

Abstract
Oxidation of d-ribulose-1,5-bisphosphate (ribulose-P2) during synthesis and/or storage produces d-glycero-2,3-pentodiulose-1,5-bisphosphate (pentodiulose-P2), a potent slow, tight-binding inhibitor of spinach (Spinacia oleracea L.) ribulose-P2 carboxylase/oxygenase (Rubisco). Differing degrees of contamination with pentodiulose-P2 caused the decline in Rubisco activity seen during Rubisco assay time courses to vary between different preparations of ribulose-P2. With some ribulose-P2 preparations, this compound can be the dominant cause of the decline, far exceeding the significance of the catalytic by-product, d-xylulose-1,5-bisphosphate. Unlike xylulose-1,5-bisphosphate, pentodiulose-P2 did not appear to be a significant by-product of catalysis by wild-type Rubisco at saturating CO2 concentration. It was produced slowly during frozen storage of ribulose-P2, even at low pH, more rapidly in Rubisco assay buffers at room temperature, and particularly rapidly on deliberate oxidation of ribulose-P2 with Cu2+. Its formation was prevented by the exclusion of transition metals and O2. Pentodiulose-P2 was unstable and decayed to a variety of other less-inhibitory compounds, particularly in the presence of some buffers. However, it formed a tight, stable complex with carbamylated spinach Rubisco, which could be isolated by gel filtration, presumably because its structure mimics that of the enediol intermediate of Rubisco catalysis. Rubisco catalyzes the cleavage of pentodiulose-P2 by H2O2, producing P-glycolate.
The photosynthetic CO2-fixing enzyme Rubisco catalyzes both carboxylation and oxygenation of ribulose-P2, producing either two molecules of P-glycerate or a molecule each of P-glycerate and P-glycolate. The catalytic reaction proceeds via enolization of ribulose-P2 following abstraction of the proton attached to C-3 by an enzymatic base. This is followed by the attack of CO2 or O2 on C-2 of the resultant 2,3-enediol. Depending on whether the attacking species is CO2 or O2, either 2′-carboxy-3-keto-arabinitol-1,5-bisphosphate or 2′-peroxy-3-keto-arabinitol-1,5-bisphosphate is formed as enzyme-bound intermediates. These intermediates are hydrated and cleaved between C-2 and C-3 to yield the products (for review, see Andrews and Lorimer, 1987; Hartman and Harpel, 1994; Gutteridge and Gatenby, 1995; Cleland et al., 1998).
The activity of higher-plant Rubisco declines during assay in vitro. The decrease in activity is approximately exponential and commences as soon as fully activated Rubisco is mixed with ribulose-P2. Depending on the conditions, it proceeds with a half-time of 5 to 10 min and eventually reaches an apparent steady state in which the final activity is 20 to 50% of the initial activity. This phenomenon, now generally called “fallover,” has been explained in terms of the production of isomers of ribulose-P2, xylulose-P2, and ketoarabinitol-P2 by stereochemically incorrect reprotonation of the enediol intermediate (Edmondson et al., 1990a, 1990b, 1990c, 1990d; Zhu and Jensen, 1991a, 1991b). However, two observations suggest that isomer production can explain fallover only partially. First, millimolar concentrations of H2O2 alleviate the decline in activity. Although H2O2 inhibits the initial rate, the subsequent decline occurs more slowly (Badger et al., 1980; Edmondson et al., 1990a). Since such concentrations of H2O2 do not rapidly destroy pentulose bisphosphates, this observation is difficult to explain in terms of inhibition by these isomers. Second, the rate and extent of the activity decline can vary between different preparations of ribulose-P2, and this is inconsistent with inhibition by reaction by-products. Both of these observations might be consistent with the variable presence of an inhibitor of Rubisco in ribulose-P2 preparations if that inhibitor was a slow, tight binder and if it was destroyed by H2O2.
A clue about the possible identity of such an inhibitor was provided by observations with certain site-directed mutants of Rhodospirillum rubrum Rubisco that were severely disabled in their ability to promote carboxylation of the enediol intermediate (Chen and Hartman, 1995; Harpel et al., 1995b). These mutants produced by-products of the oxygenase reaction that were identified as pentodiulose-P2 and its product of benzylic-acid-type rearrangement, carboxytetritol-P2. Pentodiulose-P2 has a vicinal dicarbonyl function that renders it sensitive to cleavage by H2O2, and it is possible that it might also be formed from ribulose-P2 by nonenzymatic oxidation.
This study was motivated by frustration with the variable quality of ribulose-P2 preparations. Both of our laboratories have experienced ribulose-P2 preparations that for no obvious reason have induced such rapid and extensive declines in activity of higher-plant Rubisco that the preparations were unusable. We undertook this study to investigate the identity and source of potential Rubisco inhibitors in the hope of discovering means of suppressing their formation.
MATERIALS AND METHODS
Materials
[1-14C]Ribulose-P2 was synthesized and purified according to the method of Kane et al. (1994) and stored at pH 2.8 and at −80°C for approximately 4 years before use. This preparation contained impurities arising from decay of ribulose-P2 during storage (see Results). [1-3H]Ribulose-P2 was synthesized from [2-3H]Glc (Amersham) using the same method but purified by anion-exchange chromatography on a Mono-Q 5/5 column (Pharmacia) equilibrated with 3 mm HCl, using a shallow, linear NaCl gradient rising at 20 mm column volume−1 from 0 to 300 mm. To remove NaCl, pooled fractions were concentrated 10-fold in vacuo and then gel filtered at 4°C on Sephadex G-10 equilibrated with 3 mm HCl. Ribulose-P2-containing fractions were pooled, snap frozen, and stored in liquid N2. This preparation was free of impurities initially and showed no signs of them even after storage for 1 year in liquid N2. Unlabeled ribulose-P2 was synthesized from Rib-5-phosphate (Sigma) according to the method of Horecker et al. (1958) and purified on a 4.4- × 100-cm column of AG1-X8 (Cl− form, Bio-Rad) equilibrated with 3 mm HCl, using a linear NaCl gradient rising at 24 mm column volume−1 from 100 to 250 mm. Pooled fractions containing ribulose-P2 that eluted at 150 mm NaCl were snap frozen in small aliquots and stored in liquid N2.
Rubisco was purified from spinach (Spinacia oleracea L.) (Edmondson et al., 1990a; Morell et al., 1997) and Rhodospirillum rubrum (Andrews and Kane, 1991) as described previously. Before use, spinach Rubisco was dialyzed at 4°C overnight against 50 mm Hepps (N-2-hydroxyethylpiperazine-N′-3-propanesulfonic acid)-NaOH buffer, pH 8.0, containing 15 mm MgCl2, 1 mm EDTA, and 10 mm NaHCO3, and then incubated at 50°C for 10 min and rapidly cooled. NaH14CO3 was supplied by Amersham, and scintillant, either Emulsifier Safe or Ultima Gold XR, by Canberra Packard. Sigma supplied catalase (bovine liver), creatine kinase (rabbit muscle), and carbonic anhydrase (bovine erythrocyte). All other enzyme preparations were obtained from Boehringer Mannheim as saturated (NH4)2SO4 suspensions and desalted before use.
Rubisco Fallover Assays
Rubisco assays were conducted aerobically (unless otherwise stated) for extended periods at 25°C in assay solutions containing 100 mm Hepps-NaOH buffer, pH 8.0, 20 mm MgCl2, 20 mm NaHCO3 (labeled with 14C to 2000 cpm nmol−1 in the case of 14C assays), 0.1 mg mL−1 BSA, 0.16 to 0.56 μg mL−1 spinach Rubisco, and 250 to 500 μm ribulose-P2. For spectrophotometric assays at 340 nm, based on the procedure of Lilley and Walker (1974), the following coupling enzymes and substrates were also included: 4 units mL−1 of yeast 3-phosphoglycerate kinase, 4 units mL−1 of rabbit muscle glyceraldehyde-3-phosphate dehydrogenase, 10 units mL−1 of rabbit muscle triose-phosphate isomerase, 4 units mL−1 of rabbit muscle glycerol-phosphate dehydrogenase, 4 units mL−1 of rabbit muscle creatine kinase, 0.1 mg mL−1 of bovine carbonic anhydrase, 0.2 mm NADH, 1 mm ATP, and 5 mm phosphocreatine. All components except ribulose-P2 were incubated for at least 10 min at 25°C before the ribulose-P2 preparation was added to initiate catalysis. For the radiometric assays, formic acid was added after various intervals to aliquots of the mixtures to a final concentration of 10% (v/v). The mixture was dried at 80°C and nonvolatile radioactivity was measured by scintillation spectrometry. For both assay methods, data from the resultant time courses were fitted to the following equation, which models an exponential decay of activity with time (t) from an initial, higher-activity form (Vf) to a final, lower-activity form (vf) with a half-time (t1/2) of ln 2/kobs (Edmondson et al., 1990a):
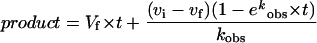 |
1 |
Analytical Anion-Exchange Chromatography
The procedure was adapted from that described by Harpel et al. (1993). A Mono-Q 5/5 column equilibrated with 10 mm Hepps-NaOH buffer, pH 8.0, containing 10 mm sodium borate and 50 mm NaCl was used to resolve 100 nmol or less of [1-14C]ribulose-P2 or [1-3H]ribulose-P2 from impurities. Samples were diluted, if necessary, with 5 mm Hepps-NaOH buffer, pH 8.0, containing 5 mm sodium borate immediately before application. Two different elution protocols, both using very shallow NaCl gradients in the equilibration buffer, were used as noted in the figures. Protocol A (analogous to that of Chen and Hartman [1995]) commenced at 50 mm NaCl, proceeded to 75 mm at 6.3 mm column volume−1, to 125 mm at 1 mm column volume−1, to 200 mm at 3.8 mm column volume−1, and finally to 500 mm at 60 mm column volume−1. Protocol B commenced at 50 mm NaCl, proceeded to 225 mm at 2.1 mm column volume−1, and finally to 500 mm at 55 mm column volume−1. For both protocols, the flow rate of the eluant was 1 mL min−1. Fractions, usually of 1-min duration, were collected and their radioactivity was measured after addition of an equal volume of Ultima Gold XR scintillant. Larger amounts of labeled ribulose-P2 (up to 1 μmol) were chromatographed on a Hema-IEC BIO 1000 Q 10U column (4.6 × 250 mm, Alltech) using the same eluant but with a steeper separating NaCl gradient (4 mm column volume−1) and a faster flow rate (2 mL min−1).
RESULTS
A H2O2-Sensitive Product of Ribulose-P2 Oxidation Is a Major Cause of Fallover in Vitro
We noticed that the rate and extent of the slow inactivation (fallover) of higher-plant Rubisco observed during extended assays varied between different preparations of ribulose-P2 (data not shown). Preparations purified by anion-exchange chromatography with shallow NaCl gradients (see Methods) showed the least fallover and did not appear to deteriorate in this respect if stored at pH 2.8 in liquid N2, even for periods as long as years. Storage of ribulose-P2 preparations at a more neutral pH or at −80°C or even a single additional freeze-thaw cycle caused fallover to be noticeably more rapid and extensive. The deterioration was particularly rapid if ribulose-P2 was stored at room temperature in buffers usually used for Rubisco assays. Effects became noticeable within 2 h of storage and worsened steadily thereafter. The initial rates of Rubisco assays were reduced, the deceleration occurred more rapidly, and the final, steady-state activity eventually attained was strongly and progressively suppressed as the period of storage of ribulose-P2 lengthened (Fig. 1A).
Figure 1.

Effect of storage of ribulose-P2 and treatment with H2O2 on the time courses of Rubisco activity assays. Assays were conducted as described in Methods using either the spectrophotometric (A and B) or the radiometric (C and D) procedure. Ribulose-P2 was derived from a freshly thawed aliquot that had been stored in liquid N2 since synthesis. It was either used without further treatment or preincubated aseptically in 125 mm Hepps-NaOH buffer, pH 8.0, containing 25 mm MgCl2, at 22°C before use. A, Effect of increasing periods of preincubation: ——, 0 min; — — —, 130 min; - - -, 240 min; — - —, 480 min; and — - - —, 1445 min. B, Effect of exclusion of O2 during ribulose-P2 storage and/or Rubisco assay: ——, fresh ribulose-P2/aerobic assay; - - -, fresh ribulose-P2/anaerobic assay; — - - —, ribulose-P2 stored aerobically for 24 h/aerobic assay; and — — —, ribulose-P2 stored anaerobically for 27 h in Mg2+-free buffer in the presence of Chelex 100 resin (100–200 mesh, Na+ form, Bio-Rad)/anaerobic assay. C, Reversal of inhibition by exposure of the stored ribulose-P2 preparation to H2O2. Ribulose-P2 was used without storage (•), stored for 24 h (▿), or stored for 24 h followed by treatment with 1 m H2O2 for 30 min and removal of H2O2 with 500 units of bovine catalase (□). D, Effect of H2O2 when present during assay of Rubisco using fresh ribulose-P2: •, no H2O2; □, 2 mm H2O2; Δ, 4 mm H2O2; and ○, 6 mm H2O2.
Whereas the presence or absence of O2 during the assay itself had no effect on the time course, the exclusion of O2 and metals from the solution in which ribulose-P2 was stored prevented the deterioration. Exclusion of either O2 or metals was partially effective (data not shown), and when both were excluded simultaneously, the time course showed slightly less deceleration even than the control with freshly thawed ribulose-P2 (Fig. 1B). Fallover induced by stored ribulose-P2 was also ameliorated by treatment of the preparation with molar concentrations of H2O2 before addition to the assay mixture (Fig. 1B). We confirmed earlier observations (Badger et al., 1980; Edmondson et al., 1990a) that millimolar concentrations of H2O2 reduced the decline in Rubisco activity if present in the assay mixture, although such concentrations also inhibit the initial activity (Fig. 1D). These lower H2O2 concentrations have little or no ameliorating effect if used as a ribulose-P2 pretreatment (data not shown).
The degree of fallover depended on the buffer in which ribulose-P2 was stored (Table I). It was worst when the storage solution was buffered with Hepps, Hepes, or Mes. However, ribulose-P2 stored in Tris, Tricine, Gly, or triethanolamine buffers showed little or no enhancement of fallover compared with controls with freshly thawed ribulose-P2. Bicine buffer gave intermediate results. Varying pH between 6.0 and 8.0 had little effect, strong inhibition being seen with Mes at pH 6.0 and Hepps and Hepes at pH 8.0. Storage of ribulose-P2 in unbuffered solution was strongly inhibitory at pH 8.2 and 6.3 (giving results similar to or worse than Hepps, Hepes, or Mes at the same pH), less inhibitory at pH 4.0, and not inhibitory at all at pH 2.8. Significantly, storage in a buffer mixture containing both Tris and Hepes caused little enhancement of fallover and the enhancement caused by storage in Hepps was substantially reversed by subsequent storage in Tris. However, the reversing effect of Tris was not instantaneous because substitution of Tris for Hepps in the assay buffer caused little reversal of the enhancement of fallover caused by storage of ribulose-P2 in Hepps (data not shown).
Table I.
Influence of the buffer used for storage of ribulose-P2 on the slow inhibition observed in subsequent Rubisco activity assays
| Buffer Used for Storage | pH | Storage Time | vi | vf/vi | t1/2 |
|---|---|---|---|---|---|
| h | s−1 | min | |||
| Control (no storage) | — | — | 2.9 | 0.39 | 15 |
| No buffer | 2.8 | 18 | 2.7 | 0.41 | 15 |
| No buffera | 4.0 | 24 | 2.4 | 0.26 | 7.8 |
| No buffera | 6.3 | 24 | 2.5 | 0.07 | 1.2 |
| No buffera | 8.2 | 30 | 0.9 | 0.06 | 0.8 |
| Hepes-NaOH (100 mm) | 8.0 | 18 | 1.3 | 0.06 | 2.2 |
| Hepps-NaOH (100 mm) | 8.0 | 18 | 1.2 | 0.07 | 3.0 |
| Mes-NaOH (100 mm) | 6.0 | 18 | 2.3 | 0.15 | 2.7 |
| Bicine-NaOH (100 mm) | 8.0 | 18 | 2.3 | 0.22 | 6.9 |
| Glycine-NaOH (100 mm) | 8.0 | 18 | 2.8 | 0.37 | 11 |
| Tricine-NaOH (100 mm) | 8.0 | 46 | 2.9 | 0.38 | 11 |
| Tris-HCl (100 mm) | 8.0 | 18 | 2.7 | 0.41 | 14 |
| Tris-HCl (100 mm), MgCl2 (20 mm) | 8.0 | 18 | 2.7 | 0.40 | 11 |
| Triethanolamine-HCl (100 mm) | 8.3 | 20 | 2.9 | 0.39 | 11 |
| Hepes-NaOH (50 mm) | 8.0 | 4 | 2.2 | 0.21 | 2.5 |
| Hepes-NaOH (50 mm), Tris-HCl (50 mm) | 8.0 | 4 | 2.7 | 0.26 | 9.6 |
| Hepps-NaOH then Tris-HClb | 8.0 | 18/18 | 2.1 | 0.49 | 7.0 |
Ribulose-P2 was stored at room temperature in aseptically filtered solutions with the stated components for the times shown and then used to initiate Rubisco activity assays according to the spectrophotometric method described in Methods. The time-course data were then fitted to Equation 1 to estimate the parameters shown.
pH adjusted with NaOH.
Ribulose-P2 was stored for 18 h in 100 mm Hepps-NaOH, then diluted 6-fold into 100 mm Tris-HCl and stored for a further 18 h before addition to an assay containing 100 mm Tris-HCl, pH 8.0.
Pi was released from ribulose-P2 during storage at room temperature at pH 6.3 and 8.0 at rates approximating 0.1% per hour regardless of the buffer present. Pi release was suppressed to barely detectable levels at pH 4.0 and below. However, there was no correlation between the amount of Pi in a stored ribulose-P2 preparation and the degree of fallover it induced (data not shown).
A Tight-Binding Impurity Accumulates during Storage of Ribulose-P2 at −80°C
Storage of [1-14C]ribulose-P2 preparations at pH 2.8 and −80°C for extended periods led to the formation of several impurities that could be resolved from ribulose-P2 by anion-exchange chromatography (Fig. 2B). The most abundant of these, designated “X,” eluted higher in the NaCl gradient than ribulose-P2 itself, suggesting that it contained at least two phosphate moieties. When such preparations were allowed to react to completion with fully activated spinach Rubisco, a residual fraction of the radioactivity remained bound to the enzyme and could be isolated by gel filtration (Fig. 2A). The binding was quite tight, with no sign of release of labeled material from the trailing side of the high-Mr peak. In this experiment, 8% of the radioactivity in the starting ribulose-P2 preparation remained bound to Rubisco, and this was equivalent to 13% of the Rubisco active sites present. The bound radioactivity released upon denaturation of the protein with SDS contained no ribulose-P2, but most of the impurities observed in the starting preparation were present (Fig. 2C). X predominated to a greater degree among these than it did in the starting preparation, and it appeared that X might be the tight-binding impurity, with the other impurities being derived from it after release from the enzyme. This interpretation was supported by the approximate agreement between the amount of X measured by anion-exchange chromatography (6.3% of the total radioactivity in Fig. 2B) and that observed bound to Rubisco (8%, Fig. 2A). In a similar experiment with another labeled ribulose-P2 preparation that contained a barely detectable amount of X (< 1%), the high-Mr complex isolated by gel filtration contained only 0.7% of the total radioactivity (data not shown). From this correspondence, we conclude that the bound material is derived from the starting preparation and is not produced in significant amounts as a by-product of catalysis at saturating CO2.
Figure 2.

An impurity in a 14C-ribulose-P2 preparation that binds tightly to Rubisco. A, [1-14C]ribulose-P2 (freshly thawed after 4 years of storage at pH 2.8 and at −80°C, final concentration 30 μm, 82,000 cpm nmol−1) was mixed with preactivated spinach Rubisco (final concentration 17 μm) in 45 mm Hepps-NaOH buffer, pH 8.0, containing 13 mm MgCl2, 1 mm EDTA, and 9 mm NaHCO3. After 10 min at 22°C, 0.56 mL of this solution was applied to a 1- × 26-cm column of Sephadex G-50 (fine) equilibrated with the same buffer components at a flow rate of 0.63 mL min−1. The effluent was monitored for radioactivity and A280. B, Anion-exchange chromatography on a Mono-Q 5/5 column (see “Materials and Methods,” elution protocol A) of the [1-14C]ribulose-P2 preparation used in A. C, Fractions comprising the high-Mr peak shown in A were pooled and SDS was added to 1% (w/v). Protein was removed by ultrafiltration and an aliquot of the filtrate was chromatographed as in B, approximately 2 h after the addition of SDS.
Studies with X isolated chromatographically from stored labeled ribulose-P2 preparations revealed that it was unstable and supported the idea that the other impurities were derived from it (Fig. 3). Rechromatography of the isolated material after overnight storage in liquid N2 revealed not only a predominant peak of X but also the other impurities present in the starting ribulose-P2 preparation (Fig. 3B). In addition to degradation during storage and the associated freeze-thaw cycle, X appeared to be degrading while bound to the anion-exchange column, giving rise to diffuse peaks of radioactivity eluting before X itself.
Figure 3.

Characterization of the tight-binding impurity in [1-14C]ribulose-P2. A, Twenty-six nanomoles of [1-14C]ribulose-P2 (82,000 cpm nmol−1) was chromatographed on a Mono-Q 5/5 column as described in Methods (elution protocol B). Fractions comprising the 52-min peak were pooled and divided into 500-μL aliquots, each of which was subjected to one of the following treatments and then rechromatographed. B, An aliquot was snap frozen, stored overnight in liquid N2, diluted to 1 mL with column-starting buffer, and rechromatographed. C, An aliquot was diluted to 1 mL with 50 mm Hepps-NaOH buffer, pH 8.0, supplemented with o-phenylenediamine to 100 mm, and rechromatographed after storage for 1 h in the dark at 22°C. D, To another aliquot, H2O2 was added to a final concentration of 1.1 m. After 1 h at room temperature, water was added to 2 mL and 2,600 units of bovine catalase was added. Thirty minutes later, the mixture was snap frozen and stored overnight in liquid N2 before rechromatography. The resulting chromatogram was aligned against a separate chromatogram of a 6:1 mixture of [3H]-P-glycolate and [3H]-P-glycerate generated from [1-3H]ribulose-P2 with R. rubrum Rubisco in a solution equilibrated with 500 μL L−1 CO2 in O2 as described by Kane et al. (1994) but omitting the phosphatase treatment. The larger peak of [3H]P-glycolate, eluting near 28 min and clearly resolved from the closely preceding [3H]P-glycerate peak, aligns precisely with the 28-min peak derived from the impurity. The peaks eluting near 4 min are presumably the products of phosphatase contamination of the enzyme preparations.
Guessing that X might be pentodiulose-P2 produced by oxidation of ribulose-P2 during storage (Scheme 1), we reacted isolated X with o-phenylenediamine, which converts vicinal dicarbonyl compounds to 2,3-substituted quinoxalines. This had two effects on the chromatographic behavior of X: it eluted slightly earlier in the NaCl gradient (consistent with the observations of Chen and Hartman [1995]) and the diffuse, early eluting peaks were suppressed (Fig. 3C).
To confirm that X was indeed pentodiulose-P2, we exposed it to 1.1 m H2O2 and then catalase. A very clean chromatogram resulted, showing only a major peak of P-glycolate and a minor peak not retarded by the column that presumably was glycolate resulting from phosphatase contamination in the catalase preparation (Fig. 3D). The other product of H2O2 cleavage of pentodiulose-P2 would be P-glycerate derived from carbons 3 to 5. This would not be labeled, since the radioisotope was originally in the carbon 1 position of ribulose-P2 (Scheme 1).
Pentodiulose-P2 Is Produced by Oxidation of Ribulose-P2 with Cu2+
Fresh preparations of [1-3H]ribulose-P2 or those stored in liquid N2 contained no detectable pentodiulose-P2 (Fig. 4, dotted line), but exposure of these preparations to millimolar concentrations of Cu2+ at pH 8.0 for 3.5 h produced pentodiulose-P2 (3.5% of starting radioactivity) and other early eluting compounds (Fig. 4, solid line). Inclusion of borate, which stabilizes pentodiulose-P2 to some extent (Chen and Hartman, 1995) during the Cu2+ oxidation process, did not increase the amount of pentodiulose-P2 recovered (data not shown), perhaps because borate also forms a complex with ribulose-P2 and therefore retards the oxidation. Column fractions in the vicinity of the pentodiulose-P2 peak were inhibitory when preincubated with fully activated spinach Rubisco before assay, and the peak of inhibition corresponded to the peak of 3H radioactivity (Fig. 4). Inhibition increased as the concentration of pentodiulose-P2 in the Rubisco assay increased but it plateaued at approximately 75% inhibition (Fig. 4, inset). This negatively cooperative pattern of inhibition is reminiscent of other tight-binding inhibitors of Rubisco, such as carboxyarabinitol-1-P (Gutteridge et al., 1986; Berry et al., 1987). Since the Kd appeared to be close to the concentration of Rubisco active sites present in the inhibition assays (35 nm), the data were fitted to an equation that modeled tight-binding inhibition (Fig. 4, inset). This estimated the Kd to be 130 nm. However, the negatively cooperative nature of the binding caused the data to fit the model quite poorly. Clearly, the binding affinity is tighter than this average estimate when site occupancy is low and looser when occupancy is high. Since 75 mm NaCl and 4 mm sodium borate were carried into the assay with the aliquots of column fractions, this estimate must be regarded as a maximum estimate.
Figure 4.

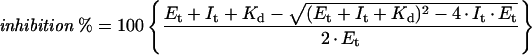 |
2 |
The behavior of pentodiulose-P2 produced by deliberate oxidation of [1-3H]ribulose-P2 with Cu2+ was identical to that of the inhibitor in stored ribulose-P2 preparations. It bound tightly to Rubisco to form a complex that could be isolated by gel filtration in a manner similar to that shown in Figure 2A. The radioactivity released from this high-Mr complex by addition of SDS chromatographed predominantly in the X peak (analogous to Fig. 2C). Alternatively, the label could be released by exposure of the complex to 5 mm H2O2, where P-glycolate and a trace of nonphosphorylated material were the only products (analogous to Fig. 3D). Cleavage of pentodiulose-P2 while it was bound to the Rubisco active site was particularly facile. Five-millimolar H2O2 effected complete cleavage in 30 min at 22°C, and this accords with the effectiveness of such low concentrations in ameliorating the deceleration of Rubisco assay time courses (Fig. 1D). H2O2 concentrations 2 to 3 orders of magnitude higher are required to cleave pentodiulose-P2 when it is free in solution (Figs. 1C and 3D); therefore, we conclude that Rubisco must catalyze the peroxidative cleavage of pentodiulose-P2 (Scheme 2).
DISCUSSION
Pentodiulose-P2 Is Produced by Nonenzymatic Oxidation of Ribulose-P2
Our data establish that pentodiulose-P2 can be produced nonenzymatically from ribulose-P2 in a reaction that depends on, or is accelerated by, O2 and Chelex-removable metals (Scheme 1). It is possible that transition metals are essential to the oxidation process and are reoxidized by O2. If so, pentodiulose-P2 might still be produced in the absence of O2 in amounts stoichiometric with the transition metals present. Although small, such amounts are still likely to be significant compared with the Rubisco concentrations present in activity assays and, therefore, inhibitory. Pretreatment of assay buffer components with Chelex would be only partially effective in removing metal contamination because Mg2+, which cannot be treated with Chelex, is required for activity and is a likely source of the contamination. Deliberate oxidation of ribulose-P2 with Cu2+ provides a ready means of synthesizing pentodiulose-P2.
The identity of the oxidation product as pentodiulose-P2 was established by its chromatographic behavior and that of its adduct with o-phenylenediamine, and the conversion of label present at C-1 to P-glycolate by cleavage with H2O2 (Figs. 2–4). Consistent with this identification, the mass spectrum (electron-impact ionization) of the silyl derivative of the quinoxaline adduct formed between o-phenylenediamine and the oxidation product showed a prominent molecular ion at m/z = 436, together with the expected m-15 and m-90 fragment ions resulting from losses of a methyl group and trimethylsilanol, respectively (H.J. Kane and T.J. Andrews, unpublished results). The oxidation product appears to be identical in all of its properties to pentodiulose-P2, previously identified as a by-product of oxygenase catalysis by mutant (but not wild-type) R. rubrum Rubisco (Chen and Hartman, 1995; Harpel et al., 1995b).
The rate of the oxidation reaction that proceeds without deliberate addition of transition metals was not strongly dependent on pH between 8.3 and 6.0 but fell to insignificant levels at pH 2.8 (Table I). This might indicate the involvement of the enediol form of ribulose-P2 in the oxidation. Enediol formation by intramolecular abstraction (Richard, 1984) of the C-3 proton of ribulose-P2 by O atoms of the phosphate groups will only occur when the latter are unprotonated. However, whereas inhibitor production at low pH was insignificant on a time scale of days, significant amounts of the inhibitor accumulated at low pH during protracted frozen storage at −80°C (Fig. 2). Perhaps the very slow rate of inhibitor production under these conditions is offset by its greater stability.
Pentodiulose-P2 Is an Excellent Analog of the Enediol Intermediate
Pentodiulose-P2 bound very tightly to carbamylated spinach Rubisco, forming a complex that could be isolated by gel filtration (Fig. 2). We suggest that this strong binding affinity is a result of the close structural resemblance between the diulose-P2 and the enediol form of ribulose-P2 (Scheme 2). All of the heavy atoms of pentodiulose-P2 can adopt the same positions as those of the enediol, and the planar configuration of C-4, C-3, O-3, C-2, O-2, and C-1 can be emulated.
Since the complex can be isolated by gel filtration, taking tens of minutes to hours without any sign of leakage of label from the trailing side of the peak (Fig. 2), pentodiulose-P2 must not be prone to further conversion on the active site to other, less tightly binding products. In this respect, the spinach enzyme differs markedly from the R. rubrum enzyme. Wild-type R. rubrum Rubisco and its K329A mutant both convert pentodiulose-P2 to carboxytetritol-P2 (Harpel et al., 1995a, 1995b). This rearrangement product did not appear to remain tightly bound to the K329A enzyme, since denaturation of the protein was not required to recover it (Harpel et al., 1995b). By contrast, the E48Q mutant of R. rubrum Rubisco apparently catalyzed the O2-dependent cleavage of pentodiulose-P2 to P-glycolate and P-glycerate (Chen and Hartman, 1995). The inability of higher-plant Rubiscos to catalyze similar transformations of pentodiulose-P2 to less tightly binding products may be a reason for their susceptibility to fallover. Most bacterial and algal Rubiscos show little sign of fallover (Gibson and Tabita, 1979; Andrews and Ballment, 1984; Yokota and Kitaoka, 1989; Lee at al., 1993; Hernandez et al., 1996), and we suggest that this might be because they resemble the R. rubrum enzyme in being able to convert pentodiulose-P2 to quickly released products.
We do not know whether, like ribulose-P2 and xylulose-P2 (Jordan and Chollet, 1983; Zhu and Jensen, 1991a), pentodiulose-P2 can also bind tightly to the uncarbamylated active site. Fallover is not accompanied by significant decarbamylation above pH 8.0 (Edmondson et al., 1990b; Zhu and Jensen, 1991b). If pentodiulose-P2 is a major cause of fallover, as our data appear to indicate, then this inhibitor may resemble carboxyarabinitol-1-P in its preference for the carbamylated active site (Seemann et al., 1985).
Pentodiulose-P2 Is Unstable
Pentodiulose-P2 decays to a variety of other compounds. The chromatographic profiles (Figs. 2–4) indicate that compounds with zero, one, and two phosphate groups are present among the products, sometimes in multiple ionic forms within each class. The label eluting higher in the NaCl gradient than pentodiulose-P2 (e.g. the 82-min peak in Fig. 2, B and C) may represent the product of the benzylic acid-type rearrangement, carboxytetritol-P2. Plausible decay pathways resulting in the successive elimination of both phosphate groups of pentodiulose-P2 may also be imagined. The evident diversity of decay products makes identifying all of them a large task and we did not attempt it. Chen and Hartman (1995) and Harpel et al. (1995a) also observed extensive decay of pentodiulose-P2 and showed that it could be slowed by complexing with borate. We also used this strategy wherever possible but found that it did not prevent the decay completely. In our studies pentodiulose-P2 never accumulated to an amount greater than 10% of that of the ribulose-P2 initially present. Apparently, a steady state was attained in which the rate of appearance of pentodiulose-P2 was balanced by the rate of its further decay. Although the presence of borate slowed the rate of decay, it slowed the oxidation leading to pentodiulose-P2 even more so that the steady-state amount of pentodiulose-P2 was reduced when borate was included during the oxidation process (data not shown).
The differences in severity of fallover induced by storage of ribulose-P2 in different buffers (Table I) seem most consistent with differences in the stability of pentodiulose-P2 in different buffers. Although it is possible that different degrees of contamination of the buffers with transition metals could give rise to differences in the amount of pentodiulose-P2 produced, serious fallover occurred even when ribulose-P2 was stored without buffer. Furthermore, buffers such as Tris were able slowly to reverse the fallover-promoting tendency after it had been induced by storage of ribulose-P2 in another buffer (Table I). This points to a role of buffers such as Tris, Tricine, Gly, and triethanolamine in accelerating the conversion of pentodiulose-P2 to less-inhibitory compounds. The worst fallover was induced after ribulose-P2 had been stored without buffer or in tertiary amine buffers. Primary and secondary amine buffers were among those inducing the least fallover. This might suggest a role for imines or enamines in the conversion were it not for the discordance with this pattern of the tertiary amine triethanolamine, which induces very little fallover (Table I). However, this discordance might be explained by the presence of approximately 1% mono- and diethanolamine in the reagent grade triethanolamine that we used. Another tertiary amine, Bicine, had intermediate fallover-inducing ability. Since diethanolamine is used in its synthesis (Good et al., 1966), Bicine may also contain traces of primary or secondary amines, explaining its intermediate status.
Reinterpretation of Earlier Observations about Fallover in the Light of Pentodiulose-P2
Our observations that pentodiulose-P2 can be a dominant cause of fallover, particularly with ribulose-P2 of indifferent quality, demand that previous explanations of fallover in terms of catalytic by-products be reconsidered. Pentodiulose-P2 resembles the fallover inhibitor(s) isolated from Rubisco reaction mixtures after complete consumption of ribulose-P2 in several respects. It binds to carbamylated Rubisco with similar affinity and its binding shows the same negative cooperativity. Furthermore, it is similarly unstable (Edmondson et al., 1990c). Edmondson et al. (1990d) showed that two inhibitory compounds were present in their preparations. One was clearly xylulose-P2 because it was destroyed by aldolase and it produced xylitol and arabinitol after reduction and dephosphorylation. They did not identify the other conclusively, but showed that it was resistant to aldolase but destroyed by brief exposure to mild alkali. Since the second inhibitor appeared to produce predominantly arabinitol on reduction and dephosphorylation, they speculated that it might be ketoarabinitol-P2, produced by misprotonation of the enediol form of ribulose-P2 at C-2. Zhu and Jensen (1990b) appeared to substantiate this speculation by detecting an inhibitor bound to Rubisco after catalysis that produced predominantly arabinitol-1,5-bisphosphate on reduction.
However, the observations of Lee et al. (1993) and Chen and Hartman (1995) that under some conditions borohydride reduction of pentodiulose-P2 can produce predominantly arabinitol-1,5-bisphosphate sound a note of caution about this interpretation. Although such reduction should produce the bisphosphates of ribitol, arabinitol, and xylitol in 1:2:1 proportions if it was stereochemically impartial, it must be concluded that there can be a strong preference for the arabinitol product under some conditions and, therefore, that evidence for the presence of ketoarabinitol-P2 based on detection of arabinitol-1,5-bisphosphate after reduction must be considered unreliable. In view of this and our present results, it appears that the second inhibitor observed by Edmondson et al. (1990d) may well have been pentodiulose-P2 present not as a catalytic by-product but by virtue of its pre-existence in the starting ribulose-P2 preparation. Its alkali lability is then readily explained by the enhancement of the benzylic acid-type rearrangement of pentodiulose-P2 expected under alkaline conditions. This reasoning, if correct, also implies that the product of the rearrangement, carboxytetritol-P2, is not a strong inhibitor of spinach Rubisco.
Another factor may have contributed to causing Edmondson et al. (1990d) to overlook pentodiulose-P2. They used Tris buffer during gel filtration of Rubisco-bound inhibitors and subsequent work-up. This may have caused the decay of much of the pentodiulose-P2 before reduction.
The relative contributions of pentodiulose-P2 and xylulose-P2 (and ketoarabinitol-P2, if it exits) to fallover is now a moot point. The effect of pentodiulose-P2 clearly dominates when it is present in significant amounts in the starting ribulose-P2. Even the reduced fallover observed with the best ribulose-P2 preparations might not be caused solely by catalytic by-products, because storage of ribulose-P2 in the absence of transition metals and O2 caused slight further alleviation of fallover (Fig. 1B). This may indicate that even our best preparations of ribulose-P2 have traces of pentodiulose-P2 that disappear due to instability when stored under conditions preventing further oxidation. Fallover observed after this pretreatment was the slightest we have ever observed (vf/vI = 0.65) and this residual level may represent the true contribution of xylulose-P2 production. However, even in this circumstance, the possible contribution of traces of pentodiulose-P2 still must not be overlooked. Even in the absence of O2, traces of transition metals introduced with the Mg2+ required for assay might result in the production of stoichiometric amounts of pentodiulose-P2 during the assay period itself.
Rubisco Catalyzes the Cleavage of Pentodiulose-P2 by H2O2
If pentodiulose-P2 emulates the enediol intermediate in binding to Rubisco's active site, its facile cleavage by low concentrations of H2O2 can be easily understood (Scheme 2). O2 addition to the enediol and H2O2 addition to pentodiulose-P2 are closely analogous and would produce the same peroxyketone intermediate that would then be cleaved to P-glycolate and P-glycerate by the normal oxygenase pathway. H2O2-assisted release of Rubisco from the otherwise dead-end complex with pentodiulose-P2 provides a satisfying explanation for the long-standing observation that H2O2 suppresses fallover (Badger et al., 1980). Earlier observations of an apparent lack of an effect of H2O2 on the isolated fallover inhibitor (Edmondson et al., 1990c) are now explicable. The millimolar H2O2 concentrations used in that study are not effective in cleaving pentodiulose-P2 when free in solution; molar H2O2 concentrations are required for this purpose. This relative resistance of unbound pentodiulose-P2 to H2O2 cleavage might be expected if one of its keto groups was predominantly hydrated in solution. Complexation with borate would also promote hydration. On the active site, however, only the diketo form would emulate the enediol intermediate. The instability of pentodiulose-P2 has so far frustrated attempts to chromatographically isolate it in quantities sufficient for more detailed kinetic studies of its Rubisco-catalyzed cleavage by H2O2.
How Should Pentodiulose-P2 Formation Be Suppressed during Synthesis and Storage of Ribulose-P2?
The need for ribulose-P2 preparations with predictable and reproducible properties gives this question some practical importance. Obviously, transition metals and O2 should be excluded whenever possible. This can be achieved easily enough during storage in liquid N2 but it is more difficult during synthesis when Mg2+, a potential source of other metals, must be present. The use of shallow NaCl gradients to separate pentodiulose-P2 from ribulose-P2 during preparative anion-exchange chromatography is therefore to be recommended. Storage at low pH assists in suppressing the oxidation but it may also improve the stability of the diulose-P2 product to some extent.
Is Pentodiulose-P2 Produced from Ribulose-P2 in Vivo?
Chloroplasts maintain high ribulose-P2 concentrations at around pH 8.0 during steady-state photosynthesis. They are well supplied with O2 and the transition metals required by the photosynthetic apparatus, such as Fe, Cu, and others, must be present at finite concentrations. Under these conditions it seems inevitable that pentodiulose-P2 must be formed and that it will accumulate on Rubisco's active sites and inhibit photosynthesis seriously unless specific mechanisms are present to prevent this from happening. There have been reports of a tight-binding Rubisco inhibitor in wheat and tobacco leaves that, unlike carboxyarabinitol-1-P, was present during the photoperiod but not in darkness (Keys et al., 1995; Paul et al., 1996; Parry et al., 1997). It was detected in amounts sufficient to inhibit 12% to 20% of the Rubisco present, shown to be neither carboxyarabinitol-1-P nor xylulose-P2, but otherwise not identified. Several of the properties of this daytime inhibitor (Keys at al., 1995; Parry et al., 1997) are suspiciously reminiscent of those of pentodiulose-P2 (this study) and the fallover inhibitor (Edmondson et al., 1990c): (a) It binds to Rubisco tightly enough to survive gel filtration but can be released by dialyzing or gel filtering the complex in 200 mm SO4−2, whereupon full recovery of the initial activity is obtained (Edmondson et al., 1990c); (b) it elutes from anion-exchange columns higher in the NaCl gradient than ribulose-P2; and (c) it is unstable at neutral pH and more stable in acidic conditions, and its inhibitory potency is diminished in Tris buffer. Reduction and dephosphorylation of the daytime inhibitor from wheat appeared to yield ribitol and arabinitol, the same products yielded by ribulose-P2 (Keys et al., 1995). In view of the potential for stereochemical bias in the reduction of pentodiulose-P2 discussed earlier, this observation is not inconsistent with the inhibitor being pentodiulose-P2. Further studies aimed at identifying the daytime inhibitor are warranted. In particular, its sensitivity to H2O2 needs to be investigated.
Study of the physiological mechanisms that limit the accumulation of pentodiulose-P2 in the chloroplast stroma is also required. Two different classes of mechanisms might be present. First, a mechanism for releasing Rubisco from its dead-end complex with pentodiulose-P2 must exist. Although cleavage of the inhibitor by H2O2 on the active site would accomplish this quite satisfactorily, it seems that millimolar concentrations of H2O2 are required for this purpose (Fig. 1D; Badger et al., 1980). Chloroplasts have a very effective ascorbate peroxidase mechanism for scavenging H2O2 that is thought to keep the steady-state pool of H2O2 below micromolar concentrations (Asada, 1994). Therefore, sufficient H2O2 probably would not be available for this release path to be feasible. Another release path could be provided by Rubisco activase, which is known to facilitate the release of a variety of inhibitors from both uncarbamylated and carbamylated Rubisco (Portis, 1992; Salvucci and Ogren, 1996). Activase is known to alleviate fallover in vitro (Robinson and Portis, 1989).
Second, if cleavage by H2O2 on Rubisco's active site is not possible, ancillary mechanisms for detoxification and disposal of pentodiulose-P2 must exist. If the daytime inhibitor discussed in the preceding paragraph is indeed pentodiulose-P2, then the small amounts detected must reflect a steady state between the rate of formation by oxidation of ribulose-P2 and the rate of disposal. A variety of possible disposal pathways may be theorized. Dephosphorylation could occur, either before or after rearrangement to carboxytetritol-P2 catalyzed by glyoxylaselike enzymes. Alternatively, H2O2-dependent cleavage to P-glycolate and P-glycerate catalyzed by a specific enzyme with a much greater affinity for H2O2 than Rubisco would not only dispose of pentodiulose-P2 safely but would also assist ascorbate peroxidase in maintaining H2O2 at a very low concentration.
Our present data for spinach Rubisco support those of Chen and Hartman (1995) for the wild-type R. rubrum enzyme in establishing that pentodiulose-P2 is not a significant catalytic by-product when CO2 is saturating. This is not surprising. Suppression of flux through the oxygenase catalytic pathway by CO2 would minimize formation of the peroxyketone intermediate (Scheme 2) from which pentodiulose-P2 might be derived by elimination of H2O2. However, reports that fallover of spinach Rubisco at pH 8.3 is exacerbated at subsaturating CO2 (Edmondson et al., 1990a), whereas decarbamylation is not (Edmondson et al., 1990b), raise suspicions that some pentodiulose-P2 might be produced enzymatically under these conditions. Further measurements of pentodiulose-P2 (and H2O2) production during Rubisco catalysis at subsaturating CO2 are required to address this issue. Any pentodiulose-P2 produced by higher-plant Rubisco under the physiologically relevant condition of CO2 undersaturation would need to be released from Rubisco and disposed of in the same manner as pentodiulose-P2 produced by nonenzymatic oxidation of ribulose-P2.
Scheme 1.

Transition-metal-catalyzed oxidation of ribulose-P2 and nonenzymatic cleavage of the resultant pentodiulose-P2 by high concentrations of H2O2. The asterisks indicate the fate of the C-1 carbon of ribulose-P2.
Scheme 2.

The structural analogy between the ribulose-P2 enediol and pentodiulose-P2 when bound within the active site of Rubisco, and the mechanistic analogy between Rubisco-catalyzed oxygenation of the enediol and Rubisco-catalyzed peroxidation of pentodiulose-P2 in the presence of low concentrations of H2O2. R = -CHOH-CH2OPO32−.
Abbreviations:
- carboxyarabinitol-1-P
2′-carboxy-d-arabinitol-1-phosphate
- carboxytetritol-P2
2′-carboxytetritol-1,4-bisphosphate
- fallover
slow inactivation of Rubisco during catalysis
- ketoarabinitol-P2
3-keto-d-arabinitol-1,5-bisphosphate
- P-glycerate
d-3-phosphoglycerate
- P-glycolate
2-phosphoglycolate
- pentodiulose-P2
d-glycero-2,3-diulose-1,5-bisphosphate
- ribulose-P2
d-ribulose-1,5-bisphosphate
- xylulose-P2
d-xylulose-1,5-bisphosphate
Footnotes
This work was supported by the Australian National University's Centre for Molecular Structure and Function.
LITERATURE CITED
- Andrews TJ, Ballment B. Active site carbamate formation and reaction-intermediate-analog binding by ribulose bisphosphate carboxylase-oxygenase in the absence of its small subunits. Proc Natl Acad Sci USA. 1984;81:3660–3664. doi: 10.1073/pnas.81.12.3660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andrews TJ, Kane HJ. Pyruvate is a by-product of catalysis by ribulosebisphosphate carboxylase/oxygenase. J Biol Chem. 1991;266:9447–9452. [PubMed] [Google Scholar]
- Andrews TJ, Lorimer GH (1987) Rubisco: structure, mechanisms, and prospects for improvement. In MD Hatch, NK Boardman, eds, The Biochemistry of Plants: A Comprehensive Treatise, Vol. 10, Photosynthesis. Academic Press, New York, pp 131–218
- Asada K. Production and action of active oxygen species in photosynthetic tissues. In: Foyer CH, Mullineaux PM, editors. Causes of Photooxidative Stress and Amelioration of Defense Systems in Plants. Boca Raton, FL: CRC Press; 1994. pp. 77–104. [Google Scholar]
- Badger MR, Andrews TJ, Canvin DT, Lorimer GH. Interaction of hydrogen peroxide with ribulose bisphosphate carboxylase-oxygenase. J Biol Chem. 1980;255:7870–7875. [PubMed] [Google Scholar]
- Berry JA, Lorimer GH, Pierce J, Seemann JR, Meek J, Freas S. Isolation, identification, and synthesis of 2-carboxyarabinitol 1-phosphate, a diurnal regulator of ribulose-bisphosphate carboxylase activity. Proc Natl Acad Sci USA. 1987;84:734–738. doi: 10.1073/pnas.84.3.734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen Y-R, Hartman FC. A signature of the oxygenase intermediate in catalysis by ribulose-bisphosphate carboxylase/oxygenase as provided by a site-directed mutant. J Biol Chem. 1995;270:11741–11744. doi: 10.1074/jbc.270.20.11741. [DOI] [PubMed] [Google Scholar]
- Cleland WW, Andrews TJ, Gutteridge S, Hartman FC, Lorimer GH. The mechanism of Rubisco: the carbamate as general base. Chem Rev. 1998;98:549–561. doi: 10.1021/cr970010r. [DOI] [PubMed] [Google Scholar]
- Edmondson DL, Badger MR, Andrews TJ. A kinetic characterization of slow inactivation of ribulosebisphosphate carboxylase during catalysis. Plant Physiol. 1990a;93:1376–1382. doi: 10.1104/pp.93.4.1376. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Edmondson DL, Badger MR, Andrews TJ. Slow inactivation of ribulosebisphosphate carboxylase during catalysis is not due to decarbamylation of the catalytic site. Plant Physiol. 1990b;93:1383–1389. doi: 10.1104/pp.93.4.1383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Edmondson DL, Badger MR, Andrews TJ. Slow inactivation of ribulosebisphosphate carboxylase during catalysis is caused by accumulation of a slow, tight-binding inhibitor at the catalytic site. Plant Physiol. 1990c;93:1390–1397. doi: 10.1104/pp.93.4.1390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Edmondson DL, Kane HJ, Andrews TJ. Substrate isomerization inhibits ribulosebisphosphate carboxylase-oxygenase during catalysis. FEBS Lett. 1990d;260:62–66. [Google Scholar]
- Gibson JL, Tabita FR. Activation of ribulose 1,5-bisphosphate carboxylase from Rhodopseudomonas sphaeroides: probable role of the small subunit. J Bacteriol. 1979;140:1023–1027. doi: 10.1128/jb.140.3.1023-1027.1979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Good NE, Winget GD, Winter W, Connolly TN, Izawa S, Singh RMM. Hydrogen ion buffers for biological research. Biochemistry. 1966;5:467–477. doi: 10.1021/bi00866a011. [DOI] [PubMed] [Google Scholar]
- Gutteridge S, Gatenby AA. Rubisco synthesis, assembly, mechanism, and regulation. Plant Cell. 1995;7:809–819. doi: 10.1105/tpc.7.7.809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gutteridge S, Parry MAJ, Burton S, Keys AJ, Mudd A, Feeney J, Servaites JC, Pierce J. A nocturnal inhibitor of carboxylation in leaves. Nature. 1986;324:274–276. [Google Scholar]
- Harpel MR, Chen Y-R, Hartman FC (1995a) The nature and alternate fates of the ribulose 1,5-bisphosphate (RuBP) carboxylase/oxygenase (Rubisco) oxygenation intermediate. In P Mathis, ed, Photosynthesis: from Light to the Biosphere. Kluwer Academic Publishers, Dordrecht, The Netherlands, pp 159–162
- Harpel MR, Lee EH, Hartman FC. Anion-exchange analysis of ribulose-bisphosphate carboxylase/oxygenase reactions: CO2/O2 specificity determination and identification of side products. Anal Biochem. 1993;209:367–374. doi: 10.1006/abio.1993.1136. [DOI] [PubMed] [Google Scholar]
- Harpel MR, Serpersu EH, Lamerdin JA, Huang ZH, Gage DA, Hartman FC. Oxygenation mechanism of ribulose-bisphosphate carboxylase/oxygenase: structure and origin of 2-carboxytetritol 1,4-bisphosphate, a novel O2-dependent side product generated by a site-directed mutant. Biochemistry. 1995b;34:11296–11306. doi: 10.1021/bi00035a039. [DOI] [PubMed] [Google Scholar]
- Hartman FC, Harpel MR. Structure, function, regulation, and assembly of d-ribulose-1,5-bisphosphate carboxylase/oxygenase. Annu Rev Biochem. 1994;63:197–234. doi: 10.1146/annurev.bi.63.070194.001213. [DOI] [PubMed] [Google Scholar]
- Hernandez JM, Baker SH, Lorbach SC, Shively JM, Tabita FR. Deduced amino acid sequence, functional expression, and unique enzymatic properties of the form I and form II ribulose bisphosphate carboxylase oxygenase from the chemoautotrophic bacterium Thiobacillus denitrificans. J Bacteriol. 1996;178:347–356. doi: 10.1128/jb.178.2.347-356.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horecker BL, Hurwitz J, Weissbach A. Ribulose diphosphate. Biochem Prep. 1958;6:83–90. [Google Scholar]
- Jordan DB, Chollet R. Inhibition of ribulose bisphosphate carboxylase by substrate ribulose 1,5-bisphosphate. J Biol Chem. 1983;258:13752–13758. [PubMed] [Google Scholar]
- Kane HJ, Viil J, Entsch B, Paul K, Morell MK, Andrews TJ. An improved method for measuring the CO2/O2 specificity of ribulosebisphosphate carboxylase-oxygenase. Aust J Plant Physiol. 1994;21:449–461. [Google Scholar]
- Keys AJ, Major I, Parry MAJ. Is there another player in the game of Rubisco regulation? J Exp Bot. 1995;46:1245–1251. [Google Scholar]
- Lee EH, Harpel MR, Chen Y-R, Hartman FC. Perturbation of reaction-intermediate partitioning by a site-directed mutant of ribulose-bisphosphate carboxylase/oxygenase. J Biol Chem. 1993;268:26583–26591. [PubMed] [Google Scholar]
- Lilley RM, Walker DA. An improved spectrophotometric assay for ribulosebisphosphate carboxylase. Biochim Biophys Acta. 1974;358:226–229. doi: 10.1016/0005-2744(74)90274-5. [DOI] [PubMed] [Google Scholar]
- Morell MK, Wilkin J-M, Kane HJ, Andrews TJ. Side reactions catalyzed by ribulose-bisphosphate carboxylase in the presence and absence of small subunits. J Biol Chem. 1997;272:5445–5451. doi: 10.1074/jbc.272.9.5445. [DOI] [PubMed] [Google Scholar]
- Parry MAJ, Andralojc PJ, Parmar S, Keys AJ, Habash D, Paul MJ, Alred R, Quick WP, Servaites JC. Regulation of Rubisco by inhibitors in the light. Plant Cell Environ. 1997;20:528–534. [Google Scholar]
- Paul MJ, Andralojc PJ, Banks FM, Parry MAJ, Knight JS, Gray JC, Keys AJ. Altered Rubisco activity and amounts of a daytime tight-binding inhibitor in transgenic tobacco expressing limiting amounts of phosphoribulokinase. J Exp Bot. 1996;47:1963–1966. [Google Scholar]
- Portis AR. Regulation of ribulose 1,5-bisphosphate carboxylase/oxygenase activity. Annu Rev Plant Physiol Plant Mol Biol. 1992;43:415–437. [Google Scholar]
- Richard JP. Acid-base catalysis of the elimination and isomerization reactions of triose phosphates. J Am Chem Soc. 1984;106:4926–4936. [Google Scholar]
- Robinson SP, Portis AR. Ribulose-1,5-bisphosphate carboxylase/oxygenase activase protein prevents the in vitro decline in activity of ribulose-1,5-bisphosphate carboxylase/oxygenase. Plant Physiol. 1989;90:968–971. doi: 10.1104/pp.90.3.968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salvucci ME, Ogren WL. The mechanism of Rubisco activase: insights from studies of the properties and structure of the enzyme. Photosynthe Res. 1996;47:1–11. doi: 10.1007/BF00017748. [DOI] [PubMed] [Google Scholar]
- Seemann JR, Berry JA, Freas S, Krump MA. Regulation of ribulose bisphosphate carboxylase activity in vivo by a light-modulated inhibitor of catalysis. Proc Natl Acad Sci USA. 1985;82:8024–8028. doi: 10.1073/pnas.82.23.8024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yokota A, Kitaoka S. Linearity and functioning forms in the carboxylase reaction of spinach ribulose 1,5-bisphosphate carboxylase/oxygenase. Plant Cell Physiol. 1989;30:183–191. [Google Scholar]
- Zhu G, Jensen RG. Xylulose 1,5-bisphosphate synthesized by ribulose 1,5-bisphosphate carboxylase/oxygenase during catalysis binds to decarbamylated enzyme. Plant Physiol. 1991a;97:1348–1353. doi: 10.1104/pp.97.4.1348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu G, Jensen RG. Fallover of ribulose 1,5-bisphosphate carboxylase/oxygenase activity. Decarbamylation of catalytic sites depends on pH. Plant Physiol. 1991b;97:1354–1358. doi: 10.1104/pp.97.4.1354. [DOI] [PMC free article] [PubMed] [Google Scholar]