

Abstract
Turris babylonia (Linnaeus, 1758) is the designated type species of Turris, the nominate genus of the family Turridae. This species has unusual taxonomic significance, since the family Turridae is a large biodiverse group that has been highly problematic in its taxonomy. In this article, we address the identity of Turris babylonia: molecular data presented here and expanded elsewhere demonstrate that two distinctive varieties with divergent shell morphology, both conventionally assigned to Turris babylonia, are in fact different species. We describe one of the forms as Turris assyria, new species. Thus, specimens previously assigned to Turris babylonia now comprise at least two taxa, Turris babylonia and Turris assyria; it remains possible that each is a multi-species complex. Some of the numerous varieties and morphologically divergent forms in each complex may prove not to be conspecific with the two species, each precisely defined in this work by a specific barcode sequence.
Keywords: Turrids, type species, taxonomy, venomous molluscs, conoideans, barcode sequences
INTRODUCTION
The molluscs in the class Gastropoda (snails and slugs) are among the most diverse and species-rich groups of marine animals. A comprehensive sampling of gastropods from several tropical Pacific sites by Bouchet and co-workers (Bouchet et al. 2002, Bouchet et al. 2009) has revealed the major contribution to gastropod biodiversity made by a group of predatory, venomous marine snails, known as the “turrids” (or “turriform conoideans”). Traditionally, these were assigned to a single family, Turridae. However, “turrids” are currently regarded as a heterogeneous, taxonomically-problematic group. The prior taxonomic scheme was based purely on shell characters; up to 13 subfamilies and hundreds of different genera were included in the family Turridae, as defined in the work of Powell more than four decades ago (Powell 1966).
A revised taxonomy for the turrids was proposed (Taylor et al. 1993) and widely adopted (Bouchet and Rocroi 2005) after other characters, notably foregut anatomy were evaluated. However, recent molecular phylogenetic data (Puillandre et al. 2008) suggest that further revision in the definition of the family Turridae is likely, as additional data are collected and analyzed. A recent proposal placed different lineages of turrids in two different superfamilies, Turroidea and Conoidea (Tucker and Tenorio 2009).
Turris (Röding, 1798), is the type genus of the Turridae, with Turris babylonia (Linnaeus, 1758), the “tower of babel”, as the designated type species. Thus, deciding which genera are properly within the family Turridae, and which species belong to the genus Turris necessarily requires evaluation of the perceived evolutionary relationship to the type species, Turris babylonia. Presumably, Turris babylonia would also be the reference species for the newly proposed superfamily Turroidea (Tucker and Tenorio 2009). However, a recent molecular phylogenetic analysis of several Turris species revealed that the very identity of this type species is ambiguous. As detailed below, two morphospecies, previously treated as variants of Turris babylonia and generally assigned to that species throughout the taxonomic literature proved to have very significantly divergent barcode sequences, establishing unequivocally that these are different species.
There are a series of specimens of Turris babylonia that were recognized by Powell as the “typical form” (see Figure 1A). There are two syntypes in the British Museum (Natural History) in London that correspond to this form, as well as a juvenile specimen, which was the holotype of Pleurotoma venusta Reeve, 1843, assigned by Powell to synonymy with Turris babylonia. Powell recognized that there were a number of morphologically-diverse forms that he had assigned to Turris babylonia; in his comprehensive treatment of the genus Turris (Powell 1964), he remarked: “Some rather distinct forms of this species occur, but since the material available to me is scant, and further, since these variants upon the available evidence do not seem to relate exclusively to geographical areas, it is not possible to judge if the several forms have arisen in response to ecological conditions, segregation, or are accountable just as variation within acceptable species limits.”
Figure 1.

A. Turris babylonia, Danajon Bank, Cebu, Philippines; B. Turris assyria, Danajon Bank, Cebu, Philippines.
The specimen shown in Figure 1B was treated by Powell (1964) in a manner that implied it was one of the distinct forms, morphologically distinguishable from typical Turris babylonia; he illustrated similar specimens (Plate 181, Figures 3 and 4) that contrasted with the typical form (illustrated in Figures 1 and 2 in the same Plate). Powell described and figured two other variants (the “sparsely pale spotted form” and the “narrow form”), these were illustrated as Figures 5 and 6 of Plate 181 in Powell’s work.
In a previous analysis of Turris species from the Philippines (Olivera 1999), the typical form of Powell was referred to as the “Babylon form”, and the form in Figure 1B was referred to as the “Niniveh form”. Because these forms were consistently separable when all the available Philippine material was examined, it was suggested at that time that the two forms would likely prove not to be conspecific upon further evaluation.
MATERIALS AND METHODS
Most of the specimens of the Turris babylonia complex described in this study were collected in the Central Philippines (Cebu – Bohol area); living specimens that provided the material for DNA analysis were mostly collected in April – December, 2009. After shells were photographed, a sample of the soft body was extracted from some specimens and preserved in ethanol for molecular studies. The analysis of the barcode sequence was carried out as described previously (Olivera et al. 2008).
A large series of specimens of Turris babylonia and related forms were examined, including those in the Academy of Natural Sciences in Philadelphia, the collection that was the source of many of the actual specimens assigned to Turris babylonia in Powell’s monograph (Powell 1964).
RESULTS
Barcode analysis
In Figure 1, specimens from which DNA was extracted and from which the sequences of three standard genes (COI, 12S and 16S) were initially obtained are illustrated. Partial barcode sequences from the two specimens shown in Figure 1 are shown in Table 1; the surprising divergence in the barcode sequences shown led to the analysis of additional specimens, shown in Figure 2, top row.
Table 1.
Partial barcode sequences of specimens shown in Figure 2, top row.
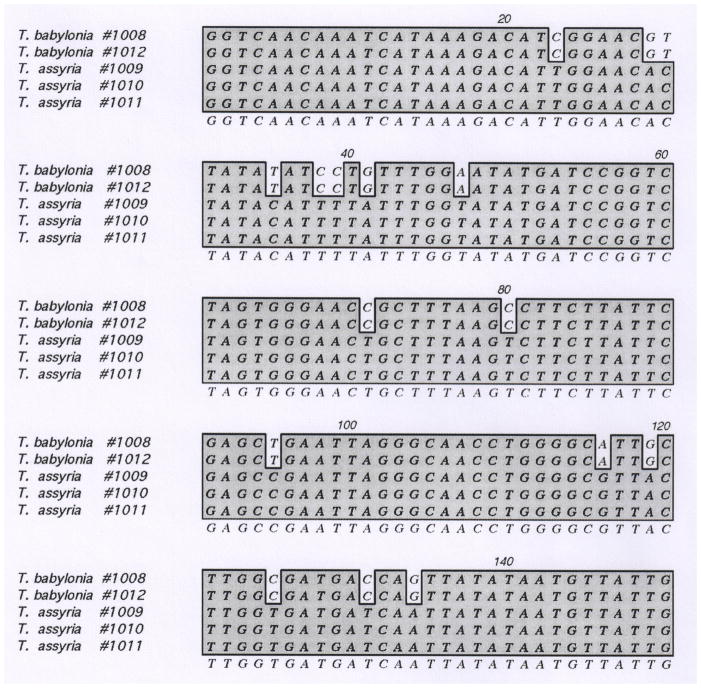
|
The first 150 nucleotides at the 5′ end of the barcode sequences (Cox-I) have been analyzed for five specimens in the Turris babylonia complex. The specimens from which these DNA sequences were derived are illustrated in Figures 1 and 2. The DNA samples were prepared as described under Materials and Methods. Note the significant divergence between the two groups of specimens (#1008 and #1012 vs. #1009–1011), providing unequivocal molecular evidence that these are not conspecific. The holotype for Turris assyria, new species (see Figure 3) was deliberately chosen to be as similar to one of the specimens analyzed above (#1009–1011) that have identical barcode sequences; it is closest to #1009. The sequences have been submitted to GenBank; #1008 is GU299968, #1010 is GU299969, #1012 is GU299970. The sequences of #1009 and #1011 are identical to #1010.
Figure 2.

Top row: Shells of specimens from which the barcode sequences shown in Table 1 were obtained. Specimens A and B are typical Turris babylonia; C, D and E are Turris assyria (see text). Bottom row: Six specimens of Turris assyria, all collected in Dec., 2009, from the Danajon Bank, Cebu, Philippines (see Table 2), the “turrid fishing grounds”. These specimens are not shown proportional to their actual size. Specimen B was from off Sogod, Cebu, Philippines.
The molecular data obtained, shown in Table 1, demonstrate that the specimen in Figure 1B and Figure 2C–E, the “Niniveh form”, is indeed a different species from the typical form of Turris babylonia (Figure 1A and Figure 2A–B). In the 150 nucleotide segment shown, there is >10% divergence in sequence between the two classes of specimens. The justification for separating the two forms into separate species is independently supported by the analysis of other standard marker genes (Fedosov et al. unpublished results). In the next section, we describe this distinctive “Niniveh form” as a new species; it is definitively separated from Turris babylonia by the significant divergence in their barcode sequences.
Systematics: Taxonomic Description of the New Species Turris assyria, new species
Range
Philippines; from the Solomon Islands and Queensland, north to Japan.
Description
Shell turriform, consists of 12 teleoconch whorls, protoconch missing in the holotype. Shell surface glossy, axial sculpture absent. First five whorls somewhat corroded. Succeeding whorls possess four distinct spiral cords, with brown maculations, among which first and third are stronger and wider than the others. The third spiral cord is notably thickened, forming a distinct keel that marks the periphery and gives the whorl its characteristic angular shape. The last whorl and canal bear 12 major maculated spiral cords, separated by minor sharp intermediate white threads, with additional white threads towards the tip of the canal. The siphonal canal is relatively long and narrow, and the distinct cords with maculations in the canal are darker brown in color than in the rest of the body whorl.
The suture is impressed and the subsutural ramp is not pronounced. The shell is white, has regular brown maculations on the upper spiral cord of each whorl followed by short vertical light-brown strokes on the succeeding sinus cord and somewhat larger maculations on the peripheral cord of each whorl (14 on the body whorl). The aperture is ovate, outer lip is evenly rounded; the anal sinus is deep, situated right above the periphery of the whorl. The aperture is white, interior smooth near the lip, with numerous parallel white spiral lirae deeper within the aperture.
Although the protoconch of Turris assyria is corroded in the holotype, one of the specimens (# 1010) for which a barcode sequence was obtained, had a reasonably well-preserved protoconch. The paucispiral protoconch is amber in color, with ~ 1.5 whorls. The early teleoconch whorls retain the amber coloration with strong whitish spiral cords that contrast with the darker amber color between the cords.
Comparison of typical Turris assyria to Turris babylonia (after Olivera 1999, and Powell 1964)
The shell of Turris assyria is proportionally narrower in outline than Turris babylonia. The color pattern is generally more sparse; in the sub-sutural cord the dark brown maculations are characteristically smaller in Turris assyria and bolder in Turris babylonia (see Figures 1 and 2). Adult shells of T. assyria are generally 60–90mm, elongate fusiform and rather smooth, with a shiny porcellaneous texture (Philippine specimens of Turris babylonia tend to have a “matte” surface). The maculations of the typical form of Turris assyria are generally more uniform in size and distribution than in Turris babylonia; the latter has very much bolder markings in the subsutural cord. There is some variation in the color of the maculations, from light brown to almost black in Turris assyria specimens (see Figure 2 for variation from the type locality). Individual whorls are generally less bulbous than in Turris babylonia.
Although in both species, the sinus cord is narrower than either the subsutural cord or the peripheral cord, the contrast in cord width is much greater in Turris babylonia, with a very thin sinus cord and an extremely broad and boldly maculated subsutural cord. In Turris assyria, the sinus cord is proportionally broader and more rounded in outline (see Figures 1 and 2). Another characteristic difference is that the region from the sinus cord to the subsutural cord is brown to off white in most (but not all) specimens of T. babylonia, and generally white in T. assyria.
Live specimens of the two forms (T. assyria and T. babylonia) from Danajon Bank and off Sogod Cebu, Philippines are illustrated in Figures 1 and 2. The two species have animals with somewhat different coloration in their foot —Turris assyria is yellowish in color, while the animal of Turris babylonia collected at the same locality is darker gray with small irregular darker spots. There is some variation in the pattern of the foot — Turris assyria specimen #1011 (Figure 2E) had large grey spots, absent in the other two specimens (see Figures 2C and D), but all three of the Turris assyria specimens shown in Figure 2 yielded identical barcode sequences.
Type locality; geographical distribution
The holotype and three paratype specimens of Turris assyria were collected at a locality in the Central Philippines known as the “turrid fishing grounds”, by divers using compressors at night at depths of 10–16 fathoms. This locality, the outer barrier reef of Danajon Bank between Cebu and Bohol province, east of Olango Island is shown in the rectangle in the map in Figure 3. On four consecutive nights, on December 3–6, 2009, of 81 Turris specimens collected, 8 were Turris assyria — other congeners found are shown in Table 2. No specimens of Turris babylonia were collected during this period, although the Turris babylonia specimen in Figure 1A was found at this locality previously. The typical substrate on which this species is collected is described as “sandy rubble”.
Figure 3.

Turris assyria, new species: holotype specimen and type locality. Top: Shell of holotype. Bottom: Shown in the white rectangle is the location of the turrid fishing ground on the western part of the outer barrier reef, Danajon Bank. The islands east of the white rectangle are Olango and Mactan (closest to Cebu).
Table 2.
Frequency of Turris Species from one field trip at the “Turrid Fishing Ground”
| Turrid Species | No. of specimens |
|---|---|
| Turris assyria | 8 |
| Turris cryptorrhaphe | 4 |
| Turris normandavisoni | 38 |
| Turris spectabilis | 17 |
| Turris garnonsii | 13 |
| Turris dollyae | 1 |
| 81 |
Specimens collected December 3–6, 2009 from the outer barrier reef, Danajon Bank, Cebu portion (see Figure 2).
- Out of the 1583 specimens of all turrids collected for 4-night harvesting (average of 5 hours per night) from the turrid fishing ground in outer barrier reef, 81 were assigned to the genus Turris.
- No Turris babylonia was collected during this harvesting period.
Turris assyria is a relatively common species in the Philippines, with significant morphological variation observed (see Figures 4 and 5). The species appears to be restricted to the Western and Central Pacific; there is no evidence for its occurrence in the Indian Ocean. Specimens tentatively assigned to Turris assyria have been found in the Solomon Islands, as has typical Turris babylonia (see Figures 5 and 6). In addition, as shown in Figure 5, specimens referred to this species also occur in Queensland, Australia; one of the specimens with this locality label appears very atypical, but is most likely an unusually light-colored, small form of Turris assyria (see Figure 5, middle row, 2nd from right). A typical specimen of Turris assyria (identified as Turris babylonia) is illustrated in a recent book on the shells of Japan (Okutani 2000) suggesting its occurrence in the Northern Pacific.
Figure 4.

Variation in Turris assyria. Top and middle: from typical to “sparsely spotted” patterns (“variety 1”, see text) are shown (all of the specimens are from the Central Philippines); Bottom: “variety 2”, with darker maculations (see text).
Figure 5.

Atypical specimens from various localities, some only provisionally assigned to Turris assyria. The wide range of variation in shell morphology and patterns is illustrated in this series. Most of the specimens are likely variants of Turris assyria; two of the more problematic, discussed specifically in the text, are the “narrow form” (leftmost specimen, top row) and the “Sulu Sea form” (rightmost specimen, middle row). Most specimens are from various Philippine localities except top row, extreme left: Moluccas, second from left: Lady Musgrave Is., Australia; Middle row, third from left, Queensland; Bottom row, extreme left, Marau Sound, Guadalcanal.
Figure 6.

Turris babylonia, illustrating morphological variation. All are from various Philippine localities except for the two on the bottom row, right, which are from the Solomon Islands.
Etymology
Since this species has been confused with Turris babylonia, we assign a name, Turris assyria that refers to the other great power in Mesopotamia at the time of Babylon.
Types
The holotype and paratypes 1–3 were collected on the outer reef, Danajon Bank between Cebu and Bohol Islands. The barcode sequences shown in Table 1 are all from Turris assyria from the type locality (because their shells were crushed to obtain tissue for analysis — see Figures 1 and 2, many of the specimens collected at the type locality could not be included in the type series). The type specimens, with shells unbroken, are summarized in Table 3.
Table 3.
Types Turris assyria, new species
| Collection Locality | Length (mm) | Maximum Diameter (mm) | Depository (Cat. #) | |
|---|---|---|---|---|
|
| ||||
| Holotype: | Outer barrier reef, Danajon Bank, Cebu, PI. | 65.2 | 16.2 | PBRC-MSI (GM1001) |
| Paratype: | ||||
| #1 | Outer barrier reef, Danajon Bank, Cebu, PI. | 65.2 | 16.0 | PBRC-MSI (GM1002) |
| #2 | Outer barrier reef, Danajon Bank, Cebu, PI. | 75.0 | 17.9 | PBRC-MSI (GM1003) |
| #3 | Outer barrier reef, Danajon Bank, Cebu, PI | 68.1 | 17.1 | MNHN (21134) |
| #4 | Off Olango Is., Cebu, PI | 60.0 | 15.4 | Field Museum, Chicago |
| #5 | Jagoliao Is., N.W. Bohol, PI (12–24 ft, 7/29/58) | 78.7 | 20.6 | ANSP (231320) |
| #6 | Calatagan, Batangas, Luzon Is., PI | 67.0 | 17.7 | ANSP (229059) |
| #7 | Calatagan, Batangas, Luzon Is., PI | 63.2 | 15.2 | ANSP (229059) |
| #8 | Calatagan, Batangas, Luzon Is., PI | 62.2 | 15.7 | ANSP (229059) |
| #9 | Kudat district, North Borneo, Malaysia | 59.8 | 15.2 | ANSP (255716) |
| #10 | Calituban, Bohol, PI | 68.7 | 18.6 | Russian Academy of Sciences |
| #11 | Mindoro, PI | 70.1 | 18.9 | Olivera collection |
| #12 | (Philippines) | 74.3 | 18.9 | Olivera collection |
| #13 | (Philippines) | 76.2 | 20.0 | Olivera collection |
| #14 | (Philippines) | 85.0 | 21.1 | Olivera collection |
| #15 | (Philippines) | 88.5 | 22.0 | Olivera collection |
| #16 | Off Olango Is., Cebu, PI | 57.7 | 14.4 | Olivera collection |
| #17 | Sogod Bay, Leyte, PI | 60.5 | 15.5 | Olivera collection |
| #18 | Albuera, Leyte, PI (G. Poppe, 12m, at night) | 64.9 | 16.7 | Olivera collection |
| #19 | North Mindinao, PI (?) | 68.3 | 17.3 | Olivera collection |
| #20 | Off Olango Is., Cebu, PI | 78.3 | 18.4 | Olivera collection |
| #21 | (Philippines) | 88.5 | 20.0 | Olivera collection |
| #22 | (Philippines) | 84.0 | 21.0 | Olivera collection |
| #23 | Mactan, Cebu, PI | 69.0 | 17.5 | Olivera collection |
PBRC MSI — Philippine Biodiversity Resources Center, Marine Science Institute, University of the Philippines, Quezon City, Philippines
MNHN — Muséum national d’Histoire naturelle, Paris, France
ANSP — Academy of Natural Sciences, Philadelphia, PA, USA
(Specimens in the Olivera collection are intended for donation to additional museum collections.)
Distinctive varieties of Turris assyria
Turris assyria specimens from the Dajanon Bank, Cebu, the type locality shown in the map in Figure 3, and from Sogod, Cebu, are mostly the typical form. A number of distinctive varieties are assigned to this species that diverge to varying extents from the typical form.
Variety 1
In this variety (see Figure 4, middle row), in contrast to the regular pattern of darker brown maculations found in the subsutural cord and the basal cords, those on the sinus cord, the peripheral cord and the more anterior major cord are either paler brown and/or much more sparse in pattern; in some specimens, maculations on these cords may be missing altogether. The cords on the canal typically have darker and bolder maculations. We regard these as varieties of Turris assyria, since there appear to be intermediate forms (see Figure 4, top row). Indeed, the holotype of Turris assyria has some intermediate characteristics. Small specimens of this variety are often found dead on the beaches of the Central Philippines. This variety is regularly found off Calituban, Bohol. Powell (1964) referred to a pale, sparsely spotted form from New Guinea, similar in pattern to this variety; the New Guinea specimens appear to be broader than the Philippine material, and may not be conspecific with Turris assyria.
Variety 2
This variety, shown in Figure 4, bottom row, has generally darker maculations than the typical form, and has a longer canal; specimens of this variety are among the largest specimens from the Philippines, with adults >85mm in length. As shown in Figure 2, this variety represents a minor fraction of specimens from the type locality (Figure 2, bottom row, 2nd from left).
Specimens provisionally assigned to Turris assyria
A variety of specimens are shown (Figure 6) that are rather divergent from the typical form of Turris assyria. However, these specimens all appear closer to T. assyria than to T. babylonia. Nevertheless, there remains the possibility that some of these will prove not to be conspecific with Turris assyria after more detailed analysis. Two classes of these specimens that appear particularly distinctive are discussed:
Narrow form
As described above, specimens assigned to Turris assyria have a relatively broad sinus cord (relative to the subsutural and peripheral cords) compared to specimens assigned to Turris babylonia (where characteristically, the sinus cord is very much narrower than the subsutural cord). There are a group of specimens with exceptionally narrow sinus ribs that resemble T. assyria in other respects (e.g., a specimen from the Moluccas, Figure 5, top row leftmost). A molecular assessment is desirable; this was also referred to by Powell (1964) as a distinctive form (see Plate 181, Figure 5).
Sulu Sea form
This distinctive form (Figure 5, middle row, leftmost specimen) is characteristic of specimens primarily collected from localities around the Sulu Sea. These specimens are generally smaller than either typical Turris assyria and Turris babylonia. Whether or not this form is conspecific with Turris assyria remains to be evaluated further.
Review of Prior Literature
Because the new species, Turris assyria, was previously regarded as a form of Turris babylonia, previous publications that either discussed or illustrated Turris babylonia were inconsistent with respect to which morph was referred to or illustrated. The most comprehensive discussion of Turris babylonia was published by Powell (1964). He documented 8 specific localities for the species in the Philippines; one was a reference to the type specimen of Pleurotoma venusta (Siquijor Island), which as discussed above, is a synonym of Turris babylonia.
The other Philippine specimens referred to by Powell were from the collection of the Academy of Natural Sciences of Philadelphia (ANSP); we have examined the specimens (comprising 13 individual shells from 7 stations). What is striking is that only one of the 13 is a typical Turris babylonia; the other 12 are, in fact, specimens tentatively assigned to Turris assyria. These include the two specimens in the color photograph published in his article (Plate 181, Figures 3 and 4). The only typical Turris babylonia from the Philippines was from Sanga-sanga, Sulu Archipelago — although Powell indicated that this was part of the Dupont/Academy expedition, it was not, in fact, collected by that expedition, but was a gift received during the expedition from John Root, an important collector of Philippine material in the 1950s. All Philippine specimens actually collected directly by the expedition and labeled Turris babylonia have proven to be Turris assyria. Thus, all of the other records of Powell from the Philippines are likely records for Turris assyria.
Powell referred to additional specimens in the ANSP collection from outside the Philippines. Two were from Malaita Island in the Solomon group, and the other was collected from the Admiralty Island; none of the three specimens were Turris assyria — these are all typical Turris babylonia.
In the first comprehensive illustration of shells from the Philippines, Springsteen and Leobrera (1986) did indeed show typical Turris babylonia (plate 76, # 1). However, in the Compendium of Seashells, Abbott and Dance (1986) identified two specimens as Turris babylonia. The rightmost specimen on page 239 is that species, but the specimen on the left is similar to the “narrow form” of Powell (1964) that is provisionally assigned to Turris assyria. In the recent book on Philippines shells by Poppe (2008), there are three specimens illustrated; two of these (plate 680, Figures 6 and 7) are typical Turris babylonia, but Figure 8 is Turris assyria. The illustrated specimen assigned to Turris babylonia in a recent book illustrating the seashells of the world by Robin (2008) (page 448, Figure 3) is Turris assyria. On Figure 8 of the same plate, there is a specimen likely to be a dark variety of Turris babylonia, identified as Turris hidalgoi, a different species (Vera-Peláez et al. 2000). Thus, the prior literature illustrates the considerable confusion with regard to which forms are properly included within the boundaries of the type species, Turris babylonia.
DISCUSSION
As Powell remarked, the specimens that he had assigned to Turris babylonia were a morphologically heterogeneous group. The molecular evidence provides unequivocal proof that two of the forms previously regarded as Turris babylonia are different species. As shown in Figure 2, shells of Turris babylonia and the new species, Turris assyria collected from Cebu in the Central Philippines, are easily distinguished by shell morphology. The potential confusion arises from variation found in these species; a large number of variants potentially belong to one or the other species (see Figures 4–6).
However, examination of a large series of specimens from the Philippines demonstrated that it was generally straightforward to separate specimens into two groups that we have tentatively assigned either to Turris babylonia or Turris assyria. We also have specimens assigned to both species from the Solomon Islands. Thus, across the central Indo-Pacific, from the Philippines to the Solomons, variants of both species are probably present. We illustrated above specimens that we assigned to Turris assyria from Queensland; typical Turris babylonia has not been reported from Australia. Similarly, Turris assyria was illustrated in the volume by Okutani (2000) on the shells of Japan (and identified as Turris babylonia); we have been unable to find a figure of a specimen of Turris babylonia from Japan. Thus, although the range of the two species mostly overlaps, the preliminary data suggest that Turris assyria may be more widely distributed across the Indo-Pacific.
The molecular data (Table 1) indicate that Turris assyria and Turris babylonia are surprisingly divergent at the molecular level. The field data at the type locality, the “turrid fishing grounds” at Danajon Bank (see Table 2 for an example) suggest that in addition to Turris assyria, Turris normandavisoni, Turris spectabilis and Turris garnonsii are also regularly found at this locality. A definition of the divergence in prey and/or microhabitat between these congeners should be feasible if systematic field collection data were carried out at this locality. At the present time, little data exists in the literature that sheds light on the species-specific biology of the different Turris species.
SUMMARY OVERVIEW
Two distinct morphospecies were previously assigned to Turris babylonia (see Poppe 2008, Powell 1964). One of these, previously called the “Niniveh form” by Olivera (1999), is designated a new species, Turris assyria. Specimens assigned to Turris assyria comprise a set with considerable variation in shell morphology. From any specific locality, Turris assyria and Turris babylonia are easily separable on the basis of shell morphology; however, each comprises forms with considerable morphological variation. The possibility that some of these are different species, but morphologically very similar to Turris assyria and Turris babylonia, cannot be eliminated at this time. A more precise phylogenetic analysis of the status of these two species within the genus Turris using multiple molecular markers has been carried out; this more extensive molecular phylogeny of Turris will be published elsewhere (Fedosov et al. unpublished results).
Acknowledgments
This work was supported in part by PharmaSeas, a grant to Dr. Giselle Concepcion from the Department of Science and Technology, Republic of the Philippines and by Grant GM48677, from the U.S. National Institute of General Medical Sciences (to BMO). We are grateful to Maren Watkins for the molecular analysis, to Mary Anne O. Astilla and Frank Heralde for their camaraderie during fieldwork and to Giselle Concepcion, Perry Aliño, and Yuri Kantor for their support and encouragement.
Kerry Matz took some of the photographs and Michael Miller measured the type specimens. We thank Gary Rosenberg and Paul Calloman for the loan of specimens from the ANSP.
Footnotes
CONFLICT OF INTEREST
There is no conflict of interest relating to this work.
CONTRIBUTION OF INDIVIDUAL AUTHORS
B. M. Olivera framed the scientific hypothesis and organized the study; R. A. Seronay contributed to the field studies; A. E. Fedosov researched prior taxonomic literature and dissected the specimens for molecular analysis.
References
- Abbott RT, Dance SP. Compendium of Seashells. Melbourne, Florida: American Malacologists, Inc; 1986. p. 239. [Google Scholar]
- Bouchet P, Lozouet P, Maestrati P, Heros V. Assessing the magnitude of species richness in tropical marine environments: High numbers of molluscs at a New Caledonia site. Biol J Linnean Soc. 2002;75:421–436. [Google Scholar]
- Bouchet P, Lozouet P, Sysoev AV. An inordinate fondness for turrids. Deep-Sea Research II. 2009;56(19–20):1724–1731. [Google Scholar]
- Bouchet P, Rocroi JP. Malacologia: International Journal of Malacology, Classification and Nomenclator of Gastropod Families. Malacologia - International Journal of Malacology ConchBooks. 2005;47(1–2):256–257. [Google Scholar]
- Okutani T. Marine Mollusks in Japan. In: Okutani T, Tomigaya, Shibuya-ku, editors. Tokyo, Japan: Tokai University Press; 2000. p. 630. [Google Scholar]
- Olivera BM. The subfamily Turrinae in the Philippines: The genus Turris (Röding, 1798) Phil J Sci. 1999;128:295–318. [Google Scholar]
- Olivera BM, Hillyard DR, Watkins M. A new species of Gemmula, Weinkauff 1875; evidence of two clades of Philippine species in the genus Gemmula. Phil Sci Lett. 2008;1(1):11–15. [Google Scholar]
- Poppe GT. Philippine Marine Mollusks. In: Poppe GT, editor. (Gastropoda - Part 2) ConchBooks. 2008. pp. 770–771. [Google Scholar]
- Powell AWB. The family Turridae in the Indo-Pacific. Part 1. The subfamily Turrinae. Indo-Pacific Mollusca. 1964;1:227–346. [Google Scholar]
- Powell AWB. The molluscan families Speightiidae and Turridae. Bull Auckland Inst Museum. 1966;5:1–184. + 123 plates. [Google Scholar]
- Puillandre N, Samadi S, Boisselier MC, Sysoev AV, Kantor YI, Cruaud C, Couloux A, Bouchet P. Starting to unravel the toxoglossan knot: Molecular phylogeny of the “Turrids” (Neogastropoda: Conoidea) Mol Phylogenet Evol. 2008;47(3):1122–1134. doi: 10.1016/j.ympev.2007.11.007. [DOI] [PubMed] [Google Scholar]
- Robin A. Encyclopedia of Marine Gastropods. Hackenheim, Germany: Xenophora and Conchbooks; 2008. p. 448. [Google Scholar]
- Springsteen FJ, Leobrera FM. Shells of the Philippines. Manila: Carfel Seashell Museum; 1986. pp. 265–266. [Google Scholar]
- Taylor JD, Kantor Y, Sysoev AV. Bulletin. 59. Natural History Museum; London: 1993. Foregut anatomy, feeding mechanisms, relationships and classification of the Conoidea (=Toxoglossa) (Gastropoda) pp. 125–170. [Google Scholar]
- Tucker JK, Tenorio MJ. Conchbooks. 2009. Systematic classification of recent and fossil Conoidean gastropods; p. 296. [Google Scholar]
- Vera-Peláez JL, Vega-Luz R, Lozano-Francisco MC. Five new species of the genus Turris Röding,1798 (Gastropoda; Turridae; Turrinae) of the Philippines and one new species of the Southern Indo-Pacific. Makakos: Monografía. 2000;2:1–29. [Google Scholar]