, Department of Applied Statistics, Operations Research and Quality, Universidad Politécnica de Valencia, using SGWIN (Statgraphics Plus for Windows version 5; Statistical Graphics, Rockville, MD). The results of each experiment were analyzed by ANOVA. We assumed a type I risk α of 0.05, and the difference between groups was evaluated using the least significance difference (LSD) intervals (95% confidence level). The difference between 2 groups was considered significant when the LSD intervals did not overlap (p-value<0.05).
Results
Extracts of astragalus root or elderberry fruit were added at dilutions 1∶18 to 1∶600 (corresponding to approximately 10, 50, 100 and 500 µg/mL dry matter, or from around 4 to 250 µg/mL pure extract) to dendritic cells 30 min before stimulation with L. acidophilus or E. coli,s or at 500 µg/mL to unstimulated cells. IL-12 (Figure 1A, 1B), TNF-α (Figure 2A, 2B) and IL-6 (Figure 2 E, 2F) levels increased in dendritic cells to the same extent on stimulation with E. coli and L. acidophilus. Conversely, IL-10 (Figure 1C, 1D) and IL-1β (Figure 2C, 2D) rose significantly higher with E. coli versus L. acidophilus (p<0.05). In immature dendritic cells, 500 µg/mL of extract did not affect IL-10 and IL-12 production and slightly increased IL-6 and TNF-α levels, albeit to a lesser extent than microbially induced cytokine production, except for IL-1β, which rose with elderberry fruit extract to comparable levels as with L. acidophilus, while astragalus root extract alone had no significant effects (data not shown).
Figure 1. Astragalus and elderberry extracts modulate IL-12 and IL10 production in dendritic cells.

Astragalus root (A, C) and elderberry fruit (B, D) extracts enhance IL-12 (A, B) and IL-10 (C, D) production when added to murine bone marrow-derived dendritic cells at concentrations from 15 to 500 µg/mL 30 min prior to stimulation with L. acidophilus (10 µg/mL) but not with E. coli (10 µg/mL). Supernatants were harvested for ELISA after 20 hr incubation. The results are representative of at least 3 experiments. Error bars are representative of the LSD intervals. The difference between 2 groups was considered significant when the LSD intervals did not overlap (p-value<0.05).
Figure 2. Astragalus and elderberry extracts modulate TNF-α, IL-1β, and IL-6 in dendritic cells.

Astragalus root (A, C, E) and elderberry fruit (B, D, F) extracts differentially modulate TNF-α (A, B), IL-1β (C, D), and IL-6 (E, F) production when added to murine bone marrow-derived dendritic cells at concentrations from 15 to 500 µg/mL 30 min prior to the addition L. acidophilus (10 µg/mL) or E. coli (10 µg/mL). Supernatants were harvested for ELISA after 20 hr incubation. The results are representative of at least 3 experiments. Error bars are representative of the LSD intervals. The difference between 2 groups was considered significant when the LSD intervals did not overlap (p-value<0.05).
After incubation with astragalus root or elderberry fruit extract and stimulation with E. coli, the production of IL-12 (Figure 1A, 1B) in dendritic cells fell dose-dependently, by 50% at the concentration 500 µg/mL. IL-10 production was unaffected by these extracts in E. coli-stimulated dendritic cells (Figure 1C, 1D).
In contrast, when the extracts were added prior to stimulation with L. acidophilus, IL-12 rose significantly. At the highest concentrations of extracts (50 to 500 µg/mL), IL-12 production increased by more than 2-fold significantly higher than the effects of L. acidophilus alone (p<0.05). Similarly, IL-10 levels climbed significantly—by 2-fold when L. acidophilus was incubated with astragalus root or elderberry fruit extract at 150 to 500 µg/mL (Figure 1C, 1D)) (p<0.05). The IL-12 stimulating effect of the two extracts lasted for at least up to two hours, while after 3 hours of incubation with extract prior to stimulation with L.acidophilus, no significant effect of the extract was observed (data not shown). The immune modulating effects were not due to physical interactions or intermolecular complex formation as washing the cells after incubation with plant extract and prior to stimulation with bacteria did not alter the immune modulatory profile of the extracts (data not shown).
The production of the proinflammatory cytokines TNF-α, IL-1β, and IL-6 is shown in Figure 2. Whereas the extracts inhibited E. coli- and increased L. acidophilus-induced TNF-α slightly (p<0.05, Figure 2A, 2B), they had robust effects on IL-1β and IL-6. Notably, L. acidophilus-induced IL-1β levels were enhanced significantly—up to 6-fold when dendritic cells were coincubated with 500 µg/mL elderberry fruit extract (p<0.05, Figure 2D) and up to 3-fold when coincubated with 500 µg/mL astragalus root extract (p<0.05, Figure 2C).
In contrast, astragalus root extract inhibited IL-1β production when added to E. coli-stimulated dendritic cells (Figure 2 C, p<0.05 with the dose 500 µg/mL astragalus root extract versus E.coli alone), whereas elderberry fruit extract did not affect such. IL-6 levels increased significantly, up to 2-fold, when astragalus root and elderberry fruit extract at the highest concentration (500 µg/mL) were incubated with L. acidophilus-stimulated dendritic cells (p<0.05). In E. coli stimulated dendritic cells, incubation with astragalus root or elderberry fruit extracts at the lowest concentration also increased by 2-fold IL-6 production (p<0.05). Increasing the concentration of the extract did not have a significant effect, suggesting active compounds with counteracting effects may have been present. The addition of the extract slightly affected the viability of the dendritic cells (figure 3). Both immature and bacteria stimulated cells had a viability of 86.5+/−2%. Addition of the extracts only slightly affected the viability of the E.coli and L.acidophilus stimulated cells (2.3–5.4% decrease in the highest concentration, while added in lower concentrations, a slight decrease in viability was only seen for elderberry and L.acidophilus stimulated cells (2.5%)). Interestingly, the extracts increased the viability of the unstimulated cells, even when added at the lowest concentration. Taken together, these results demonstrate that Astragalus root and Elderberry extract both modulated the cytokine responses to E.coli and L.acidophilus differently and that this is not due to toxic effects of the extracts.
Figure 3. Astragalus root and Elderberry extracts only slightly affect the viability of DCs.
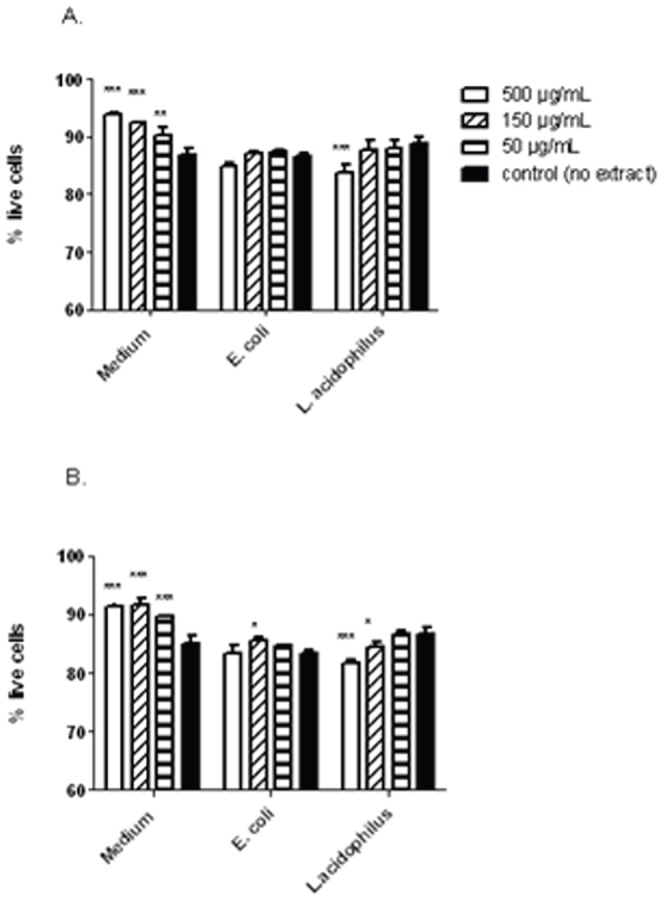
DCs were incubated with increasing concentrations of the astragalus extract (A) and elderberry extract (B) and left unstimulated or stimulated with E.coli Nissle (10 µg/mL) or L.acidophilus NCFM (10 µg/mL). After 20 hr the cells were harvested and stained for viability with 7AAD for 15 min., washed and analysed by flow cytometry. (*: p<0.05; **: p<0.01; ***: p<0.001).
We have demonstrated that much of the IL-12 that is induced by L. acidophilus, but not E. coli, is attributed to strong induction of IFN-β, which, by binding the IFN-α receptor (IFNAR), upregulates IL-12 and other genes that mediate viral defense and stimulation of Th1 responses. Thus, we hypothesized that the IL-12-enhancing effects of astragalus root and elderberry fruit extracts are the result of enhanced Ifnβ transcription.
Astragalus root (Figure 4A) and elderberry fruit (Figure 4B) extract dose-dependently enhanced IFN-β levels, as measured in cell culture supernatants after 20 hr in response to L. acidophilus (p<0.05). Ifnβ transcription was unaffected by elderberry fruit extract alone but increased significantly when the extract was added to L. acidophilus-stimulated cells at 4 and 6 hr compared with L. acidophilus alone (p<0.05, Figure 5A). Tlr3 expression rose at 4 and 6 hr in cells that were stimulated with elderberry fruit extract and L. acidophilus versus L. acidophilus alone; at 10 hr after stimulation, the expression in the 2 samples was comparable (Figure 5B).
Figure 4. Astragalus and elderberry extracts potentiate the IFN-β-inducing capacity of L. acidophilus in dendritic cells.

Extracts of astragalus root (A) and elderberry fruit (B) were added to murine bone marrow-derived dendritic cells at the concentrations 15 and 150 µg/mL prior to the addition of L. acidophilus (10 µg/mL) and incubated at 2, 4, 10, and 20 hr. Supernatants from the cells that were incubated for 20 hr were harvested, and IFN-β was measured by ELISA. Error bars are representative of the LSD intervals. The difference between 2 groups was considered significant when the LSD intervals did not overlap (p-value<0.05).
Figure 5. Elderberry extract potentiates Ifn-β, Trl3, Il10 and Il12 gene expression by L. acidophilus in dendritic cells.

Extract of elderberry fruit was added to murine bone marrow-derived dendritic cells at the concentration 500 µg/mL prior to the addition of L. acidophilus (10 µg/mL) and incubated at 2, 4, 10, and 20 hr. RNA was harvested at various time points and analyzed for Ifnβ, (A), Tlr3 (B), Il12 (C) and Il10 (D) expression by RT-PCR relative to β-actin. Cells only stimulated with L.acidophilus or elderberry extract were used as controls. Error bars are representative of the LSD intervals. The difference between 2 groups was considered significant when the LSD intervals did not overlap (p-value<0.05).
Similarly, Il10 (Figure 5C) and Il12 (Figure 5D) expression increased significantly with the elderberry and L. acidophilus combination versus L. acidophilus alone, peaking at 6 and 24 hr, respectively. In summary, the increase in IFN-β demonstrates that astragalus root and elderberry fruit extracts contain components that upregulate Ifnβ, potentially enhancing the production of viral defense genes that are induced by the IFN-β/IFNAR pathway in response to L. acidophilus.
Immature dendritic cells constantly survey the environment by endocytosing external fluid. We examined the effects of elderberry fruit and astragalus root extract on the endocytotic activity of dendritic cells by measuring the uptake of FITC-conjugated dextran, which are inert nanoparticles that do not activate pathogen recognition receptors on dendritic cells, and L. acidophilus, the uptake of which is facilitated by ligation of bacterial components to various receptors on dendritic cells.
Elderberry fruit and astragalus root extract significantly enhanced endocytotic activity in unstimulated (immature) cells, as evidenced by uptake of FITC-conjugated dextran, versus control (p<0.05) (Figure 6A, 6B). This increase, however, was less robust than the effect of LPS. The addition of fluorescence-labeled L. acidophilus to cells that were incubated with elderberry fruit or astragalus root extract also increased the percentage of fluorescent cells (p<0.05) but not their mean fluorescence intensity (Figure 6C, 6D).
Figure 6. Elderberry and astragalus increase dextran and L. acidophilus uptake in immature dendritic cells.

Immature murine bone marrow-derived dendritic cells were incubated with elderberry fruit extract, astragalus root extract, or LPS for 30 min prior to the addition of FITC-labeled 10 kD-dextran molecules (A, B) or Alexa Fluor 647-labeled L. acidophilus (C, D). Results are expressed as % of fluorescent cells (A, C) and as mean fluorescence intensity (MFI) (B, D). Error bars are representative of the LSD intervals. The difference between 2 groups was considered significant when the LSD intervals did not overlap (p-value<0.05). Lower panel show examples of histograms of which A–B were made; Fitc-dextran (left) or Alexa Fluor 647-conjugated L.acidophilus (right) incubated cells with or without prior incubation with elderberry fruit extract.
Discussion
Certain foods and food components are traditionally used to maintain or improve immune activity, but their mechanisms of action remain to be determined. In this study, the modulatory effects of 2 botanical extracts on dendritic cells that were stimulated with E. coli or L. acidophilus were examined. Astragalus root and elderberry fruit extracts modulated immune cell responses, albeit disparately, depending on the bacteria. Both extracts enhanced L. acidophilus-induced production of IL-12 and mitigated IL-12 upregulation by E. coli. These results reflect the differential responses of these extracts to various microbes and demonstrate the Th1-skewing effects of astragalus root and elderberry fruit extracts. To our knowledge, this is the first time the effect of plant extracts on gram-positive and gram-negative bacteria stimulated dendritic cells has been compared and stresses the importance of the choice of stimuli for the outcome of such investigations.
Astragalus root and elderberry fruit extracts per se slightly upregulated the proinflammatory cytokines IL-6 and TNF-α in dendritic cells and elderberry fruit extract also induced IL-1β production. Generally, however, these responses are substantially lower than those to bacterial stimulation. Moreover, no IL-10 or IL-12 production was induced. As IL-10 and IL-12 are both induced in dendritic cells by the presence of even minor LPS contaminations [30], we can exclude significant LPS contaminations in the two extracts.
As demonstrated, the gram-negative bacterium E. coli induces IL-10 production and upregulates surface markers [31], whereas the gram-positive L. acidophilus induces IFN-β production, in turn upregulating genes that activate adaptive immunity and viral defense [3], [32]. Hence, disparate signaling pathways appear to be induced by the 2 bacteria, which might explain the differential effects of the extracts on the responses to the microbes. IFN-β has anti-inflammatory activity; type 1 IFN signaling through IFNAR and STAT1 inhibits IL-1β through upregulation of IL-10 production and repression of inflammasome activity [33]. In addition, IFN-β induces the production of IL-1β receptor antagonist in vivo, which inhibits IL-1β production [5]. The effects of the extracts on the cellular responses cannot be caused by a cytotoxic effect on the cells as the extracts only in the highest concentration showed a slight effect on the viability.
The viability of the cells was actually slightly increased in cells only treated with extracts and no bacteria, and cells stimulated with bacteria showed less viability, indicating that the presence of extracts or bacteria only have a minimal cytotoxic effect on the dendritic cells.
The response to L. acidophilus, but not E. coli, is associated with robust IFN-β synthesis [3]; thus, we expected to observe a reduction in IL-1β after adding IFN-β-enhancing extracts to L. acidophilus-stimulated cells. However, incubation of L. acidophilus in dendritic cells with the botanical extracts increased IFN-β, IL-10, and IL-1β production, indicating that IL-1β is induced by a different pathway on L. acidophilus stimulation that is not negatively affected by IFN-β or IL-10. Nevertheless, IFN-β production might still affect that of IL-1β by other cells.
The modulation of cytokine production by astragalus root and elderberry fruit extracts might be attributed to the activation of different signaling pathways in dendritic cells, the mechanisms of which are partially understood. Further, this immunomodulation might be caused by a specific component of the botanical extract or by synergistic effects of several components.
Although we did not address the identification of the active compounds in the present study it is tempting to speculate which compounds that might be involved. Polysaccharide fractions of astragalus root extracts contain bioactive components that influence the immune response [14]–[17]. Several nonstarch plant polysaccharides modulate dendritic cells. β-glucans and galactomannan guar gum synergistically increase IL-10 and TNF-α production and suppress IL12p70 on lipopolysaccharide (LPS) stimulation [30]. Thus, in our set-up, the combination of astragalus polysaccharides and E.coli, which containd LPS, might cause the IL-12 reducing effects. As regards the stimulating effect of the combination of astragalus extract and L.acidophilus, it was recently shown that the humoral immune response was enhanced upon administration of a combination of astragalus polysaccharides and lactobacillus [34]. Likewise, several immune modulating components in elderberry fruit extracts have been identified, including anthocyanins [21] and lectins [23], [25].
Identification of the active compound(s) of the two extracts, in particular of the IFN-β enhancing compounds, is however, not the scope of this study, which aimed to assess the effect of plant extracts on dendritic cells stimulated with intact bacteria. To this end, we have previously shown that in particular whole gram-positive bacteria induce distinct responses in dendritic cells as compared to bacterial ligands such as peptidoglycan and lipoteichoic acid [28]. Accordingly, our study is not comparable with previous studies using peptidoglycan or other bacterial cell wall constituents as stimulating ligands.
Overall, the 2 extracts have similar effects, despite their distinct content of putative bioactive compounds. In an initial screen, many other botanical extracts did not have immune-enhancing effects (unpublished data). Nevertheless, we cannot exclude the possibility that a specific component present in both extracts exerts the IFN-β enhancing effect, a hypothesis that remains to be examined.
Both extracts increased the uptake of the relatively small (10 kD) and inert dextran particles, as well as whole bacteria that express ligands for TLRs and other pathogen recognition receptors. But, whereas LPS, which enhances endocytotic activity in dendritic cells [35], had a stronger effect on the endocytosis of dextran, the 2 extracts were as potent as LPS in increasing the uptake of L. acidophilus. This enhancement manifested as an increase in the number of cells that took up fluorescently labeled bacteria, whereas that of dextran was observed as increases in the number of cells and the amount of endocytosed dextran. This disparity might be explained by the underlying endocytotic mechanisms of such diverse entities as dextran and L. acidophilus. In accordance with our previous findings of the importance of the endocytotic pathway for the cytokine response [3], these data could indicate that the extracts affect the cytokine signaling by affecting the uptake of L.acidophilus.
We used extract concentrations in the range of 15–500 µg/mL, but as these commercial extracts were stabilized with either maltodextrin (elderberry) or gum arabicum (astragalus root), the real extracts concentration used in the cell experiemens were actually 7–250 µg/mL(astragalus root) and 5–170 µg/ml (elderberry fruit), respectively. The two carrier materials did not per se affect the cell responses (data not shown). In general, we found that extract concentrations down to around 20–25 µg/mL excerted significant effects on the cytokine production. Whether these concentrations are too high to be realistic in vivo cannot be concluded as long as the concentration of the active compound(s) is not known. Although we acknowledge that the bioavailability of the active compounds are of major importance in order to be able to understand the impact of our presented data, the identity of the compounds as well as the amounts present in the extracts have to be established before we can relate our results to in vivo conditions.
In conclusion, astragalus root extract and elderberry fruit extract enhance the production of Th1-skewing cytokines, IL-12 and IFN-β, in murine bone marrow-derived dendritic cells that have been stimulated by L. acidophilus but not E. coli. This is the first study to demonstrate distinct immunomodulatory effects of plant extract in combination with stimulating bacteria and also the first report of the enhancing activity of plant extracts on the L.acidophilus induced IL-12 and IFN-β production. These extracts may find clinical use in boosting immunity and preventing viral infections, but the active compounds remain to be identified.
Acknowledgments
We would like to thank Maria Angeles Pasamar and Nicolas Feuillère (NATUREX Spain) for performing quality control of the extracts, Dr. Antoine Bily (NATUREX USA) for producing the extracts, and Anni Mehlsen for invaluable technical assistance in the cell culture experiments.
Funding Statement
Naturex is involved in the research/development and marketing/sales of astragalus root and elderberry fruit extracts as ingredients for the food, cosmetic, and nutraceutical industries. Therefore, Naturex has a commercial interest in this publication. The University of Copenhagen was paid by Naturex to perform and report the scientific work that formed the basis of this publication. The University of Copenhagen and Naturex declare that the data in this report represent a true and faithful representation of the work that has been performed. The financial assistance from Naturex is gratefully acknowledged. The study was designed in collaboration between Naturex and the University of Copenhagen, data collection and analysis was conducted by the University of Copenhagen, the decision to publish and preparation of the manuscript was done in collaboration between the two institutions.
References
- 1. Cummings JH, Antoine JM, Azpiroz F, Bourdet-Sicard R, Brandtzaeg, et al. (2004) PASSCLAIM–gut health and immunity. Eur J Nutr 43 Suppl 2: II118–II173. [DOI] [PubMed] [Google Scholar]
- 2. vanBaarlen P, Troost F, van der Meer C, Hooiveld G, Boekschoten M, et al. (2011) Human mucosal in vivo transcriptome responses to three lactobacilli indicate how probiotics may modulate human cellular pathways. Proc Natl Acad Sci U S A 108 Suppl 1: 4562–4569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Weiss G, Rasmussen S, Zeuthen LH, Nielsen BN, Jarmer H, et al. (2010) Lactobacillus acidophilus induces virus immune defence genes in murine dendritic cells by a Toll-like receptor-2-dependent mechanism. Immunology 131: 268–281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Katze MG, He Y, Gale M Jr (2002) Viruses and interferon: a fight for supremacy. Nat Rev Immunol 2: 675–687. [DOI] [PubMed] [Google Scholar]
- 5. Sciacca FL, Canal N, Grimaldi LM (2000) Induction of IL-1 receptor antagonist by interferon beta: implication for the treatment of multiple sclerosis. J Neurovirol 6 Suppl 2: S33–S37. [PubMed] [Google Scholar]
- 6. Jungo F, Dayer JM, Modoux C, Hyka N, Burger D (2001) IFN-beta inhibits the ability of T lymphocytes to induce TNF-alpha and IL-1beta production in monocytes upon direct cell-cell contact. Cytokine 14: 272–282. [DOI] [PubMed] [Google Scholar]
- 7. Banchereau J, Steinman RM (1998) Dendritic cells and the control of immunity. Nature 392: 245–252. [DOI] [PubMed] [Google Scholar]
- 8. Iwasaki A, Medzhitov R (2004) Toll-like receptor control of the adaptive immune responses. Nat Immunol 5: 987–995. [DOI] [PubMed] [Google Scholar]
- 9. Christensen HR, Frokiaer H, Pestka JJ (2002) Lactobacilli differentially modulate expression of cytokines and maturation surface markers in murine dendritic cells. J Immunol 168: 171–178. [DOI] [PubMed] [Google Scholar]
- 10. Pan MH, Lai CS, Dushenkov S, Ho CT (2009) Modulation of inflammatory genes by natural dietary bioactive compounds. J Agric Food Chem 57: 4467–4477. [DOI] [PubMed] [Google Scholar]
- 11. Lien EJ, Lien LL, Wang J (2010) Longevity depends on a balance between proinflammatory and anti-inflammatory factors: use of TCMS and natural products. Curr Drug Discov Technol 7: 13–21. [DOI] [PubMed] [Google Scholar]
- 12. Anonymous (2003) Astragalus membranaceus. Monograph. Altern Med Rev 8: 72–77. [PubMed] [Google Scholar]
- 13. Denzler KL, Waters R, Jacobs BL, Rochon Y, Langland JO (2010) Regulation of inflammatory gene expression in PBMCs by immunostimulatory botanicals. PLoS One 5: e12561 10.1371/journal.pone.0012561 [doi]. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Liu J, Hu X, Yang Q, Yu Z, Zhao Z, et al. (2010) Comparison of the immunoregulatory function of different constituents in radix astragali and radix hedysari. J Biomed Biotechnol 2010: 479426 10.1155/2010/479426 [doi]. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Liu QY, Yao YM, Zhang SW, Sheng ZY (2010) Astragalus polysaccharides regulate T cell-mediated immunity via CD11c(high)CD45RB(low) DCs in vitro. J Ethnopharmacol S0378–8741. [DOI] [PubMed] [Google Scholar]
- 16. Mao XF, Piao XS, Lai CH, Li DF, Xing JJ, et al. (2005) Effects of beta-glucan obtained from the Chinese herb Astragalus membranaceus and lipopolysaccharide challenge on performance, immunological, adrenal, and somatotropic responses of weanling pigs. J Anim Sci 83: 2775–2782. [DOI] [PubMed] [Google Scholar]
- 17. Shao BM, Xu W, Dai H, Tu P, Li Z, et al. (2004) A study on the immune receptors for polysaccharides from the roots of Astragalus membranaceus, a Chinese medicinal herb. Biochem Biophys Res Commun 320: 1103–1111. [DOI] [PubMed] [Google Scholar]
- 18. Takei M, Tachikawa E, Hasegawa H, Lee JJ (2004) Dendritic cells maturation promoted by M1 and M4, end products of steroidal ginseng saponins metabolized in digestive tracts, drive a potent Th1 polarization. Biochem Pharmacol 68: 441–452. [DOI] [PubMed] [Google Scholar]
- 19. Puertollano MA, Puertollano E, de Cienfuegos GA, de Pablo MA (2011) Dietary antioxidants: immunity and host defense. Curr Top Med Chem 11: 1752–1766. [DOI] [PubMed] [Google Scholar]
- 20. Rogers J, Perkins I, van OA, Burdash N, Klein TW, et al. (2005) Epigallocatechin gallate modulates cytokine production by bone marrow-derived dendritic cells stimulated with lipopolysaccharide or muramyldipeptide, or infected with Legionella pneumophila. Exp Biol Med (Maywood) 230: 645–651 230/9/645 [pii]. [DOI] [PubMed] [Google Scholar]
- 21. Anonymous (2005) Monograph. Sambucus nigra (elderberry). Altern Med Rev 10: 51–54. [PubMed] [Google Scholar]
- 22. Vlachojannis JE, Cameron M, Chrubasik S (2010) A systematic review on the sambuci fructus effect and efficacy profiles. Phytother Res 24: 1–8. [DOI] [PubMed] [Google Scholar]
- 23. Mumcuoglu M, Manor D, Slavin S (1986) Enrichment for GM-CFU from human bone marrow using Sambucus nigra agglutinin: potential application to bone marrow transplantation. Exp Hematol 14: 946–950. [PubMed] [Google Scholar]
- 24. Mach L, Kerschbaumer R, Schwihla H, Glossl J (1996) Elder (Sambucus nigra L.)-fruit lectin (SNA-IV) occurs in monomeric, dimeric and oligomeric isoforms. Biochem J 315: 1061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Girbes T, Citores L, De Benito FM, Inglesias R, Ferreras JM (1996) A non-toxic two-chain ribosome-inactivating protein co-exists with a structure-related monomeric lectin (SNA III) in elder (Sambucus nigra) fruits. Biochem J 315: 343–344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Durgnat JM, Heuser J, Andrey D, Perrin C (2005) Quality and safety assessment of ginseng extracts by determination of the contents of pesticides and metals. Food Addit Contam 22: 1224–1230. [DOI] [PubMed] [Google Scholar]
- 27. Lutz MB, Kukutsch N, Ogilvie AL, Rössner S, Koch F, et al. (1999) An advanced culture method for generating large quantities of highly pure dendritic cells from mouse bone marrow. J Immunol Methods 223 (1) 77–92. [DOI] [PubMed] [Google Scholar]
- 28. Zeuthen LH, Fink LN, Frokiaer H (2008) Toll-like receptor 2 and nucleotide-binding oligomerization domain-2 play divergent roles in the recognition of gut-derived lactobacilli and bifidobacteria in dendritic cells. Immunology 124: 489–502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Pfaffl MW (2001) A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Research 29: 2002–2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Wismar R, Brix S, Laerke HN, Frokiaer H (2011) Comparative analysis of a large panel of non-starch polysaccharides reveals structures with selective regulatory properties in dendritic cells. Mol Nutr Food Res 55: 443–454. [DOI] [PubMed] [Google Scholar]
- 31. Zeuthen LH, Christensen HR, Frokiaer H (2006) Lactic acid bacteria inducing a weak interleukin-12 and tumor necrosis factor alpha response in human dendritic cells inhibit strongly stimulating lactic acid bacteria but act synergistically with gram-negative bacteria. Clin Vaccine Immunol 13: 365–375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Weiss G, Rasmussen S, Nielsen FL, Jarmer H, Nohr NB, et al. (2010) Bifidobacterium bifidum actively changes the gene expression profile induced by Lactobacillus acidophilus in murine dendritic cells. PLoS One 5: e11065 10.1371/journal.pone.0011065 [doi]. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Guarda G, Braun M, Staehli F, Tardivel A, Mattmann C, et al. (2011) Type I interferon inhibits interleukin-1 production and inflammasome activation. Immunity 34: 213–223. [DOI] [PubMed] [Google Scholar]
- 34. Li SP, Zhao XJ, Wang JY (2009) Synergy of Astragalus polysaccharides and probiotics (Lactobacillus and Bacillus cereus) on immunity and intestinal microbiota in chicks. Poult Sci 88: 519–525. [DOI] [PubMed] [Google Scholar]
- 35. West MA, Wallin RP, Matthews SP, Svensson HG, Zaru R, et al. (2004) Enhanced dendritic cell antigen capture via toll-like receptor-induced actin remodeling. Science 305: 1153–1157. [DOI] [PubMed] [Google Scholar]