

Abstract
Aberrant neuronal activity in injured peripheral nerves is believed to be an important factor in the development of neuropathic pain. Pharmacological blockade of that activity has been shown to mitigate the onset of associated molecular events in the nervous system. However, results in preventing onset of pain behaviors by providing prolonged nerve blockade have been mixed. Furthermore, the experimental techniques used to date to provide that blockade were limited in clinical potential in that they would require surgical implantation. To address these issues, we have used liposomes (SDLs) containing saxitoxin (STX), a site 1 sodium channel blocker, and the glucocorticoid agonist dexamethasone to provide nerve blocks lasting ∼1 wk from a single injection. This formulation is easily injected percutaneously. Animals undergoing spared nerve injury (SNI) developed mechanical allodynia in 1 wk; nerve blockade with a single dose of SDLs (duration of block 6.9 ± 1.2 d) delayed the onset of allodynia by 2 d. Treatment with three sequential SDL injections resulting in a nerve block duration of 18.1 ± 3.4 d delayed the onset of allodynia by 1 mo. This very prolonged blockade decreased activation of astrocytes in the lumbar dorsal horn of the spinal cord due to SNI. Changes in expression of injury-related genes due to SNI in the dorsal root ganglia were not affected by SDLs. These findings suggest that formulations of this kind, which could be easy to apply clinically, can mitigate the development of neuropathic pain.
Keywords: conduction block, drug delivery, hyperalgesia, prolonged anesthesia
Neuropathic pain is caused by a primary lesion or dysfunction in the nervous system (1); nerve trauma is a common cause (in addition to many medical conditions) (2). The prevalence of chronic neuropathic pain cases is ∼3.75 million in the United States alone (3), and the management of such cases is a significant burden in health care spending (4). Despite availability of several pharmacological therapies, a significant number of patients have unsatisfactory pain control, and/or experience undesirable side effects from existing pain medication, making it imperative to examine new therapeutic strategies to prevent chronic neuropathic pain.
One major approach that has been investigated is the blockade of the enhanced neuronal activity from injured primary afferents that contributes to the development of neuropathic pain from nerve injury (5–7). It has been argued that excitatory neurotransmitters such as glutamate, released in response to injury-induced aberrant neuronal impulses, lead to calcium-mediated excitotoxicity and subsequent cellular injury at the level of the dorsal root ganglion (DRG) and spinal cord (8–10). It is clear that the inhibition of injury-induced neuronal discharges can mitigate the subsequent development of pain behavior (11, 12).
However, research in this area has encountered two major difficulties. First, the methods developed to provide prolonged blockade to date—a continuous infusion of tetrodotoxin from an osmotic pump (13), a depot of bupivacaine free base (14), and a bupivacaine–dexamethasone microspheres contained in fibrin glue, loaded within a silicone tube (15)—would be difficult to implement clinically, as they would require surgical implantation of a device on the nerve. Second, attempts to block neuropathic pain by using those devices to inhibit injury-induced neuronal discharges for ∼1 wk have had mixed results, even though such blockade resulted in a mitigation, or at least a delaying, of molecular events associated with the development of neuropathic pain (13, 16, 17). A continuous infusion of tetrodotoxin from an osmotic pump or a depot of bupivacaine free base were able to inhibit the onset of pain behaviors (13) after nerve injury. In contrast, bupivacaine–dexamethasone microspheres contained in fibrin glue, loaded within a silicone tube, had no effect on the development of pain behaviors (15). There is no obvious explanation for these discrepancies, but the reports used different drugs [with different effects on ion channels (13, 18, 19) and on local tissue reaction (20)], and sustained release formulations (also with potentially different effects on tissue reaction; ref. 21).
Here, we have used liposomes containing the site 1 sodium channel blocker saxitoxin (STX) with dexamethasone (SDLs) (22) as the means of providing prolonged nerve blockade to address the crucially important issue of the ability of prolonged duration local anesthesia to mitigate the onset of neuropathic pain. STX is a potent local anesthetic that acts at site 1 of the voltage-sensitive sodium channel (23); dexamethasone has been shown to prolong the duration of nerve blockade in several sustained release formulations (24–27), although the mechanism is not understood. SDLs are easy to administer percutaneously, and single injections produce nerve blocks lasting ∼1 wk in the rat (26). The SDLs are advantageous in that STX (and the SDLs) does not cause myo- or neurotoxicity (26). This lack of toxicity is in contrast to the encapsulated amino-amide local anesthetics used in previous attempts to mitigate neuropathic pain (15); those compounds are myo- and neurotoxic as free drugs (28, 29) and, when encapsulated, potentially cause local tissue injury (21, 30, 31). To gauge the impact of prolonged blockade with SDLs on neuropathic pain, we have studied their effect on the development of allodynia, and on biological events in the dorsal root ganglia and spinal cord, in the spared nerve injury (SNI) model in the rat.
Results
Sciatic Nerve Blockade with SDLs.
Multilamellar/multivesicular SDLs 5.4 ± 1.4 μm in diameter were prepared using the thin-lipid film technique as reported (26) (Fig. S1 A and B). Injection of 0.3 mL of the SDL formulation at the sciatic nerve increased hind-paw thermal latency in the injected limb for 5–6 d (Fig. S1C). Liposomes made in exactly the same way with (dexamethasone and citrate buffer, pH 4.5, but no STX) had no effect on withdrawal latency.
Effect of SDLs on Nerve Conduction.
Nerve conduction studies were performed in animals with intact sciatic nerves to determine the physical extent of SDL-induced conduction block along the sciatic nerve (Fig. 1A), specifically to confirm that the SDLs remained at the site of injection vs. spreading along the length of the nerve.
Fig. 1.

SDLs produce focal nerve conduction deficits in the sciatic nerve. (A) Schematic showing the anatomy of the sciatic nerve in relation to the site of SDL injection and stimulating/recording sites for nerve conduction studies. (B) Evoked compound muscle action potential measurements after simulation of the sciatic nerve at the ankle (black) and at the hip (red) were recorded from the plantaris muscle at day 4 in the SDL-treated (Right) and untreated/pristine (Left) hind limbs. M and H wave latency and M wave amplitudes are shown in the table. Data are means ± SD, n = 5.
In the pristine hind limb (no SDL), (Fig. 1B) electrical stimulation of the sciatic nerve at the hip (Fig. 1A) evoked a compound muscle action potential that traveled anterograde and was recorded at the plantaris muscle (M wave, Fig. 1B). Stimulation at the hip also evoked retrograde conduction of the electrical impulse to the spinal cord, which was then transmitted back to the periphery monosynaptically via the sciatic nerve where it was measured at the plantaris muscle (H wave, Fig. 1B). Similarly, stimulation of the tibial nerve at the ankle evoked robust M and H waves, at different latencies compared with M and H wave latencies evoked by stimulation at the hip (Fig. 1B, table).
On day 4 after injection with SDL, electrical stimulation at the hip (proximal to the site of SDL injection), failed to generate the M and H wave in the plantaris muscle, suggesting a complete block of the proximal sciatic nerve segment. Stimulation of the tibial nerve at the ankle evoked a robust M wave, but not the H wave, suggesting normal nerve function in the tibial segment distal to the SDLs, but confirming block in the proximal sciatic nerve segment. Amplitude of the M wave elicited by ankle stimulation in the SDL-treated limb was similar (P > 0.05, n = 5) to the M wave elicited from the pristine limb, confirming normal axonal function in the distal tibial segment (Fig. 1B, table).
The localized distribution of SDL liposomes was also confirmed by postmortem dissection of the injection sites, where localized liposomal residue was found at a segment of the sciatic nerve near the greater trochanter even 30 d after administration.
Effect of Prolonged Nerve Block on Onset of SNI-Induced Tactile Hypersensitivity.
To determine the effect of extended nerve blockade on the onset of neuropathic pain, SNI rats received nerve block injections with SDLs in the left hind-limb, ipsilateral and rostral to the nerve injury. Rats were tested for tactile sensitivity, an indicator of neuropathic pain, over a period of 9 wk. Thermal withdrawal latencies were used to monitor SDL-induced nerve conduction block. Untreated SNI animals demonstrated a thermal withdrawal latency of under 3 s, i.e., no nerve block.
Effect of 7-d Nerve Blockade on SNI-Induced Tactile Hypersensitivity.
Administration of a single dose of SDLs immediately after nerve injury achieved nerve block duration of 6.9 ± 1.2 d, as assessed by thermal latency testing (Fig. 2A). This duration was similar to those obtained in other studies on the effect of prolonged duration local anesthetics (15, 16) on neuropathic pain. Hind-limb mechanical withdrawal thresholds were determined to assess the effect of SDLs on SNI-induced tactile sensitivity (Fig. 2 B and C) in the ipsilateral (nerve-injured) and the uninjured contralateral limb. (In these and all experiments, the SNI and the SDLs were in the same extremity.) In SNI animals, mechanical withdrawal thresholds dropped progressively following injury. During (and because of) nerve blockade from a single dose of SDLs, mechanical withdrawal thresholds remained elevated in the injected extremity in SNI rats (P < 0.05, n = 5) compared with animals that did not receive SDL (Fig. 2B). That difference resolved (P > 0.05) 2 d after recovery from block (i.e., there was a delay in the onset of hypersensitivity by 2 d). Mechanical withdrawal thresholds in the leg contralateral from the SNI decreased following SNI; this occurred irrespective of whether SDL were injected (Fig. 2C; P > 0.05 for the comparison of SNI vs. SNI with SDL).
Fig. 2.

Effect of 7-d nerve blockade from SDLs on the onset of tactile hypersensitivity in the SNI model. (A) Thermal latency measurements in the ipsilateral limb. Mechanical withdrawal threshold measurements were obtained from the ipsilateral (B) and contralateral (C) hind limbs. Animals received SDLs on day 0. Shaded areas in B and C represent duration of nerve block (see A). Data are mean ± SD; *P < 0.05, compared with untreated SNI animals, n = 5 rats per group.
A single injection of liposomes containing only dexamethasone (and citrate buffer without STX) had no effect on SNI-induced decrease in mechanical withdrawal threshold in the injured leg.
Effect of 18-d Nerve Blockade on SNI-Induced Paw Hypersensitivity in the Ipsilateral Limb.
The 2-d delay in the onset of tactile hypersensitivity following 7 d of nerve blockade prompted an examination of the effect of longer durations of block. To attain very prolonged continuous nerve block, SNI rats received injections with SDLs on day 0 (immediately after injury), and on days 5 and 12 (Fig. 3A). Thermal nociceptive block in the repeatedly injected extremities lasted 18.1 ± 3.4 d (Fig. 3A). In the absence of SDL injection, SNI rats exhibited decreased mechanical withdrawal thresholds in the injured limb compared with preinjury levels beginning at week 2 (P < 0.05, n = 5), and continuing till week 9 (Fig. 3B, P < 0.05, n = 5, one-way ANOVA). In contrast, mechanical withdrawal thresholds in the injured limb in SNI animals treated with SDLs were similar (P > 0.05, n = 5) to preinjury threshold values until week 6 (P > 0.05, n = 5), i.e., more than 3 wk after nerve block from the SDLs had worn off. (During nerve blockade, the elevated mechanical withdrawal thresholds were likely due to effects of the SLDs, rather than reflecting a lack of hypersensitivity.) Furthermore, mechanical withdrawal thresholds were significantly higher in SDL-injected than noninjected SNI animals during weeks 2–6 (P > 0.05, n = 5), again long after nerve block from SDLs had worn off. After week 6, mechanical withdrawal thresholds from the nerve-injured extremity of SDL-treated rats were comparable to those obtained from untreated SNI animals (P > 0.05, n = 5; Fig. 3B).
Fig. 3.

Effect of 18-d nerve blockade from SDLs on the onset of tactile hypersensitivity in the SNI model. (A) Thermal latency measurements in the ipsilateral limb. Mechanical withdrawal threshold measurements in the ipsilateral (B) and contralateral (C) hind limbs. Animals received SDLs on days 0, 5, and 12 (arrows in A). Shaded areas in B and C represent duration of nerve block (see A). Note that the time scale in A is different from those in panels B and C; dotted lines bridging A and B show the correspondence of the time frames. Data are means ± SD; *, †P < 0.05, n = 5 rats per group. (*, comparison between SNI- and SDL-treated SNI groups; †, comparison between SNI or SDL + SNI and respective preinjury levels at day 0.)
Effect of 18-d Nerve Blockade on SNI-Induced Tactile Hypersensitivity in the Contralateral Limb.
Mechanical withdrawal thresholds were decreased from week 2 to week 7 in the uninjured (contralateral) limb of SNI animals (P < 0.05 compared with preinjury threshold values; Fig. 3C). In SNI animals administered SDLs, withdrawal thresholds in the uninjured limb were unchanged at all time points tested (P < 0.05, compared with untreated animals at weeks 2–7).
Effect of 18-d Nerve Block on SNI-Induced Astrocyte Response in the Lumbar Spinal Cord.
Maintenance of neuropathic pain is facilitated by the activation of astrocytes in the central nervous system. To determine if prolonged nerve block (18.1 ± 3.4 d in these experiments) can alter astrocyte activation, we analyzed the expression activation of glial fibrillary acidic protein (GFAP, a marker for astrocytes) in the pain signal processing area of the dorsal horn of the lumbar spinal cord, 5 and 60 d after SNI. Those time points were chosen to represent acute and chronic responses to nerve injury respectively. GFAP-positive glial cells from the dorsal horn of uninjured naïve rats appeared star shaped with thin processes (Fig. 4A Inset), whereas those from dorsal horns ipsilateral to nerve injury, 5 d after SNI exhibited a stubby morphology with thick processes suggestive of astrocyte activation (32) (Fig. 4B Inset). At the same time point, astrocyte morphology in SNI animals treated with SDLs was similar to that from uninjured naïve animals (Fig. 4C Inset).
Fig. 4.

Prolonged duration nerve block attenuates SNI-induced astrocyte (GFAP) activation. Representative images of GFAP-positive cells (white arrows) in the L4, L5 dorsal horn of spinal cord sections from uninjured (A), SNI-treated (B), and SNI + SDL-treated (C) animals. (Insets) Representative pictures depicting astrocyte morphology in each group. Tissues were harvested 5 d after nerve injury and/or first SDL administration. Quantitative analysis of GFAP staining in dorsal horn ipsilateral to injury (D) and contralateral to injury (E) is expressed as the percentage of area that is GFAP positive. Data are means ± SD, *P < 0.05, n = 5 rats per group.
The average total area covered by GFAP-positive cells (33) was increased at 5 and 60 d in the ipsilateral dorsal horn after SNI, compared with uninjured naïve rats (Fig. 4D, P < 0.05, n = 5). In contrast, SDL-treated SNI animals at days 5 and 60 exhibited ipsilateral GFAP staining that was similar in extent (P > 0.05) to the saline-treated naïve group, but significantly lower (P < 0.05) than that observed in the untreated SNI group (Fig. 4D).
GFAP staining in the contralateral dorsal horn of SNI animals that did not receive SDLs was increased (P < 0.05) compared with uninjured animals at day 5 but not at day 60 (Fig. 4E). There was no such increase in the GFAP-positive area in SDL-treated animals.
Effect of 18-d Nerve Block on Nerve Injury-Induced Gene Expression Changes in the DRG.
In addition to astrocyte activation in the CNS, gene expression changes in the cell bodies of peripheral afferent neurons are consistently observed in nerve-injured neuropathic pain models (34). To assess whether prolonged conduction block (18.1 ± 3.4 d in these experiments) of afferent neurons can alter injury-dependent changes in gene expression, we used a customized RT2 Profiler PCR array to profile genes whose expression is known to be altered in the DRG of SNI animals (35, 36), with or without SDL treatment at day 5 and 60. Gene expression in all experimental groups was studied as a multiple of the expression in uninjured, unblocked animals.
SDL treatment alone (in uninjured animals) did not induce an increase in the expression of any genes examined in ipsilateral DRGs on day 5 (Table 1). SNI increased the expression of several injury-related genes in ipsilateral DRG’s at 5 and 60 d, whereas treatment with SDLs did not prevent SNI-induced increase in expression of most genes at either time point.
Table 1.
Effect of 18-d nerve block on the expression of selected genes related to nerve injury in the L4, L5 DRG
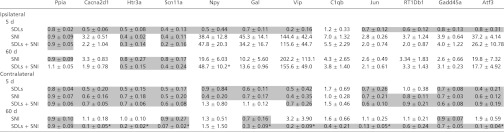 |
Data are means ± SD of mRNA fold changes over expression levels in pristine animals (n = 5 rats per group). *P < 0.05 in comparison of SDL-treated SNI group (SDLs + SNI) and untreated SNI group by one-way ANOVA with Bonferroni post hoc comparison. Decreases in mRNA expression levels (numbers <1, being the inverse of the fold decrease) are shaded, whereas increases are unshaded. Gene abbreviations: Ppia (control), Cyclophilin A; Cacna2d1, Calcium channel α2/δ1; Htr3a, 5-hyrdoxytryptamine receptor 3a; Scn11a, Sodium channel IX α; Npy, Neuropeptide Y; Gal, Galanin prepropeptide; Vip, Vasoactive intestinal peptide; C1qb, Complement component 1q βchain; Jun, Jun oncogene; RT1Db1, RT1 class II, locus Db1; Gadd45a, Growth arrest and DNA damage-inducible α; Atf3, activating transcription factor 3.
Contralateral DRGs harvested from SNI animals showed no changes in gene expression after 5 and 60 d. However, at 60 d, DRGs harvested from SDL-treated SNI rats exhibited down-regulation of many genes tested (Table 1) (P < 0.05) compared with SNI animals that did not receive SDL.
Discussion
Aberrant spontaneous activity in the sensory afferents is consistently observed in animal models of painful neuropathy (37, 38). Peripheral nerve associated injury-induced discharges are thought to be responsible for irreversible changes in the central nervous system (8), leading to maintenance of chronic pain. Previous efforts to mitigate the development of neuropathic pain by prolonged neural blockade have had conflicting results. Bupivacaine–dexamethasone particles contained within a silicone tube provided nerve blockade lasting 8 d, but did not impact the development of allodynia or hyperalgesia in SNI (15) even though the same formulation was shown to prevent the rise in p38 mitogen-activated protein kinase seen in spinal microglia after SNI (17) and prevented the induction of apoptosis in the dorsal horn (16). In contrast, mitigation of pain behavior and reduced injury-induced glial cell activation in the spinal cord was seen following ∼1-wk block with bupivacaine free base powder or a tetrodotoxin infusion (13, 14). Different parts of our results were in agreement with both reports. Blockade with a single dose of SDL, resulting in 7-d block, did not prevent allodynia. However, 18-d blockade with repeated dosing resulted in a 4-wk delay in the onset of allodynia. Furthermore, spinal astrocyte activation, a facilitator of central sensitization in chronic neuropathic pain models (34, 39), was markedly decreased in SDL-treated SNI animals; this reduction was still in effect 60 d after SNI, 42 d after nerve block wore off. These results are consistent with the view that very prolonged nerve blockade could mitigate the development of neuropathic pain, and that such could be provided by an easily injectable sustained release system.
There has been interest for several decades in using drug delivery technologies to affect neuronal function locally, in the central (40) and peripheral nervous systems (31), particularly in the development of prolonged duration local anesthetics. A wide range of devices have been used for the latter application (24, 26, 27, 30, 41–45). Few have achieved blocks lasting even a few days (24, 25). Those that did often caused severe difficulties with local inflammation and/or tissue toxicity (21), particularly myo- and neurotoxicity (21, 25). SDLs produce prolonged nerve blockade (∼1 wk from a single injection) with only mild inflammation and without evidence of myo- or neurotoxicity (26).
One possible explanation for the ability of SDLs to mitigate the development of mechanical allodynia where the bupivacaine–dexamethasone particles had failed is that the latter were neurotoxic and injured the nerves during blockade. That explanation is not consistent with the reports that nerve block lasting ∼1 wk did not mitigate the development of allodynia whether achieved by the bupivacaine–dexamethasone particles (15) or the SDLs, whereas bupivacaine free base powder (presumably more tissue-toxic) was able to mitigate pain behavior (13). Duration of block seems to be a key factor, as seen by the fact that repeated administration of SDL did impact the onset of allodynia. The repeated administration of SDL may be blocking injury-induced late-phase electrical discharges (46) that are important for pain maintenance. However, duration may not be the only factor. Whereas 1 wk of nerve block with SDLs did not mitigate pain behavior, a similar duration of block with tetrodotoxin or bupivacaine free base has been reported to do so (13). The question arises why the SLDs had less effect on the development of allodynia than was reported with some other approaches (13). One possibility is that nerve blockade may not have been as consistently dense with the SDLs as was in some other reports (13). It is also possible that although dexamethasone prolongs nerve blockade, it also exacerbates the development of neuropathic pain; this would also explain why bupivacaine free base prevented neuropathic pain (13, 14) but bupivacaine–dexamethasone particles did not (15).
Some studies demonstrate the consistent onset of contralateral hyperalgesia in nerve-injured animal models of neuropathic pain (47–51), whereas others report a mild or no hyperalgesic response (52, 53). Here, we observed transient contralateral hyperalgesia in SNI animals that was both slower in onset and lower in intensity compared with its ipsilateral counterpart. At the spinal cord level, astrocyte activation was increased in the contralateral dorsal horn, but it was milder compared with activation in the ipsilateral dorsal horn. The mechanism for contralateral pain is largely unknown, but altered spinal processing of sensory signals has been proposed (54). Mirror image pain, a well documented feature of human causalgia, has been equated with experimental contralateral hyperalgesia (49); the findings of this study raise the possibility of using prolonged ipsilateral nerve conduction block in preventing mirrored pain syndromes.
Nerve injury-induced gene expression changes in the DRG cell soma have been well documented, with the majority of injury-related genes changing expression 3 d after nerve injury (35) and remaining altered even after 13 d (36). In the present study, both 7- and 18-d nerve block failed to prevent SNI-induced gene expression changes in the ipsilateral L4, L5 DRG, even though nerve block mitigated the development of allodynia. These findings raise the possibility that activity-dependent changes in gene expression are not by themselves sufficient to cause neuropathic pain. In the contralateral DRGs, prolonged nerve block induced a significant decrease in expression in 7 of 11 genes, 2 mo after nerve injury. The significance of that finding is unclear; as noted above, contralateral pain following nerve injury is not well understood (nor universally reported) at this time.
Ultra-long-duration nerve block obtained by repeated administration of SDLs delays nerve injury-induced tactile hypersensitivity and attenuates the central astrocyte activation response. There would appear to be many important factors with potential impact on the effectiveness of formulations in mitigating pain behaviors, including type of drug, type of formulation, and duration and intensity of nerve blockade. SDLs provide an approach that is potentially clinically practical for preventing neuropathic pain conditions.
Materials and Methods
Liposome Preparation.
Liposomes were produced by the thin lipid film technique, using 1,2-distearoyl-sn-glycero-3 phosphocholine (DSPC) and 1,2-distearoyl-sn-glycero-3-phoshatidylglycerol (DSPG) (Genzyme) as described (26). Size of liposomes was determined using a multisizer 3 Coulter Counter (Beckman Coulter). Intraliposome STX content was determined after separating the lipid fraction using the Bligh and Dryer method (55). Determination of STX content was based on a previously published method by Bates et al. (56). STX was received as a generous gift from Sherwood Hall (Food and Drug Administration, College Park, MD).
Nerve Conduction Studies.
For assessing the effect of SDLs on nerve function, we measured evoked compound muscle action potential latencies and amplitudes from the sciatic nerve (57) at the peak of nerve block (day 4 after SDL treatment, as characterized by the thermal paw withdrawal test). Rats were weighed daily and their body weight monitored (Fig. S2).
Detailed protocols for nerve conduction studies, immunohistochemistry, PCR array, spared nerve injury, behavioral tests for nociception, and histological and statistical methods are described in SI Materials and Methods.
Supplementary Material
Footnotes
The authors declare no conflict of interest.
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1214634109/-/DCSupplemental.
References
- 1.Classification of chronic pain. Descriptions of chronic pain syndromes and definitions of pain terms. Prepared by the International Association for the Study of Pain, Subcommittee on Taxonomy. Pain Suppl. 1986;3:S1–S226. [PubMed] [Google Scholar]
- 2.Zimmermann M. Pathobiology of neuropathic pain. Eur J Pharmacol. 2001;429:23–37. doi: 10.1016/s0014-2999(01)01303-6. [DOI] [PubMed] [Google Scholar]
- 3.Harden RN. Chronic neuropathic pain. Mechanisms, diagnosis, and treatment. Neurologist. 2005;11:111–122. doi: 10.1097/01.nrl.0000155180.60057.8e. [DOI] [PubMed] [Google Scholar]
- 4.Turk DC. Clinical effectiveness and cost-effectiveness of treatments for patients with chronic pain. Clin J Pain. 2002;18:355–365. doi: 10.1097/00002508-200211000-00003. [DOI] [PubMed] [Google Scholar]
- 5.Wu G, et al. Early onset of spontaneous activity in uninjured C-fiber nociceptors after injury to neighboring nerve fibers. J Neurosci. 2001;21:RC140. doi: 10.1523/JNEUROSCI.21-08-j0002.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Govrin-Lippmann R, Devor M. Ongoing activity in severed nerves: Source and variation with time. Brain Res. 1978;159:406–410. doi: 10.1016/0006-8993(78)90548-6. [DOI] [PubMed] [Google Scholar]
- 7.Wall PD, Devor M. Sensory afferent impulses originate from dorsal root ganglia as well as from the periphery in normal and nerve injured rats. Pain. 1983;17:321–339. doi: 10.1016/0304-3959(83)90164-1. [DOI] [PubMed] [Google Scholar]
- 8.Coggeshall RE, Lekan HA, White FA, Woolf CJ. A-fiber sensory input induces neuronal cell death in the dorsal horn of the adult rat spinal cord. J Comp Neurol. 2001;435:276–282. doi: 10.1002/cne.1029. [DOI] [PubMed] [Google Scholar]
- 9.Choi DW. Calcium and excitotoxic neuronal injury. Ann N Y Acad Sci. 1994;747:162–171. doi: 10.1111/j.1749-6632.1994.tb44407.x. [DOI] [PubMed] [Google Scholar]
- 10.Woolf CJ, Salter MW. Neuronal plasticity: Increasing the gain in pain. Science. 2000;288:1765–1769. doi: 10.1126/science.288.5472.1765. [DOI] [PubMed] [Google Scholar]
- 11.Dougherty PM, Garrison CJ, Carlton SM. Differential influence of local anesthetic upon two models of experimentally induced peripheral mononeuropathy in the rat. Brain Res. 1992;570:109–115. doi: 10.1016/0006-8993(92)90570-y. [DOI] [PubMed] [Google Scholar]
- 12.Sotgiu ML, Castagna A, Lacerenza M, Marchettini P. Pre-injury lidocaine treatment prevents thermal hyperalgesia and cutaneous thermal abnormalities in a rat model of peripheral neuropathy. Pain. 1995;61:3–10. doi: 10.1016/0304-3959(94)00120-4. [DOI] [PubMed] [Google Scholar]
- 13.Xie W, Strong JA, Zhang JM. Early blockade of injured primary sensory afferents reduces glial cell activation in two rat neuropathic pain models. Neuroscience. 2009;160:847–857. doi: 10.1016/j.neuroscience.2009.03.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Xie W, Strong JA, Meij JTA, Zhang J-M, Yu L. Neuropathic pain: Early spontaneous afferent activity is the trigger. Pain. 2005;116:243–256. doi: 10.1016/j.pain.2005.04.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Suter MR, et al. Development of neuropathic pain in the rat spared nerve injury model is not prevented by a peripheral nerve block. Anesthesiology. 2003;99:1402–1408. doi: 10.1097/00000542-200312000-00025. [DOI] [PubMed] [Google Scholar]
- 16.Scholz J, et al. Blocking caspase activity prevents transsynaptic neuronal apoptosis and the loss of inhibition in lamina II of the dorsal horn after peripheral nerve injury. J Neurosci. 2005;25:7317–7323. doi: 10.1523/JNEUROSCI.1526-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Wen Y-R, et al. Nerve conduction blockade in the sciatic nerve prevents but does not reverse the activation of p38 mitogen-activated protein kinase in spinal microglia in the rat spared nerve injury model. Anesthesiology. 2007;107:312–321. doi: 10.1097/01.anes.0000270759.11086.e7. [DOI] [PubMed] [Google Scholar]
- 18.Seltzer Z, Cohn S, Ginzburg R, Beilin B. Modulation of neuropathic pain behavior in rats by spinal disinhibition and NMDA receptor blockade of injury discharge. Pain. 1991;45:69–75. doi: 10.1016/0304-3959(91)90166-U. [DOI] [PubMed] [Google Scholar]
- 19.Yoon YW, Na HS, Chung JM. Contributions of injured and intact afferents to neuropathic pain in an experimental rat model. Pain. 1996;64:27–36. doi: 10.1016/0304-3959(95)00096-8. [DOI] [PubMed] [Google Scholar]
- 20.Padera RF, Tse JY, Bellas E, Kohane DS. Tetrodotoxin for prolonged local anesthesia with minimal myotoxicity. Muscle Nerve. 2006;34:747–753. doi: 10.1002/mus.20618. [DOI] [PubMed] [Google Scholar]
- 21.Kohane DS, Langer R. Biocompatibility and drug delivery systems. Chem Sci. 2010;1:441–446. [Google Scholar]
- 22.Kohane DS, et al. The local anesthetic properties and toxicity of saxitonin homologues for rat sciatic nerve block in vivo. Reg Anesth Pain Med. 2000;25:52–59. doi: 10.1016/s1098-7339(00)80011-5. [DOI] [PubMed] [Google Scholar]
- 23.Zhang M-M, et al. Cooccupancy of the outer vestibule of voltage-gated sodium channels by micro-conotoxin KIIIA and saxitoxin or tetrodotoxin. J Neurophysiol. 2010;104:88–97. doi: 10.1152/jn.00145.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Curley J, et al. Prolonged regional nerve blockade. Injectable biodegradable bupivacaine/polyester microspheres. Anesthesiology. 1996;84:1401–1410. doi: 10.1097/00000542-199606000-00017. [DOI] [PubMed] [Google Scholar]
- 25.Kohane DS, et al. Prolonged duration local anesthesia from tetrodotoxin-enhanced local anesthetic microspheres. Pain. 2003;104:415–421. doi: 10.1016/s0304-3959(03)00049-6. [DOI] [PubMed] [Google Scholar]
- 26.Epstein-Barash H, et al. Prolonged duration local anesthesia with minimal toxicity. Proc Natl Acad Sci USA. 2009;106:7125–7130. doi: 10.1073/pnas.0900598106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Colombo G, Padera R, Langer R, Kohane DS. Prolonged duration local anesthesia with lipid-protein-sugar particles containing bupivacaine and dexamethasone. J Biomed Mater Res A. 2005;75:458–464. doi: 10.1002/jbm.a.30443. [DOI] [PubMed] [Google Scholar]
- 28.Gold MS, Reichling DB, Hampl KF, Drasner K, Levine JD. Lidocaine toxicity in primary afferent neurons from the rat. J Pharmacol Exp Ther. 1998;285:413–421. [PubMed] [Google Scholar]
- 29.Radwan IAM, Saito S, Goto F. The neurotoxicity of local anesthetics on growing neurons: A comparative study of lidocaine, bupivacaine, mepivacaine, and ropivacaine. Anesth Analg. 2002;94:319–324. doi: 10.1097/00000539-200202000-00016. [DOI] [PubMed] [Google Scholar]
- 30.Padera R, Bellas E, Tse JY, Hao D, Kohane DS. Local myotoxicity from sustained release of bupivacaine from microparticles. Anesthesiology. 2008;108:921–928. doi: 10.1097/ALN.0b013e31816c8a48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Kohane DS, et al. Biocompatibility of lipid-protein-sugar particles containing bupivacaine in the epineurium. J Biomed Mater Res. 2002;59:450–459. doi: 10.1002/jbm.1261. [DOI] [PubMed] [Google Scholar]
- 32.Wieseler-Frank J, Maier SF, Watkins LR. Glial activation and pathological pain. Neurochem Int. 2004;45:389–395. doi: 10.1016/j.neuint.2003.09.009. [DOI] [PubMed] [Google Scholar]
- 33.Blackbeard J, et al. Quantification of the rat spinal microglial response to peripheral nerve injury as revealed by immunohistochemical image analysis and flow cytometry. J Neurosci Methods. 2007;164:207–217. doi: 10.1016/j.jneumeth.2007.04.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Guo W, et al. Glial-cytokine-neuronal interactions underlying the mechanisms of persistent pain. J Neurosci. 2007;27:6006–6018. doi: 10.1523/JNEUROSCI.0176-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Costigan M, et al. Replicate high-density rat genome oligonucleotide microarrays reveal hundreds of regulated genes in the dorsal root ganglion after peripheral nerve injury. BMC Neurosci. 2002;3:16. doi: 10.1186/1471-2202-3-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Wang H, et al. Chronic neuropathic pain is accompanied by global changes in gene expression and shares pathobiology with neurodegenerative diseases. Neuroscience. 2002;114:529–546. doi: 10.1016/s0306-4522(02)00341-x. [DOI] [PubMed] [Google Scholar]
- 37.Roza C, Laird JMA, Souslova V, Wood JN, Cervero F. The tetrodotoxin-resistant Na+ channel Nav1.8 is essential for the expression of spontaneous activity in damaged sensory axons of mice. J Physiol. 2003;550:921–926. doi: 10.1113/jphysiol.2003.046110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Han HC, Lee DH, Chung JM. Characteristics of ectopic discharges in a rat neuropathic pain model. Pain. 2000;84:253–261. doi: 10.1016/s0304-3959(99)00219-5. [DOI] [PubMed] [Google Scholar]
- 39.Gao YJ, Ji RR. Chemokines, neuronal-glial interactions, and central processing of neuropathic pain. Pharmacol Ther. 2010;126:56–68. doi: 10.1016/j.pharmthera.2010.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Tamargo RJ, Epstein JI, Reinhard CS, Chasin M, Brem H. Brain biocompatibility of a biodegradable, controlled-release polymer in rats. J Biomed Mater Res. 1989;23:253–266. doi: 10.1002/jbm.820230209. [DOI] [PubMed] [Google Scholar]
- 41.Kopacz DJ, et al. The dose response and effects of dexamethasone on bupivacaine microcapsules for intercostal blockade (T9 to T11) in healthy volunteers. Anesth Analg. 2003;96:576–582. doi: 10.1097/00000539-200302000-00050. [DOI] [PubMed] [Google Scholar]
- 42.Kohane DS. Microparticles and nanoparticles for drug delivery. Biotechnol Bioeng. 2007;96:203–209. doi: 10.1002/bit.21301. [DOI] [PubMed] [Google Scholar]
- 43.Kohane DS, Lipp M, Kinney RC, Lotan N, Langer R. Sciatic nerve blockade with lipid-protein-sugar particles containing bupivacaine. Pharm Res. 2000;17:1243–1249. doi: 10.1023/a:1026470831256. [DOI] [PubMed] [Google Scholar]
- 44.Rose JS, Neal JM, Kopacz DJ. Extended-duration analgesia: Update on microspheres and liposomes. Reg Anesth Pain Med. 2005;30:275–285. doi: 10.1016/j.rapm.2005.01.004. [DOI] [PubMed] [Google Scholar]
- 45.Kuzma PJ, Kline MD, Calkins MD, Staats PS. Progress in the development of ultra-long-acting local anesthetics. Reg Anesth. 1997;22:543–551. [PubMed] [Google Scholar]
- 46.Suzuki R, Dickenson AH. Differential pharmacological modulation of the spontaneous stimulus-independent activity in the rat spinal cord following peripheral nerve injury. Exp Neurol. 2006;198:72–80. doi: 10.1016/j.expneurol.2005.10.032. [DOI] [PubMed] [Google Scholar]
- 47.Erichsen HK, Blackburn-Munro G. Pharmacological characterisation of the spared nerve injury model of neuropathic pain. Pain. 2002;98:151–161. doi: 10.1016/s0304-3959(02)00039-8. [DOI] [PubMed] [Google Scholar]
- 48.Kim SH, Chung JM. An experimental model for peripheral neuropathy produced by segmental spinal nerve ligation in the rat. Pain. 1992;50:355–363. doi: 10.1016/0304-3959(92)90041-9. [DOI] [PubMed] [Google Scholar]
- 49.Seltzer Z, Dubner R, Shir Y. A novel behavioral model of neuropathic pain disorders produced in rats by partial sciatic nerve injury. Pain. 1990;43:205–218. doi: 10.1016/0304-3959(90)91074-S. [DOI] [PubMed] [Google Scholar]
- 50.Takaishi K, Eisele JH, Jr, Carstens E. Behavioral and electrophysiological assessment of hyperalgesia and changes in dorsal horn responses following partial sciatic nerve ligation in rats. Pain. 1996;66:297–306. doi: 10.1016/0304-3959(96)03023-0. [DOI] [PubMed] [Google Scholar]
- 51.Yasuda T, Miki S, Yoshinaga N, Senba E. Effects of amitriptyline and gabapentin on bilateral hyperalgesia observed in an animal model of unilateral axotomy. Pain. 2005;115:161–170. doi: 10.1016/j.pain.2005.02.026. [DOI] [PubMed] [Google Scholar]
- 52.Decosterd I, Woolf CJ. Spared nerve injury: An animal model of persistent peripheral neuropathic pain. Pain. 2000;87:149–158. doi: 10.1016/S0304-3959(00)00276-1. [DOI] [PubMed] [Google Scholar]
- 53.Broom DC, et al. Cyclooxygenase 2 expression in the spared nerve injury model of neuropathic pain. Neuroscience. 2004;124:891–900. doi: 10.1016/j.neuroscience.2004.01.003. [DOI] [PubMed] [Google Scholar]
- 54.Koltzenburg M, Wall PD, McMahon SB. Does the right side know what the left is doing? Trends Neurosci. 1999;22:122–127. doi: 10.1016/s0166-2236(98)01302-2. [DOI] [PubMed] [Google Scholar]
- 55.Bligh EG, Dyer WJ. A rapid method of total lipid extraction and purification. Can J Biochem Physiol. 1959;37:911–917. doi: 10.1139/o59-099. [DOI] [PubMed] [Google Scholar]
- 56.Bates HA, Kostriken R, Rapoport H. A chemical assay for saxitoxin. Improvements and modifications. J Agric Food Chem. 1978;26:252–254. doi: 10.1021/jf60215a060. [DOI] [PubMed] [Google Scholar]
- 57.Shankarappa SA, Piedras-Rentería ES, Stubbs EB., Jr Forced-exercise delays neuropathic pain in experimental diabetes: Effects on voltage-activated calcium channels. J Neurochem. 2011;118:224–236. doi: 10.1111/j.1471-4159.2011.07302.x. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.