

Abstract
Path integration refers to the ability to integrate continuous information of the direction and distance travelled by the system relative to the origin. Previous studies have investigated path integration through blindfolded walking along simple paths such as straight line and triangles. However, limited knowledge exists regarding the role of path complexity in path integration. Moreover, little is known about how information from different sensory input systems (like vision and proprioception) contributes to accurate path integration. The purpose of the current study was to investigate how sensory information and curved path complexity affect path integration. Forty blindfolded participants had to accurately reproduce a curved path and return to the origin. They were divided into four groups that differed in the curved path, circle (simple) or figure-eight (complex), and received either visual (previously seen) or proprioceptive (previously guided) information about the path before they reproduced it. The dependent variables used were average trajectory error, walking speed, and distance travelled. The results indicated that (a) both groups that walked on a circular path and both groups that received visual information produced greater accuracy in reproducing the path. Moreover, the performance of the group that received proprioceptive information and later walked on a figure-eight path was less accurate than their corresponding circular group. The groups that had the visual information also walked faster compared to the group that had proprioceptive information. Results of the current study highlight the roles of different sensory inputs while performing blindfolded walking for path integration.
Keywords: human navigation, vision, proprioception, spatial performance, locomotion, gait
Introduction
Humans can utilize two distinct strategies for navigation: allocentric and egocentric navigation. Allocentric navigation is associated with the knowledge or memory of landmarks and the ability to orient with respect to a known object or vista of a scene [1]. Animals like honeybees, utilize landmark navigation to locate their hive while humans utilize distinct landmarks when driving [2]. Egocentric navigation is associated with path integration which is the ability to navigate in space using the system itself as a reference [3]. Continuous information of the distance and direction travelled from the system itself are integrated through path integration. Additionally, a homing vector from the starting point is created and updated until reaching the desired endpoint location. It has been demonstrated that desert ants rely on the ability of path integration by foraging along novel routes until they find a food source [4,5]. After reaching the site, desert ants calculate the homing vector to guide them back to the nest. If the desert ant is placed on a new starting location, it will continue to travel along the same (now incorrect) homing vector, demonstrating that distance and direction are updated by egocentric movement cues [6,7]. Similar behavior has been found in birds [8] and mammals [9].
Humans can use different sensory systems for path integration. These sensory systems include visual (optic flow), proprioceptive (feedback from the muscles and the tendons) and vestibular (translational and rotational accelerations) systems. However, the nature of this multi-sensory integration for path integration is unknown. The most commonly used method of investigating path integration is walking blindfolded to a previously seen target (for a review see [10]). In the past, path integration has been studied by estimating the distance and direction travelled from a starting point while walking blindfolded mostly on either a straight [11–13] or a triangular path [14,15]. The accuracy of path integration in these processes is addressed on the basis of the endpoint of the path. The differences observed between the distance of the actual path and the distance travelled of participants’ return path gives a measure of perceived distance, and the angular difference between the direction of that path and the required direction provides a measure of perceived heading.
While path integration-based research has focused on straight line and triangular paths, limited information exists regarding path integration using a circular path. Takei and colleagues found that a circular path was more demanding and required additional attentional control involving multi-sensory inputs [16–17]. The authors suggested that different sensory processes were utilized for the estimation of the length and the curvature (direction) of the path. In theory, otolith stimulation due to rotational forces (i.e. centrifugal) and/or angular position of the lower extremities can provide information about the constant change in the curvature of these paths. Proprioceptive information directly from the feet and/or information from the semicircular canals based on the head orientation could be used to update instantaneous position. However, research in the area of path complexity and how this interacts with sensory information is still scanty. It has been proposed that the proprioceptive system can be used not only to adopt a specific locomotor path but to estimate how far someone rotates during turning [18].
The purpose of the current study was to investigate how sensory information and path complexity affect path integration. Four groups of blindfolded subjects walked on a circular or a figure-eight path which they previously saw or on which they were previously guided. We hypothesized that visual information of the path (previously seen path) would lead to greater accuracy (path length and trajectory) than proprioceptive information (previously guided path). We also hypothesized that in comparison to the more complex figure-eight path, accuracy would be greater on the circular path. Finally, we hypothesized that as complexity of the path increased the difference in accuracy between the groups with visual and proprioceptive information will decrease.
Methods
Forty healthy university students from psychology and physical education majors, aged between 19 to 32 years gave informed consent according to the University guidelines (Table 1). The sample size was determined based on our pilot data. We calculated that a sample size of 10 subjects per groups in each of the four groups was sufficient to achieve an 80% power to test the effect of both sensory system and complexity of curved path. Exclusion criteria were neuromuscular or musculoskeletal disorders that could alter gait or present a safety issue, vestibular or ataxic disorders, history of dizziness or medications that can cause dizziness, synesthesia or other disorders affecting the subject’s orientation in space. Dizziness was assessed with the Dizziness Handicap Inventory (DHI) questionnaire [19].
Table 1.
Subject demographics for all groups/conditions; each group had ten subjects.
| Age (yrs) | Height (cm) | Mass (kg) | Sex (F/M) | DHI | |
|---|---|---|---|---|---|
| Circular Path - Previously Seen | 22.4 ± 3.03 | 175.2 ± 11.3 | 68.3 ± 14.22 | 5/5 | 8/100 |
| Circular Path - Guided | 21.8 ± 3.08 | 168.9 ± 6.68 | 61.4 ± 6.90 | 6/4 | 10/100 |
| Figure of eight Path - Previously Seen | 23.5 ± 4.19 | 169.8 ± 5.78 | 59.5 ± 7.13 | 7/3 | 8/100 |
| Figure of eight Path - Guided | 22.2 ± 4.02 | 172 ± 7.70 | 62.1 ± 10.24 | 7/3 | 12/100 |
Note: DHI = Dizziness Handicap Inventory
The subjects were randomly assigned into four groups/conditions. In the first group (previously seen-circular path condition), the subjects first saw the circular path and then were asked to walk blindfolded on the path while data was collected. In the second group (previously guided-circular path condition), the subjects were blindfolded upon entering into the laboratory and were hand-guided along the circular path. Then, they were asked to walk blindfolded on the path while data was collected. In the third group (previously seen-figure-eight condition), the subjects first saw the figure-eight path and then they were asked to walk blindfolded on the path while data was collected. In the fourth group (previously guided-figure-eight condition), the subjects were blindfolded upon entering into the laboratory and were hand-guided through the figure-eight path. Then they were asked to walk blindfolded on the path while data was collected. Each subject performed only one trial of the respective condition and walked with their shoes. The circular path had a radius of 1.2m. The figure-eight path had a radius of 1.2m for each semicircular component and a distance of 1.2m from the center of the figure to the center of each semicircle [17]. The experiments were conducted in a quiet environment. All the subjects were instructed to retrace the path at their self-selected speed. They were also assured that in lieu of their safety, the experimenter would inform them well in advance if they get close to any of the cameras or the wall while walking blindfolded. The nearest camera tripod was 3.1m, the nearest wall in the room was 2.87m and the nearest object (data collection station) was 1.57m from the perimeter of the circular path. The nearest camera tripod was 1.7m, the nearest wall in the room was 2.72m and the nearest object (data collection station) was 1.57m from the perimeter of the figure-eight path. In addition, all the subjects wore earplugs to avoid auditory interference.
An eight-camera system (Motion Analysis Corp, Santa Rosa, CA) was used to capture the 3D coordinates of a reflective marker placed on the sacrum of the subjects while walking. The data was exported and processed using custom-made Matlab (Mathworks Inc., Natick, MA) routine. This software was used to calculate the dependent measures of average trajectory error, walking speed, and distance travelled from the acquired coordinates for each subject during each condition. The ideal trajectory of the paths was inscribed on the laboratory floor (Figure 1).
Figure 1.
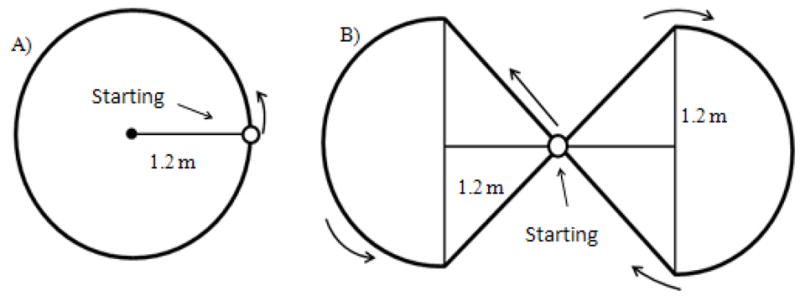
The circular (A) and figure of eight (B) paths used. The starting point is represented by the white (inside) circle and the movement direction by the direction of the arrows.
The average trajectory error was calculated as the summation of the deviation error of each point of the walked trajectory from each point of the true predefined trajectory of the path divided by the length of data points of the corresponding trial. The distance travelled was calculated as the overestimation or underestimation of the walked trajectory with the true total distance (7.53m for the circular path and 14.32m for the figure-eight path) of the predefined path. Smaller values of trajectory error and distance travelled indicate greater accuracy. Walking speed was calculated as the first derivative of the position data.
A 2×2 ANOVA was used to identify differences between the group means for the dependent variables of average trajectory error and walking speed. The two factors were complexity of the curved path (circular versus figure-eight) and sensory system (visual versus proprioception; previously seen versus previously guided). Post hoc Tukey tests were performed when a significant interaction was identified. For the dependent measure distance travelled, and due to the actual difference between the two paths (7.53m for circular and 14.32m for figure-eight), we performed separate independent t-tests for each path to compare the groups under previously seen and previously guided conditions. Statistical analysis was performed using SPSS (International Business Machines, Armonk, NY) and the level of significance was set at 0.05.
Results
Average trajectory error
The ANOVA results revealed a significant main effect for the complexity of the curved path factor [F (1, 36) = 69.75, p < 0.0001]. Both groups of the previously seen and previously guided conditions of the circular path produced much smaller values than the corresponding groups of the figure-eight path (Table 2). There was a significant main effect for the sensory system factor [F (1, 36) = 14.27, p < 0.001; Table 2]. On an average, subjects produced smaller errors while retracing the path relying on their visual system (previously seen condition) compared to subjects’ performance relying on the proprioceptive information (previously guided condition). In addition, these differences resulted in a significant interaction between the two factors [F (1, 36) = 26.47, p < 0.0001] (Table 2). Practically, while the trajectory errors for the circular path were relatively similar using both sensory systems, the error for the figure-eight path was greater while using the proprioceptive system (previously guided condition; Figure 2).
Table 2.
Group means (standard deviation) for circular and figure of eight path for all conditions (p < .05) for direction travelled, distance travelled, and walking speed.
| Trajectory error (m) | Distance Travelled Error (m) | Walking Speed (m/s) | |
|---|---|---|---|
| Circular Path - Previously Seen | 0.45 ± 0.26*,¥,‡ | 10.70 ± 1.9† | 0.31 ± 0.09¥ |
| Circular Path - Guided | 0.32 ± 0.19* | 9.95 ± 2.17† | 0.26 ± 0.07 |
| Figure of Eight Path - Previously Seen | 0.75 ± 0.22¥ | 13.83 ± 2.03 NS | 0.33 ± 0.10¥ |
| Figure of Eight Path - Guided | 1.59 ± 0.61 | 15.43 ± 2.89 NS | 0.26 ± 0.08 |
main effect of the curved path factor,
main effect of the sensory system factor,
significant differences with actual distance,
significant interactions, NS no significant differences.
Figure 2.

Sacral trajectory (thick line) of a subject walking a circular (A) and figure of eight (B) path in the previously seen condition. The thin line indicates the ideal trajectory.
Walking speed
The ANOVA results revealed significant differences for the walking speed (Table 2). There was a significant main effect for the sensory system factor [F (1, 36) = 5.67, p <.05] but not for the complexity of the curved path factor [F (1, 36) = 0.062, p = 0.805]. Subjects relying on the visual system (previously seen condition) walked faster than those relying on the proprioceptive system (previously guided condition). There was no significant interaction [F (1, 36) = 0.179, p = 0.675].
Distance travelled
The distance travelled was significantly larger than the true total distance of 7.53m for the circular previously seen condition [t (9) = 5.26, p < 0.001; Table 2]. In addition, the distance travelled was significantly larger than the true total distance of 7.53m, for the circular previously guided condition [t (9) = 3.53, p < 0.01]. No significant differences were found with the true distance of 14.32m for the figure-eight path in both the previously seen and the previously guided conditions (Table 2). Lastly, no significant differences were found for the distance travelled between the previously seen and the previously guided conditions for both the circular (10.70m versus. 9.95m) and figure-eight paths (13.83 versus 15.43m; Table 2).
Discussion
The purpose of the current study was to investigate how sensory information (visual versus proprioceptive) and path complexity (circular versus figure-eight) affect path integration. We hypothesized that visual information of the path would lead to greater accuracy than proprioceptive information. We also hypothesized that the simpler circular path would have greater accuracy than the more complex figure-eight path. Our results indicated that our hypotheses for main effects were true for the trajectory error. Further the hypothesis for the sensory system (visual vs proprioceptive) was true for the walking speed. For the interaction, we hypothesized that the differences between vision and proprioception would minimize as the path complexity increases. This hypothesis was false for the average trajectory error.
Our results showed that proprioceptive information is not as crucial as visual information for path integration when we walk a complex curved path like figure-eight. In a previous study, researchers found that healthy human subjects can accurately reproduce three circular paths of different radii even when they walk in dim lighting conditions concluding that reproduction of circular paths is possible without visual information [17]. However, their subjects had both visual and proprioceptive cues available questioning their conclusions. Our results demonstrated that during walking on curved paths, path integration relies heavily on visual information. In contrast with our results, other studies have found that information from proprioception can provide an accurate representation of the imposed distance [12,20,21]. In these studies, subjects actively or passively reproduced straight line paths that they were previously guided through. The simplicity of this type of the path could lead to the predominance of proprioceptive information for path integration. Therefore, our results suggest that proprioception can provide only a gross orientation in space when we are walking along curved paths. This explanation agrees with others who suggested a similar role for proprioception for path integration [10,22–24].
Our results demonstrated that vision is more dominant than proprioception in a figure-eight path. Specifically, when we compared the figure-eight previously guided condition with the circular previously guided condition, a significant increase was found in the trajectory error for the figure-eight path. This suggests that the complexity of the path can significantly affect the contribution of proprioception to path integration. Support for these results is provided by studies that explored path integration in more complex paths, such as in longer straight lines and triangles [12,25]. These studies suggested that returning to the starting location required some type of an additional record of the outbound path (i.e. hypotenuse) which requires the usage of additional neural resources from higher control centers [12,25].
The distance travelled demonstrated significant differences when we compared the actual trajectories with the one that the subjects reproduced. Originally, significant differences were anticipated for both paths and conditions. However, significant differences were found only for the circular path as the subjects walked more than the actual distance. Participants traversed 0.5m lesser (in previously seen condition) and 1.1m more (in previously guided condition) compared to the true total distance (14.32m) on the figure-eight path. Though the differences between the distances travelled in these conditions with the total true distance were not statistically significant, these numbers show that the subjects were always away from the true total distance during figure-eight path conditions as well. There was a tendency to undershoot the distance travelled in the previously seen condition and overshoot the distance travelled in the guided condition.
Walking speed decreased for both paths in the previously guided condition and was similar across conditions. Bredin et al. found that healthy subjects walking on a straight line tended to decrease the distance travelled with an increase in walking speed suggesting that the total distance travelled and speed are related [11]. However, our results demonstrated that differences in walking speed were not the same as that in distance travelled. Distance travelled did not change between the conditions, while walking speed changed across paths for the previously seen condition. This difference in walking speed may be an effect of fear of bumping into the surrounding cameras or even the wall, as the subjects did not have access to a guide. A similar effect was observed in another study where blindfolded subjects had to complete a triangular path while driving a mobile robot [26].
The differences seen for the average trajectory error and distance travelled variables between the two curved path conditions (circular and figure-eight) could be attributed to the difference in complexity of both the paths. The figure-eight is more complex than the circular path and this complexity could be in terms of the length of information that needs to be processed or familiarity with the shape/path. Certainly both the paths differ in the amount of information that needs to be processed as the circular trajectory has only the angular component but the figure-eight path has both the angular and linear components.
In our experiments, subjects had to retrieve information from their different sensory systems to complete the circular or the figure-eight path. This could have occurred through two distinct mechanisms. The first mechanism was associated with a visual representation of the intended path trajectory. The second mechanism was associated with proprioceptive-motor representation during which the subjects were blindfolded from the beginning and were guided through the paths. Based on the goal/specifications of the task a motor command was sent to the lower limbs to fulfill the requirements of the task. The motor command based on the motor output was reinforced from sensory information gathered from the environment and the organism itself. Then, the modified command with the reinforced information was transferred to the memory processes. However, there is the possibility that the modified command could also transfer to the path integration process and provide a new motor output that again could get reinforced from sensory feedback and get consolidated into memory. Therefore, we hypothesize that the processing centers of visual and proprioceptive information share a common biological substrate that affect the planning and execution of a motor command.
The results of this study can benefit specific populations. For example, visual- and hearing-impaired individuals as well as the elderly with vision problems can benefit through the designing of instruments that will enhance the sensory systems contribution to path integration and the formulation of cognitive maps. Another group that can benefit is miners. According to the Mine Safety and Health Administration, the annual average rate of fatalities associated with underground incidents in 1997 in mining was 56.9% (Mine Safety and Health Administration). Miners work in risky and hazardous environments. In emergency situations like a mine collapse, miners may be trapped in the dark trying to find their way to safer areas. Understanding the contribution of sensory systems in path integration can help in the development of equipment that will help miners utilize their senses other than vision in order to find their way in the dark and survive until help arrives. Our future studies will explore the contribution of other sensory systems like tactile and vestibular systems on path integration.
Conclusion
The current study investigated the contribution of vision and proprioception on path integration during simple and complex curved locomotion. The results indicate that (a) visual information minimizes error in path integration however complexity of path affects this ability and (b) the more complex the curved path, the larger the error in path integration and consequently more is the dependence on external feedback.
Figure 3.

Means (SE) of the average trajectory error showing significant interaction between factors of path complexity and memory systems.
Highlights.
The purpose of this study was to investigate how sensory information and curved path complexity affect path integration.
Participants walking on a circular path and with visual information were more accurate in reproducing the path.
Participants walking on a figure-eight path with proprioceptive information were less accurate.
These results highlight the roles of different sensory inputs while performing blindfolded walking for path integration.
Acknowledgments
Support for this work provided from an Alexander S. Onassis Public Benefit Foundation scholarship given to PK, the Nebraska Research Initiative, and the NIH (Grant No. 1R01AG034995-01A1).
Footnotes
Conflict of interest statement
None.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Foo P, Warren WH, Duchon A, Tarr MJ. Do humans integrate routes into a cognitive map? Map- versus landmark-based navigation of novel shortcuts. J Exp Psychol Learn Mem Cogn. 2005;31:195–215. doi: 10.1037/0278-7393.31.2.195. [DOI] [PubMed] [Google Scholar]
- 2.Srinivasan MV, Zhang SW. Visual control of honeybee flight. EXS. 1997;84:95–113. doi: 10.1007/978-3-0348-8878-3_4. [DOI] [PubMed] [Google Scholar]
- 3.Mittelstaedt H, Mittelstaedt ML. Avian Navigation. 1982. Homing by path integration; pp. 290–297. [Google Scholar]
- 4.Muller M, Wehner R. Path integration in desert ants, Cataglyphis fortis. Proc Natl Acad Sci U S A. 1988;85:5287–5290. doi: 10.1073/pnas.85.14.5287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Muller M, Wehner R. Wind and sky as compass cues in desert ant navigation. Naturwissenschaften. 2007;94:589–594. doi: 10.1007/s00114-007-0232-4. [DOI] [PubMed] [Google Scholar]
- 6.Sommer S, Wehner R. Vector navigation in desert ants, Cataglyphis fortis: celestial compass cues are essential for the proper use of distance information. Naturwissenschaften. 2005;92:468–471. doi: 10.1007/s00114-005-0020-y. [DOI] [PubMed] [Google Scholar]
- 7.Wehner R. Handbook of Sensory Physiology. New York: Springer; 1981. Spatial vision in arthropods; pp. 287–616. [Google Scholar]
- 8.Wallraff H. Seven theses on pigeon homing deduced from empirical findings. J Exp Biol. 1996;199:105–111. doi: 10.1242/jeb.199.1.105. [DOI] [PubMed] [Google Scholar]
- 9.Etienne AS, Maurer R, Boulens V, Levy A, Rowe T. Resetting the path integrator: a basic condition for route-based navigation. J Exp Biol. 2004;207:1491–1508. doi: 10.1242/jeb.00906. [DOI] [PubMed] [Google Scholar]
- 10.Loomis JM, Klatzky RL, Golledge RG. Navigating without vision: basic and applied research. Optom Vis Sci. 2001;78:282–289. doi: 10.1097/00006324-200105000-00011. [DOI] [PubMed] [Google Scholar]
- 11.Bredin J, Kerlirzin Y, Israel I. Path integration: is there a difference between athletes and non-athletes? Exp Brain Res. 2005;167:670–674. doi: 10.1007/s00221-005-0251-3. [DOI] [PubMed] [Google Scholar]
- 12.Durgin FH, Akagi M, Gallistel CR, Haiken W. The precision of locomotor odometry in humans. Exp Brain Res. 2009;193:429–436. doi: 10.1007/s00221-008-1640-1. [DOI] [PubMed] [Google Scholar]
- 13.Ellard CG, Shaughnessy SC. A comparison of visual and nonvisual sensory inputs to walked distance in a blind-walking task. Perception. 2003;32:567–578. doi: 10.1068/p5041. [DOI] [PubMed] [Google Scholar]
- 14.Kearns MJ, Warren WH, Duchon AP, Tarr MJ. Path integration from optic flow and body senses in a homing task. Perception. 2002;31:349–374. doi: 10.1068/p3311. [DOI] [PubMed] [Google Scholar]
- 15.Peruch P, Borel L, Magnan J, Lacour M. Direction and distance deficits in path integration after unilateral vestibular loss depend on task complexity. Brain Res Cogn Brain Res. 2005;25:862–872. doi: 10.1016/j.cogbrainres.2005.09.012. [DOI] [PubMed] [Google Scholar]
- 16.Takei Y, Grasso R, Berthoz A. Quantitative analysis of human walking trajectory on a circular path in darkness. Brain Res Bull. 1996;40:491–5. doi: 10.1016/0361-9230(96)00147-5. discussion 495–6. [DOI] [PubMed] [Google Scholar]
- 17.Takei Y, Grasso R, Amorim MA, Berthoz A. Circular trajectory formation during blind locomotion: a test for path integration and motor memory. Expl Brain Res. 1997;115:361–368. doi: 10.1007/pl00005705. [DOI] [PubMed] [Google Scholar]
- 18.Becker W, Nasios G, Raab S, Jurgens R. Fusion of vestibular and podokinesthetic information during self-turning towards instructed targets. Exp Brain Res. 2002;144:458–474. doi: 10.1007/s00221-002-1053-5. [DOI] [PubMed] [Google Scholar]
- 19.Jacobson GP, Newman CW. The development of the Dizziness Handicap Inventory (DHI) Archives of Otolaryngology-Head and Neck Surgery. 1990;116:424–427. doi: 10.1001/archotol.1990.01870040046011. [DOI] [PubMed] [Google Scholar]
- 20.Grasso R, Glasauer S, Takei Y, Berthoz A. The predictive brain: anticipatory control of head direction for the steering of locomotion. Neuroreport. 1996;7:1170–1174. [PubMed] [Google Scholar]
- 21.Berthoz A, Israël I, Georges-François G, Grasso R, Tsuzuku T. Spatial Memory of Body Linear Displacement: What Is Being Stored? Science. 1995;269:95–98. doi: 10.1126/science.7604286. [DOI] [PubMed] [Google Scholar]
- 22.Israel I, Lecoq C, Capelli A, Golomer E. Vestibular memory-contingent whole-body return: brave exocentered dancers. Ann N Y Acad Sci. 2005;1039:306–313. doi: 10.1196/annals.1325.029. [DOI] [PubMed] [Google Scholar]
- 23.Marlinsky VV. Vestibular and vestibulo-proprioceptive perception of motion in the horizontal plane in blindfolded man--I. Estimations of linear displacement. Neuroscience. 1999;90:389–394. doi: 10.1016/s0306-4522(98)00450-3. [DOI] [PubMed] [Google Scholar]
- 24.Mittelstaedt ML, Mittelstaedt H. Idiothetic navigation in humans: estimation of path length. Exp Brain Res. 2001;139:318–332. doi: 10.1007/s002210100735. [DOI] [PubMed] [Google Scholar]
- 25.Fujita N, Klatzky RL, Loomis JM, Golledge RG. The encoding-error model of pathway completion without vision. Geogr Anal. 1993:295–314. [Google Scholar]
- 26.Nico D, Israel I, Berthoz A. Interaction of visual and idiothetic information in a path completion task. Exp Brain Res. 2002;146:379–382. doi: 10.1007/s00221-002-1184-8. [DOI] [PubMed] [Google Scholar]