

Abstract
Vertebrate metamorphosis is often marked by dramatic morphological and physiological changes of the alimentary tract, along with major shifts in diet following development from larva to adult. Little is known about how these developmental changes impact the gut microbiome of the host organism. The metamorphosis of the sea lamprey (Petromyzon marinus) from a sedentary filter-feeding larva to a free-swimming sanguivorous parasite is characterized by major physiological and morphological changes to all organ systems. The transformation of the alimentary canal includes closure of the larval esophagus and the physical isolation of the pharynx from the remainder of the gut, which results in a nonfeeding period that can last up to 8 months. To determine how the gut microbiome is affected by metamorphosis, the microbial communities of feeding and nonfeeding larval and parasitic sea lamprey were surveyed using both culture-dependent and -independent methods. Our results show that the gut of the filter-feeding larva contains a greater diversity of bacteria than that of the blood-feeding parasite, with the parasite gut being dominated by Aeromonas and, to a lesser extent, Citrobacter and Shewanella. Phylogenetic analysis of the culturable Aeromonas from both the larval and parasitic gut revealed that at least five distinct species were represented. Phenotypic characterization of these isolates revealed that over half were capable of sheep red blood cell hemolysis, but all were capable of trout red blood cell hemolysis. This suggests that the enrichment of Aeromonas that accompanies metamorphosis is likely related to the sanguivorous lifestyle of the parasitic sea lamprey.
INTRODUCTION
Studies of the mammalian gut have highlighted the importance of the gut microbiome to the metabolism, behavior, physiology and general health of the host (5, 10, 12, 31). The microbiome can prevent the establishment of bacterial pathogens, aid in digestion and nutrient uptake, and even assist in the development of host tissue such as the brain, which in turn affects behavior (8, 11, 14). In vertebrates that undergo metamorphosis, however, the gut can be subject to major morphological and physiological changes, with postmetamorphic juveniles having a diet drastically different from that of their larval stage. The impact of physiological and dietary changes on the gut microbiomes of organisms that undergo metamorphosis remains largely uncharacterized.
Studies of metamorphosing anurans (frogs and toads) have evaluated changes in bacterial species composition and proportion in different life stages of the host. A survey of both tadpoles and adults of northern leopard frogs (Rana pipiens) yielded a prevalence of members of the Enterobacteriaceae, including Escherichia coli, Citrobacter, Klebsiella, Enterobacter, Serratia, and Yersinia (9). Citrobacter freundii and Aeromonas hydrophila were the predominant microbes in tadpole and adult intestines, with both being found in about 45% of sampled specimens. Another study also observed shifts in the gut microbiomes of southern toads (Bufo terrestris) and spring peepers (Pseudacris crucifer) (4). While these studies provide a glimpse into some of the possible changes in the composition of the gut microbiome during anuran metamorphosis, it is unclear how the gut microbiome is affected when metamorphosis is also accompanied by a shift to a highly specialized diet.
The sea lamprey (Petromyzon marinus) is a jawless vertebrate having a complex life cycle that involves a shift from a diverse diet of detritus and microbes to a highly specialized diet of fish blood. After hatching, newly emerged larvae burrow into the sand, where they feed on detritus and microbes in the water column for 3 to 7 years (36, 37) (Fig. 1). Once metamorphosis has begun, the larvae cannot feed due to major changes in the digestive system, which includes blockage of the larval esophagus and physical isolation from the pharynx (35). This nontrophic (nonfeeding) period can last up to 8 months (37). As metamorphosis nears completion, a new esophagus is formed and the gut develops longitudinal mucosal folds for greater absorption of nutrients (Fig. 1) (37). The digestive system also undergoes several other changes, including formation of a suctorial disc for parasitic feeding and the degeneration of the bile ducts, bile canaliculi, and gallbladder (34). To compensate for the loss of a functional blood-bile barrier, parasites use circulating bile-binding proteins to transport toxins through the circulatory system, depositing them in the posterior end of the intestine and resulting in bile accumulation in this region (34). The newly metamorphosed sea lamprey is sanguivorous, having a diet of predominately fish blood and body fluids (37).
Fig 1.
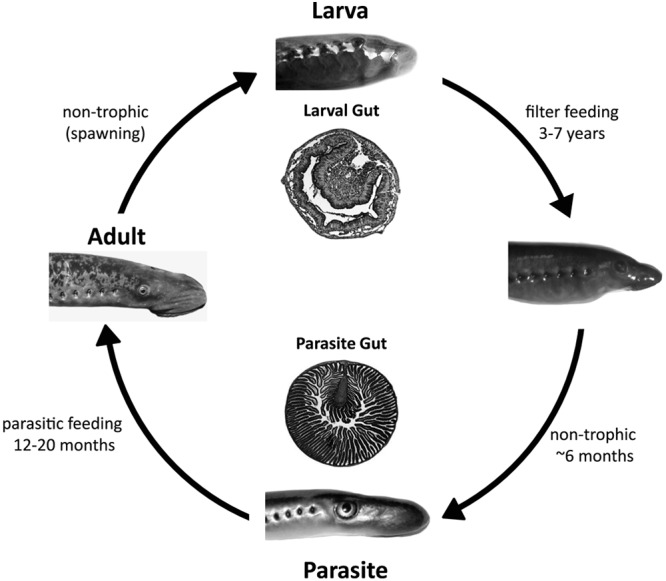
Life cycle of the sea lamprey. Adult sea lamprey spawn in the river headwaters. Embryogenesis lasts approximately 17 days, and larval lamprey hatch, float downstream, and burrow in the sandy river bed, where they remain as relatively sedentary filter feeders for 3 to 7 years. The end of the larval period is marked with a metamorphosis, which involves major morphological and physiological changes to all organ systems. Metamorphosis is a nontrophic period which lasts approximately 6 months. Postmetamorphic juveniles (parasites) migrate downstream to a large body of water (a lake or the Atlantic Ocean) and commence parasitic feeding on the blood and body fluids of boney fishes. The parasitic feeding phase lasts 12 to 20 months, after which juveniles commence upstream migration, sexually mature into adults, spawn, and die. The cross sections of the larval and parasite intestines depicted in the figure highlight the dramatic morphological changes that occur during metamorphosis.
Because the sea lamprey undergoes major morphological and physiological changes during metamorphosis and also goes through a nontrophic stage before acquiring a highly specialized diet, it represents an ideal system to evaluate the impact of changes to host physiology on the gut bacterial community. Only one published study to date has examined the lamprey gut microbiome. This particular work examined the composition of gut bacteria specifically in the larval stage of the pouched lamprey (Geotria australis), although no comparisons were made to the parasitic stage (24). In this study, we compared the gut microbiota of the larval and parasitic life stages using 16S rRNA gene sequencing and also characterized a collection of culturable isolates with respect to hemolysin production, protease secretion, and bile tolerance. Our results highlight a significant shift in the relative composition of microbes between larval and parasitic sea lamprey stages that is reflective of the shift to a more specialized diet.
MATERIALS AND METHODS
Animal and tissue collection.
Larval sea lamprey were collected from Oshawa Creek (Ontario, Canada), Harlow Creek (Michigan), and Little Garlic River (Michigan) and transported by truck to the University of Regina. When possible, sea lampreys were sampled immediately upon arrival at the University of Regina. To rear parasites, larvae were housed in aquaria with 12 to 15 cm of sterile, washed, and screened sand, filled with dechlorinated city tap water (kept at 18 to 21°C), which was aerated and filtered continuously. Larvae were fed a suspension of baker's yeast (equivalent to 1 g of yeast per animal) twice weekly until the onset of metamorphosis. Lampreys were not fed during the nontrophic phase of metamorphosis. When metamorphosis was complete, postmetamorphic juveniles were transferred to 1,700-liter aquaria containing aerated and continuously filtered dechlorinated water at 11 to 15°C. Rainbow trout (Oncorhynchus mykiss) were introduced into the tank as a food source. Prior to tissue collection, animals were anesthetized in 0.05% tricaine methanesulfonate buffered with 1% sodium bicarbonate. Following anesthesia, animals were euthanized by decapitation prior to harvesting the gut. Tissue samples were sectioned into anterior, medial, and posterior thirds and placed in 10 mM MgSO4 solution (100 μl for larvae and 400 μl for parasites). Samples from postmetamorphic sea lamprey were obtained from animals that had metamorphosed in lab aquaria. All animal handling and procedures were approved by the President's Committee on Animal Care at the University of Regina and were consistent with the guidelines of the Canadian Council on Animal Care.
DNA extraction and sequencing.
Bacteria from larval or parasitic phase sea lamprey were harvested by vortexing a pool of 10 anterior or posterior gut sections from 10 individual lampreys in 10 mM MgSO4 solution and then physically removing the tissue fragments. This process was repeated 5 times (50 animals total), producing 5 independent samples. Genomic DNA was extracted from the bacterial suspension immediately using the Qiagen Puregene Core Kit Yeast/Bacteria Genomic DNA Kit according to the manufacturer's instructions. Genomic DNA was used as a template for PCR to amplify a 1,065-bp fragment of the 16S rRNA gene using the following primers: forward 16S-335 (5′-ACTCCTACGGGAGGCAGC-3′) and reverse 16S-1400 (5′-ACGGGCGGTGTGTACAA-3′). Twenty-five-microliter PCR mixtures were prepared using 1× standard Taq buffer (10 mM Tris-HCl, 50 mM KCl, 1.5 mM MgCl2) (NEB, Mississauga, Ontario, Canada), 0.2 mM deoxynucleoside triphosphates (dNTPs), 0.2 μM forward primer, 0.2 μM reverse primer, 0.625 unit of standard Taq polymerase (NEB), and 1 μl DNA as the template. Cycling parameters were as follow: 94°C for 4 min; 32 cycles of 94°C for 30 s, 55°C for 30 s, and 72°C for 60 s; and a final 10-min extension at 72°C. PCR amplicons were visualized by gel electrophoresis using 1% agarose gels prepared in sodium borate (SB) buffer (5 mM NaOH, 20 mM boric acid, pH 8.5). PCRs were performed on bacterial genomic DNA extracts from intestinal fragments of different life history stages: larval anterior, larval posterior, parasite anterior, and parasite posterior. Five independent PCRs were performed for each, and pools were generated by combining 17 μl from each of the five independent PCRs, each of which was generated from a bacterial DNA sample derived from a pool of 10 different tissue samples. The EZ-10 spin column PCR purification kit (BioBasic, Markham, Ontario, Canada) and the accompanying protocol were used to purify the PCR product pools. Samples were then quantified on a NanoDrop spectrophotometer. Prior to cloning, PCR product pools were dA tailed by combining 1× NEB standard buffer, 0.2 mM dATP, 0.5 unit NEB standard Taq polymerase, and 3.9 μl pooled PCR product, incubating at 72°C for 15 min, and then snap-cooling on ice. The parasite samples were ligated into pGEM-T Vector using the protocol from the pGEM-T and pGEM-T Easy Vector Systems technical manual (Promega, Madison, WI). The vector was then transformed into chemically competent TOP10 cells (Invitrogen, Burlington, Ontario, Canada) according to the manufacturer's instructions. Transformed cells were plated onto lysogeny broth (LB) plates containing 150 μg/ml ampicillin and top coated with 50 μl of 2% X-Gal (5-bromo-4-chloro-3-indolyl-β-d-galactopyranoside). The plates were incubated at 37°C for 12 to 16 h. Larval samples were processed as for parasite samples with the following changes. Pooled PCR products were ligated into pCR2.1-TOPO vector (Invitrogen) according to manufacturer's instructions and then plated onto LB plates containing 50 μg/ml kanamycin. Individual colonies were selected and grown for 16 h at 37°C in 150 μl of LB containing 10% glycerol and either 150 μg/ml ampicillin or 50 μg/ml kanamycin (for parasite and larval samples, respectively). One hundred microliters from each liquid culture was transferred to a 384-well plate, frozen at −20°C overnight, packaged on dry ice, and shipped to the Plant Biotechnology Institute in Saskatoon, Saskatchewan, Canada, for DNA sequencing. Given preliminary evidence that suggested several predominant microbes in the parasite gut, we sampled more sequences from the parasite to increase the chance of sampling the rare microbes. In total, 451 random 16S rRNA clones were sequenced and analyzed for the parasite anterior and posterior gut samples and 239 for the larval anterior and posterior samples.
Isolation and phenotypic characterization of culturable isolates.
Culturable isolates were obtained by plating washes of gut tissue samples onto LB agar. A total of 62 bacterial isolates were selected from larval as well as blood-fed and unfed postmetamorphic parasites. Random isolates were selected and identified to genus level by 16S rRNA gene sequencing using the primer set described above. Hemolysin assays were carried out with 48-h incubations at 30°C on sheep blood agar (TSA II Trypticase soy agar with 10% citrated sheep blood) (BD, Mississauga, Ontario, Canada) and trout blood agar (TSA with 10% citrated trout blood) (Fort Qu'appelle Provincial Trout Hatchery, Fort Qu'appelle, Saskatchewan, Canada). Beta-hemolysin activity was expressed as the diameter of lysis (clearing) relative to colony diameter. Protease assays were conducted using skim milk agar (nutrient agar with 10% [wt/vol] skim milk powder). Casein digestion (protease secretion) was scored as positive if clearing was observed following 24 h of incubation. Bile tolerance was evaluated on MacConkey agar (50 g/liter Mikrobiolie MacConkey agar) with various concentrations of bile salts (1×, 2×, 4×, 6×, and 10×) (Sigma-Aldrich [B8756], St. Louis, MO), with incubation at 30°C for 12 to 16 h. Isolates were scored as bile tolerant if any growth was apparent.
DNA sequencing of the gyrB genes from culturable Aeromonas isolates.
Genomic DNA was extracted from liquid LB cultures grown at 37°C for 15 to 20 h using the Qiagen Puregene Core Kit Yeast/Bacteria Genomic DNA Kit according to the manufacturer's instructions. Genomic DNA was used as the template to amplify a portion of the gyrB gene using the following primers: forward 334 (5′-TCCGGCGGTCTGCACGGCGT-3′) and reverse 1464 (5′-TTGTCCGGGTTGTACTCGTC-3′) (32). Twenty-five-microliter PCR mixtures were prepared using 5× KAPA2G buffer A (1.5 mM MgCl2) (Kapabiosystems, Woburn, MA), 0.2 mM dNTPs, 0.2 μM forward primer, 0.2 μM reverse primer, 0.5 unit of KAPA2G Robust DNA polymerase (Kapabiosystems), and 1 μl DNA template. Cycling parameters were as follows: 94°C for 4 min; 35 cycles of 94°C for 30 s, 60°C for 30 s, and 72°C for 60 s; and a final 10-min extension at 72°C. PCR amplicons were analyzed using 1% SB agarose gel electrophoresis. Following PCR amplification, samples were processed for automated DNA sequencing by adding 1 unit each calf intestinal phosphatase (NEB) and exonuclease I (NEB) to each sample and incubating at 37°C for 15 min followed by 15 min at 80°C. Eight microliters of each sample was placed in a 96-well plate and combined with 3.3 μM forward primer (forward 334). Sequencing was performed by Eurofins MWG Operon.
Sequence analysis and phylogenetic reconstruction.
Sequences were evaluated for both vector contamination and quality. DNA sequences were trimmed using the Sequencher 4.8 (GeneCodes Corporation) program with the following parameters: for the 5′ end, trimming no more than 25%, trim until the first 25 bases contain fewer than 3 bases with confidence below 25; for the 3′ end, trim from the 3′ end until the last 25 bases contain fewer than 3 bases with confidence below 25; and for postfix, remove leading and trailing ambiguous bases (but some sequences had additional sequence kept with lower confidence scores). The 16S rRNA gene sequences were compared to the Ribosomal Database Project using Classifier, 16S rRNA training set 9, with a cutoff value of 50% (3), while gyrB sequences were analyzed using a BLASTN search of GenBank (1). For phylogenetic reconstruction, the 16S rRNA gene and gyrB sequences were aligned separately using ClustalX, version 2.0 (15), along with known sequences gathered from the Ribosomal Database and GenBank, respectively (2, 3). From these alignments, neighbor-joining phylogenetic trees were constructed with MEGA5 (28).
Nucleotide sequence accession numbers.
The sequences of the 16S rRNA genes and gyrB genes have been deposited under GenBank accession numbers JX453764 to JX454446 and JX453730 to JX453763, respectively.
RESULTS
Comparison of microbial communities in larvae and parasites.
A comparison of 16S rRNA gene sequences from both the larval and parasitic sea lamprey revealed a higher bacterial diversity in larvae, with members of the Proteobacteria (36%), Bacteroidetes (30%), and Tenericutes (26%) comprising the majority of diversity sampled. The predominant genera included Ureaplasma (15%), Paludibacter (11%), Acinetobacter (8%), and Legionella (5%) (Fig. 2). Taxa that were less abundant included Coxiella (4%), Aeromonas (4%), Mycoplasma (3%), Exiguobacterium (2%), and Helicobacter (2%), along with approximately 20 other genera and unclassified taxa (see Table S1 in the supplemental material). An analysis of the distribution of these bacteria along the length of the larval gut revealed some evidence of microbial partitioning, where the larval anterior gut contained predominantly Acinetobacter and members of the Bacteriodetes. These groups were underrepresented in the posterior gut, which was dominated by Ureaplasma and Paludibacter.
Fig 2.

Distribution of bacterial taxa found in the gut of the lamprey. (A) Predominant bacterial groups in the larval stage. “Tenericutes” comprises Ureaplasma, Mycoplasma, and unclassified Tenericutes; “Other Bacteroidetes” comprises three genera along with related, unclassified taxa; “Other Proteobacteria” comprises 10 genera along with unclassified taxa; and “Other” comprises seven genera, along with unclassified taxa. (B) Prevalent bacterial genera in the parasitic stage. “Other” comprises six taxa, along with unclassified taxa.
In contrast, the parasite gut was dominated primarily by Aeromonas (84%), with a smaller proportion of Shewanella (11%) and Citrobacter (2%) species (Fig. 2). Other taxa identified in the parasite included Curvibacter and Raoultella, as well as several others, but these represented only about 3% of the sampled diversity (see Table S1 in the supplemental material). The parasite anterior and posterior bacterial populations were quite similar, with most taxa being found throughout; however, Citrobacter isolates were more abundant in the posterior than in the anterior gut.
Isolation and phenotypic characterization of culturable isolates.
Culturable bacteria were isolated from gut tissue samples of the three different stream populations of larval and lab-reared parasitic (fed and unfed) sea lampreys, and each isolate was identified by 16S rRNA gene typing. Isolates of Aeromonas, Citrobacter, and Enterococcus were cultured from both wild-caught larval and lab-reared parasite gut tissues. The parasite gut tissues also yielded isolates of Shewanella, Deinococcus, Micrococcus, Nitrosomonas, and Rhodococcus. With the exception of Aeromonas, Citrobacter, and Shewanella, the remaining isolates were not represented in the 16S rRNA metagenomic clone library. Cultured isolates were characterized phenotypically for several environment-specific biochemical capabilities. First, the ability to grow on bile, which is found at high concentrations in the parasite posterior gut (33), was tested using MacConkey agar containing a range of bile salt concentrations. All Aeromonas and Citrobacter isolates, which were found in anterior and posterior sections of the gut in both larvae and parasites, were able to grow on all bile salt concentrations tested, whereas Enterococcus isolates, which were also distributed throughout the gut in both lamprey life stages, were not bile tolerant (Table 1). Shewanella strains were largely bile tolerant and capable of growing on bile salt concentrations of up to 6× (Table 1).
Table 1.
Phenotypic characteristics of culturable isolates from lamprey
| Stage | Isolate no. | Feedinga | Gut regionb | Taxon | Bile tolerance on MacConkey agarc |
Hemolysis ond: |
Proteolysis | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sheep blood |
Trout blood |
|||||||||||||
| 1× | 2× | 4× | 6× | 10× | α, β, γ | Ratio | α, β, γ | Ratio | ||||||
| Larval | 11 | F | P | Aeromonas | + | + | + | + | + | γ | β | 1.71 | + | |
| 21 | F | M | Aeromonas | + | + | + | + | + | β | 1.44 | β | 1.63 | + | |
| 25 | F | A | Aeromonas | + | + | + | + | + | β | 1.45 | β | 2.08 | + | |
| 27 | F | C | A. allosaccharophilae | + | + | + | + | + | β | 1.20 | β | 1.55 | + | |
| 6c | F | C | A. bestiarum | + | + | + | + | + | β | 1.13 | β | 2.33 | + | |
| 17c | F | C | A. bestiarum | + | + | + | + | + | β | 1.28 | β | 2.31 | + | |
| 3c | F | C | A. bestiarum | + | + | + | + | + | γ | β | 2.33 | + | ||
| 16c | F | C | A. bestiarum | + | + | + | + | + | γ | β | 2.43 | + | ||
| 7c | F | C | A. media | + | + | + | + | + | γ | β | 2.00 | + | ||
| 8c | F | C | A. media | + | + | + | + | + | γ | β | 1.87 | + | ||
| 14 | F | P | A. media | + | + | + | + | + | γ | β | 1.89 | + | ||
| 18 | F | M | A. media | + | + | + | + | + | γ | β | 1.94 | + | ||
| 20 | F | M | A. media | + | + | + | + | + | γ | β | 2.13 | + | ||
| 22 | F | M | A. media | + | + | + | + | + | γ | β | 2.19 | + | ||
| 23 | F | A | A. media | + | + | + | + | + | γ | β | 2.92 | + | ||
| 29 | F | C | A. mediae | + | + | + | + | + | γ | β | 2.07 | + | ||
| 24 | F | A | A. salmonicida | + | + | + | + | + | β | 2.75 | β | 2.33 | + | |
| 16 | F | M | A. veronii | + | + | + | + | + | β | 1.50 | β | 1.60 | + | |
| 15 | F | P | Citrobacter | + | + | + | + | + | β | 1.45 | β | 2.31 | + | |
| 19 | F | M | Citrobacter | + | + | + | + | + | γ | α | − | |||
| 28 | F | C | Citrobactere | + | + | + | + | + | γ | α | − | |||
| 30 | F | C | Citrobactere | + | + | + | + | + | γ | α | − | |||
| 12 | F | P | Enterococcus | − | − | − | − | − | α | α | − | |||
| 13 | F | P | Enterococcus | − | − | − | − | − | NDf | α | − | |||
| 10 | F | P | Enterococcus | − | − | − | − | − | α | γ | − | |||
| 17 | F | M | Enterococcus | − | − | − | − | − | α | γ | − | |||
| 26 | F | A | Enterococcus | − | − | − | − | − | α | γ | − | |||
| Parasite | 101 | F | C | Aeromonas | + | + | + | + | + | β | 2.23 | β | 2.18 | + |
| 1 | U | P | A. media | + | + | + | + | + | β | 2.67 | β | 2.07 | + | |
| 6 | U | M | A. media | + | + | + | + | + | β | 2.82 | β | 2.19 | + | |
| 7 | U | M | A. media | + | + | + | + | + | β | 2.23 | β | 2.29 | + | |
| 8 | U | M | A. media | + | + | + | + | + | β | 2.64 | β | 2.13 | + | |
| 32 | U | A | A. media | + | + | + | + | + | β | 2.82 | β | 2.80 | + | |
| 33 | U | A | A. media | + | + | + | + | + | β | 2.64 | β | 2.73 | + | |
| 35 | U | A | A. media | + | + | + | + | + | β | 2.55 | β | 2.21 | + | |
| 38 | F | C | A. media | + | + | + | + | + | β | 2.64 | β | 2.50 | + | |
| 41 | F | C | A. media | + | + | + | + | + | β | 1.07 | β | 2.50 | + | |
| 36 | U | A | A. media | + | + | + | + | + | β | 2.64 | β | 2.54 | + | |
| 40 | F | C | A. media | + | + | + | + | + | γ | β | 2.92 | + | ||
| 74 | F | C | A. media | + | + | + | + | + | γ | β | 2.13 | − | ||
| 91 | F | C | A. media | + | + | + | + | + | γ | β | 2.13 | − | ||
| 95 | F | C | A. salmonicida | + | + | + | + | + | β | 2.33 | β | 2.83 | + | |
| 96 | F | C | A. salmonicida | + | + | + | + | + | β | 2.92 | β | 2.62 | + | |
| 102 | F | C | A. sobria | + | + | + | + | + | γ | β | 1.12 | + | ||
| 110 | F | C | A. sobria | + | + | + | + | + | γ | β | 1.11 | + | ||
| 117 | F | C | A. sobria | + | + | + | + | + | γ | β | 1.07 | + | ||
| 2 | U | C | Citrobacter | + | + | + | + | + | γ | α | − | |||
| 9 | U | M | Citrobacter | + | + | + | + | + | γ | α | + | |||
| 37 | F | C | Citrobacter | + | + | + | + | + | γ | α | − | |||
| 4 | U | P | Citrobacter | + | + | + | + | + | γ | α | − | |||
| 113 | F | C | Deinococcus | − | − | − | − | − | γ | β | 2.00 | − | ||
| 89 | F | C | Ensifer | + | + | + | + | + | γ | γ | − | |||
| 3 | U | P | Enterococcus | − | − | − | − | − | α | α | − | |||
| 92 | F | C | Micrococcus | − | − | − | − | − | γ | γ | + | |||
| 31 | U | A | Nitrosomonas | + | + | + | + | + | β | 2.82 | β | 2.73 | + | |
| 111 | F | C | Rhodococcus | + | + | + | + | + | α | β | 1.08 | − | ||
| 39 | F | C | Shewanella | − | + | + | + | − | γ | β | 1.07 | − | ||
| 93 | F | C | Shewanella | − | + | − | − | − | γ | β | 1.07 | − | ||
| 34 | U | A | Shewanella | + | + | + | + | − | α | γ | − | |||
| 42 | F | C | Shewanella | − | + | + | + | − | α | γ | − | |||
| 112 | F | C | Shewanella | − | + | + | + | − | α | γ | − | |||
| 119 | F | C | Shewanella | − | + | + | + | + | α | γ | − | |||
F, fed; U, unfed.
A, anterior gut segment; M, medial gut segment; P, posterior gut segment; C, complete gut.
MacConkey agar results seen at bile concentrations of 1×, 2×, 4×, 6×, and 10×, where + indicates growth and − indicates no growth.
Blood agar hemolysis results indicated as γ, α, or β; if beta-hemolysis was observed, the ratio of diameter clearing to diameter of colony is given.
Isolated from yeast pellet.
ND, not detected.
Isolates were evaluated for their ability to produce enzymes related to digestion. Growth on skim milk agar, an assay developed for evaluating protease secretion, showed that 63% of cultured isolates, including all but two Aeromonas isolates (isolates 74 and 91 from the parasite) could hydrolyze casein, whereas all of the Enterococcus and Shewanella isolates and most of the Citrobacter isolates could not (Table 1). Isolates were also evaluated for production of hemolysins using both sheep and trout blood. On sheep blood agar, 31% of the 26 larval isolates tested showed beta-hemolysis (39% of the Aeromonas isolates), and 54% were gamma-hemolytic/nonhemolytic (61% of the Aeromonas isolates). The parasite tended to have more strains that were beta-hemolytic on sheep blood agar (40% of all isolates) but a much greater proportion of beta-hemolytic Aeromonas isolates (68%). On the trout blood agar, approximately 70% of larval and 70% of parasite isolates showed beta-hemolysis, with 100% of Aeromonas isolates being beta-hemolytic on trout blood (Table 1). Citrobacter and Shewanella isolates recovered from the parasite did not exhibit beta-hemolytic abilities.
Phylogeny of Aeromonas.
Given the prevalence of Aeromonas species in the parasitic lamprey, the species diversity of the cultured aeromonads from both larval and parasite samples was evaluated by a phylogenetic approach using the gyrB gene (Fig. 3). The gyrB gene has a mean synonymous (conserved) substitution rate almost four times that of the 16S rRNA gene, making it more suitable for establishing species relationships (32). A. allosaccharophila, A. bestiarum, A. media, A. salmonicida, and A. veronii were identified in the larval gut, while A. media, A. salmonicida, and A. sobria were found in the parasite gut (Fig. 3). There was no apparent correlation between microbial species and either lamprey population (natal stream) or life stage. However, all Aeromonas isolates collected from the unfed parasites were A. media, with approximately 66% of the cultured isolates from both larvae and parasites being identified as A. media.
Fig 3.

Neighbor-joining tree of 34 Aeromonas isolates based on gyrB. Reference taxa include Aeromonas allosaccharophila (accession number AY101777), Aeromonas bestiarum (AY101774), Aeromonas media (AY101782), Aeromonas popoffii (AY101801), Aeromonas salmonicida (AY101773), Aeromonas sobria (AY101781), Aeromonas veronii bv. sobria (AY101775), Aeromonas veronii bv. veronii (AY101787), and Escherichia coli (HQ660623).
Aeromonas strains having similar phenotypic characteristics tend to cluster together in the phylogenetic tree. All Aeromonas isolates had beta-hemolytic activity on trout blood, whereas sheep blood hemolysis was variable (beta or gamma). The A. media group has both beta- and gamma-hemolytic groups, with the beta-hemolytic strains, which were all isolated from the parasite, being monophyletic (Fig. 3). In addition, the two isolates that do not produce casein hydrolase, 91 and 74, are also monophyletic within the A. media group. The A. bestiarum group contains both beta- and gamma-hemolytic groups, again with beta-hemolytic strains being monophyletic. Isolates from the A. salmonicida, A. allosaccharophila, and A. veronii were all beta-hemolytic, while the A. sobria isolates were all gamma-hemolytic.
DISCUSSION
The unusually complex life cycle of the sea lamprey, which includes a nontrophic metamorphic stage and a dramatic shift to a highly specialized diet, provides a unique glimpse into how changes in host physiology impact the microbial communities in the gut. A comparison of the community compositions of the larval and parasitic stages using both culture-dependent and culture-independent methods revealed a higher bacterial diversity in the filter-feeding larvae, which is consistent with previous studies that examined microbial diversity in the larval pouched lamprey (24). The predominant bacterial species of the larval pouched lamprey included Bacillus mycoides and Aeromonas hydrophila, although the genera Enterobacter, Pseudomonas, Clostridium, and Corynebacterium were also represented (24). Although Aeromonas, Enterobacter, and Clostridium were identified in the larval sea lamprey, the majority of diversity was represented by the Tenericutes (Ureaplasma and Mycoplasma), as well as members of the Bacteroidetes (Paludibacter). It is interesting, however, that Aeromonas hydrophila was not recovered from the sea lamprey. The microbial composition of the lamprey, however, has been shown to be correlated with the microbial communities of the river bed and the water in and around the larval burrow (24).
The parasitic juvenile, which is not a filter feeder and subsists predominantly on a diet of fish blood, has lower microbial diversity than the larval stage, with predominant species of Aeromonas, Citrobacter, and Shewanella. Culture-dependent methods recovered additional species that were not present in the 16S rRNA gene library. This difference may have been due to culturing bias, which favors fast-growing species, 16S rRNA gene amplification bias, which could have led to the overrepresentation of the predominant groups such as Aeromonas, Citrobacter and Shewanella, and low sampling of clones. The microbiome of parasitic phase lamprey might also be affected in part by the fact they were reared under laboratory conditions, since this environment could potentially expose the lamprey to other microbial consortia. Sampling of wild-caught feeding parasites could shed more light on the influence of lab rearing on microbial communities, but these animals are extremely difficult to obtain. Nonetheless, there was a clear difference in microbial community structure during the transition from larva to parasite, with a pronounced enrichment of Aeromonas. This community shift may be attributable to the metabolic capabilities of Aeromonas, since many species produce aerolysins that lyse red blood cells and may therefore gain ready access to nutrients in addition to aiding the parasitic host in the digestion of the blood meal and/or nutrient absorption (13, 21). Indeed, our phenotypic tests of culturable isolates revealed that just over half of the culturable Aeromonas isolates tested were capable of lysing sheep blood, while all isolates were capable of lysing trout blood. This suggests that the hemolysins produced by these Aeromonas strains are more specialized to fish blood. Not all isolates have this capability, though. The outgroup (isolate 4), a putative Citrobacter isolate, exhibits alpha-hemolytic activity on trout blood. Furthermore, the majority of culturable Aeromonas isolates from both the larval and parasite guts also show protease secretion, indicating possible involvement in the digestion of the protein-rich blood diet. Interestingly, the medicinal leech, vampire bat, and mosquito, all of which are sanguivorous, have been found to have Aeromonas in their guts (6, 16, 19, 21).
The shift in gut community composition from high bacterial diversity to one in which Aeromonas is predominant raises several interesting questions about the origin of these isolates and the specific mechanisms underlying their enrichment during metamorphosis. Aeromonads are present in the gut microbiomes of many fish (21–23), which is not surprising given the prevalence of Aeromonas in aquatic environments (26). At least five distinct Aeromonas species were found to be represented in the larval stage and at least three in the parasite gut, but the same Aeromonas species are found in both life stages. This could suggest that Aeromonas may colonize the larval stage from the general environment and persist in the gut during the nontrophic metamorphic period through to adulthood. This is supported by the fact that larvae collected from different localities carry species of Aeromonas that are found in the parasite and by the fact that there was no evidence of particular phylogenetic groups being represented more in either life stage. Alternatively, the gut microbiome could be purged completely during metamorphosis, allowing for recolonization by free-living bacteria from the general environment. When mosquitoes undergo metamorphosis from larva to adult, they utilize a gut sterilization mechanism, which purges the insect midgut of bacteria (18). Meconial peritrophic membranes that sequester bacteria are formed and surround the larval midgut epithelium, and they are eventually sloughed off during metamorphosis (18, 25). Such processes may be accompanied by other specificity mechanisms that promote the establishment of specific bacteria from the general environment. In the squid, which forms a mutualistic association with Vibrio fischeri, nitric oxide is produced in the light organ tissues that enables colonization of the symbiont through a series of signaling steps (29, 30). For the medicinal leech, the ingested blood meal contains an active complement system capable of killing sensitive bacteria (13, 20, 27). Similar systems could function in the sea lamprey to eliminate some groups of bacteria. It is also possible that the accumulation of toxic bile during metamorphosis could be functioning to impose selection on specific species or groups of bacteria. Aeromonas strains tested were all tolerant to high bile concentrations; however, cultured isolates from the parasite were no more bile tolerant than those in the larva, suggesting that this alone cannot be responsible for the enrichment observed. Shewanella and Citrobacter strains that were more prevalent in the parasite tended to be bile tolerant, with the Shewanella strains tolerant to various degrees. In fact, Citrobacter strains were found to be more prevalent in the parasite posterior gut, where bile is known to accumulate. Still, there were several isolates in the parasite that were bile intolerant, suggesting that bile accumulation may not be a primary selective mechanism. If particular host specificity determinants that allow particular strains to persist within the sea lamprey do function, the colonization by these bacteria may actively exclude other microbes from establishing, possibly through the action of specific antimicrobial compounds. The medicinal leech, for example, has been suggested to possess a single Aeromonas species, Aeromonas veronii bv. sobria (6), which prevents other bacteria from colonizing the digestive tract through the production of antimicrobial compounds (7, 13, 17). Still, it is unclear whether similar, specific mechanisms function in the sea lamprey to promote the persistence and/or colonization of particular strains during sea lamprey metamorphosis. Nonetheless, this study has established that sea lamprey experience a pronounced change in their gut microflora following the transition from filter-feeding larvae to sanguivorous parasites. Our results have formed the basis for investigating specific mechanisms that control gut microbiota development and for establishing the importance of particular species of gut bacteria, such as Aeromonas, in host blood meal digestion and nutrient absorption. Determining the impact of Aeromonas on host fitness may provide a unique target for the development of new lampricidal compounds.
Supplementary Material
ACKNOWLEDGMENTS
We thank Tyler Boa, Caristin McDougal, and Irene Chair for technical assistance.
This work was supported by Natural Sciences and Engineering Council of Canada (NSERC) Discovery Grants to R.G.M., J.S., and C.K.Y. and by a University of Regina Faculty of Science Research Grant to R.G.M. A.T. was supported in part by an NSERC Undergraduate Student Research Award.
Footnotes
Published ahead of print 24 August 2012
Supplemental material for this article may be found at http://aem.asm.org/.
REFERENCES
- 1. Altschul SF, et al. 1997. Gapped blast and psi-blast: A new generation of protein database search programs. Nucleic Acids Res. 25:3389–3402 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Benson DA, Karsch-Mizrachi I, Lipman DJ, Ostell J, Sayers EW. 2011. Genbank. Nucleic Acids Res. 39:D32–D37 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Cole JR, et al. 2009. The ribosomal database project: improved alignments and new tools for rRNA analysis. Nucleic Acids Res. 37:D141–D145 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Fedewa LA. 2006. Fluctuating gram-negative microflora in developing anurans. J. Herpetol. 40:131–135 [Google Scholar]
- 5. Gill SR, et al. 2006. Metagenomic analysis of the human distal gut microbiome. Science 312:1355–1359 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Graf J. 1999. Symbiosis of Aeromonas veronii biovar sobria and Hirudo medicinalis, the medicinal leech: a novel model for digestive tract associations. Infect. Immun. 67:1–7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Graf J, Kikuchi Y, Rio RVM. 2006. Leeches and their microbiota: naturally simple symbiosis models. Trends Microbiol. 14:365–371 [DOI] [PubMed] [Google Scholar]
- 8. Heijtza RD, et al. 2011. Normal gut microbiota modulates brain development and behavior. Proc. Natl. Acad. Sci. U. S. A. 108:3047–3052 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Hird DW, et al. 1983. Enterobacteriaceae and Aeromonas hydrophila in Minnesota frogs and tadpoles (Rana pipiens). Appl. Environ. Microbiol. 46:1423–1425 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Hooper LV, Bry L, Falk PG, Gordon JI. 1998. Host-microbial symbiosis in the mammalian intestine: exploring an internal ecosystem. Bioessays 20:336–343 [DOI] [PubMed] [Google Scholar]
- 11. Hooper LV, Gordon JI. 2001. Commensal host-bacterial relationships in the gut. Science 292:1115–1118 [DOI] [PubMed] [Google Scholar]
- 12. Hooper LV, Midtvedt T, Gordon JI. 2002. How host-microbial interactions shape the nutrient environment of the mammalian intestine. Annu. Rev. Nutr. 22:283–307 [DOI] [PubMed] [Google Scholar]
- 13. Indergand S, Graf J. 2000. Ingested blood contributes to the specificity of the symbiosis of Aeromonas veronii biovar sobria and Hirudo medicinalis, the medicinal leech. Appl. Environ. Microbiol. 66:4735–4741 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Kelly D, Conway S, Aminov R. 2005. Commensal gut bacteria: mechanisms of immune modulation. Trends Immunol. 26:326–333 [DOI] [PubMed] [Google Scholar]
- 15. Larkin MA, et al. 2007. Clustal W and Clustal X version 2.0. Bioinformatics 23:2947–2948 [DOI] [PubMed] [Google Scholar]
- 16. Mackay DR, et al. 1999. Aeromonas species isolated from medicinal leeches. Ann. Plas. Surg. 42:275–279 [DOI] [PubMed] [Google Scholar]
- 17. Messi P, Guerrieri E, Bondi M. 2003. Bacteriocin-like substance (bls) production in Aeromonas hydrophila water isolates. FEMS Microbiol. Lett. 220:121–125 [DOI] [PubMed] [Google Scholar]
- 18. Moll RM, Romoser WS, Modrzakowski MC, Moncayo AC, Lerdthusnee K. 2001. Meconial peritrophic membranes and the fate of midgut bacteria during mosquito (Diptera culicidae) metamorphosis. J. Med. Entomol. 38:29–32 [DOI] [PubMed] [Google Scholar]
- 19. Muller HE, Pinus M, Schmidt U. 1980. Aeromonas hydrophila as a normal intestinal bacterium of the vampire bat (Desmodus rotundus). Zentralbl. Veterinarmed. B 27:419–424 [PubMed] [Google Scholar]
- 20. Nelson M, Graf J. 2012. Bacterial symbioses of the medicinal leech Hirudo verbana. Gut Microbes doi:10.4161/gmic.20227 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Pidiyar V, Kaznowski A, Narayan NB, Patole M, Shouche YS. 2002. Aeromonas culicicola sp nov., from the midgut of Culex quinquefasciatus. Int. J. Syst. Evol. Microbiol. 52:1723–1728 [DOI] [PubMed] [Google Scholar]
- 22. Ringo E, Birkbeck TH. 1999. Intestinal microflora of fish larvae and fry. Aquacult. Res. 30:73–93 [Google Scholar]
- 23. Ringo E, Olsen RE. 1999. The effect of diet on aerobic bacterial flora associated with intestine of arctic charr (Salvelinus alpinus L.). J. Appl. Microbiol. 86:22–28 [DOI] [PubMed] [Google Scholar]
- 24. Rogers PA, Glenn AR, Potter IC. 1980. The bacterial flora of the gut contents and environment of larval lampreys. Acta Zool. 61:23–27 [Google Scholar]
- 25. Romoser WS, Moll RM, Moncayo AC, Lerdthusnee K. 2000. The occurrence and fate of the meconium and meconial peritrophic membranes in pupal and adult mosquitoes (Diptera culicidae). J. Med. Entomol. 37:893–896 [DOI] [PubMed] [Google Scholar]
- 26. Seshadri R, et al. 2006. Genome sequence of Aeromonas hydrophila ATCC 7966(t): jack of all trades. J. Bacteriol. 188:8272–8282 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Silver A, Graf J. 2011. Innate and procured immunity inside the digestive tract of the medicinal leech. Invertebrate Surviv. J. 8:173–178 [PMC free article] [PubMed] [Google Scholar]
- 28. Tamura K, et al. 2011. Mega5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 28:2731–2739 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Visick KL, Ruby EG. 2006. Vibrio fischeri and its host: it takes two to tango. Curr. Opin. Microbiol. 9:632–638 [DOI] [PubMed] [Google Scholar]
- 30. Wang YL, et al. 2010. Vibrio fischeri flavohaemoglobin protects against nitric oxide during initiation of the squid-vibrio symbiosis. Mol. Microbiol. 78:903–915 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Willing BP, Antunes LCM, Keeney KM, Ferreira RBR, Finlay BB. 2011. Harvesting the biological potential of the human gut microbiome. Bioessays 33:414–418 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Yanez MA, Catalan V, Apraiz D, Figueras MJ, Martinez-Murcia AJ. 2003. Phylogenetic analysis of members of the genus Aeromonas based on gyrb gene sequences. Int. J. Syst. Evol. Microbiol. 53:875–883 [DOI] [PubMed] [Google Scholar]
- 33. Youson JH. 1981. The alimentary canal, p 95–189 In Hardisty MW, Potter IC. (ed), The biology of lampreys, vol 3 Academic Press, London, United Kingdom [Google Scholar]
- 34. Youson JH. 1999. The lability and endocrinology of lamprey metamorphosis: an ancient developmental strategy, p 285–288 In Roubos EW, Wendelaar-Bonga SE, Vaudry H, Loof D. (ed), Recent developments in comparative endocrinology and neurobiology. Shaker, Maastrict, The Neatherlands [Google Scholar]
- 35. Youson JH. 2003. The biology of metamorphosis in sea lampreys: endocrine, environmental, and physiological cues and events, and their potential application to lamprey control. J. Great Lakes Res. 29(Suppl 1):26–49 [Google Scholar]
- 36. Youson JH. 2004. The impact of environmental and hormonal cues on the evolution of fish metamorphosis, p 239–278 In Hall BK, Pearson RD, Muller GB. (ed), Environment, development, and evolution toward a synthesis. Massachusetts Institute of Technology, Cambridge, MA [Google Scholar]
- 37. Youson JH, Manzon RG. 2012. Lamprey metamorphosis, p 12–75 In Dufour S, Rousseau K, Kapoor BG. (ed), Metamorphosis in fish. Science Publishers, Enfield, NH [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.