

Abstract
Pigeons trained on successive AB symbolic matching show emergent BA antisymmetry if they are also trained on successive AA oddity and BB identity (Urcuioli, 2008, Experiment 4). In other words, when tested on BA probe trials following training, they respond more to the comparisons on the reverse of the nonreinforced AB baseline trials than on the reverse of the reinforced AB baseline trials (the opposite of an associative symmetry pattern). The present experiment replicated this finding. In addition, it showed that antisymmetry also emerged after baseline training on successive AB symbolic matching, AA identity, and BB oddity, consistent with the prediction from Urcuioli's (2008) theory of pigeons' stimulus-class formation. Together, these results provide further empirical support for that theory including the proposition that the functional stimuli in pigeons' successive matching consist of the nominal stimuli plus their ordinal positions within a trial.
Keywords: antisymmetry, emergent relations, stimulus classes, successive matching, pigeons, key peck
In the conditional discrimination and equivalence literature, associative symmetry refers to the finding that after symbolic matching training in which subjects learn to match sample stimuli (A) to comparison stimuli (B) that bear no physical resemblance to one another (AB matching), they can then do the reverse (BA matching) without any explicit training to do so (Lionello-DeNolf, 2009). Associative symmetry (or “symmetry” for short) is thus an emergent relation, and it indicates that conditional discrimination training has yielded more than just a specific set of learned “if, then” relations (e.g., “If sample A1, then respond to comparison B1”). In particular, symmetry suggests that training has yielded a class of stimuli in which samples and comparisons are interchangeable with one another (Sidman & Tailby, 1982). As a result, subjects now exhibit new performances based on relations derived from those they explicitly learned—for example, by matching sample B1 to comparison A1.
Although humans often exhibit symmetry (e.g., Fields, Arntzen, Nartey, & Eilifsen, 2012; Sidman, Rauzin, Lazar, Cunningham, Tailby, & Carrigan, 1982, Experiment 3; Tomanari, Sidman, Rubio, & Dube, 2006), pigeons and other nonhuman animals frequently do not if they are trained and tested in two-alternative choice matching-to-sample (D'Amato, Salmon, Loukas, & Tomie, 1985; Dugdale & Lowe, 2000; Lionello-DeNolf & Urcuioli, 2002; Lipkens, Kop, & Matthijs, 1988; Sidman et al., 1982; Urcuioli, 2008, Experiments 1 and 2; although see Schusterman & Kastak, 1993, and Velasco, Huziwara, Machado, & Tomanari, 2010). One explanation for the pigeon findings is that they are sensitive to the location at which the samples and comparisons appear, so the location “swap” that accompanies a symmetry test (viz., center-key sample stimuli now appear as side-key comparison alternatives, and vice versa) is disruptive. More specifically, for them, a stimulus at one location is functionally different from the same nominal stimulus appearing at a different location. For example, Lionello and Urcuioli (1998) demonstrated that pigeons that have learned to match A center-key samples to A side-key comparisons (identity matching) are unable to continue to match accurately when those same samples later appear on a side key and the comparisons appear on the remaining keys (see also Iversen, 1997; Iversen, Sidman, & Carrigan, 1986; Urcuioli, 2007).
Controlling for the effects of stimulus location, however, does not by itself yield symmetry (Lionello-DeNolf & Urcuioli, 2002; Richards, 1988; see also Frank, 2007). One reason appears to be that in addition to location, pigeons “code” stimuli in terms of when they appear during a trial—that is, first (sample) or second (comparison). In other words, the functional matching stimuli also have a temporal or ordinal-position component. Thus, a red sample stimulus is properly described as red-in-the-first-ordinal-position and a reinforced triangle comparison stimulus is properly described as triangle-in-the-second-ordinal-position (ignoring for the moment their spatial locations). The symmetry test, however, involves triangle-in-the-first-ordinal-position (sample) and red-in-the-second-ordinal-position (comparison), differences vis-à-vis baseline training that are likely to impact test performances.
Interestingly, Frank and Wasserman (2005) and Urcuioli (2008, Experiment 3) found evidence for associative symmetry in pigeons after baseline training that controlled for both spatial location and ordinal position. Each study employed a successive or go/no-go matching-to-sample procedure (Wasserman, 1976) in which samples and comparisons appeared singly and sequentially on the same (e.g., center) response key, thus holding spatial location constant. To negate the possible disruptive effects of ordinal position in the shift from AB training to BA testing, symbolic (AB) matching was supplemented by concurrent training on AA and BB identity matching. In this arrangement, pigeons saw each A and B matching stimulus in both the first and second ordinal positions during training (i.e., both as a sample and as a comparison). Consequently, this avoided having to present these stimuli in novel ordinal positions (i.e., as novel samples or novel comparisons) during testing. The net result was that pigeons showed emergent BA matching (symmetry): They responded more to the comparisons on the reverse of the reinforced AB baseline trials than to the comparisons on the reverse of the nonreinforced AB baseline trials.
However, this associative symmetry effect does not appear to result from pigeons learning that ordinal position is irrelevant. For instance, Frank (2007) found no evidence for symmetry when pigeons were concurrently trained on AB, CA, and BD successive matching. Note that in this set of conditional relations, each A and B stimulus also appears both as a sample and as a comparison in training, yet BA matching does not emerge. Furthermore, Urcuioli (2008, Experiment 4) found that training pigeons on AB successive matching plus AA oddity and BB identity yields an “antisymmetry” effect in testing. In other words, pigeons respond more to the comparisons on the reverse of the nonreinforced AB baseline trials than on the reverse of the reinforced AB baseline trials. Clearly, these findings indicate that some other process besides “learning to ignore” ordinal position is at work. Indeed, the peculiar antisymmetry effect is predicted in part from an assumption in Urcuioli's (2008) theory of pigeons' stimulus-class formation stating that ordinal position within a matching trial is part of a stimulus' functional properties (i.e., that pigeons are sensitive to ordinal position just as they are to spatial location). The following paragraphs review this theory's assumptions and the predictions tested in the current experiment.
Urcuioli (2008) proposed that successive (go/no-go) matching facilitates the formation of stimulus classes because, besides holding spatial location constant, one half of all matching trials in each training session end in nonreinforcement independently of a pigeon's performance. In other words, each session contains an equal number of reinforced and nonreinforced sample–comparison combinations no matter whether pigeons respond nondifferentially to the comparisons (as they do at the start of training) or differentially (viz., preferentially) to the comparisons on reinforced trials (as training progresses). Second, the classes arising from such training consist of the elements of the reinforced combinations. Thus, if a red sample followed by a triangle comparison ends in reinforcement, a [red sample, triangle comparison] stimulus class will develop. Similarly, if a green sample followed by a horizontal-lines comparison ends in reinforcement, a [green sample, horizontal comparison] stimulus class will also develop. This segregation of stimuli into classes is ostensibly enhanced by the fact that the remaining sample–comparison combinations are repeatedly nonreinforced (Urcuioli, 2010). Thus, the red sample and horizontal-lines comparison are in different classes, as are the green sample and triangle comparison. Third, Urcuioli (2008) assumes that the functional stimuli are composed of each nominal stimulus plus its ordinal position within a matching trial (again ignoring spatial location). In essence, this assumption states that, for pigeons, a red sample (i.e., red-in-the-first-ordinal-position) is different from a red comparison (i.e., red-in-the-second-ordinal position). Finally, the theory proposes that elements common to more than one stimulus class cause their respective classes to merge, and that the composition of those merged classes determines what emergent relations, if any, will be observed.
Figures 1, 2 and 3 depict how these assumptions predict the antisymmetry effect reported by Urcuioli (2008, Experiment 4) and its attempted replication in the present experiment (see Group HO in Table 1). Baseline training involves symbolic (AB) successive matching with red and green samples (R1 and G1, respectively, where “1” = first ordinal position) and triangle and horizontal-lines comparisons (T2 and H2, respectively, where “2” = second ordinal position). The red sample – triangle comparison (R1→T2) and green sample – horizontal comparison (G1→H2) combinations are reinforced; the remaining combinations are not. Concurrent training on hue (AA) oddity and form (BB) identity using the same stimuli is also conducted. For hue oddity, responding to a hue comparison is reinforced only if it differs from its preceding sample (viz., R1→G2 and G1→R2). For form identity, responding to a form comparison is reinforced only if it matches its preceding sample (viz., T1→T2 and H1→H2).
Fig. 1.
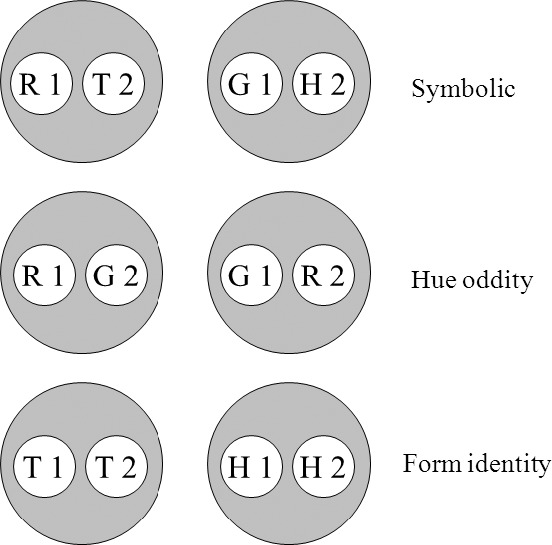
The six stimulus classes hypothesized to result from hue–form symbolic, hue oddity and form identity successive matching training (Group HO). R = red, G = green, T = triangle, H = horizontal, 1 = first ordinal position (sample stimulus), 2 = second ordinal position (comparison stimulus). Copyright 2008 by the Society for the Experimental Analysis of Behavior, Inc.
Fig. 2.
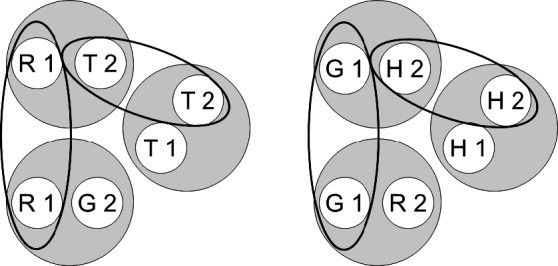
A rearrangement of the six stimulus classes shown in Figure 1 in order to highlight common class elements (via the ellipses). R = red, G = green, T = triangle, H = horizontal, 1 = first ordinal position (sample stimulus), 2 = second ordinal position (comparison stimulus). Copyright 2008 by the Society for the Experimental Analysis of Behavior, Inc.
Fig. 3.
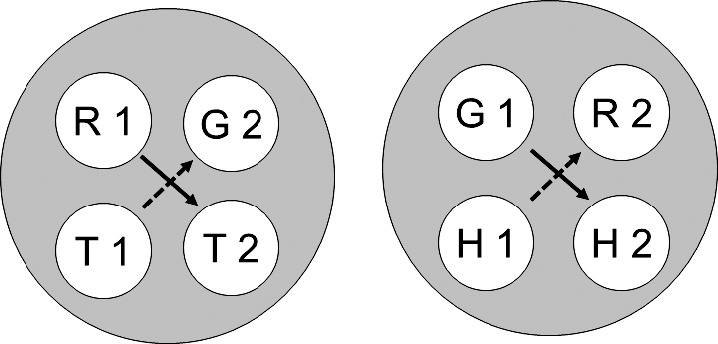
The two 4-member stimulus classes hypothesized to result from class merger via common elements after training on hue–form symbolic, hue oddity, and form identity successive matching (cf. Figure 2). Solid and broken arrows denote explicitly trained and predicted emergent symbolic relations, respectively. R = red, G = green, T = triangle, H = horizontal, 1 = first ordinal position (sample stimulus), 2 = second ordinal position (comparison stimulus). Copyright 2008 by the Society for the Experimental Analysis of Behavior, Inc.
Table 1.
Successive Matching Training Contingencies for the Two Groups

|
Hue-Form (AB) Matching |
Hue-Hue (AA) Oddity |
Form-Form (BB) Identity |
| Group HO | ||
| R → T - FI 5 s | R → R - EXT | T → T - FI 5 s |
| R → H - EXT | R → G - FI 5 s | T → H - EXT |
| G → T - EXT | G → R - FI 5 s | H → T - EXT |
| G → H - FI 5 s | G → G - EXT | H → H - FI 5 s |
| Hue-Form (AB) Matching | Hue-Hue (AA) Identity | Form-Form (BB) Oddity |
| Group FO | ||
| R → T - FI 5 s | R → R - FI 5 s | T → T - EXT |
| R → H - EXT | R → G - EXT | T → H - FI 5 s |
| G → T - EXT | G → R - EXT | H → T - FI 5 s |
| G → H - FI 5 s | G → G - FI 5 s | H → H - EXT |
Note. R = red, G = green, T = triangle, H = horizontal lines, FI = fixed interval schedule, EXT = nonreinforced. The first stimulus in the trial sequence (the sample) is shown to the left of the arrows, and the second stimulus (the comparison) is shown to the right. Counterbalancing of the hue–form matching contingencies has been omitted.
Figure 1 (cf. Figure 11 in Urcuioli, 2008) shows the two stimulus classes hypothesized to arise from each of the three successive matching tasks just described. Each class contains a sample and a comparison from one reinforced combination (e.g., the red sample [R1] and triangle comparison [T2] in symbolic matching). Figure 2 (cf. Figure 12 in Urcuioli, 2008) rearranges the six classes of Figure 1 in order to highlight (via the ellipses) elements common to more than one class. Lastly, Figure 3 (cf. Figure 13 in Urcuioli, 2008) shows the two 4-member classes hypothesized to result from the merging of classes sharing common elements. The solid arrows indicate the explicitly reinforced symbolic (AB) matching combinations, and the broken arrows indicate the predicted emergent BA relations. Note that the emergent relations (viz., T1→G2 and H1→R2) are not the symmetrical versions of the reinforced baseline relations. Instead, they are the symmetrical versions of the nonreinforced baseline relations. In other words, they represent “antisymmetry”, the effect reported by Urcuioli (2008, Experiment 4).
The present experiment was designed in part to provide a direct replication of this unusual but theoretically important finding. Consequently, one group of pigeons (Group HO in Table 1) was concurrently trained on AB symbolic matching, AA oddity, and BB identity and then tested on BA relations to see if the BA results would reproduce those previously obtained. The second purpose was to test another prediction from Urcuioli's (2008) theory that antisymmetry should also emerge in testing if baseline training involves form (rather than hue) oddity. In other words, higher comparison response rates on the reverse of the nonreinforced AB baseline trials should also be obtained after concurrent training on AB symbolic matching, AA identity, and BB oddity (see Group FO in Table 1).
Figures 4, 5 and 6 visually illustrate the derivation of this latter prediction. Figure 4 shows the six individual stimulus classes hypothesized to arise from baseline successive matching training. The classes shown in this figure resemble those in Figure 1 except that 1) the AA (hue identity) classes contain the elements of the matching sample–comparison combinations (R1 and R2, and G1 and G2), and 2) the BB (form oddity) classes contain the elements of the mismatching sample–comparison combinations (H1 and T2, and T1 and H2). Figure 5 rearranges these six classes to highlight (via the ellipses) common across-class elements, and Figure 6 shows the two 4-member classes hypothesized to result from the resulting class mergers. Note the antisymmetry prediction: Although symbolic (AB) training consists of explicitly reinforced R1→T2 and G1→H2 relations (solid arrows), testing should reveal more frequent responding on the reverse of the nonreinforced baseline relations (viz. to the comparisons on the H1→R2 and T1→G2 test trials; broken arrows). If this effect also materializes, it would extend the conditions from which antisymmetry results and provide additional support for Urcuioli's (2008) theory.
Fig. 4.
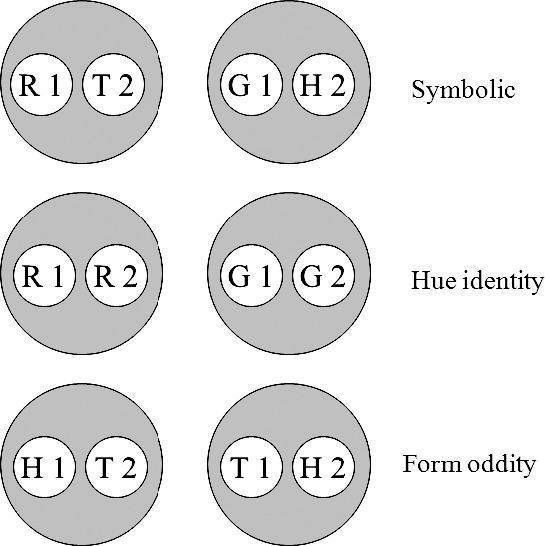
The six stimulus classes hypothesized to result from hue–form symbolic, hue identity and form oddity successive matching training (Group FO). R = red, G = green, T = triangle, H = horizontal, 1 = first ordinal position (sample stimulus), 2 = second ordinal position (comparison stimulus).
Fig. 5.
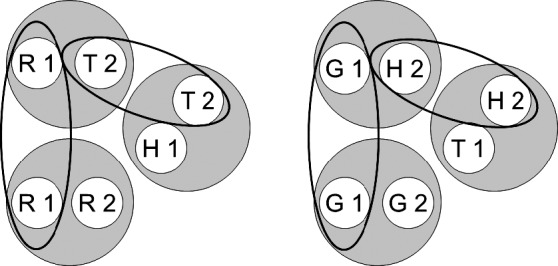
A rearrangement of the six stimulus classes shown in Figure 3 in order to highlight common class elements (via the ellipses). R = red, G = green, T = triangle, H = horizontal, 1 = first ordinal position (sample stimulus), 2 = second ordinal position (comparison stimulus).
Fig. 6.
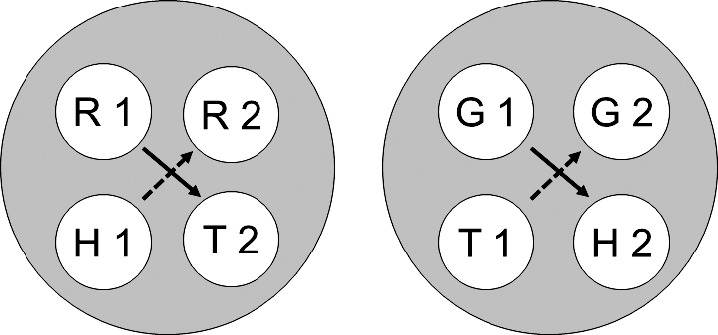
The two 4-member stimulus classes hypothesized to result from class merger via common elements after training on hue–form symbolic, hue identity, and form oddity successive matching (cf. Figure 5). Solid and broken arrows denote explicitly trained and predicted emergent symbolic relations, respectively. R = red, G = green, T = triangle, H = horizontal, 1 = first ordinal position (sample stimulus), 2 = second ordinal position (comparison stimulus).
METHOD
Subjects
Eight experimentally naive White Carneau pigeons from the Double “T” Farm (Glenwood, IA) were used. They were approximately 1–2 yrs old and were housed individually in stainless-steel wire-mesh cages in a colony room on a 14h-10h light–dark cycle (lights on at 07:00). Free-feeding weights were established immediately upon arrival in the laboratory by providing unrestricted access to Purina ProGrains over a period of 7-21 days. Prior to experimental participation, pigeons were gradually reduced to 80% of their free-feeding weights and were maintained at that level during the experiment by confining food access to the experimental sessions. Home-cage feedings were provided on days the experiment was not run and when pigeons did not obtain sufficient food in a session to maintain their 80% body weights. Water and grit were always available in the home cage. Pigeons were randomly divided into two groups of 4 (Groups HO and FO) prior to the experiment, and an equal number from each group were assigned to be run in each experimental chamber (see below).
Apparatus
Two pigeon operant chambers consisting of Model PIP-016 three-key panels inside Model SEC-002 enclosures (BRS/LVE, Laurel MD) were used. Only the center key of each chamber was active during the experiment. A stimulus projector mounted behind the center key could display a solid inverted white triangle, three white horizontal lines, and three small diagonally oriented white dots all on black backgrounds, and red and green homogeneous fields (BRS/LVE Pattern 692). The house light (GE No. 1829 bulb) was located 7.6 cm above the 2.5-cm-diameter center key and its light was directed toward the ceiling by a metal housing partially covering the bulb. A 5.8 cm x 5.8 cm opening directly below the center key provided access to a rear-mounted food hopper which, when raised, was illuminated by a miniature bulb (ESB-28). A continuously running blower fan attached to the outside of each chamber provided ventilation and masking noise. IBM-compatible computers controlled and recorded all experimental events.
Procedure
Preliminary training
After each pigeon learned to eat quickly from a periodically raised and lit food hopper, the method of successive approximations was used to shape pecking to the white dots on the center key. This was followed by one 60-trial session in which single pecks to a center-key triangle and horizontal lines were reinforced, and one 60-trial session in which single pecks to red and green on the center key were reinforced. Each form or hue stimulus appeared an equal number of times in these sessions with successive trials separated by a 10-s intertrial interval (ITI). The house light remained on throughout each session, and reinforcement consisted of 3-s access to grain.
Next, pigeons were trained over the course of ten 60-trial sessions to obtain food by pecking each stimulus on fixed-interval (FI) schedules. The triangle and horizontal lines, and red and green hues, appeared equally often on the center key during the first and second block of five sessions, respectively. The FI value was 2 s for the first session in each block, 3 s for the next two sessions, and 5 s for the last two sessions. Successive trials were separated by a 15-s ITI, the first 14 s of which was dark. The house light was turned on the last 1 s of the ITI and remained on until the end of the reinforcement cycle. Reinforcement duration was constant within a session for a given pigeon but could vary between 1.8 and 6.0 s across sessions in a manner that maintained 80% body weights.
Successive matching acquisition
Table 1 summarizes the successive matching training contingencies for each group. Both were trained on hue–form (AB) successive matching with red and green samples and triangle and horizontal-lines comparisons. For half of the pigeons in each group, reinforcement was contingent on responding to the triangle (T) comparison on red (R)-sample trials (R→T) and on responding to the horizontal-lines (H) comparison on green (G)-sample trials (G→H). The remaining hue sample – form comparison combinations (viz., R→H and G→T) ended without reinforcement. For the other half of the pigeons in each group, the opposite contingencies were in effect.
The groups differed from one another in the two other successive matching tasks on which they were concurrently trained. The Group HO (hue oddity) pigeons learned hue–hue (AA) oddity and form–form (BB) identity, whereas the Group FO (form oddity) pigeons learned form–form (BB) oddity and hue–hue (AA) identity. For the identity matching tasks, reinforcement was contingent upon responding to comparisons that were identical to their preceding samples (i.e., on T→T and H→H trials or on R→R and G→G trials), whereas mismatching sample–comparison combinations ended without reinforcement. For the oddity tasks, reinforcement was contingent upon responding to comparisons that did not match their preceding samples (i.e., on R→G and G→R trials or on T→H and H→T trials), whereas matching sample–comparison combinations ended without reinforcement.
Each 96-trial training session consisted of 32 trials of each successive matching task, with trials equally divided among the four possible sample–comparison combinations of each task. Trials were randomized with the constraint that no combination could occur more than three times in a row. The 5-s sample stimulus duration began with the first peck to the sample and ended with the first peck after 5 s. Sample offset coincided with a blank 1-s interstimulus interval after which a single comparison on the same (center key) location appeared. On reinforced trials, the first comparison–stimulus peck after 5 s turned off the comparison and produced food; on nonreinforced trials, the comparison went off automatically after 5 s. [Note that an FI schedule of reinforcement has been successfully used to study emergent relations in pigeons in several studies reported by this laboratory (e.g., Sweeney & Urcuioli, 2010; Urcuioli, 2008; Urcuioli & Swisher, 2012) and by others (Frank & Wasserman, 2005)]. The house light was turned off after the end of the reinforcement cycle or at comparison offset on nonreinforced trials. The next matching trial began after a 15-s ITI, the first 14 s of which was spent in darkness. The house light was turned on 1 s prior to sample presentation. Once again, the duration of food access on reinforced trials was constant within a session but could vary across sessions and across pigeons in such a way that 80% body weights were maintained.
Pigeons were trained on their respective successive matching tasks until they achieved a discrimination ratio (DR) of at least .80 on each of the three tasks for five of six consecutive sessions (“criterion”). A minimum of 10 overtraining sessions (to help ensure stable performances) then followed and ended when criterion was maintained or was again met. The DRs were calculated by dividing the total number of comparison pecks on reinforced trials by the total number of comparison pecks on both reinforced and nonreinforced trials. Only pecks occurring within 5 s of comparison onset entered into these calculations.
One pigeon in each group (HO1 and FO3) had difficulty meeting the acquisition criterion but was nonetheless moved into overtraining after 145 and 133 sessions, respectively, because the performances on their respective successive matching tasks were consistently close to and often exceeded a .80 DR. For example, pigeon HO1's DRs for hue–form matching, hue–hue oddity, and form–form identity averaged over the last five sessions prior to overtraining were .82, .79, and .79, respectively. This pigeon then received 30 overtraining sessions; its DRs averaged over the last five of those sessions were .77, .85, and .79, respectively. For pigeon FO3, its DRs for hue–form matching, hue–hue identity, and form–form oddity over the last five acquisition sessions were .82, .86, and .78, respectively. It then received 13 overtraining sessions and was moved into testing (see below) given that the corresponding DRs for the last five of those sessions were .83, .83, and .80, respectively. Finally, the baseline performances of 1 pigeon (HO4) that met criterion rather quickly in acquisition inexplicably became unstable during overtraining. It was finally moved into the test phase after over 300 overtraining sessions; its DRs for hue–form matching, hue–hue oddity, and form–form identity for the last five of those sessions were .75, .84, and .78, respectively.
Successive matching testing
Testing for possible emergent form–hue (BA) relations began following overtraining. A total of eight test sessions were run in blocks of two sessions, with each block separated by a minimum of five baseline training sessions (see Table 1) until criterion performance (viz., DRs ≥ 0.80 for all three baseline tasks for five of six consecutive sessions) was met. Each 104-trial test session consisted of 96 baseline trials, 32 with each successive matching training task, and eight nonreinforced probe trials on which the sample was either the triangle or horizontal lines and the comparison was either red or green. The comparison on these trials went off automatically after 5 s without food. The four probe-trial combinations (viz., T→R, T→G, H→R, and H→G) occurred equally often in each test session. They were distributed randomly throughout a test session with the restrictions that all of the 12 possible baseline trials occur at least once before the first probe trial and that at least six baseline trials separate successive probe trials. The dependent variable of interest was the number of comparison responses occurring on the reverse of the reinforced or “positive” AB baseline trials and on the reverse of the nonreinforced or “negative” AB baseline trials.
RESULTS
Analysis of variance (ANOVA) and, where appropriate, post-hoc contrasts were used to analyze group data and individual subject data. Statistical results were compared to the tabled F values reported by Rodger (1975) to control Type I error on a per-decision basis.
Acquisition and baseline performances
Although most pigeons met the acquisition criterion of a .80 DR or higher on all three baseline successive matching tasks, 2 did not, although their performances generally met or exceeded .80 on one or two tasks at different times during acquisition. For Group HO, the average numbers of sessions to criterion levels of performance on the three baseline tasks were 31.8, 49.0 and 36.0 for hue–form symbolic, hue oddity, and form identity matching, respectively. The numerical difference was not statistically significant, F(2, 6) = .53. For Group FO, the average sessions-to-criterion were 33.0, 62.5, and 29.8 for hue–form symbolic, form oddity, and hue identity matching, F(2, 6) = 6.55. The pigeons in this group were slowest to acquire form oddity relative to the other two tasks, F(2, 6) = 6.49. Performances over the last five overtraining sessions by the Group FO pigeons mirrored the acquisition differences: The average DR for form oddity (.85) was significantly lower than for hue–form symbolic (.88) and hue identity matching (.90), F(2, 6) = 10.26. Nevertheless, all baseline DRs were above the criterion level. The corresponding DRs for Group HO showed no significant between-task differences: .84, .88, and .85 for hue-form symbolic, hue oddity, and form identity matching, F(2, 6) = 1.74.
Most baseline DRs during the test sessions themselves was at or above .80 in both groups. There were only two instances in which a DR fell below .70, and most sub-.80 DRs were in the .75-.79 range.
Test performances
Figure 7 plots the number of comparison pecks/sec for Group HO on the hue–form symbolic baseline trials (open circles) and the nonreinforced form–hue probe trials (filled circles) averaged over the eight test sessions. Figure 8 plots the corresponding results from Group FO. The probe data are the averages of the four test trials in each test session in which the reinforced baseline combinations were reversed (“positive”) and the four test trials in each test session in which the nonreinforced baseline combinations were reversed (“negative”). The baseline data represent the average numbers of pecks for four randomly selected “positive” (reinforced) and four randomly selected “negative” (nonreinforced) trials (two of each positive and negative hue–form combination) from each test session. Only four randomly selected baseline trials of each type from each session entered into the calculations in order to maintain comparability with the numbers of positive and negative probe trials per test session.
Fig. 7.

Comparison–response rates in pecks/sec (± 1 SEM) on the symbolic matching baseline trials (open circles) and the nonreinforced symmetry probe trials (filled circles) averaged over the eight test sessions for each Group HO pigeon. Positive = reinforced symbolic baseline trials and test trials in which the samples and comparisons of the reinforced baseline trials were reversed. Negative = nonreinforced symbolic baseline trials and test trials on which the samples and comparisons of the nonreinforced baseline trials were reversed.
Fig. 8.

Comparison–response rates in pecks/sec (± 1 SEM) on the symbolic matching baseline trials (open circles) and the nonreinforced symmetry probe trials (filled circles) averaged over the eight test sessions for each Group FO pigeon. Positive = reinforced symbolic baseline trials and test trials in which the samples and comparisons of the reinforced baseline trials were reversed. Negative = nonreinforced symbolic baseline trials and test trials on which the samples and comparisons of the nonreinforced baseline trials were reversed.
Throughout testing, the hue–form baseline discriminations remained intact for all pigeons: They pecked at much higher rates to the comparison stimuli on positive than on negative trials. By contrast, the probe-trial test results mostly showed the opposite pattern. For Group HO (whose baseline training included hue oddity), comparison–response rates by all 4 pigeons were numerically higher on the negative than positive probe trials. In other words, they pecked more frequently to the comparisons on the reverse of the nonreinforced hue–form baseline trials than on the reverse of the reinforced hue–form baseline trials. For 2 pigeons (HO1 and HO4), this difference in probe-trial rates was significant, Fs(1, 62) = 52.26 and 13.94, respectively; for the other 2 (HO2 and HO3), it was not, Fs(1, 62) = .57 and .16, respectively. For Group FO (whose baseline training included form oddity), 3 pigeons (FO1, FO3, and FO4) pecked the probe-trial comparisons more frequently on the reverse of the negative than the positive baseline trials, and the difference was significant for all 3, Fs(1, 62) = 11.99, 12.06, and 12.66, respectively. The remaining Group FO pigeon (FO2) pecked the comparisons more frequently on positive than negative probe trials, but this difference was not significant, F(1, 62) = 2.50.
DISCUSSION
This experiment accomplished the two aims for which it was designed. First, it replicated in 2 of 4 pigeons the theoretically important antisymmetry effect originally reported by Urcuioli (2008, Experiment 4). Baseline training on AB symbolic matching along with AA oddity and BB identity yielded an emergent BA effect in which pigeons responded more to the comparisons on the reverse of the nonreinforced (negative) than reinforced (positive) AB trials. Second, it demonstrated this same pattern of test results when baseline training involved concurrent AB matching, AA identity, and BB oddity. This emergent antisymmetry effect is also predicted by Urcuioli's (2008) theory of pigeons' stimulus-class formation.
One of the key assumptions of that theory is that the functional stimuli in successive matching are compounds consisting of the nominal stimuli plus their ordinal position within a trial. In fact, another component is surely the location at which the nominal stimuli appear (Lionello & Urcuioli, 1998). However, this potential variable is typically held constant in successive matching (e.g., Campos, Debert, Barros, & McIlvane, 2011; Debert, Matos, & McIlvane, 2007; Frank & Wasserman, 2005; Nelson & Wasserman, 1978; Strasser, Ehrlinger, & Bingman, 2004; see also Cullinan, Barnes, & Smeets, 1998 and Zentall & Hogan, 1975), so theoretical derivations of possible emergent relations after such constant-location training can safely ignore where the matching stimuli appear.
It would not be possible to derive the antisymmetry prediction without taking into account each stimulus' ordinal position. Indeed, if all of the assumptions of Urcuioli's (2008) theory were retained except for the one about the nature of the functional stimuli (i.e., their ordinal position component), no emergent effect of any kind would be predicted. The reason is that the class-merger-via-common-elements assumption leads to the prediction that baseline training on either set of successive matching tasks would result in a single stimulus class containing all four nominal matching stimuli (viz., R, G, T, and H). Consequently, the novel T→R, T→G, H→R, and H→G test sequences should yield comparable rates of comparison responding. Alternatively, if the only functions of the two baseline tasks trained concurrently with AB symbolic matching in Group HO and Group FO were 1) to familiarize pigeons with seeing each stimulus both as a sample and as a comparison, and 2) to insure the requisite sample and comparison discriminations (Saunders & Green, 1999) prior to testing, then symmetry would be predicted: Pigeons should subsequently respond more frequently to the comparisons on the reverse of the positive (reinforced) than negative (nonreinforced) AB trials (Frank, 2007; Frank & Wasserman, 2005). Clearly, this was not the case—the opposite pattern of results occurred.
Why, then, did some pigeons not show the predicted antisymmetry effect when one of their baseline tasks was hue oddity or form oddity? It could be that, for them, the functional stimuli did not include an ordinal position component. Once again, if the nominal stimuli themselves were the functional stimuli and the baseline relations gave rise to new derived relations, the probable outcome would be associative symmetry. Instead, these pigeons mostly responded non-differentially on the positive and negative test trials. Apparently, for them, baseline training did not yield the individual stimulus classes shown in Figures 1 and 4 or the individual classes did not merge despite having elements in common. In either case, no emergent effect would be predicted.
The results from this experiment are also important in the context of some failures of Urcuioli's (2008) theory to predict recently published data. For instance, although Sweeney and Urcuioli (2010) reported that pigeons trained on AB symbolic, BA symbolic, and BB identity matching showed the predicted emergent AA matching, other pigeons trained on the same two symbolic tasks and BB oddity did not exhibit the predicted emergent AA oddity relations. The latter is akin to the antisymmetry prediction confirmed here and it, too, relies on the theoretical assumption regarding ordinal position. Likewise, Urcuioli and Swisher (2012) recently reported that some pigeons trained solely on AB and BA symbolic matching showed emergent AA matching in testing. A stimulus class analysis like that portrayed in Figures 1–3 (and Figures 4–6) does not predict this result. There may be other factors at work responsible for the former failure-to-confirm and the latter theoretical disconfirmation (viz., a possible identity bias—Hogan & Zentall, 1981). Consequently, it is important to provide additional data that bear directly on the functional-stimulus assumption. That was done here.
Urcuioli (2008, Experiment 3) trained AB symbolic matching along with AA and BB identity and found, like Frank and Wasserman (2005), associative symmetry in testing. Here, we have reported that training on AB symbolic matching plus one identity and one oddity task using the A and B stimuli (see also Urcuioli, 2008, Experiment 4) yields emergent antisymmetry. A manipulation which would “round out” the four ways to supplement AB symbolic successive matching is to concurrently train both AA oddity and BB oddity. The theoretical prediction (that awaits testing) is that pigeons trained in this fashion will exhibit emergent BA symmetry. Such a finding would lend additional support to the assumption that the functional stimuli in pigeons' successive matching consist not only of their physical features (e.g., their color or form) but also their ordinal positions within a trial; that is, whether they appear first (as a sample) or second (as a comparison).
Footnotes
This research was supported by NICHD Grant R01 HD061322. The authors thank Heloisa Campos, Greg Erhardt, Lisa Macklin, and Blake Polak for their assistance in conducting this research.
REFERENCES
- Campos H. C., Debert P., Barros R., McIlvane W. J. Relational discriminations by pigeons in a go/no-go procedure with compound stimuli: A methodological note. Journal of the Experimental Analysis of Behavior. (2011);96:417–426. doi: 10.1901/jeab.2011.96-413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cullinan V. A., Barnes D., Smeets P. M. A precursor to the relational evaluation procedure: Analyzing stimulus equivalence. The Psychological Record. (1998);48:121–145. [Google Scholar]
- D'Amato M. R., Salmon D. P., Loukas E., Tomie A. Symmetry and transitivity in the conditional relations in monkeys (Cebus apella) and pigeons (Columba livia) Journal of the Experimental Analysis of Behavior. (1985);44:35–47. doi: 10.1901/jeab.1985.44-35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Debert P., Matos M. A., McIlvane W. Conditional relations with compound abstract stimuli using a go/no-go procedure. Journal of the Experimental Analysis of Behavior. (2007);87:89–96. doi: 10.1901/jeab.2007.46-05. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dugdale N., Lowe C. F. Testing for symmetry in the conditional discriminations of language-trained chimpanzees. Journal of the Experimental Analysis of Behavior. (2000);73:5–22. doi: 10.1901/jeab.2000.73-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fields L., Arntzen E., Nartey R. K., Eilifsen C. Effects of a meaningful, a discriminative, and a meaningless stimulus on equivalence class formation. Journal of the Experimental Analysis of Behavior. (2012);97:163–181. doi: 10.1901/jeab.2012.97-163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frank A. J. An examination of the temporal and spatial stimulus control in emergent symmetry in pigeons. Unpublished doctoral dissertation; University of Iowa: (2007). [Google Scholar]
- Frank A. J., Wasserman E. A. Associative symmetry in the pigeon after successive matching-to-sample training. Journal of the Experimental Analysis of Behavior. (2005);84:147–165. doi: 10.1901/jeab.2005.115-04. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hogan D. E., Zentall T. R. The role of identity in the learning and memory of a matching-to-sample problem by pigeons. Bird Behaviour. (1981);3:27–36. [Google Scholar]
- Iversen I. Matching-to-sample performance in rats: A case of mistaken identity? Journal of the Experimental Analysis of Behavior. (1997);68:27–47. doi: 10.1901/jeab.1997.68-27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iversen I., Sidman M., Carrigan P. Stimulus definition in conditional discriminations. Journal of the Experimental Analysis of Behavior. (1986);45:297–304. doi: 10.1901/jeab.1986.45-297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lionello K. M., Urcuioli P. J. Control by sample location in pigeons' matching to sample. Journal of the Experimental Analysis of Behavior. (1998);70:235–251. doi: 10.1901/jeab.1998.70-235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lionello-DeNolf K. M. The search for symmetry: 25 years in review. Learning & Behavior. (2009);37:188–203. doi: 10.3758/LB.37.2.188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lionello-DeNolf K. M., Urcuioli P. J. Stimulus control topographies and tests of symmetry in pigeons. Journal of the Experimental Analysis of Behavior. (2002);78:467–495. doi: 10.1901/jeab.2002.78-467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lipkens R., Kop P. F. M., Matthijs W. A test of symmetry and transitivity in the conditional discrimination performances of pigeons. Journal of the Experimental Analysis of Behavior. (1988);49:395–409. doi: 10.1901/jeab.1988.49-395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nelson K. R., Wasserman E. A. Temporal factors influencing the pigeon's successive matching-to-sample performance: Sample duration, intertrial interval, and retention interval. Journal of the Experimental Analysis of Behavior. (1978);30:153–162. doi: 10.1901/jeab.1978.30-153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richards R. W. The question of bidirectional associations in pigeons' learning of conditional discrimination tasks. Bulletin of the Psychonomic Society. (1988);26:577–579. [Google Scholar]
- Rodger R. S. The number of non-zero, post hoc contrasts from ANOVA and error-rate. I. British Journal of Mathematical and Statistical Psychology. (1975);28:71–78. [Google Scholar]
- Saunders R. R., Green G. A discrimination analysis of training structure effects on stimulus equivalence outcomes. Journal of the Experimental Analysis of Behavior. (1999);72:117–137. doi: 10.1901/jeab.1999.72-117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schusterman R. J., Kastak D. A California sea lion (Zalophus Californianus) is capable of forming equivalence relations. The Psychological Record. (1993);43:823–839. [Google Scholar]
- Sidman M., Rauzin R., Lazar R., Cunningham S., Tailby W., Carrigan P. A search for symmetry in the conditional discrimination of rhesus monkeys, baboons, and children. Journal of the Experimental Analysis of Behavior. (1982);37:23–44. doi: 10.1901/jeab.1982.37-23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sidman M., Tailby W. Conditional discrimination vs. matching to sample: An expansion of the testing paradigm. Journal of the Experimental Analysis of Behavior. (1982);37:5–22. doi: 10.1901/jeab.1982.37-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strasser R., Ehrlinger J. M., Bingman V. P. Transitive behavior in hippocampal-lesioned pigeons. Brain, Behavior, and Evolution. (2004);63:181–188. doi: 10.1159/000076442. [DOI] [PubMed] [Google Scholar]
- Sweeney M. M., Urcuioli P. J. A reflexivity effect in pigeons. Journal of the Experimental Analysis of Behavior. (2010);94:267–282. doi: 10.1901/jeab.2010.94-267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tomanari G. Y., Sidman M., Rubio A. R., Dube W. V. Equivalence classes with requirements for short response latencies. Journal of the Experimental Analysis of Behavior. (2006);85:349–369. doi: 10.1901/jeab.2006.107-04. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Urcuioli P. J. Sample and comparison location as factors in matching acquisition, transfer, and acquired equivalence. Learning & Behavior. (2007);35:252–261. doi: 10.3758/bf03206431. [DOI] [PubMed] [Google Scholar]
- Urcuioli P. J. Associative symmetry, antisymmetry, and a theory of pigeons' equivalence-class formation. Journal of the Experimental Analysis of Behavior. (2008);90:257–282. doi: 10.1901/jeab.2008.90-257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Urcuioli P. J. Associative symmetry and stimulus-class formation by pigeons: The role of nonreinforced baseline relations. Behavioural Processes. (2010);85:226–235. doi: 10.1016/j.beproc.2010.07.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Urcuioli P. J., Swisher M. Emergent identity matching after successive matching training. II: Reflexivity or transitivity? Journal of the Experimental Analysis of Behavior. (2012);97:5–27. doi: 10.1901/jeab.2012.97-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Velasco S. M., Huziwara E. M., Machado A., Tomanari G. Y. Associative symmetry by pigeons after few-exemplar training. Journal of the Experimental Analysis of Behavior. (2010);94:283–295. doi: 10.1901/jeab.2010.94-283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wasserman E. A. Successive matching-to-sample in the pigeon: Variation on a theme by Konorski. Behavior Research Methods & Instrumentation. (1976);8:278–282. [Google Scholar]
- Zentall T. R., Hogan D. E. Concept learning in the pigeon: Transfer to new matching and nonmatching stimuli. American Journal of Psychology. (1975);88:233–244. [Google Scholar]