

Abstract
Pancreatic cancer remains a leading cause of death despite its relatively low incidence. As in many other solid tumors, angiogenesis is critical to the growth and metastasis of this cancer. Through serial in vivo passages in mice, we have developed a highly aggressive variant of human pancreatic cancer cell line XPA-1 which demonstrates more rapid primary tumor growth, faster time to metastasis, and more rapid lethality than the parental cell line. The aggressive variant developed a much denser tumor vasculature early during growth within the pancreas. Interestingly, examination of the in vitro growth of this aggressive variant yielded no significant difference from the parental cell line. Real-time PCR evaluation of genes involved in angiogenesis revealed a 24-fold increase in Thrombospondin-1 expression in cells derived from the aggressive variant when compared to the parental cell line. These findings provide direct evidence that elevated capability for angiogenesis, mediated by specific changes in gene expression, can lead to a large increase in cancer aggressiveness and resulting metastasis. These findings have important implications for the treatment of metastatic disease.
Keywords: RFP, Angiogenesis, Thrombospondin-1, Pancreatic Cancer
Introduction
Despite its relatively low incidence in the population, pancreatic adenocarcinoma remains the fourth leading cause of cancer-related death in the United States (1), with overall 5-year survival as low as 5% (2). Recent advances in surgical and critical care have had little impact on the epidemiology of this disease, largely due to its often late stage at presentation (3) and its aggressive biology. The precise mechanisms involved in local tumor progression and metastasis in this disease remain poorly understood. Local invasion and distant metastasis occur within complex tumor-host microenvironments and involve signal exchange between cancer cells and host stroma. The tumor microenvironment promotes proliferation and survival of cancer cells in addition to stimulating cancer cell migration and invasiveness (4). Pancreatic adenocarcinoma, in particular, is characterized by a dense stromal reaction, which has been postulated to support tumor invasion (5).
The oxygen and nutrients which are central to cell survival are supplied to tissues by functioning vasculature. New blood vessel formation (angiogenesis) is critical to continued local and systemic progression of rapidly-growing tumors. The formation of new blood vessels is initiated by signaling from cancer and stromal cells, including the release of stimulatory factors which begin a series of events ultimately leading to endothelial cell degradation of extracellular matrix, migration, and proliferation (6, 7). A number of factors involved in angiogenesis are often perturbed in pancreatic cancer, including upregulation of pro-angiogenic cytokines (8). Pancreatic cancer has been shown to metastasize via hematogenous spread (9), and clinical studies in patients with pancreatic cancer have linked increased tumor angiogenesis with poorer prognosis (10).
The realization of the critical nature of angiogenesis in tumor growth and metastasis has led to the development of pharmacologic agents that block various steps in the process of tumor neovascularization (11). Growing experience with these drugs has underscored the complex nature of tumor angiogenesis which involves a dynamic interplay between positive and negative secreted regulators in addition to local tissue modulators (6). Indeed, both pro- and antiangiogenic mediators may play a role in tumor vasculature maturation and function (12). We describe here a highly aggressive human pancreatic cancer cell line generated through repeated in vivo passage to select highly metastatic cells, which demonstrates both increased tumor vascularity and upregulation of Thrombospondin-1 (TSP-1), a molecule with whose role in angiogenesis remains incompletely understood (13, 14).
Materials and Methods
Cell Culture
Cells were maintained in RPMI 1640 media supplemented with 10% fetal bovine serum (FBS) from Gibco-BRL, Life Technologies, Inc. (Grand Island, NY.) The human pancreatic cancer cell line XPA-1 was a gift from Dr. Anirban Maitra at Johns Hopkins University. All media was supplemented with penicillin/streptomycin (Gibco-BRL), L-glutamine (Gibco-BRL), MEM nonessential amino acids (Gibco-BRL), sodium bicarbonate (Cellgro, Herndon VA), and sodium pyruvate (Gibco-BRL). All cell lines were cultured at 37°C with 5% CO2.
Production of a Stable Red Fluorescent Cell Line (XPA1-RFP)
The pDSRed-2 vector from Clontech Laboratories, Inc. (Palo Alto, CA) was used for stable expression of RFP in the human pancreatic cancer cell line XPA-1. The HindIII/NotI fragment of pDsRed2 (Clontech Laboratories, Inc.) was inserted into the HindIII/NotI site of the pLNCX2 plasmid (Clontech Laboratories, Inc.) containing the neomycin resistance gene to produce the PLNCX2-DsRed2 plasmid. PLNCX2-DsRed2 plasmids were transfected into PT67 packaging cells using the LipofectAMINE system (Gibco-BRL), and vector production was accomplished via growth in PT67 packaging cells cultured in the presence of 200-1000 μg/mL Geneticin (G418) from Invitrogen Corporation (Carlsbad, CA) for 7 days. 20% confluent XPA-1 cells were incubated with retroviral supernatants of the packaging cells until high levels of DsRed2 expression were observed under fluorescence microscopy. The cells were then harvested and subcultured in selective medium containing G418. The G418 was increased in a stepwise fashion from 200 μg/mL to 2,000 μg/mL. Clones with high DsRed2 expression were isolated and grown for 10 passages in the absence of G418 to select for stable in vitro expression of DsRed2.
Animal Care
Athymic nu/nu nude mice and transgenic nu/nu mice with the nestin-driven GFP transgene (which express GFP in nestin-positive tissues) (15), both between 4 and 6 weeks of age, were maintained in a barrier facility on high efficiency particulate air (HEPA)-filtered racks. The animals were fed with autoclaved laboratory rodent diet (Teckland LM-485; Western Research Products, Orange, CA). All invasive procedures and imaging were performed with the animals anesthetized by intramuscular injection of 0.02 mL of a solution of 50% ketamine, 38% xylazine and 12% acepromazine maleate. All surgical procedures were performed under sterile conditions. All animal studies were conducted in accordance with the principles and procedures outlined in the NIH Guide for the Care and Use of Animals under Assurance #A3873-01.
Establishing Orthotopic Pancreatic Tumors
Human pancreatic cancer xenografts from the pancreatic cancer cell line XPA1-RFP were established in nude mice by surgical orthotopic implantation (SOI), a procedure developed in our laboratory(16). Briefly, small subcutaneous tumors were initiated in 4-6 week old female nu/nu mice via subcutaneous injection of 1×106 XPA1-RFP cells into the left flank. When these tumors reached approximately 1 cm diameter, the animals were euthanized and the tumor was harvested and sectioned into 1 mm3 pieces. Recipient 4-6 week old female nu/nu mice were then anesthetized as described, and a small transverse incision was then made in the left lateral flank through the skin and peritoneum. The tail of the pancreas was exposed, and a 1 mm3 piece of tumor tissue was sutured into the pancreatic tail using 8-0 nylon surgical suture from US Surgical. The pancreas was then returned to the abdomen, and the peritoneum and skin were closed using 6-0 polysorb surgical suture (US Surgical).
Establishing the High Metastatic Variant
Orthotopic human pancreatic cancer xenografts from the pancreatic cancer cell line XPA1-RFP were initially established in nude mice by SOI as described above. These tumors were allowed to grow until the animals developed widespread metastases and malignant ascites. At the time of sacrifice, the ascitic fluid was harvested and placed on ice. 4-6 week old female nu/nu mice were anesthetized and a small transverse incision was made in the left lateral flank through the skin and peritoneum. The tail of the pancreas was exposed, and 50 μL of malignant ascites were injected into the pancreatic tail. The pancreas was then returned to the abdomen, and the peritoneum and skin were closed using 6-0 polysorb surgical sutures (US Surgical). After six serial passages in this fashion, the resulting primary tumor and malignant ascites were harvested. The primary tumor and malignant ascites were gently dissociated and plated for in vitro culture. After a single in vitro passage for expansion, the passage-6 ascites and passage-6 primary cells were stored in liquid nitrogen for later analysis.
Animal Imaging
Mice were imaged using the Olympus OV100 Small Animal Imaging System (Olympus Corp. Tokyo, Japan), containing an MT-20 light source (Olympus Biosystems Planegg, Germany) and DP70 CCD camera (Olympus Corp. Tokyo, Japan) based on the expression of fluorescent proteins, a procedure developed in our laboratory (17, 18). All images were analyzed using Image-J (National Institute of Health Bethesda, MD) and were processed for contrast and brightness with the use of Photoshop element -4 (Adobe Systems Inc. San Jose, CA).
Blood Vessel Measurement
Tumors harvested from nestin-GFP mice, which express GFP nude in nascent blood vessels as established by our laboratory(15), were weighed and measured for volume and gently compressed between two glass slides as previously described (19). Sequential images at high magnification using the GFP narrow bandpass filter were obtained using the OV100 Small Animal Imaging System. All GFP-expressing blood vessels were measured using Image-J software and calculated as blood vessel density (blood vessel length/tumor volume = mm/mm3). Tumor volume was calculated as length x height x width x 0.52.
Histology
Primary and metastatic tumor samples were removed en bloc with surrounding tissue at the time of sacrifice, fixed in Bouin’s Solution, and embedded in paraffin prior to sectioning and staining with H&E for standard light microscopy. All slides were imaged using an inverted Nikon DE-300 microscope and Spot camera RD. All images were analyzed using Image-J (National Institute of Health Bethesda, MD) and were processed for contrast and brightness with the use of Photoshop element -4 (Adobe Systems Inc. San Jose, CA).
In Vitro Proliferation
All cell lines evaluated were thawed from frozen stocks and assayed within two passages after thaw. The aggressive (generated from either primary tumor or malignant ascites) and parental cell lines (XPA1-RFP) were plated in 96-well plates in triplicate at 5,000 cells per well. Four days later, proliferation was quantified using an XTT assay from Biotium Inc. (Hayward, CA). The XTT developing reagent was added to the wells and allowed to incubate for 4 hours at 37°C. Plates were then read using a standard plate reader at OD 490 nm. OD readings between the three cell lines were evaluated by ANOVA using NCSS Statistical Software (Kaysville, Utah) for statistically significant differences in cell proliferation.
RNA Isolation from Cultured Cells
Total NRA was isolated from aggressive and parental cell lines using the RNeasy kit from Qiagen (Valencia, CA). Briefly, cells were grown in RPMI as described. At 75% confluence, the cells were scraped from the dish, washed once using ice-cold Phosphate Buffered Saline (PBS) from Gibco-BRL, and counted. The cells were then pelleted and homogenized. An equal volume of 70% ethanol was added to the homogenized lysate, and the mixture was applied to an RNeasy midi-spin column. The RNA was washed several times while on the column according to the manufacturer’s instructions and eluted using RNAse-free water. Total RNA concentration and purity in the eluted samples obtained was quantitated using a NanoDrop ND-100 UV spectrophotometer from NanoDrop Technologies, LLC (Wilmington, DE).
cDNA Preparation
Following RNA preparation, the samples were treated with DNase to ensure elimination of genomic DNA, and the extracted RNA was converted to cDNA using the RT2 First Strand Kit from SuperArray Bioscience Corporation (Frederick, MD). Briefly, 25-50 ng RNA was combined with 2 uL gDNA elimination buffer and brought up to a final volume of 10 uL using RNAse-free H20. This mixture was incubated at 42 °C for 5 minutes, then chilled on ice. 10 uL of RT Cocktail was then added to this mixture and incubated at 42 °C for exactly 15 minutes followed by 5 minutes at 95 °C. 91 uL ddH20 was added to each 20 uL cDNA synthesis reaction, and the diluted cDNA mixture was stored at −20 °C until used for gene expression profiling.
Gene Expression Profiling
The RT2 Profiler polymerase chain reaction (PCR) Array System (SuperArray Bioscience Corp.) was used to evaluate the aggressive and parental cell lines for differential gene expression. The genes evaluated in this PCR array include multiple genes involved in angiogenesis and vascular homeostasis (Table 1). Real-time PCR detection was carried out per the manufacturer’s instructions. The experiment cocktail was prepared by adding 550 uL of the SuperArray RT2 qPCR master mix and 448 uL ddH20 to 102 uL of the diluted cDNA mixture. For real-time PCR detection, 10 uL of this cocktail was added to each well of the 384-well PCR array. The array was then cycled on a real-time thermal cycler through the following program: 1 cycle of 10 minutes at 95 °C followed by 40 cycles of 15 seconds at 95 °C and 1 minute at 60 °C. SYBR Green fluorescence was detected from each well during the annealing step of each cycle, and values were exported to a template Excel file for analysis. Analysis of the raw data was performed through the Superarray Data Analysis Web Portal (SuperArray Bioscience Corp.).
Table 1. Multiple genes involved in angiogenesis, including growth factors and receptors, adhesion molecules, proteases, matrix proteins, and transcription factors, were evaluated for expression using real-time PCR via the RT2 Profiler PCR Array System.
Table 1 lists the genes of interest evaluated and their fold increase or decrease in the aggressive cell variant as compared to the parental cell line. In contrast to many of the angiogenesis-related genes evaluated, thrombospondin-1 demonstrated a 24-fold increase in expression in these cells. Complete abbreviations of genes can be found at http://www.sabiosciences.com/.
| Type of Gene | Fold difference (P6/P0) | |||||
|---|---|---|---|---|---|---|
| 0.1-0.5 | 0.5-1.0 | 1.0-2.0 | 2.0-10 | 10-20 | >20 | |
| Angiogenic Factors | VEGFC | PGF | ANGPT 1 | ANPEP | ||
| STAB1 | ANGPT 2 | EREG | ||||
| VEGF A | FGF1 | IL8 | ||||
| TYMP | ||||||
| FGF-2 | ||||||
| FIGF | ||||||
| FLT-1 | ||||||
| JAG-1 | ||||||
| KDR | ||||||
| LAMA5 | ||||||
| NRP1 | ||||||
| NRP2 | ||||||
| PLXDC1 | ||||||
| Adhesion molecules | ANGPTL3 | |||||
| BAI 1 | ||||||
| COL4A3 | ||||||
| Proteases, inhibitors and other molecules | ANGPT4 | PECAM1 | ||||
| PF4 | ||||||
| PROK2 | ||||||
| SERPINF1 | ||||||
| TNFAIP2 | ||||||
| Transcription factors and others | HAND2 | |||||
| SPHK1 | ||||||
| Other factors involved in angiogenesis | ||||||
| Cytokines and chemokines | CXCL10 | CXCL9 | CCL11 | CXCL1 | ||
| IFNB1 | IFNA1 | CCL2 | ||||
| IL6 | CXCL3 | |||||
| MDK | CXCL5 | |||||
| TNF | CXCL6 | |||||
| Other growth factors and receptors | HGF | EFNA1 | EGF | EDG-1 | ||
| ITGB3 | EFNA3 | |||||
| TGFBR1 | EFNB2 | |||||
| EPHB4 | ||||||
| FGFR3 | ||||||
| IGF1 | ||||||
| PDGFA | ||||||
| TEK | ||||||
| TGFA | ||||||
| TGFB1 | ||||||
| TGFB2 | ||||||
| Adhesion molecules | ITGB3 | CDH5 | TSP | |||
| COL18A1 | ||||||
| ENG | ||||||
| ITGAV | ||||||
| THBS2 | ||||||
| Protease inhibitors and other matrix proteins | TIMP2 | LECT1 | LEP | PLAU | ||
| MMP2 | ||||||
| MMP9 | ||||||
| PLG | ||||||
| TIMP1 | ||||||
| TIMP3 | ||||||
| Transcription factors and others | AKT1 | HPSE | ||||
| HIF1A | ID3 | |||||
| NOTCH 4 | ID1 | |||||
| PTGSI1 | ||||||
Results and Discussion
Serial in vivo passage of malignant ascites from human pancreatic cancer-bearing nu/nu mice was used to generate a highly aggressive passage-6 generation cell line with much more rapid primary tumor growth and faster time to distant metastasis. The tumors generated from the parental cell line XPA1-RFP grew slowly (Figure 1A) with eventual metastasis to multiple extrapancreatic sites. Mice bearing these tumors showed widespread metastasis 9 to 12 weeks after implantation (Figure 1B) and died within 12 to 14 weeks (Figure 1C). Repeated in-vivo passage of malignant ascites from XPA1-RFP tumor-bearing mice into the pancreas of recipient nude mice yielded a much more aggressive cell line. Tumors generated from the aggressive variant grew rapidly (Figure 1A) and demonstrated widespread metastasis within 3 to 5 weeks (Figure 1B). Time to death in this group was from 3 to 8 weeks (Figure 1C).
Figure 1. Malignant ascites were isolated from tumor-bearing animals and serially passaged into the pancreas of 4-6 week of female nu/nu mice.
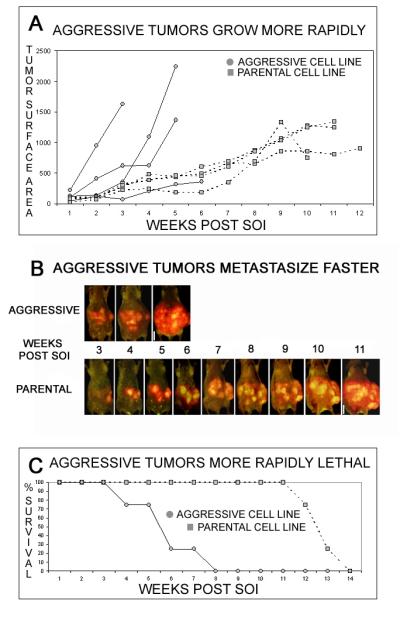
The high in vivo passage cell line (aggressive variant) grew more rapidly in vivo and was more rapidly lethal than the parental cell line XPA1-RFP. Measurement of tumor surface area (measured in mm2) via whole body imaging demonstrated much more rapid primary growth of tumors generated from the aggressive cell line (A). Representative animals from each group imaged using the Olympus OV-100 are shown and demonstrate the earlier time to widespread metastasis in the aggressive variant (B). Survival data from each group demonstrates more rapid mortality in animals bearing tumors from the aggressive cell line (C). The solid line indicates growth of tumors in animals implanted with the aggressive variant, while the dotted line represents the parental line.
Orthotopic parental and aggressive-variant tumors metastasized to distant sites in a similar distribution and yielded histologically similar lesions. Mice bearing tumors generated from the parental cell line XPA1-RFP developed lesions in the liver, spleen, abdominal lymph nodes, mesentery, and lung by 9 to 12 weeks after implantation. Similarly, the aggressive variant metastasized to liver, spleen, abdominal lymph nodes, mesentery, and lung. While the metastatic lesions in the animals bearing tumors from the aggressive variant were found in the same distribution, they developed much more rapidly than tumors generated from the parental cell line, within 3 to 5 weeks of tumor implantation. Microscopic evaluation of the metastatic and primary tumors from animals after implantation of either the parental or aggressive cell lines revealed lesions that were histologically indistinguishable (Figure 2).
Figure 2. The aggressive and parental variant tumors demonstrate similar histology.

Tumors generated from the parental and aggressive cell lines were harvested and evaluated using light microscopy. Primary tumors from the parental cell line (A) and the aggressive cell line (B) were virtually indistinguishable by light microscopy. Likewise, diaphragmatic metastases from the parental cell line (C) and the aggressive cell line (D) demonstrated very similar histology.
In addition to faster growth in vivo, tumors from the aggressive cell line had greater blood vessel density at 8 days after implantation. Cells generated from the aggressive variant were assayed for their early in vivo growth and angiogenesis within the pancreas of transgenic nude mice with nestin-driven GFP in which nascent blood vessels express GFP. At 8 days after implantation, tumors generated from the parental cell line were harvested and compared to those generated from the aggressive variant. The aggressive tumors demonstrated more rapid early growth (Figure 3A) and a greater density of nascent blood vessels within the developing tumor (Figure 3B-D).
Figure 3. Tumors generated from the aggressive and parental cell lines were implanted into the pancreas of nestin-GFP nu/nu mice and allowed to grow for 8 days.
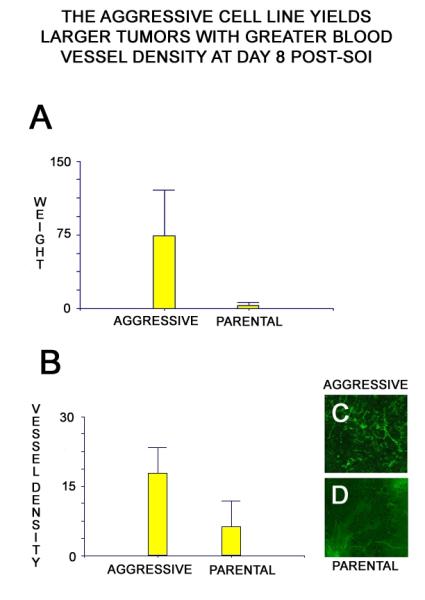
At 8 days post-implantation, the tumors were harvested, weighed, and imaged. Tumors from the aggressive variant were significantly larger at day 8. All tumor weights are measured in mg (A). (p = 0.016 by paired t-test) Tumors from the aggressive cell line demonstrate significantly greater density of GFP-expressing nascent blood vessels at day 8. Tumor blood vessel density is measured in mm/mm3 (B). (p = 0.009 by paired t-test) Representative images from aggressive (C) and parental (D) tumors illustrate the disparity in blood vessel density.
Cell lines generated from the aggressive variant show comparable in vitro morphology and proliferation when compared to the parental cell line. In vitro testing of cell proliferation yielded no significant difference between the parental cell line and cell lines derived from either the primary tumor or malignant ascites of the aggressive variant (Figure 4).
Figure 4. Cell lines derived from the aggressive variant show similar in vitro proliferation when compared to the parental cell line (XPA1-RFP).
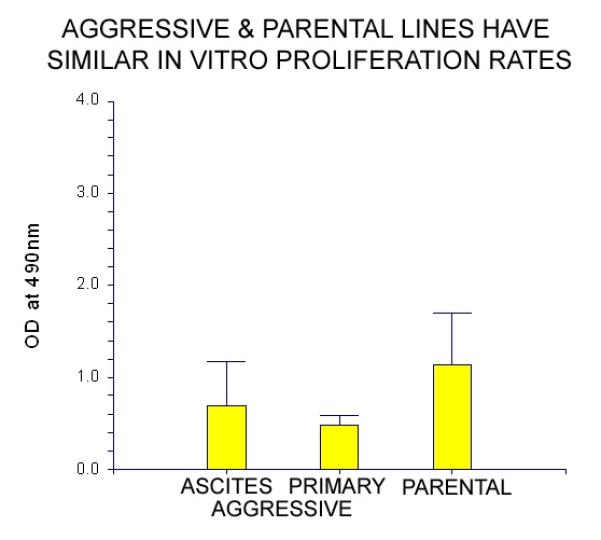
In vitro proliferation was evaluated under standard culture conditions using an XTT assay. No significant difference was seen when cell lines derived from either the primary tumor or the malignant ascites derived from an animal implanted with the aggressive variant were compared to the parental cell line XPA1-RFP. Comparison between the three groups was determined by ANOVA (p=0.15).
Evaluation of gene expression demonstrated elevated levels of thrombospondin-1 expression in the aggressive variant. After a single passage in vitro, RNA was harvested from both the parental (XPA1-RFP) and the aggressive variant. Real-time RT-PCR was used to assay for multiple angiogenic factors including growth factors and receptors, adhesion molecules, proteases, matrix proteins, and transcription factors (Table 1). Upregulation of throbmospondin-1 was demonstrated in cells from the aggressive variant with a 24-fold increase in thrombospondin-1 RNA compared to the parental cell line (Table 1, Figure 5). Other genes showed no remarkable changes in the variant.
Figure 5. The aggressive variant demonstrates increased Thrombospondin-1 (TSP-1) expression at the RNA level.

Cells derived from the aggressive variant and cells from the parental cell line were harvested and RNA was isolated. The RNA expression of a number of genes involved in angiogenesis was then quantified using real-time PCR. Thrombospondin-1 expression was increased 24-fold when compared to the parental cell line (arrow). All bars represent expression in the aggressive cell line divided by expression in the parental cell line (P6/P0).
Multiple-passage selection of ascites of the human pancreatic cancer cell line XPA1-RFP has thereby yielded a population of cells with highly aggressive in vivo growth and metastasis. Although this aggressive variant grew faster within the pancreas of living animals and metastasized earlier to distant sites, the cell line generated from this variant did not grow more rapidly in culture. This suggested that the aggressive nature of this variant was related to something other than increased cell proliferation, and the adaptations responsible for its aggressive in vivo behavior may be related to the pancreatic microenvironment present within the living animal. Indeed, evaluation of early orthotopic tumors generated from the aggressive and parental cell lines revealed larger tumors at 8 days post-implantation with greater blood vessel density in the aggressive tumors. Interestingly, real-time PCR evaluation of cells from this aggressive variant demonstrated a 24-fold increase in Thrombospondin-1 RNA expression when compared to the parental, less aggressive, cell line.
Thrombospondin-1 (TSP-1) is a trimeric 450-kDa glycoprotein present in platelets and the extracellular matrix. This protein participates in platelet aggregation as well as in the tissue response to injury (20, 21). The role of TSP-1 in angiogenesis has been somewhat controversial, with both stimulatory and inhibitory functions reported (13, 14, 22, 23). Qian et al. found that bovine aortic endothelial (BAE) cell tube formation was enhanced by low levels of TSP-1 but inhibited at higher levels (14). TSP-1 mediated this effect on tube formation through the upregulation of matrix metalloproteinase-9 (MMP-9) in the BEA cells. Nicosia et al. showed that matrix-bound TSP-1 was able to promote growth of microvessels and fibroblast-like cells in culture in a dose-dependent fashion (22). TSP-1 has also been shown to be involved in platelet-stimulated smooth muscle proliferation (24), which may play a role in vascular remodeling. Our results suggest an important role for TSP-1 in elevated angiogenesis and metastasis. These findings have important implications for the treatment of metastatic disease.
Future experiments will evaluate protein expression of TSP-1 in the aggressive cell line as compared to both the parental cell line and normal pancreatic tissue, as well as gene and protein expression of TSP-1 in tumors growing within the mouse pancreas. In addition, the time-dependent induction of TSP-1 during tumor development and metastasis will be evaluated, as will the effect of in vitro and in vivo down-regulation of the Thrombospondin-1 gene.
Acknowledgements
This work was supported in part by the following grants: Cancer Therapeutics Training Program (T32 CA121938) (M.B.); National Institutes of Health (CA109949-03) (M.B.); American Cancer Society (RSG-05-037-01-CCE) (M.B.); and National Institutes of Health (CA103563) (AntiCancer Inc.).
Financial Support: Cancer Therapeutics Training Program (T32 CA121938) (M.B.)
National Institutes of Health (CA109949-03) (M.B.)
American Cancer Society (RSG-05-037-01-CCE) (M.B.)
National Institutes of Health (CA103563) (AntiCancer Inc.)
Abbreviation List
- TSP-1
Thrombospondin-1
- RFP
Red Fluorescent Protein
- ANOVA
Analysis of Variance
Footnotes
Conflict of Interest Statement: The authors have no conflicts of interest to report.
References
- 1.Jemal A, Siegel R, Ward E, Murray T, Xu J, Thun MJ. Cancer statistics, 2007. CA Cancer J Clin. 2007;57:43–66. doi: 10.3322/canjclin.57.1.43. [DOI] [PubMed] [Google Scholar]
- 2.Yeo C, Yeo T, Hruban R, editors. Cancer of the Pancreas. 7 ed Lippincott Williams & Wilkins; Philadelphia: 2004. [Google Scholar]
- 3.Sener SF, Fremgen A, Menck HR, Winchester DP. Pancreatic cancer: a report of treatment and survival trends for 100,313 patients diagnosed from 1985-1995, using the National Cancer Database. J Am Coll Surg. 1999;189:1–7. doi: 10.1016/s1072-7515(99)00075-7. [DOI] [PubMed] [Google Scholar]
- 4.Hanahan D, Weinberg RA. The hallmarks of cancer. Cell. 2000;100:57–70. doi: 10.1016/s0092-8674(00)81683-9. [DOI] [PubMed] [Google Scholar]
- 5.Qian LW, Mizumoto K, Maehara N, et al. Co-cultivation of pancreatic cancer cells with orthotopic tumor-derived fibroblasts: fibroblasts stimulate tumor cell invasion via HGF secretion whereas cancer cells exert a minor regulative effect on fibroblasts HGF production. Cancer Lett. 2003;190:105–12. doi: 10.1016/s0304-3835(02)00517-7. [DOI] [PubMed] [Google Scholar]
- 6.Gasparini G. The rationale and future potential of angiogenesis inhibitors in neoplasia. Drugs. 1999;58:17–38. doi: 10.2165/00003495-199958010-00003. [DOI] [PubMed] [Google Scholar]
- 7.Beckermann BM, Kallifatidis G, Groth A, et al. VEGF expression by mesenchymal stem cells contributes to angiogenesis in pancreatic carcinoma. Br J Cancer. 2008;99:622–31. doi: 10.1038/sj.bjc.6604508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Yamanaka Y, Friess H, Buchler M, et al. Overexpression of acidic and basic fibroblast growth factors in human pancreatic cancer correlates with advanced tumor stage. Cancer Res. 1993;53:5289–96. [PubMed] [Google Scholar]
- 9.Kamisawa T, Isawa T, Koike M, Tsuruta K, Okamoto A. Hematogenous metastases of pancreatic ductal carcinoma. Pancreas. 1995;11:345–9. doi: 10.1097/00006676-199511000-00005. [DOI] [PubMed] [Google Scholar]
- 10.Karademir S, Sokmen S, Terzi C, et al. Tumor angiogenesis as a prognostic predictor in pancreatic cancer. J Hepatobiliary Pancreat Surg. 2000;7:489–95. doi: 10.1007/s005340070020. [DOI] [PubMed] [Google Scholar]
- 11.Homsi J, Daud AI. Spectrum of activity and mechanism of action of VEGF/PDGF inhibitors. Cancer Control. 2007;14:285–94. doi: 10.1177/107327480701400312. [DOI] [PubMed] [Google Scholar]
- 12.Fukumura D, Jain RK. Tumor microvasculature and microenvironment: targets for anti-angiogenesis and normalization. Microvasc Res. 2007;74:72–84. doi: 10.1016/j.mvr.2007.05.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Volpert OV. Modulation of endothelial cell survival by an inhibitor of angiogenesis thrombospondin-1: a dynamic balance. Cancer Metastasis Rev. 2000;19:87–92. doi: 10.1023/a:1026560618302. [DOI] [PubMed] [Google Scholar]
- 14.Qian X, Wang TN, Rothman VL, Nicosia RF, Tuszynski GP. Thrombospondin-1 modulates angiogenesis in vitro by up-regulation of matrix metalloproteinase-9 in endothelial cells. Exp Cell Res. 1997;235:403–12. doi: 10.1006/excr.1997.3681. [DOI] [PubMed] [Google Scholar]
- 15.Amoh Y, Yang M, Li L, et al. Nestin-linked green fluorescent protein transgenic nude mouse for imaging human tumor angiogenesis. Cancer Res. 2005;65:5352–7. doi: 10.1158/0008-5472.CAN-05-0821. [DOI] [PubMed] [Google Scholar]
- 16.Hoffman RM. Orthotopic metastatic mouse models for anticancer drug discovery and evaluation: a bridge to the clinic. Invest New Drugs. 1999;17:343–59. doi: 10.1023/a:1006326203858. [DOI] [PubMed] [Google Scholar]
- 17.Chishima T, Miyagi Y, Wang X, et al. Cancer invasion and micrometastasis visualized in live tissue by green fluorescent protein expression. Cancer Res. 1997;57:2042–7. [PubMed] [Google Scholar]
- 18.Hoffman RM. The multiple uses of fluorescent proteins to visualize cancer in vivo. Nat Rev Cancer. 2005;5:796–806. doi: 10.1038/nrc1717. [DOI] [PubMed] [Google Scholar]
- 19.Amoh Y, Li L, Tsuji K, et al. Dual-color imaging of nascent blood vessels vascularizing pancreatic cancer in an orthotopic model demonstrates antiangiogenesis efficacy of gemcitabine. J Surg Res. 2006;132:164–9. doi: 10.1016/j.jss.2005.12.028. [DOI] [PubMed] [Google Scholar]
- 20.Lawler JW, Slayter HS, Coligan JE. Isolation and characterization of a high molecular weight glycoprotein from human blood platelets. J Biol Chem. 1978;253:8609–16. [PubMed] [Google Scholar]
- 21.Raugi GJ, Olerud JE, Gown AM. Thrombospondin in early human wound tissue. J Invest Dermatol. 1987;89:551–4. doi: 10.1111/1523-1747.ep12461198. [DOI] [PubMed] [Google Scholar]
- 22.Nicosia RF, Tuszynski GP. Matrix-bound thrombospondin promotes angiogenesis in vitro. J Cell Biol. 1994;124:183–93. doi: 10.1083/jcb.124.1.183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Sharghi-Namini S, Fan H, Sulochana KN, et al. The first but not the second thrombospondin type 1 repeat of ADAMTS5 functions as an angiogenesis inhibitor. Biochem Biophys Res Commun. 2008;371:215–9. doi: 10.1016/j.bbrc.2008.04.047. [DOI] [PubMed] [Google Scholar]
- 24.Ichii T, Koyama H, Tanaka S, et al. Thrombospondin-1 mediates smooth muscle cell proliferation induced by interaction with human platelets. Arterioscler Thromb Vasc Biol. 2002;22:1286–92. doi: 10.1161/01.atv.0000024684.67566.45. [DOI] [PubMed] [Google Scholar]