

Synopsis
The development of sedative/hypnotic molecules has been empiric rather than rational. The empiric approach has produced clinically useful drugs but for no drug is the mechanism of action completely understood. All available sedative/hypnotic medications have unwanted side effects and none of these medications creates a sleep architecture that is identical to the architecture of naturally occurring sleep. This chapter reviews recent advances in research aiming to elucidate the neurochemical mechanisms regulating sleep and wakefulness. One promise of rational drug design is that understanding the mechanisms of sedative/hypnotic action will significantly enhance drug safety and efficacy.
Keywords: Neurotransmitters, Receptors, Translational Research, Drug Development
Sleep states are comprised of a constellation of physiological and behavioral traits, and the mechanisms by which sedative/hypnotic medications alter these traits remain unclear. Drugs that enhance states of sleep also alter autonomic physiology, behavior, cognition, and affect. The complexities of brain neurochemistry and the extensive neural circuits regulating levels of behavioral arousal contribute to the present inability to understand exactly how sedative/hypnotics promote sleep. An additional complexity is that many sedative/hypnotic drugs have behavioral state-specific actions. For example, some sedative/hypnotic drugs promote the non-rapid eye movement (NREM) phase of sleep at the expense of decreasing the rapid eye movement (REM) phase of sleep. In spite of the foregoing limitations, there has been progress in developing sleep medications that maximize desired actions such as rapid sleep onset, minimal next day effect, low or no abuse potential, and creation of a drug-induced state that is indistinguishable from physiological sleep. To date, however, no sedative/hypnotic produces all of these desired effects.
Rational drug design is an approach that has been successful in the development of antibiotic medications. Rational drug development of sedative/hypnotic medications is an approach based on understanding the receptor-binding properties of a molecule and how a molecule alters ligand binding, neurotransmitter synthesis, release, reuptake, and degradation. All of the foregoing cellular mechanisms can then be interpreted in the context of the overall drug effect. For sedative/hypnotic medications the desired action is, of course, promoting a safe and restorative sleep-like state. This chapter and Figure 1 provide an overview of neurotransmitters and brain regions currently known to modulate states of sleep and wakefulness. This overview of sleep neuropharmacology is an update of a précis1 of a book chapter2 and interested readers are referred elsewhere for detailed reviews on sleep.3–10
Figure 1. Brain regions modulating sleep and wakefulness.

Sagittal drawing of the rat brain (modified from222) schematizes the location, shape, and size of some brain regions that regulate sleep and wakefulness. The name of each brain region appears in bold print, the major neurotransmitters used for signaling to other brain regions are in parentheses, and neurochemical analytes relevant for arousal-state control that have been measured in that brain region are listed under the header “Quantified”. The microdialysis probe is drawn to scale and is shown sampling from the prefrontal cortex. Abbreviations: XII – hypoglossal nucleus; BF – basal forebrain; DRN – dorsal raphé nucleus; LC – locus coeruleus; LDT – laterodorsal tegmental nucleus; LH – lateral hypothalamus; MPO – medial preoptic area; PFC – prefrontal cortex; PPT – pedunculopontine tegmental nucleus; PnC – pontine reticular formation, caudal part; PnO – pontine reticular formation, oral part; TMN – tuberomamillary nucleus; TNC – trigeminal nucleus complex; VLPO – ventrolateral preoptic area; VTA – ventral tegmental area; 5HT – serotonin; ACh – acetylcholine; Ado – adenosine; Asp – aspartate; DA – dopamine; GABA – γ-aminobutyric acid; Glu – glutamate; Gly – glycine; His – histamine; Hcrt – hypocretin; NE – norepinephrine; NO – nitric oxide; Noc – nociceptin; Ser – serine; 5HT – serotonin; Tau – taurine. Figure reprinted from Watson et al., 20102 with permission.
γ-AMINOBUTYRIC ACID (GABA)
GABA is the major inhibitory neurotransmitter in the brain. Although GABA transporters11 and GABAB12 receptors can modulate sleep and wakefulness, most research into GABAergic regulation of behavioral arousal focuses on the GABAA receptor. Activation of GABAA receptors causes neuronal inhibition by increasing chloride ion conductance. Because of their powerful inhibitory effects, GABAA receptors are the targets of most sedative/hypnotic and general anesthetic drugs. GABAA receptors exist as multiple subtypes (reviewed in13) and these subtypes are differentially located throughout the brain (reviewed in14). The differences in clinical effects caused by various benzodiazepine (e.g., diazepam) and non-benzodiazepine (e.g., eszopiclone) sedative/hypnotics are attributed to the relative selectivity of these drugs for different GABAA receptor subtypes.14 The complexity imparted by the numerous GABAA receptor subtypes is humbling. Although there is detailed knowledge about the many subunit isoforms that comprise GABAA receptor subtypes,13 information is lacking about which of the many possible subtypes actually are expressed in specific brain regions,15–17 and which subtypes are localized synaptically verses extrasynaptically.18 Extrasynaptically localized GABAA receptors possess a delta subunit and have particular relevance for sleep medicine.19, 20
A better understanding of the in vivo characteristics and anatomical localization of GABAA receptor subtypes will contribute to rationale drug development. The preclinical studies described in this section illustrate the complexity of the problem and provide examples of how the effects of GABAergic drugs on behavior vary as a function of brain region. For example, although systemic administration of GABAmimetic drugs promotes sleep, sedation, or general anesthesia, enhancing GABAergic transmission with the pontine reticular formation actually increases wakefulness and decreases sleep. The pontine reticular formation is part of the ascending reticular activating system and contributes to the generation of REM sleep. Direct administration into the pontine reticular formation of drugs that increase GABAergic transmission increases wakefulness and inhibits sleep.21–24 Similarly, pharmacologically increasing the concentration of endogenous GABA within the pontine reticular formation increases the time required for isoflurane to induce general anesthesia.25 Consistent with this finding are data showing that endogenous GABA levels in the pontine reticular formation are greater during wakefulness than during REM sleep26, 27 (Figure 2) or during the loss of wakefulness caused by isoflurane.25 Inhibiting GABAergic signaling at GABAA receptors within the pontine reticular formation causes an increase in REM sleep and a decrease in wakefulness.22, 23, 28, 29 Likewise, decreasing extracellular GABA levels in the pontine reticular formation of rat decreases wakefulness and increases sleep,24 and shortens the time required for isoflurane to induce loss of consciousness.25 Furthermore, blocking GABAA receptors in the pontine reticular formation increases time needed to regain wakefulness after isoflurane anesthesia.22 Considered together, these data demonstrate a wakefulness-promoting role for GABA in the pontine reticular formation.
Figure 2. GABA levels in pontine reticular formation during wakefulness, NREM sleep, and REM sleep.
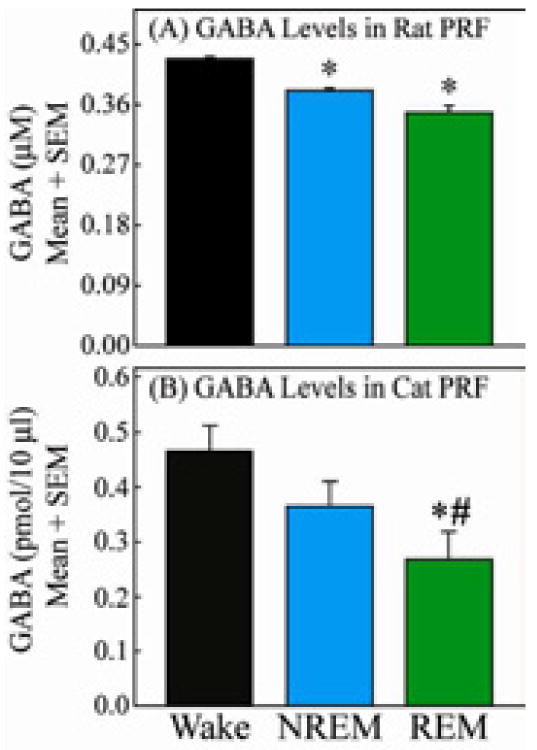
These comparative data illustrate two key points. First, that in both rat (A) and cat (B) there are parallel, state-dependent changes in GABA levels. In rat and cat GABA levels are significantly lower in REM sleep than during wakefulness. Second, methodological differences in the collection of GABA preclude direct comparison of GABA levels between these two species. GABA levels shown in A and B reflect differences in microdialysis flowrate (0.4 μL/min for rat and 2.0 μL/min for cat), molecular weight cut-off of the microdialysis probe membrane (18000 Daltons for rat and 6 Daltons for cat) and possibly membrane material (regenerated cellulose for rat and cuprophane for cat). Figures modified from Watson et al., 201127 and Vanini et al., 201126 with permission.
In brain regions containing neurons that promote wakefulness, GABAergic inhibition has been shown to cause an increase in sleep. These brain regions include the dorsal raphé nucleus (Fig. 1; DRN), tuberomamillary nucleus of the posterior hypothalamus (Fig. 1; TMN), medial preoptic area (Fig. 1; MPO), and ventrolateral periaqueductal gray30 (for reviews see8, 31, 32).
ACETYLCHOLINE
Acetylcholine is distinguished as being the first identified neurotransmitter. Although the first neurochemical theory of sleep33 correctly posited that acetylcholine plays a primary role in generating the brain-activated states of wakefulness and REM sleep, cholinergic drugs are not part of the standard pharmacological armamentarium of sleep disorders medicine. Nonetheless, understanding the mechanisms by which cholinergic neurotransmission generates and maintains REM sleep is crucial, because acetylcholine interacts with other transmitter systems that are targets of sleep pharmacotherapy (e.g., GABAergic and monoaminergic). Much of the research on the regulation of sleep by acetylcholine has focused on transmission mediated by muscarinic cholinergic receptors. Five subtypes (M1–M5) of the muscarinic receptor have been identified,34 and the M2 subtype plays a key role in the generation of REM sleep.35
Cholinergic signaling originating from the laterodorsal tegmental and pedunculopontine tegmental nuclei (LDT/PPT) and the basal forebrain (see Fig. 1) promotes the cortically activated states of wakefulness and REM sleep (reviewed in36). LDT/PPT neurons can be divided into two populations based on discharge pattern. One population discharges maximally during wakefulness and REM sleep (referred to as Wake-On/REM-On) and another population fires only during wakefulness (Wake-On/REM-Off) (reviewed in3). This finding helps explain how acetylcholine can promote both wakefulness and REM sleep. LDT/PPT neurons project to numerous wakefulness-promoting brain regions.3 Cholinergic terminals in the pontine reticular formation arise from the LDT/PPT,3 and muscarinic receptors are present in the pontine reticular formation.35, 37, 38 Many studies have administered cholinomimetics to the pontine reticular formation and have demonstrated that cholinergic transmission in the pontine reticular formation induces REM sleep (reviewed in3, 36). Electrically stimulating the LDT/PPT increases acetylcholine release in the pontine reticular formation39 and increases REM sleep.40 The release of endogenous acetylcholine in the pontine reticular formation is significantly greater during REM sleep than during wakefulness or NREM sleep.41–43 Taken together, these data demonstrate that cholinergic projections from the LDT/PPT to the pontine reticular formation promote REM sleep.
Recent in vivo data obtained from normal rats demonstrate that the sedative/hypnotics zolpidem, diazepam, and eszopiclone differentially alter acetylcholine release in the pontine reticular formation.44 Intravenous administration of eszopiclone prevented the REM phase of sleep, increased EEG delta power, and decreased acetylcholine release in rat pontine reticular formation (Figure 3).44 These data provide the first functional evidence for a heterogeneous distribution of GABAA receptor subtypes within the pontine reticular formation. The different effects of GABAA receptor agonists on sleep have been attributed to brain region-specific distributions of GABAA receptors and differences in sedative/hypnotic affinities for GABAA receptor subtypes.45 These preclinical data can be contrasted with human psychopharmacology where there has been no study convincingly demonstrating differential GABAA subtype binding among benzodiazepine and non-benzodiazepine sleeping medications.45 To date, the non-benzodiazepine, benzodiazepine-receptor agonist eszopiclone remains the only sleeping medication for which the long-term (6 months) effects have been characterized.46, 47
Figure 3. Intravenous administration of eszopiclone to intact, behaving rats decreases acetylcholine (ACh) release in the pontine reticular formation (PRF).

Top: schematic coronal section of rat brain stem illustrates placement of a microdialysis probe in the PRF. Ringer’s solution is pumped into the probe and samples are collected for quantification of ACh. Schematized at top right of brain are electrodes and an amplifier for recording the cortical electroencephalogram (EEG), and a representative trace showing EEG activity after intravenous administration of eszopiclone. Bottom: Histograms summarize the significant decrease in ACh release within the PRF caused by intravenous administration of eszopiclone. Data reprinted from Hambrecht-Wiedbusch et al., 201044 with permission.
Cholinergic neurons originating in the basal forebrain project throughout the entire cerebral cortex (reviewed in48). Acetylcholine release in the basal forebrain is highest during REM sleep, lower during quiet wakefulness, and lowest during NREM sleep.49 Cortical acetylcholine release is increased during wakefulness48, 50, 51 and REM sleep50 as compared to NREM sleep. These support the interpretation that cholinergic transmission from the basal forebrain promotes cortical activation during wakefulness and REM sleep.
ADENOSINE
Adenosine is a breakdown product of adenosine triphosphate (ATP). Increases in endogenous adenosine levels in a specific brain region during a period of prolonged wakefulness indicate that the region has been metabolically active. Direct biochemical measures show that ATP levels increase during sleep in areas of the brain that are most active during wakefulness.52 This finding provides direct support for the hypothesis that sleep serves a restorative function.53
Four subtypes of adenosine receptors, A1, A2A, A2B, and A3, have been identified and are distributed widely throughout the brain. Adenosine A1 and A2A receptors are antagonized by caffeine and the idea that adenosine promotes sleep is supported by the ubiquitous consumption of caffeine to maintain wakefulness and enhance alertness. In humans, oral administration of caffeine prior to nocturnal sleep increases sleep latency and reduces sleep efficiency.54 Furthermore, morning caffeine ingestion has been shown to decrease sleep efficiency and overall sleep during the subsequent night.55 No adenosine agonists are presently available to promote sleep. Adenosine, however, is relevant for sleep medicine, as insomnia can be caused by consumption of caffeine or by the respiratory stimulant theophylline. Interestingly, adenosine can have analgesic effects and this action shows promise for clinical use.56
Adenosinergic transmission in brain regions that regulate sleep and wakefulness has been extensively investigated (reviewed in3, 57–60). Activating adenosine A1 receptors causes neuronal inhibition, and A1 is the most abundant adenosine receptor subtype in brain. This section highlights selected studies supporting the interpretation that adenosine promotes sleep, at least in part, by inhibiting neurons in several key wakefulness-promoting brain areas.
Prolonged wakefulness increases adenosine levels selectively in the basal forebrain (Fig. 1; BF) and cortex,61, 62 and increases adenosine A1 receptor binding in human63 and rat64 brain. Pharmacologically increasing adenosine levels in the basal forebrain65 or administering adenosine A1 receptor agonists to the basal forebrain59 causes an increase in sleep. Intravenous administration of buprenorphine decreases adenosine levels in the basal forebrain and increases wakefulness.66 Inactivating adenosine A1 receptors in the basal forebrain decreases EEG delta power and NREM sleep time,67 and immunohistochemical studies reveal that the basal forebrain contains A1 receptors, but not A2A receptors.68 Cholinergic neurons in the basal forebrain project to the cortex and contribute to the EEG activation characteristic of wakefulness and REM sleep. Adenosine directly inhibits cholinergic neurons in the basal forebrain by activating A1 receptors.69 Adenosine indirectly inhibits wakefulness-promoting hypocretin (orexin)-containing neurons in the lateral hypothalamus (Fig. 1; LH) by activating A1 receptors.70 Blocking adenosine A1 receptors in the lateral hypothalamus causes an increase in wakefulness and a decrease in sleep.71 Histaminergic neurons in the tuberomammillary nucleus (Fig. 1; TMN) express adenosine A1 receptors, and activating those receptors increases NREM sleep.72 These complementary data suggest that adenosine promotes sleep by inhibiting wakefulness-promoting neurons localized to the basal forebrain, lateral hypothalamus, and tuberomammillary nucleus.
Adenosine also exerts sleep-promoting effects by actions at the level of the prefrontal cortex (Fig. 1; PFC) and the pontine reticular formation (Fig. 1; PnO, PnC). In vivo microdialysis experiments in mouse73 have shown that adenosine acting at A1 receptors in the prefrontal cortex inhibits traits that characterize wakefulness (including acetylcholine release in the prefrontal cortex and activation of the EEG), as well as the state of wakefulness. Activation of adenosine A1 receptors in the prefrontal cortex also causes a decrease in the release of acetylcholine in the pontine reticular formation. These findings demonstrate that in the prefrontal cortex, adenosine A1 receptors mediate a descending inhibition of wakefulness-promoting systems. Within the pontine reticular formation, activation of adenosine A2A receptors increases time needed to recover from general anesthesia,74 increases acetylcholine release,74, 75 and increases the amount of time spent in NREM sleep75 and REM sleep.75, 76 The increase in REM sleep may be a result of the A2A-mediated increase in acetylcholine release, because coadministration of a muscarinic receptor antagonist with the A2A agonist blocks the REM sleep increase.76 Studies examining the effects on sleep of adenosine receptor antagonists are required in order to conclude that endogenous adenosine within the pontine reticular formation modulates sleep. The finding that clinically used opioids, such as morphine, fentanyl and buprenorphine, decrease adenosine levels in the pontine reticular formation66, 77 {added one reference} and disrupt REM sleep66 (also reviewed in78) suggests the possibility that adenosinergic transmission within the pontine reticular formation participates in REM sleep generation.
BIOGENIC AMINES
The monoamines have long been known to promote wakefulness. Serotonin (5-hydroxytryptamine; 5HT)-containing neurons of the dorsal raphé nucleus (Fig. 1; DRN), norepinephrine-containing neurons of the locus coeruleus (Fig. 1; LC), and histamine-containing neurons of the tuberomammillary nucleus (Fig. 1; TMN) discharge at their fastest rates during wakefulness, slow their firing in NREM sleep, cease discharging prior to and during REM sleep, and resume firing prior to the onset of wakefulness (reviewed in3). Dopaminergic neurons, by contrast, do not show major changes in firing rates across the sleep-wakefulness cycle.
Serotonin
Serotonin release in the dorsal raphé nucleus79 and preoptic area80 of rat is highest during wakefulness. Furthermore, electrical stimulation of the dorsal raphé nucleus increases wakefulness.81 Serotonin receptors are divided into seven families (5HT1–5HT7).82 Systemic administration of agonists for 5HT1A, 5HT1B, 5HT2A/2C or 5HT3 receptors causes an increase in wakefulness and a decrease in sleep (reviewed in7). Local administration of a 5HT1A receptor agonist to the dorsal raphé nucleus increases wakefulness in rat83 but increases REM sleep in cat.84 Microinjection of a 5HT2A/2C receptor agonist into rat dorsal raphé nucleus also decreases REM sleep with no significant effect on wakefulness.85 These incongruent findings may be due to species differences, or may indicate that in addition to promoting wakefulness, serotonin plays a permissive role in the generation of REM sleep. Systemic administration of antagonists for the 5HT2A receptor or the 5HT6 receptor to rat during the dark phase of the light/dark cycle (active period) decreases wakefulness, increases NREM sleep, and has no effect on REM sleep.86 These data are consistent with the view that serotonin is wakefulness-promoting. Genetically modified mice also have been used to explore the role of serotonin in sleep and wakefulness. Mice lacking the genes for the 5HT1A87 or 5HT1B88 receptor showed an increase in REM sleep. Administration of a 5HT1A87, 89, a 5HT1B88, or a 5HT2A/2C90 receptor agonist decreased REM sleep in rodent and human. These data indicate that serotonin acting at 5HT1A, 5HT1B, and 5HT2A/2C receptors plays a role in suppressing REM sleep. The forgoing data underlie the fact that insomnia can be secondary to the use of selective serotonin reuptake inhibitors (SSRI) or serotonin, norepinephrine reuptake inhibitors (SNRI).
Norepinephrine
Noradrenergic cells of the locus coeruleus inhibit REM sleep, promote wakefulness, and project to a variety of other arousal-regulating brain regions (Fig. 1) including the hypothalamus, thalamus, basal forebrain, and cortex (reviewed in91). Noradrenergic receptors include α1-, α2-, and β-adrenergic subtypes.92 Administration of noradrenaline or α- and β-receptor agonists to the medial septal area93, 94 or the medial preoptic area95, 96 increases wakefulness. Stimulation of locus coeruleus neurons increases noradrenaline in the prefrontal cortex of anesthetized rat,97, 98 and contributes to cortical activation. These data are consistent with the view that noradrenaline promotes wakefulness. However, bilateral microinjection of an α1-antagonist (prazosin), an α2-agonist (clonidine), or a β-antagonist (propranolol) into the pedunculopontine tegmental nucleus increases REM sleep with little to no effect on NREM sleep or wakefulness.99 The arousal-regulating effects of noradrenaline are brain-region specific. The treatment of hypertension with blockers of α- and/or β-adrenergic receptors can disrupt normal sleep.
Histamine
Histaminergic cell bodies, which are located in the tuberomamillary nucleus of the posterior hypothalamus have diffuse projections throughout the brain (reviewed in100, 101). Data from posterior hypothalamic lesion studies and from single unit recordings indicate that the tuberomamillary nucleus promotes wakefulness.100, 101 Three histaminergic receptors, denoted H1, H2, and H3, are present in the brain (for review see102). First generation H1 receptor antagonists, such as diphenhydramine, cause drowsiness (sedation) and impaired performance in humans103 and rats.104 Newer antagonists that are relatively selective for the H1 histamine receptor, such as the potent antagonist doxepin, improve subjective and objective measures of sleep in insomnia patients without causing sedation or psychomotor impairments the next day.105 Systemic administration of the H1 receptor antagonists mepyramine106 and cyproheptadine107 caused a significant increase in NREM sleep in cat and rat, respectively. Decreasing brain histamine levels by inhibiting synthesis significantly decreases wakefulness and increases NREM sleep in rat108, 109 and cat.106 These data suggest that histaminergic signaling via the H1 receptor promotes wakefulness. New therapies for sleep disorders and for maintaining vigilance include H3 receptor antagonists and inverse agonists.110–114
Dopamine
Stimulants such as amphetamine, cocaine, and methylphenidate increase wakefulness and counter hypersomnia by increasing levels of endogenous dopamine (reviewed in115). In vivo imaging studies suggest that sleep deprivation increases dopamine levels in human brain.116 The cell bodies of dopaminergic neurons that regulate arousal reside in the ventral tegmental area (Fig. 1; VTA) and the substantia nigra pars compacta.117 These dopaminergic neurons project to the dorsal raphé nucleus, basal forebrain, locus coeruleus, thalamus, and LDT (reviewed in118). There are also dopaminergic neurons in the ventrolateral periaqueductal gray that are active during wakefuless and have reciprocal connections with sleep-regulating brain areas.119
Five dopaminergic receptors have been cloned (D1–D5). Dopaminergic neurons of the substantia nigra and ventral tegmental area do not change firing rates as a function of states of sleep and wakefulness(reviewed in3). Dopamine does promote wakefulness and dopamine-transporter-knockout mice display increased wakefulness and decreased NREM sleep compared to controls.120 Systemic administration of D1 receptor agonists or antagonists causes an increase or decrease, respectively, in wakefulness.121 Intracerebroventricular administration of a D1 or D2 receptor agonist to rat increases wakefulness.122 Systemic administration of a D2 receptor agonist causes biphasic effects with low doses decreasing wakefulness and high doses increasing wakefulness.123, 124 Systemic administration of D-amphetamine to rat increases wakefulness and decreases NREM sleep and REM sleep.125 The mechanisms by which modafinil counters excessive daytime sleepiness remain to be specified. There is evidence that modafinil enhances synaptic release of dopamine and norepinephrine.126
GLUTAMATE
Glutamate is the main excitatory neurotransmitter in the brain and acts at α-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid (AMPA), kainate, and N-methyl-D-aspartate (NMDA) ionotropic receptors. Surprisingly, little is known about glutamatergic regulation of sleep and wakefulness. Sleep state-dependent changes in levels of endogenous glutamate change differentially across the brain (see Table 8 of127). For example, glutamate levels in some areas of rat cortex show increases in concentration during wakefulness and REM sleep, and decreases during NREM sleep,128 and glutamate concentrations in rat pontine reticular formation are higher during wakefulness than during NREM sleep and REM sleep.27 Sleep deprivation increases glutamate concentrations in rat dorsal hippocampus and medial thalamus.129 Microinjection and electrophysiological studies provide evidence that glutamate acts within the laterodorsal tegmental nucleus and pedunculopontine tegmental nucleus130–132 (Fig. 1; PPT), the pontine reticular formation31, 133, 134 (Fig. 1; PnO, PnC), the medial preoptic area,135 the insular cortex,136 and medial portions of the medullary reticular formation137, 138 to modulate traits and states of arousal. Glutamatergic neurons are present in rat pontine reticular formation139 and neurons in the pontine reticular formation are capable of synthesizing glutamate for use as a neurotransmitter.140 Glutamate elicits excitatory responses from pontine reticular formation neurons,141, 142 and glutamatergic and cholinergic transmission in the pontine reticular formation interact synergistically to potentiate catalepsy.143 Given individually, agonists for AMPA, kainate, and NMDA receptors evoke excitatory responses from pontine reticular formation neurons.133 Dialysis delivery of the NMDA receptor antagonists ketamine or MK-801 to cat pontine reticular formation decreases acetylcholine release in the pontine reticular formation and disrupts breathing.43
PEPTIDES
Many peptides are known to modulate sleep (reviewed in144). The present chapter focuses on hypocretin (orexin), leptin, and ghrelin because of their relevance for sleep disorders medicine.
Hypocretin-1 and -2
Numerous lines of evidence support a role for hypocretin-1 and -2 (also called orexin A and B) in the maintenance of wakefulness. The cell bodies of hypocretin-producing neurons are localized to the dorsolateral hypothalamus145, 146 and send projections to all the major brain regions that regulate arousal.147, 148 Hypocretinergic neurons discharge with the highest frequency during active wakefulness and show almost no discharge activity during sleep.149, 150 Hypocretin-1 levels in the hypothalamus of cat are greater during wakefulness and REM sleep than during NREM sleep.151 Dogs displaying a narcoleptic phenotype have a mutation of the hypocretin receptor-2 gene,152 and hypocretin mRNA and peptide levels are greatly reduced in human narcoleptic patients.153, 154 Patients presenting with narcolepsy-cataplexy also have greatly reduced levels of hypocretin in their cerebrospinal fluid compared to controls.155 Preclinical studies have demonstrated that selective lesions of hypocretin-containing neurons156, 157 or genetic removal of the peptide158 result in a narcoleptic phenotype. By what mechanisms might hypocretin enhance wakefulness?
Two receptors for the hypocretin peptides have been identified. Hypocretin-1 and -2 receptors have been localized to the LDT/PPT, pontine reticular formation, dorsal raphé nucleus, and locus coeruleus.159–163 Electrophysiological studies demonstrate that hypocretin-1 and/or hypocretin-2 excite neurons in these same brain regions.164–172 Hypocretin-1 and -2 also excite tuberomamillary neurons173, 174 and cholinergic neurons of the basal forebrain.175 Studies using intracerebroventricular injection in wild-type and knock-out mice (OX1R−/−, OX2R−/−, and OX1R−/−;OX2R−/−) suggest a differential regulation of arousal state via each hypocretin receptor subtype.176
Intracerebroventricular administration of hypocretin-1 increases wakefulness and decreases NREM sleep and REM sleep in rat.177, 178 When administered into the lateral preoptic area,179 the LDT,180 pontine reticular formation,24, 181 or basal forebrain,182, 183 hypocretin-1 causes an increase in wakefulness. In cat, microinjection of hypocretin-1 into the pontine reticular formation increases REM sleep if delivered during NREM sleep,166 but suppresses REM sleep if delivered during wakefulness.181 The wakefulness-promoting effect of hypocretin in the pontine reticular formation is further supported by evidence that delivery of antisense oligionucleotides against the hypocretin-2 receptor to the pontine reticular formation of rat enhance REM sleep and induce cataplexy.184
Measuring the effect of hypocretin-1 on the release of other arousal-regulating transmitters may provide insight into how hypocretin-1 promotes wakefulness. Microinjection of hypocretin-1 into the basal forebrain of rat increases cortical acetylcholine release.185 Intracerebroventricular delivery of hypocretin-increases histamine in rodent frontal cortex186 and anterior hypothalamus.187 Microinjection of hypocretin-1 into the ventricles or the ventral tegmental area increases dopamine release in rat prefrontal cortex.178 Hypocretin-1 delivered to rat dorsal raphé nucleus increases serotonin release in the dorsal raphé nucleus,188 and dialysis delivery of hypocretin-1 to rat pontine reticular formation increases acetylcholine release189 and GABA levels24 in the pontine reticular formation. The increase in wakefulness produced by microinjecting hypocretin-1 into the pontine reticular formation is prevented by blocking GABAA receptors.190 This finding suggests that hypocretin may increase wakefulness, in part, by increasing GABA levels in the pontine reticular formation. Considered together, these data support the classification of hypocretin-1 as a wakefulness-promoting neuropeptide.
An alternative hypothesis is that a primary function of hypocretin is to enhance activity in motor systems and the increase in wakefulness is secondary. This hypothesis is supported by data showing that hypocretin-1 concentrations in the cerebrospinal fluid are significantly greater during active wakefulness with movement than during quiet wakefulness with no movement.151 Hypocretinergic neurons also have very low firing rates during quiet wakefulness (without movement) compared to active wakefulness.149, 150 Oral administration of the hypocretin-1 and -2 receptor antagonists ACT-078573, DORA-22 or MK-6096 increases NREM sleep and/or REM sleep in mouse,191 rat,191, 192 dog,191, 192 and human,192 suggesting a direct, wakefulness-promoting effect of endogenous hypocretin.
Leptin and Ghrelin
Due to the ongoing epidemic of obesity and the association between metabolic syndrome and sleep disorders, many studies aim to understand the sleep-related roles of leptin and ghrelin. Decreased levels of leptin (a hormone that suppresses appetite) and increased levels of ghrelin (a hormone that stimulates appetite) are associated with short sleep duration in humans.193, 194 Obese patients with obstructive apnea sleep syndrome (OSAS) have increased plasma levels of leptin195 and ghrelin196 compared to age-matched obese patients without OSAS. Obese patients with OSAS and excessive day time sleepiness have significantly lower levels of ghrelin and a trend for lower plasma levels of leptin compared to obese patients with OSAS but without excessive day time sleepiness.197 These data suggest that there is a complex relationship between leptin, ghrelin, obesity, and sleep disruption that warrants further investigation. {New paragraph break}
Rodent models that may increase the understanding of the link between metabolic syndrome, leptin, and sleep disorders include ob/ob mice (obese mice with reduced levels of leptin) and db/db mice (which are also obese but are resistant to leptin). Leptin deficient mice have attenuated responses to certain drug treatments when compared to control species. When dialyzed into the prefrontal cortex of mouse, the atypical antipsychotic olanzapine increases acetylcholine release in the prefrontal cortex.198 The increase in acetylcholine release is significantly greater in C57BL/6J mice than in leptin-deficient mice. However, when leptin is restored to leptin-deficient mice, the olanzapine-induced increase in acetylcholine release is the same as that in the C57BL/6J mouse (Figure 4). Similarly, leptin deficient mice have a reduced antinociceptive response to supraspinal administration of neostigmine (an acetylcholinesterase inhibitor that increases levels of acetylcholine) when compared to C57BL/6J mice.199 Leptin replacement restores the antinociceptive responses of the leptin-deficient mice to that of the C57BL/6J mice. These data indicate that a possible link between leptin and cholinergic signaling within the prefrontal cortex and the pontine reticular formation – two brain areas that play a role in the regulation of sleep and wakefulness (see Figure 1).
Figure 4. Leptin replacement restores the olanzapine-induced increase of acetylcholine (ACh) release in the prefrontal cortex of leptin-deficient mice.
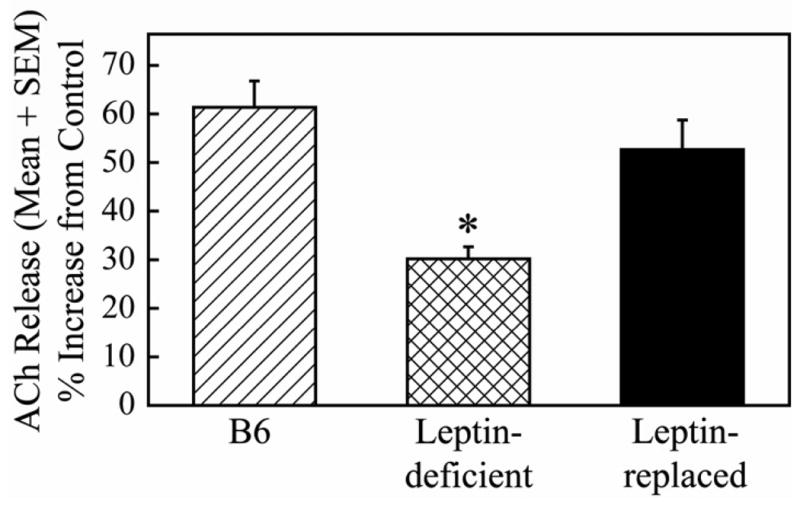
Dialysis administration of olanzapine (100 μM) to the prefrontal cortex of C57BL/6J (B6), leptin-deficient, or leptin-replaced mice caused an increase in ACh release in the prefrontal cortex. The increase in ACh release was significantly smaller in leptin-deficient mice compared to B6 controls. The olanzanpine-induced increase in ACh release was not significantly different between B6 controls and leptin-replaced mice. This suggests that leptin modulates the release of ACh within the prefrontal cortex and may also play a role in the cortical activation that occurs during wakefulness and REM sleep. Data reprinted from Wathen et al., 2012198 with permission.
The sleep of ob/ob mice is characterized by an increase in number of arousals and a decrease in the duration of sleep bouts compared to wild type controls.200 The ob/ob mice also have an impaired response to the cholinergic enhancement of REM sleep.201 Similarly, db/db mice have significant alterations in sleep architecture compared to wild type control mice that include, but are not limited to, increases in NREM sleep and REM sleep during the dark phase and decreases in wakefulness and NREM sleep bout duration.202 Local administration of ghrelin into rat lateral hypothalamus, medial preoptic area, or paraventricular nucleus increases wakefulness, decreases NREM sleep, and increases food intake.203 Together, these findings suggest that leptin and ghrelin, hormones that are important for appetite regulation, significantly influence sleep and are significantly modulated by sleep.
OPIOIDS
Opioids are the major class of drugs used to treat acute and chronic pain, and one side effect of opioids is sleep disruption. Sleep disruption, in turn, exacerbates pain204–206 and increases the dose of opioids required for successful pain management (reviewed in77, 78, 206). Clinically relevant doses of opioids given to naïve rats66 or to otherwise healthy humans (reviewed in207) disrupt sleep. For example, a single intravenous infusion of morphine in healthy volunteers decreases stages 3 and 4 NREM sleep, decreases REM sleep, and increases stage 2 NREM sleep.208 A nighttime dose of morphine or methadone also decreases stages 3 and 4 NREM sleep while increasing stage 2 NREM sleep.209 Constant infusion of analgesic doses of remifentanil overnight decreases REM sleep in healthy volunteers.210 Patients receiving methadone treatment for opioid dependence experience sleep disturbances including insomnia, decreases in total sleep time, slow wave sleep, and sleep efficiency as well as increases in the number of awakings.211–213 The cycle of opioid-induced sleep disruption leading to increased pain and increased opioid requirement is recognized as a significant clinical problem that must be addressed at the mechanistic level.214
Opioid-induced disruption of REM sleep is mediated, at least in part, by decreasing acetylcholine release in the pontine reticular formation.78 Opioids also decrease adenosine levels in the basal forebrain and in the pontine reticular formation,77 two brain regions where adenosine has sleep-promoting effects. Local administration of morphine into the pontine reticular formation of cat215 or rat216 increases wakefulness and decreases REM sleep.
FUTURE DIRECTIONS
This selective overview was completed during the summer of 2010, a date also marking the 20th anniversary of the human genome project. The stunning successes – and unmet hopes – of genomic approaches to medicine were highlighted in the June 12th and 14th issues of The New York Times.217, 218 These two articles offer a sobering reminder that taking a molecule from pre-clinical discovery to commercially available drug typically requires 15 or more years. This time interval is without any mandate to understand the mechanisms of drug action. As a former director of research and development at Wyeth noted218 “Genomics did not speed up drug development. It gave us more rapid access to new molecular targets.” Potential molecular targets can be rapidly interrogated with high throughput screening programs that use a cell line transfected to contain a reporter construct. But identifying potential molecular targets leaves unanswered the question of whether the candidate targets will be druggable in vivo. This complexity is exemplified by sedative/hypnotic medications commonly used in sleep medicine. GABAA receptors are drug targets that promotes a sleep-like state by unknown actions45 when they are activated in some brain regions, yet GABAA receptors enhance wakefulness when activated selectively in the posterior hypothalamus219 or pontine reticular formation.22, 23, 25 As busy as Fig. 1 may seem, it barely hints at the complexity of data that must be logically integrated if we are to derive a coherent model of the endogenous neurochemical processes that regulate states of sleep and wakefulness.
Recent progress in understanding the basic neuropharmacology of sleep can be appreciated by comparing the 1990 and the 2005 editions of Brain Control of Wakefulness and Sleep.3 The incorporation of basic neuropharmacology into sleep disorders medicine is readily apparent by comparing the first and most recent editions of Principles and Practice of Sleep Medicine.220 Future progress is most likely to come from a systems biology approach that seeks to integrate genomic, cellular, network, and behavioral levels of analysis.221 The focus on sleep medications in the Clinics of North America series demonstrates the cross-cutting relevance of sleep for the practice of medicine. The pressing clinical problem of sleep disorders medicine will continue to stimulate advances in understanding the neurochemical regulation of sleep.
Key Points.
Development of sedative/hypnotic molecules has been empiric rather than rational; the empiric approach has produced clinically useful drugs but for no drug is the mechanism of action completely understood.
All available sedative/hypnotic medications have unwanted side effects and none of these medications creates a sleep architecture that is identical to the architecture of naturally occurring sleep.
This chapter reviews recent advances in research aiming to elucidate the neurochemical mechanisms regulating sleep and wakefulness.
Footnotes
Disclosure Statement: This work supported by National Institutes of Health grants: HL40881, HL65272, MH45361, and the Department of Anesthesiology. We thank Mary A. Norat, and Sarah L. Watson for critical comments on this chapter. This work was not an industry-supported study and the authors have no financial conflicts of interest.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Watson CJ, Baghdoyan HA, Lydic R. Neuropharmacology of Sleep and Wakefulness. Sleep Med Clin. 2010;5(4):513–528. doi: 10.1016/j.jsmc.2010.08.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Watson CJ, Baghdoyan HA, Lydic R. A neurochemical perspective on states of consciousness. In: Hudetz AG, Pearce RA, editors. Suppressing the Mind: Anesthetic Modulation of Memory and Consciousness. New York: Springer/Humana Press; 2010. pp. 33–80. [Google Scholar]
- 3.Steriade M, McCarley RW, editors. Brain Control of Wakefulness and Sleep. New York: Kluwer Academic/Plenum Publishers; 2005. [Google Scholar]
- 4.Datta S, MacLean RR. Neurobiological mechanisms for the regulation of mammalian sleep-wake behavior: reinterpretation of historical evidence and inclusion of contemporary cellular and molecular evidence. Neurosci Biobehav Rev. 2007;31(5):775–824. doi: 10.1016/j.neubiorev.2007.02.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.McCarley RW. Neurobiology of REM and NREM sleep. Sleep Med. 2007;8(4):302–330. doi: 10.1016/j.sleep.2007.03.005. [DOI] [PubMed] [Google Scholar]
- 6.Stenberg D. Neuroanatomy and neurochemistry of sleep. Cell Mol Life Sci. 2007;64(10):1187–1204. doi: 10.1007/s00018-007-6530-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Monti JM, Pandi-Perumal SR, Sinton CM, editors. Neurochemistry of sleep and wakefulness. New York: Cambridge University Press; 2008. [Google Scholar]
- 8.Szymusiak R, McGinty D. Hypothalamic regulation of sleep and arousal. Ann N Y Acad Sci. 2008;1129:275–286. doi: 10.1196/annals.1417.027. [DOI] [PubMed] [Google Scholar]
- 9.Mallick BN, Pandi-Perumal SR, McCarley RW, et al. Rapid Eye Movement Sleep: Regulation and Function. New York: Cambridge University Press; 2010. [Google Scholar]
- 10.Espana RA, Scammell TE. Sleep neurobiology from a clinical perspective. Sleep. 2011;34(7):845–858. doi: 10.5665/SLEEP.1112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Narita M, Niikura K, Nanjo-Niikura K, et al. Sleep disturbances in a neuropathic pain-like condition in the mouse are associated with altered GABAergic transmission in the cingulate cortex. Pain. 2011;152(6):1358–1372. doi: 10.1016/j.pain.2011.02.016. [DOI] [PubMed] [Google Scholar]
- 12.Vienne J, Bettler B, Franken P, et al. Differential effects of GABAB receptor subtypes, {gamma}-hydroxybutyric Acid, and Baclofen on EEG activity and sleep regulation. J Neurosci. 2010;30(42):14194–14204. doi: 10.1523/JNEUROSCI.3145-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Olsen RW, Sieghart W. International Union of Pharmacology. LXX. Subtypes of gamma-aminobutyric acid(A) receptors: classification on the basis of subunit composition, pharmacology, and function. Update Pharmacol Rev. 2008;60(3):243–260. doi: 10.1124/pr.108.00505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Winsky-Sommerer R. Role of GABAA receptors in the physiology and pharmacology of sleep. Eur J Neurosci. 2009;29(9):1779–1794. doi: 10.1111/j.1460-9568.2009.06716.x. [DOI] [PubMed] [Google Scholar]
- 15.Fritschy JM, Mohler H. GABAA-receptor heterogeneity in the adult rat brain: differential regional and cellular distribution of seven major subunits. J Comp Neurol. 1995;359(1):154–194. doi: 10.1002/cne.903590111. [DOI] [PubMed] [Google Scholar]
- 16.Heldt SA, Ressler KJ. Forebrain and midbrain distribution of major benzodiazepine-sensitive GABAA receptor subunits in the adult C57 mouse as assessed with in situ hybridization. Neuroscience. 2007;150(2):370–385. doi: 10.1016/j.neuroscience.2007.09.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Pirker S, Schwarzer C, Wieselthaler A, et al. GABAA receptors: immunocytochemical distribution of 13 subunits in the adult rat brain. Neuroscience. 2000;101(4):815–850. doi: 10.1016/s0306-4522(00)00442-5. [DOI] [PubMed] [Google Scholar]
- 18.Farrant M, Nusser Z. Variations on an inhibitory theme: phasic and tonic activation of GABAA receptors. Nat Rev Neurosci. 2005;6(3):215–229. doi: 10.1038/nrn1625. [DOI] [PubMed] [Google Scholar]
- 19.Orser BA. Extrasynaptic GABAA receptors are critical targets for sedative-hypnotic drugs. J Clin Sleep Med. 2006;2(2):S12–18. [PubMed] [Google Scholar]
- 20.Walsh JK, Deacon S, Dijk DJ, et al. The selective extrasynaptic GABAA agonist, gaboxadol, improves traditional hypnotic efficacy measures and enhances slow wave activity in a model of transient insomnia. Sleep. 2007;30(5):593–602. doi: 10.1093/sleep/30.5.593. [DOI] [PubMed] [Google Scholar]
- 21.Camacho-Arroyo I, Alvarado R, Manjarrez J, et al. Microinjections of muscimol and bicuculline into the pontine reticular formation modify the sleep-waking cycle in the rat. Neurosci Lett. 1991;129(1):95–97. doi: 10.1016/0304-3940(91)90728-c. [DOI] [PubMed] [Google Scholar]
- 22.Flint RR, Chang T, Lydic R, et al. GABAA receptors in the pontine reticular formation of C57BL/6J mouse modulate neurochemical, electrographic, and behavioral phenotypes of wakefulness. J Neurosci. 2010;30(37):12301–12309. doi: 10.1523/JNEUROSCI.1119-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Xi MC, Morales FR, Chase MH. Evidence that wakefulness and REM sleep are controlled by a GABAergic pontine mechanism. J Neurophysiol. 1999;82(4):2015–2019. doi: 10.1152/jn.1999.82.4.2015. [DOI] [PubMed] [Google Scholar]
- 24.Watson CJ, Soto-Calderon H, Lydic R, et al. Pontine reticular formation (PnO) administration of hypocretin-1 increases PnO GABA levels and wakefulness. Sleep. 2008;31(4):453–464. doi: 10.1093/sleep/31.4.453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Vanini G, Watson CJ, Lydic R, et al. ®-aminobutyric acid-mediated neurotransmission in the pontine reticular formation modulates hypnosis, immobility, and breathing during isoflurane anesthesia. Anesthesiology. 2008;109(6):978–988. doi: 10.1097/ALN.0b013e31818e3b1b. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Vanini G, Wathen BL, Lydic R, et al. Endogenous GABA levels in the pontine reticular formation are greater during wakefulness than during rapid eye movement sleep. J Neurosci. 2011;31(7):2649–2656. doi: 10.1523/JNEUROSCI.5674-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Watson CJ, Lydic R, Baghdoyan HA. Sleep duration varies as a function of glutamate and GABA in rat pontine reticular formation. J Neurochem. 2011;118(4):571–580. doi: 10.1111/j.1471-4159.2011.07350.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Sanford LD, Tang X, Xiao J, et al. GABAergic regulation of REM sleep in reticularis pontis oralis and caudalis in rats. J Neurophysiol. 2003;90(2):938–945. doi: 10.1152/jn.00993.2002. [DOI] [PubMed] [Google Scholar]
- 29.Marks GA, Sachs OW, Birabil CG. Blockade of GABA, type A, receptors in the rat pontine reticular formation induces rapid eye movement sleep that is dependent upon the cholinergic system. Neuroscience. 2008;156(1):1–10. doi: 10.1016/j.neuroscience.2008.06.067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Vanini G, Torterolo P, McGregor R, et al. GABAergic processes in the mesencephalic tegmentum modulate the occurrence of active (rapid eye movement) sleep in guinea pigs. Neuroscience. 2007;145(3):1157–1167. doi: 10.1016/j.neuroscience.2006.12.051. [DOI] [PubMed] [Google Scholar]
- 31.Vanini G, Lydic R, Baghdoyan HA. GABAergic modulation of REM sleep. In: Mallick BN, Pandi-Perumal SR, McCarley RW, et al., editors. Rapid Eye Movement Sleep: Regulation and Function. New York: Cambridge University Press; 2010. pp. 206–213. [Google Scholar]
- 32.Vanini G, Baghdoyan HA, Lydic R. Relevance of sleep neurobiology for cognitive neuroscience and anesthesiology. In: Mashour GA, editor. Consciousness, Awareness, and Anesthesia. New York: Cambridge University Press; 2010. pp. 1–23. [Google Scholar]
- 33.Jouvet M. The role of monoamines and acetylcholine-containing neurons in the regulation of the sleep-waking cycle. Ergeb Physiol. 1972;64:166–307. doi: 10.1007/3-540-05462-6_2. [DOI] [PubMed] [Google Scholar]
- 34.Ishii M, Kurachi Y. Muscarinic acetylcholine receptors. Curr Pharm Des. 2006;12(28):3573–3581. doi: 10.2174/138161206778522056. [DOI] [PubMed] [Google Scholar]
- 35.Baghdoyan HA, Lydic R. M2 muscarinic receptor subtype in the feline medial pontine reticular formation modulates the amount of rapid eye movement sleep. Sleep. 1999;22(7):835–847. doi: 10.1093/sleep/22.7.835. [DOI] [PubMed] [Google Scholar]
- 36.Lydic R, Baghdoyan HA. Acetylcholine modulates sleep and wakefulness: a synaptic perspective. In: Monti JM, Pandi-Perumal SR, Sinton CM, editors. Neurochemistry of Sleep and Wakefulness. Cambridge: Cambridge University Press; 2008. pp. 109–143. [Google Scholar]
- 37.Baghdoyan HA. Location and quantification of muscarinic receptor subtypes in rat pons: implications for REM sleep generation. Am J Physiol. 1997;273(3 Pt 2):R896–904. doi: 10.1152/ajpregu.1997.273.3.R896. [DOI] [PubMed] [Google Scholar]
- 38.Demarco GJ, Baghdoyan HA, Lydic R. Differential cholinergic activation of G proteins in rat and mouse brainstem: relevance for sleep and nociception. J Comp Neurol. 2003;457(2):175–184. doi: 10.1002/cne.10548. [DOI] [PubMed] [Google Scholar]
- 39.Lydic R, Baghdoyan HA. Pedunculopontine stimulation alters respiration and increases ACh release in the pontine reticular formation. Am J Physiol. 1993;264(3 Pt 2):R544–554. doi: 10.1152/ajpregu.1993.264.3.R544. [DOI] [PubMed] [Google Scholar]
- 40.Thakkar M, Portas C, McCarley RW. Chronic low-amplitude electrical stimulation of the laterodorsal tegmental nucleus of freely moving cats increases REM sleep. Brain Res. 1996;723(1–2):223–227. doi: 10.1016/0006-8993(96)00256-9. [DOI] [PubMed] [Google Scholar]
- 41.Kodama T, Takahashi Y, Honda Y. Enhancement of acetylcholine release during paradoxical sleep in the dorsal tegmental field of the cat brain stem. Neurosci Lett. 1990;114(3):277–282. doi: 10.1016/0304-3940(90)90576-u. [DOI] [PubMed] [Google Scholar]
- 42.Leonard TO, Lydic R. Pontine nitric oxide modulates acetylcholine release, rapid eye movement sleep generation, and respiratory rate. J Neurosci. 1997;17(2):774–785. doi: 10.1523/JNEUROSCI.17-02-00774.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Lydic R, Baghdoyan HA. Ketamine and MK-801 decrease acetylcholine release in the pontine reticular formation, slow breathing, and disrupt sleep. Sleep. 2002;25(6):617–622. [PubMed] [Google Scholar]
- 44.Hambrecht-Wiedbusch VS, Gauthier EA, Baghdoyan HA, et al. Benzodiazepine receptor agonists cause drug-specific and state-specific alterations in EEG power and acetylcholine release in rat pontine reticular formation. Sleep. 2010;33(7):909–918. doi: 10.1093/sleep/33.7.909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Krystal AD. In vivo evidence of the specificity of effects of GABAA receptor modulating medications. Sleep. 2010;33(7):859–860. doi: 10.1093/sleep/33.7.859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Krystal AD, Walsh JK, Laska E, et al. Sustained efficacy of eszopiclone over 6 months of nightly treatment: results of a randomized, double-blind, placebo-controlled study in adults with chronic insomnia. Sleep. 2003;26(7):793–799. doi: 10.1093/sleep/26.7.793. [DOI] [PubMed] [Google Scholar]
- 47.Walsh JK, Krystal AD, Amato DA, et al. Nightly treatment of primary insomnia with eszopiclone for six months: effect on sleep, quality of life, and work limitations. Sleep. 2007;30(8):959–968. doi: 10.1093/sleep/30.8.959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Sarter M, Bruno JP. Cortical cholinergic inputs mediating arousal, attentional processing and dreaming: differential afferent regulation of the basal forebrain by telencephalic and brainstem afferents. Neuroscience. 2000;95(4):933–952. doi: 10.1016/s0306-4522(99)00487-x. [DOI] [PubMed] [Google Scholar]
- 49.Vazquez J, Baghdoyan HA. Basal forebrain acetylcholine release during REM sleep is significantly greater than during waking. Am J Physiol Regul Integr Comp Physiol. 2001;280(2):R598–601. doi: 10.1152/ajpregu.2001.280.2.R598. [DOI] [PubMed] [Google Scholar]
- 50.Marrosu F, Portas C, Mascia MS, et al. Microdialysis measurement of cortical and hippocampal acetylcholine release during sleep-wake cycle in freely moving cats. Brain Res. 1995;671(2):329–332. doi: 10.1016/0006-8993(94)01399-3. [DOI] [PubMed] [Google Scholar]
- 51.Materi LM, Rasmusson DD, Semba K. Inhibition of synaptically evoked cortical acetylcholine release by adenosine: an in vivo microdialysis study in the rat. Neuroscience. 2000;97(2):219–226. doi: 10.1016/s0306-4522(00)00075-0. [DOI] [PubMed] [Google Scholar]
- 52.Dworak M, McCarley RW, Kim T, et al. Sleep and brain energy levels: ATP changes during sleep. J Neurosci. 2010;30(26):9007–9016. doi: 10.1523/JNEUROSCI.1423-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Benington JH, Heller HC. Restoration of brain energy metabolism as the function of sleep. Prog Neurobiol. 1995;45(4):347–360. doi: 10.1016/0301-0082(94)00057-o. [DOI] [PubMed] [Google Scholar]
- 54.Landolt HP, Dijk DJ, Gaus SE, et al. Caffeine reduces low-frequency delta activity in the human sleep EEG. Neuropsychopharmacology. 1995;12(3):229–238. doi: 10.1016/0893-133X(94)00079-F. [DOI] [PubMed] [Google Scholar]
- 55.Landolt HP, Werth E, Borbely AA, et al. Caffeine intake (200 mg) in the morning affects human sleep and EEG power spectra at night. Brain Res. 1995;675(1–2):67–74. doi: 10.1016/0006-8993(95)00040-w. [DOI] [PubMed] [Google Scholar]
- 56.Gan TJ, Habib AS. Adenosine as a non-opioid analgesic in the perioperative setting. Anesth Analg. 2007;105(2):487–494. doi: 10.1213/01.ane.0000267260.00384.d9. [DOI] [PubMed] [Google Scholar]
- 57.Radulovacki M. Adenosine sleep theory: how I postulated it. Neurol Res. 2005;27(2):137–138. doi: 10.1179/016164105X21814. [DOI] [PubMed] [Google Scholar]
- 58.Basheer R, Strecker RE, Thakkar MM, et al. Adenosine and sleep-wake regulation. Prog Neurobiol. 2004;73(6):379–396. doi: 10.1016/j.pneurobio.2004.06.004. [DOI] [PubMed] [Google Scholar]
- 59.Strecker RE, Morairty S, Thakkar MM, et al. Adenosinergic modulation of basal forebrain and preoptic/anterior hypothalamic neuronal activity in the control of behavioral state. Behav Brain Res. 2000;115(2):183–204. doi: 10.1016/s0166-4328(00)00258-8. [DOI] [PubMed] [Google Scholar]
- 60.Porkka-Heiskanen T, Kalinchuk AV. Adenosine, energy metabolism and sleep homeostasis. Sleep Med Rev. 2011;15(2):123–135. doi: 10.1016/j.smrv.2010.06.005. [DOI] [PubMed] [Google Scholar]
- 61.Porkka-Heiskanen T, Strecker RE, McCarley RW. Brain site-specificity of extracellular adenosine concentration changes during sleep deprivation and spontaneous sleep: an in vivo microdialysis study. Neuroscience. 2000;99(3):507–517. doi: 10.1016/s0306-4522(00)00220-7. [DOI] [PubMed] [Google Scholar]
- 62.Kalinchuk AV, McCarley RW, Porkka-Heiskanen T, et al. The time course of adenosine, nitric oxide (NO) and inducible NO synthase changes in the brain with sleep loss and their role in the non-rapid eye movement sleep homeostatic cascade. J Neurochem. 2011;116(2):260–272. doi: 10.1111/j.1471-4159.2010.07100.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Elmenhorst D, Meyer PT, Winz OH, et al. Sleep deprivation increases A1 adenosine receptor binding in the human brain: a positron emission tomography study. J Neurosci. 2007;27(9):2410–2415. doi: 10.1523/JNEUROSCI.5066-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Elmenhorst D, Basheer R, McCarley RW, et al. Sleep deprivation increases A1 adenosine receptor density in the rat brain. Brain Res. 2009;1258:53–58. doi: 10.1016/j.brainres.2008.12.056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Porkka-Heiskanen T, Strecker RE, Thakkar M, et al. Adenosine: a mediator of the sleep-inducing effects of prolonged wakefulness. Science. 1997;276(5316):1265–1268. doi: 10.1126/science.276.5316.1265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Gauthier EA, Guzick SE, Brummett CM, et al. Buprenorphine disrupts sleep and decreases adenosine concentrations in sleep-regulating brain regions of Sprague Dawley rat. Anesthesiology. 2011;115(4):743–753. doi: 10.1097/ALN.0b013e31822e9f85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Thakkar MM, Winston S, McCarley RW. A1 receptor and adenosinergic homeostatic regulation of sleep-wakefulness: effects of antisense to the A1 receptor in the cholinergic basal forebrain. J Neurosci. 2003;23(10):4278–4287. doi: 10.1523/JNEUROSCI.23-10-04278.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Basheer R, Halldner L, Alanko L, et al. Opposite changes in adenosine A1 and A2A receptor mRNA in the rat following sleep deprivation. Neuroreport. 2001;12(8):1577–1580. doi: 10.1097/00001756-200106130-00013. [DOI] [PubMed] [Google Scholar]
- 69.Arrigoni E, Chamberlin NL, Saper CB, et al. Adenosine inhibits basal forebrain cholinergic and noncholinergic neurons in vitro. Neuroscience. 2006;140(2):403–413. doi: 10.1016/j.neuroscience.2006.02.010. [DOI] [PubMed] [Google Scholar]
- 70.Liu ZW, Gao XB. Adenosine inhibits activity of hypocretin/orexin neurons by the A1 receptor in the lateral hypothalamus: a possible sleep-promoting effect. J Neurophysiol. 2007;97(1):837–848. doi: 10.1152/jn.00873.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Thakkar MM, Engemann SC, Walsh KM, et al. Adenosine and the homeostatic control of sleep: effects of A1 receptor blockade in the perifornical lateral hypothalamus on sleep-wakefulness. Neuroscience. 2008;153(4):875–880. doi: 10.1016/j.neuroscience.2008.01.017. [DOI] [PubMed] [Google Scholar]
- 72.Oishi Y, Huang ZL, Fredholm BB, et al. Adenosine in the tuberomammillary nucleus inhibits the histaminergic system via A1 receptors and promotes non-rapid eye movement sleep. Proc Natl Acad Sci U S A. 2008;105(50):19992–19997. doi: 10.1073/pnas.0810926105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Van Dort CJ, Baghdoyan HA, Lydic R. Adenosine A1 and A2A receptors in mouse prefrontal cortex modulate acetylcholine release and behavioral arousal. J Neurosci. 2009;29(3):871–881. doi: 10.1523/JNEUROSCI.4111-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Tanase D, Baghdoyan HA, Lydic R. Dialysis delivery of an adenosine A1 receptor agonist to the pontine reticular formation decreases acetylcholine release and increases anesthesia recovery time. Anesthesiology. 2003;98(4):912–920. doi: 10.1097/00000542-200304000-00018. [DOI] [PubMed] [Google Scholar]
- 75.Coleman CG, Baghdoyan HA, Lydic R. Dialysis delivery of an adenosine A2A agonist into the pontine reticular formation of C57BL/6J mouse increases pontine acetylcholine release and sleep. J Neurochem. 2006;96(6):1750–1759. doi: 10.1111/j.1471-4159.2006.03700.x. [DOI] [PubMed] [Google Scholar]
- 76.Marks GA, Shaffery JP, Speciale SG, et al. Enhancement of rapid eye movement sleep in the rat by actions at A1 and A2a adenosine receptor subtypes with a differential sensitivity to atropine. Neuroscience. 2003;116(3):913–920. doi: 10.1016/s0306-4522(02)00561-4. [DOI] [PubMed] [Google Scholar]
- 77.Nelson AM, Battersby AS, Baghdoyan HA, et al. Opioid-induced decreases in rat brain adenosine levels are reversed by inhibiting adenosine deaminase. Anesthesiology. 2009;111(6):1327–1333. doi: 10.1097/ALN.0b013e3181bdf894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Lydic R, Baghdoyan HA. Neurochemical mechanisms mediating opioid-induced REM sleep disruption. In: Lavigne G, Sessle B, Choinière M, et al., editors. Sleep and Pain. Seattle: IASP Press; 2007. pp. 99–122. [Google Scholar]
- 79.Portas CM, Bjorvatn B, Fagerland S, et al. On-line detection of extracellular levels of serotonin in dorsal raphe nucleus and frontal cortex over the sleep/wake cycle in the freely moving rat. Neuroscience. 1998;83(3):807–814. doi: 10.1016/s0306-4522(97)00438-7. [DOI] [PubMed] [Google Scholar]
- 80.Python A, Steimer T, de Saint Hilaire Z, et al. Extracellular serotonin variations during vigilance states in the preoptic area of rats: a microdialysis study. Brain Res. 2001;910(1–2):49–54. doi: 10.1016/s0006-8993(01)02477-5. [DOI] [PubMed] [Google Scholar]
- 81.Houdouin F, Cespuglio R, Jouvet M. Effects induced by the electrical stimulation of the nucleus raphe dorsalis upon hypothalamic release of 5-hydroxyindole compounds and sleep parameters in the rat. Brain Res. 1991;565(1):48–56. doi: 10.1016/0006-8993(91)91735-j. [DOI] [PubMed] [Google Scholar]
- 82.Fink KB, Gothert M. 5-HT receptor regulation of neurotransmitter release. Pharmacol Rev. 2007;59(4):360–417. doi: 10.1124/pr.107.07103. [DOI] [PubMed] [Google Scholar]
- 83.Monti JM, Jantos H. Dose-dependent effects of the 5-HT1A receptor agonist 8-OH-DPAT on sleep and wakefulness in the rat. J Sleep Res. 1992;1(3):169–175. doi: 10.1111/j.1365-2869.1992.tb00033.x. [DOI] [PubMed] [Google Scholar]
- 84.Portas CM, Thakkar M, Rainnie D, et al. Microdialysis perfusion of 8-hydroxy-2-(di-n-propylamino)tetralin (8-OH-DPAT) in the dorsal raphe nucleus decreases serotonin release and increases rapid eye movement sleep in the freely moving cat. J Neurosci. 1996;16(8):2820–2828. doi: 10.1523/JNEUROSCI.16-08-02820.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Monti JM, Jantos H. Effects of activation and blockade of 5-HT2A/2C receptors in the dorsal raphe nucleus on sleep and waking in the rat. Prog Neuropsychopharmacol Biol Psychiatry. 2006;30(7):1189–1195. doi: 10.1016/j.pnpbp.2006.02.013. [DOI] [PubMed] [Google Scholar]
- 86.Morairty SR, Hedley L, Flores J, et al. Selective 5HT2A and 5HT6 receptor antagonists promote sleep in rats. Sleep. 2008;31(1):34–44. doi: 10.1093/sleep/31.1.34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Boutrel B, Monaca C, Hen R, et al. Involvement of 5-HT1A receptors in homeostatic and stress-induced adaptive regulations of paradoxical sleep: studies in 5-HT1A knock-out mice. J Neurosci. 2002;22(11):4686–4692. doi: 10.1523/JNEUROSCI.22-11-04686.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Boutrel B, Franc B, Hen R, et al. Key role of 5-HT1B receptors in the regulation of paradoxical sleep as evidenced in 5-HT1B knock-out mice. J Neurosci. 1999;19(8):3204–3212. doi: 10.1523/JNEUROSCI.19-08-03204.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Wilson SJ, Bailey JE, Rich AS, et al. The use of sleep measures to compare a new 5HT1A agonist with buspirone in humans. J Psychopharmacol. 2005;19(6):609–613. doi: 10.1177/0269881105058775. [DOI] [PubMed] [Google Scholar]
- 90.Monti JM, Jantos H. Effects of the serotonin 5-HT2A/2C receptor agonist DOI and of the selective 5-HT2A or 5-HT2C receptor antagonists EMD 281014 and SB-243213, respectively, on sleep and waking in the rat. Eur J Pharmacol. 2006;553(1–3):163–170. doi: 10.1016/j.ejphar.2006.09.027. [DOI] [PubMed] [Google Scholar]
- 91.Berridge CW, Waterhouse BD. The locus coeruleus-noradrenergic system: modulation of behavioral state and state-dependent cognitive processes. Brain Res Brain Res Rev. 2003;42(1):33–84. doi: 10.1016/s0165-0173(03)00143-7. [DOI] [PubMed] [Google Scholar]
- 92.Hein L. Adrenoceptors and signal transduction in neurons. Cell Tissue Res. 2006;326(2):541–551. doi: 10.1007/s00441-006-0285-2. [DOI] [PubMed] [Google Scholar]
- 93.Berridge CW, Foote SL. Enhancement of behavioral and electroencephalographic indices of waking following stimulation of noradrenergic beta-receptors within the medial septal region of the basal forebrain. J Neurosci. 1996;16(21):6999–7009. doi: 10.1523/JNEUROSCI.16-21-06999.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Berridge CW, Isaac SO, Espana RA. Additive wake-promoting actions of medial basal forebrain noradrenergic alpha1- and beta-receptor stimulation. Behav Neurosci. 2003;117(2):350–359. doi: 10.1037/0735-7044.117.2.350. [DOI] [PubMed] [Google Scholar]
- 95.Kumar VM, Datta S, Chhina GS, et al. Alpha adrenergic system in medial preoptic area involved in sleep-wakefulness in rats. Brain Res Bull. 1986;16(4):463–468. doi: 10.1016/0361-9230(86)90174-7. [DOI] [PubMed] [Google Scholar]
- 96.Sood S, Dhawan JK, Ramesh V, et al. Role of medial preoptic area beta adrenoceptors in the regulation of sleep-wakefulness. Pharmacol Biochem Behav. 1997;57(1–2):1–5. doi: 10.1016/s0091-3057(96)00384-x. [DOI] [PubMed] [Google Scholar]
- 97.Berridge CW, Abercrombie ED. Relationship between locus coeruleus discharge rates and rates of norepinephrine release within neocortex as assessed by in vivo microdialysis. Neuroscience. 1999;93(4):1263–1270. doi: 10.1016/s0306-4522(99)00276-6. [DOI] [PubMed] [Google Scholar]
- 98.Florin-Lechner SM, Druhan JP, Aston-Jones G, et al. Enhanced norepinephrine release in prefrontal cortex with burst stimulation of the locus coeruleus. Brain Res. 1996;742(1–2):89–97. doi: 10.1016/s0006-8993(96)00967-5. [DOI] [PubMed] [Google Scholar]
- 99.Pal D, Mallick BN. Role of noradrenergic and GABA-ergic inputs in pedunculopontine tegmentum for regulation of rapid eye movement sleep in rats. Neuropharmacology. 2006;51(1):1–11. doi: 10.1016/j.neuropharm.2006.02.006. [DOI] [PubMed] [Google Scholar]
- 100.Haas HL, Sergeeva OA, Selbach O. Histamine in the nervous system. Physiol Rev. 2008;88(3):1183–1241. doi: 10.1152/physrev.00043.2007. [DOI] [PubMed] [Google Scholar]
- 101.Thakkar MM. Histamine in the regulation of wakefulness. Sleep Med Rev. 2011;15(1):65–74. doi: 10.1016/j.smrv.2010.06.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Haas H, Panula P. The role of histamine and the tuberomamillary nucleus in the nervous system. Nat Rev Neurosci. 2003;4(2):121–130. doi: 10.1038/nrn1034. [DOI] [PubMed] [Google Scholar]
- 103.Nicholson AN, Stone BM. Antihistamines: impaired performance and the tendency to sleep. Eur J Clin Pharmacol. 1986;30(1):27–32. doi: 10.1007/BF00614191. [DOI] [PubMed] [Google Scholar]
- 104.Kaneko Y, Shimada K, Saitou K, et al. The mechanism responsible for the drowsiness caused by first generation H1 antagonists on the EEG pattern. Methods Find Exp Clin Pharmacol. 2000;22(3):163–168. [PubMed] [Google Scholar]
- 105.Roth T, Rogowski R, Hull S, et al. Efficacy and safety of doxepin 1 mg, 3 mg, and 6 mg in adults with primary insomnia. Sleep. 2007;30(11):1555–1561. doi: 10.1093/sleep/30.11.1555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Lin JS, Sakai K, Jouvet M. Evidence for histaminergic arousal mechanisms in the hypothalamus of cat. Neuropharmacology. 1988;27(2):111–122. doi: 10.1016/0028-3908(88)90159-1. [DOI] [PubMed] [Google Scholar]
- 107.Tokunaga S, Takeda Y, Shinomiya K, et al. Effects of some H1-antagonists on the sleep-wake cycle in sleep-disturbed rats. J Pharmacol Sci. 2007;103(2):201–206. doi: 10.1254/jphs.fp0061173. [DOI] [PubMed] [Google Scholar]
- 108.Monti JM, D’Angelo L, Jantos H, et al. Effects of a-fluoromethylhistidine on sleep and wakefulness in the rat. Short note J Neural Transm. 1988;72(2):141–145. doi: 10.1007/BF01250237. [DOI] [PubMed] [Google Scholar]
- 109.Kiyono S, Seo ML, Shibagaki M, et al. Effects of α-fluoromethylhistidine on sleep-waking parameters in rats. Physiol Behav. 1985;34(4):615–617. doi: 10.1016/0031-9384(85)90057-5. [DOI] [PubMed] [Google Scholar]
- 110.Parmentier R, Anaclet C, Guhennec C, et al. The brain H3-receptor as a novel therapeutic target for vigilance and sleep-wake disorders. Biochem Pharmacol. 2007;73(8):1157–1171. doi: 10.1016/j.bcp.2007.01.002. [DOI] [PubMed] [Google Scholar]
- 111.Ligneau X, Perrin D, Landais L, et al. BF2. 649 [1-{3-[3-(4-Chlorophenyl)propoxy]propyl}piperidine, hydrochloride], a nonimidazole inverse agonist/antagonist at the human histamine H3 receptor: Preclinical pharmacology. J Pharmacol Exp Ther. 2007;320(1):365–375. doi: 10.1124/jpet.106.111039. [DOI] [PubMed] [Google Scholar]
- 112.Le S, Gruner JA, Mathiasen JR, et al. Correlation between ex vivo receptor occupancy and wake-promoting activity of selective H3 receptor antagonists. J Pharmacol Exp Ther. 2008;325(3):902–909. doi: 10.1124/jpet.107.135343. [DOI] [PubMed] [Google Scholar]
- 113.James LM, Iannone R, Palcza J, et al. Effect of a novel histamine subtype-3 receptor inverse agonist and modafinil on EEG power spectra during sleep deprivation and recovery sleep in male volunteers. Psychopharmacology (Berl) 2011;215(4):643–653. doi: 10.1007/s00213-010-2158-3. [DOI] [PubMed] [Google Scholar]
- 114.Lin JS, Sergeeva OA, Haas HL. Histamine H3 receptors and sleep-wake regulation. J Pharmacol Exp Ther. 2011;336(1):17–23. doi: 10.1124/jpet.110.170134. [DOI] [PubMed] [Google Scholar]
- 115.Boutrel B, Koob GF. What keeps us awake: the neuropharmacology of stimulants and wakefulness-promoting medications. Sleep. 2004;27(6):1181–1194. doi: 10.1093/sleep/27.6.1181. [DOI] [PubMed] [Google Scholar]
- 116.Volkow ND, Wang GJ, Telang F, et al. Sleep deprivation decreases binding of [11C]raclopride to dopamine D2/D3 receptors in the human brain. J Neurosci. 2008;28(34):8454–8461. doi: 10.1523/JNEUROSCI.1443-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Monti JM, Jantos H. The roles of dopamine and serotonin, and of their receptors, in regulating sleep and waking. Prog Brain Res. 2008;172:625–646. doi: 10.1016/S0079-6123(08)00929-1. [DOI] [PubMed] [Google Scholar]
- 118.Monti JM, Monti D. The involvement of dopamine in the modulation of sleep and waking. Sleep Med Rev. 2007;11(2):113–133. doi: 10.1016/j.smrv.2006.08.003. [DOI] [PubMed] [Google Scholar]
- 119.Lu J, Jhou TC, Saper CB. Identification of wake-active dopaminergic neurons in the ventral periaqueductal gray matter. J Neurosci. 2006;26(1):193–202. doi: 10.1523/JNEUROSCI.2244-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Wisor JP, Nishino S, Sora I, et al. Dopaminergic role in stimulant-induced wakefulness. J Neurosci. 2001;21(5):1787–1794. doi: 10.1523/JNEUROSCI.21-05-01787.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Monti JM, Fernandez M, Jantos H. Sleep during acute dopamine D1 agonist SKF 38393 or D1 antagonist SCH 23390 administration in rats. Neuropsychopharmacology. 1990;3(3):153–162. [PubMed] [Google Scholar]
- 122.Isaac SO, Berridge CW. Wake-promoting actions of dopamine D1 and D2 receptor stimulation. J Pharmacol Exp Ther. 2003;307(1):386–394. doi: 10.1124/jpet.103.053918. [DOI] [PubMed] [Google Scholar]
- 123.Monti JM, Hawkins M, Jantos H, et al. Biphasic effects of dopamine D-2 receptor agonists on sleep and wakefulness in the rat. Psychopharmacology (Berl) 1988;95(3):395–400. doi: 10.1007/BF00181955. [DOI] [PubMed] [Google Scholar]
- 124.Monti JM, Jantos H, Fernandez M. Effects of the selective dopamine D-2 receptor agonist, quinpirole on sleep and wakefulness in the rat. Eur J Pharmacol. 1989;169(1):61–66. doi: 10.1016/0014-2999(89)90817-0. [DOI] [PubMed] [Google Scholar]
- 125.Andersen ML, Margis R, Frey BN, et al. Electrophysiological correlates of sleep disturbance induced by acute and chronic administration of D-amphetamine. Brain Res. 2009;1249:162–172. doi: 10.1016/j.brainres.2008.10.023. [DOI] [PubMed] [Google Scholar]
- 126.Minzenberg MJ, Carter CS. Modafinil: a review of neurochemical actions and effects on cognition. Neuropsychopharmacology. 2008;33(7):1477–1502. doi: 10.1038/sj.npp.1301534. [DOI] [PubMed] [Google Scholar]
- 127.Brevig HN, Baghdoyan HA. Neurotransmitters and neuromodulators regulating sleep and wakefulness. In: Koob GF, Le Moa M, Thompson RF, editors. Encyclopedia of Behavioral Neuroscience. Vol. 3. Oxford: Academic Press; 2010. pp. 456–463. [Google Scholar]
- 128.Dash MB, Douglas CL, Vyazovskiy VV, et al. Long-term homeostasis of extracellular glutamate in the rat cerebral cortex across sleep and waking states. J Neurosci. 2009;29(3):620–629. doi: 10.1523/JNEUROSCI.5486-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Cortese BM, Mitchell TR, Galloway MP, et al. Region-specific alteration in brain glutamate: possible relationship to risk-taking behavior. Physiol Behav. 2010;99(4):445–450. doi: 10.1016/j.physbeh.2009.12.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Datta S, Patterson EH, Spoley EE. Excitation of the pedunculopontine tegmental NMDA receptors induces wakefulness and cortical activation in the rat. J Neurosci Res. 2001;66(1):109–116. doi: 10.1002/jnr.1202. [DOI] [PubMed] [Google Scholar]
- 131.Datta S, Spoley EE, Patterson EH. Microinjection of glutamate into the pedunculopontine tegmentum induces REM sleep and wakefulness in the rat. Am J Physiol Regul Integr Comp Physiol. 2001;280(3):R752–759. doi: 10.1152/ajpregu.2001.280.3.R752. [DOI] [PubMed] [Google Scholar]
- 132.Datta S, Spoley EE, Mavanji VK, et al. A novel role of pedunculopontine tegmental kainate receptors: a mechanism of rapid eye movement sleep generation in the rat. Neuroscience. 2002;114(1):157–164. doi: 10.1016/s0306-4522(02)00250-6. [DOI] [PubMed] [Google Scholar]
- 133.Stevens DR, McCarley RW, Greene RW. Excitatory amino acid-mediated responses and synaptic potentials in medial pontine reticular formation neurons of the rat in vitro. J Neurosci. 1992;12(11):4188–4194. doi: 10.1523/JNEUROSCI.12-11-04188.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Onoe H, Sakai K. Kainate receptors: a novel mechanism in paradoxical (REM) sleep generation. Neuroreport. 1995;6(2):353–356. [PubMed] [Google Scholar]
- 135.Kaushik MK, Kumar VM, Mallick HN. Glutamate microinjection at the medial preoptic area enhances slow wave sleep in rats. Behav Brain Res. 2011;217(1):240–243. doi: 10.1016/j.bbr.2010.11.007. [DOI] [PubMed] [Google Scholar]
- 136.Cui L, Wang JH, Wang M, et al. Injection of L: -glutamate into the insular cortex produces sleep apnea and serotonin reduction in rats. Sleep Breath. 2011 doi: 10.1007/s11325-011-0586-x. [DOI] [PubMed] [Google Scholar]
- 137.Lai YY, Siegel JM. Medullary regions mediating atonia. J Neurosci. 1988;8(12):4790–4796. doi: 10.1523/JNEUROSCI.08-12-04790.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Lai YY, Siegel JM. Pontomedullary glutamate receptors mediating locomotion and muscle tone suppression. J Neurosci. 1991;11(9):2931–2937. doi: 10.1523/JNEUROSCI.11-09-02931.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Kaneko T, Itoh K, Shigemoto R, et al. Glutaminase-like immunoreactivity in the lower brainstem and cerebellum of the adult rat. Neuroscience. 1989;32(1):79–98. doi: 10.1016/0306-4522(89)90109-7. [DOI] [PubMed] [Google Scholar]
- 140.Jones BE. Arousal systems. Front Biosci. 2003;8:s438–451. doi: 10.2741/1074. [DOI] [PubMed] [Google Scholar]
- 141.Núñez A, Buño W, Reinoso-Suárez F. Neurotransmitter actions on oral pontine tegmental neurons of the rat: an in vitro study. Brain Res. 1998;804(1):144–148. doi: 10.1016/s0006-8993(98)00681-7. [DOI] [PubMed] [Google Scholar]
- 142.Greene RW, Carpenter DO. Actions of neurotransmitters on pontine medical reticular formation neurons of the cat. J Neurophysiol. 1985;54(3):520–531. doi: 10.1152/jn.1985.54.3.520. [DOI] [PubMed] [Google Scholar]
- 143.Elazar Z, Berchanski A. Glutamatergic-cholinergic synergistic interaction in the pontine reticular formation. Effects on catalepsy Naunyn Schmiedebergs. Arch Pharmacol. 2001;363(5):569–576. doi: 10.1007/s002100100400. [DOI] [PubMed] [Google Scholar]
- 144.de Lecea L. Neuropeptides and sleep-wake regulation. In: Monti JM, Pandi-Perumal SR, Sinton CM, editors. Neurochemistry of Sleep and Wakefulness. New York: Cambridge University Press; 2008. pp. 387–401. [Google Scholar]
- 145.de Lecea L, Kilduff TS, Peyron C, et al. The hypocretins: hypothalamus-specific peptides with neuroexcitatory activity. Proc Natl Acad Sci U S A. 1998;95(1):322–327. doi: 10.1073/pnas.95.1.322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Sakurai T, Amemiya A, Ishii M, et al. Orexins and orexin receptors: a family of hypothalamic neuropeptides and G protein-coupled receptors that regulate feeding behavior. Cell. 1998;92(4):573–585. doi: 10.1016/s0092-8674(00)80949-6. [DOI] [PubMed] [Google Scholar]
- 147.Peyron C, Tighe DK, van den Pol AN, et al. Neurons containing hypocretin (orexin) project to multiple neuronal systems. J Neurosci. 1998;18(23):9996–10015. doi: 10.1523/JNEUROSCI.18-23-09996.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Zhang JH, Sampogna S, Morales FR, et al. Distribution of hypocretin (orexin) immunoreactivity in the feline pons and medulla. Brain Res. 2004;995(2):205–217. doi: 10.1016/j.brainres.2003.10.004. [DOI] [PubMed] [Google Scholar]
- 149.Lee MG, Hassani OK, Jones BE. Discharge of identified orexin/hypocretin neurons across the sleep-waking cycle. J Neurosci. 2005;25(28):6716–6720. doi: 10.1523/JNEUROSCI.1887-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Mileykovskiy BY, Kiyashchenko LI, Siegel JM. Behavioral correlates of activity in identified hypocretin/orexin neurons. Neuron. 2005;46(5):787–798. doi: 10.1016/j.neuron.2005.04.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Kiyashchenko LI, Mileykovskiy BY, Maidment N, et al. Release of hypocretin (orexin) during waking and sleep states. J Neurosci. 2002;22(13):5282–5286. doi: 10.1523/JNEUROSCI.22-13-05282.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Lin L, Faraco J, Li R, et al. The sleep disorder canine narcolepsy is caused by a mutation in the hypocretin (orexin) receptor 2 gene. Cell. 1999;98(3):365–376. doi: 10.1016/s0092-8674(00)81965-0. [DOI] [PubMed] [Google Scholar]
- 153.Thannickal TC, Moore RY, Nienhuis R, et al. Reduced number of hypocretin neurons in human narcolepsy. Neuron. 2000;27(3):469–474. doi: 10.1016/s0896-6273(00)00058-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Peyron C, Faraco J, Rogers W, et al. A mutation in a case of early onset narcolepsy and a generalized absence of hypocretin peptides in human narcoleptic brains. Nat Med. 2000;6(9):991–997. doi: 10.1038/79690. [DOI] [PubMed] [Google Scholar]
- 155.Nishino S, Kanbayashi T. Symptomatic narcolepsy, cataplexy and hypersomnia, and their implications in the hypothalamic hypocretin/orexin system. Sleep Med Rev. 2005;9(4):269–310. doi: 10.1016/j.smrv.2005.03.004. [DOI] [PubMed] [Google Scholar]
- 156.Beuckmann CT, Sinton CM, Williams SC, et al. Expression of a poly-glutamine-ataxin-3 transgene in orexin neurons induces narcolepsy-cataplexy in the rat. J Neurosci. 2004;24(18):4469–4477. doi: 10.1523/JNEUROSCI.5560-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Murillo-Rodriguez E, Liu M, Blanco-Centurion C, et al. Effects of hypocretin (orexin) neuronal loss on sleep and extracellular adenosine levels in the rat basal forebrain. Eur J Neurosci. 2008;28(6):1191–1198. doi: 10.1111/j.1460-9568.2008.06424.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Willie JT, Chemelli RM, Sinton CM, et al. Distinct narcolepsy syndromes in Orexin receptor-2 and Orexin null mice: molecular genetic dissection of Non-REM and REM sleep regulatory processes. Neuron. 2003;38(5):715–730. doi: 10.1016/s0896-6273(03)00330-1. [DOI] [PubMed] [Google Scholar]
- 159.Bernard R, Lydic R, Baghdoyan HA. Hypocretin-1 causes G protein activation and increases ACh release in rat pons. Eur J Neurosci. 2003;18(7):1775–1785. doi: 10.1046/j.1460-9568.2003.02905.x. [DOI] [PubMed] [Google Scholar]
- 160.Greco MA, Shiromani PJ. Hypocretin receptor protein and mRNA expression in the dorsolateral pons of rats. Brain Res Mol Brain Res. 2001;88(1–2):176–182. doi: 10.1016/s0169-328x(01)00039-0. [DOI] [PubMed] [Google Scholar]
- 161.Hervieu GJ, Cluderay JE, Harrison DC, et al. Gene expression and protein distribution of the orexin-1 receptor in the rat brain and spinal cord. Neuroscience. 2001;103(3):777–797. doi: 10.1016/s0306-4522(01)00033-1. [DOI] [PubMed] [Google Scholar]
- 162.Marcus JN, Aschkenasi CJ, Lee CE, et al. Differential expression of orexin receptors 1 and 2 in the rat brain. J Comp Neurol. 2001;435(1):6–25. doi: 10.1002/cne.1190. [DOI] [PubMed] [Google Scholar]
- 163.Brischoux F, Mainville L, Jones BE. Muscarinic-2 and orexin-2 receptors on GABAergic and other neurons in the rat mesopontine tegmentum and their potential role in sleep-wake state control. J Comp Neurol. 2008;510(6):607–630. doi: 10.1002/cne.21803. [DOI] [PubMed] [Google Scholar]
- 164.Burlet S, Tyler CJ, Leonard CS. Direct and indirect excitation of laterodorsal tegmental neurons by Hypocretin/Orexin peptides: implications for wakefulness and narcolepsy. J Neurosci. 2002;22(7):2862–2872. doi: 10.1523/JNEUROSCI.22-07-02862.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Takahashi K, Koyama Y, Kayama Y, et al. Effects of orexin on the laterodorsal tegmental neurones. Psychiatry Clin Neurosci. 2002;56(3):335–336. doi: 10.1046/j.1440-1819.2002.00967.x. [DOI] [PubMed] [Google Scholar]
- 166.Xi MC, Fung SJ, Yamuy J, et al. Induction of active (REM) sleep and motor inhibition by hypocretin in the nucleus pontis oralis of the cat. J Neurophysiol. 2002;87(6):2880–2888. doi: 10.1152/jn.2002.87.6.2880. [DOI] [PubMed] [Google Scholar]
- 167.Liu RJ, van den Pol AN, Aghajanian GK. Hypocretins (orexins) regulate serotonin neurons in the dorsal raphe nucleus by excitatory direct and inhibitory indirect actions. J Neurosci. 2002;22(21):9453–9464. doi: 10.1523/JNEUROSCI.22-21-09453.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Soffin EM, Gill CH, Brough SJ, et al. Pharmacological characterisation of the orexin receptor subtype mediating postsynaptic excitation in the rat dorsal raphe nucleus. Neuropharmacology. 2004;46(8):1168–1176. doi: 10.1016/j.neuropharm.2004.02.014. [DOI] [PubMed] [Google Scholar]
- 169.Brown RE, Sergeeva OA, Eriksson KS, et al. Convergent excitation of dorsal raphe serotonin neurons by multiple arousal systems (orexin/hypocretin, histamine and noradrenaline) J Neurosci. 2002;22(20):8850–8859. doi: 10.1523/JNEUROSCI.22-20-08850.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Bourgin P, Huitron-Resendiz S, Spier AD, et al. Hypocretin-1 modulates rapid eye movement sleep through activation of locus coeruleus neurons. J Neurosci. 2000;20(20):7760–7765. doi: 10.1523/JNEUROSCI.20-20-07760.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Hagan JJ, Leslie RA, Patel S, et al. Orexin A activates locus coeruleus cell firing and increases arousal in the rat. Proc Natl Acad Sci U S A. 1999;96(19):10911–10916. doi: 10.1073/pnas.96.19.10911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Horvath TL, Peyron C, Diano S, et al. Hypocretin (orexin) activation and synaptic innervation of the locus coeruleus noradrenergic system. J Comp Neurol. 1999;415(2):145–159. [PubMed] [Google Scholar]
- 173.Eriksson KS, Sergeeva O, Brown RE, et al. Orexin/hypocretin excites the histaminergic neurons of the tuberomammillary nucleus. J Neurosci. 2001;21(23):9273–9279. doi: 10.1523/JNEUROSCI.21-23-09273.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Bayer L, Eggermann E, Serafin M, et al. Orexins (hypocretins) directly excite tuberomammillary neurons. Eur J Neurosci. 2001;14(9):1571–1575. doi: 10.1046/j.0953-816x.2001.01777.x. [DOI] [PubMed] [Google Scholar]
- 175.Eggermann E, Serafin M, Bayer L, et al. Orexins/hypocretins excite basal forebrain cholinergic neurones. Neuroscience. 2001;108(2):177–181. doi: 10.1016/s0306-4522(01)00512-7. [DOI] [PubMed] [Google Scholar]
- 176.Mieda M, Hasegawa E, Kisanuki YY, et al. Differential roles of orexin receptor-1 and -2 in the regulation of non-REM and REM sleep. J Neurosci. 2011;31(17):6518–6526. doi: 10.1523/JNEUROSCI.6506-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Piper DC, Upton N, Smith MI, et al. The novel brain neuropeptide, orexin-A, modulates the sleep-wake cycle of rats. Eur J Neurosci. 2000;12(2):726–730. doi: 10.1046/j.1460-9568.2000.00919.x. [DOI] [PubMed] [Google Scholar]
- 178.Vittoz NM, Berridge CW. Hypocretin/orexin selectively increases dopamine efflux within the prefrontal cortex: involvement of the ventral tegmental area. Neuropsychopharmacology. 2006;31(2):384–395. doi: 10.1038/sj.npp.1300807. [DOI] [PubMed] [Google Scholar]
- 179.Methippara MM, Alam MN, Szymusiak R, et al. Effects of lateral preoptic area application of orexin-A on sleep-wakefulness. Neuroreport. 2000;11(16):3423–3426. doi: 10.1097/00001756-200011090-00004. [DOI] [PubMed] [Google Scholar]
- 180.Xi MC, Morales FR, Chase MH. Effects on sleep and wakefulness of the injection of hypocretin-1 (orexin-A) into the laterodorsal tegmental nucleus of the cat. Brain Res. 2001;901(1–2):259–264. doi: 10.1016/s0006-8993(01)02317-4. [DOI] [PubMed] [Google Scholar]
- 181.Moreno-Balandran E, Garzon M, Bodalo C, et al. Sleep-wakefulness effects after microinjections of hypocretin 1 (orexin A) in cholinoceptive areas of the cat oral pontine tegmentum. Eur J Neurosci. 2008;28(2):331–341. doi: 10.1111/j.1460-9568.2008.06334.x. [DOI] [PubMed] [Google Scholar]
- 182.Espana RA, Baldo BA, Kelley AE, et al. Wake-promoting and sleep-suppressing actions of hypocretin (orexin): basal forebrain sites of action. Neuroscience. 2001;106(4):699–715. doi: 10.1016/s0306-4522(01)00319-0. [DOI] [PubMed] [Google Scholar]
- 183.Thakkar MM, Ramesh V, Strecker RE, et al. Microdialysis perfusion of orexin-A in the basal forebrain increases wakefulness in freely behaving rats. Arch Ital Biol. 2001;139(3):313–328. [PubMed] [Google Scholar]
- 184.Thakkar MM, Ramesh V, Cape EG, et al. REM sleep enhancement and behavioral cataplexy following orexin (hypocretin)-II receptor antisense perfusion in the pontine reticular formation. Sleep Res Online. 1999;2(4):112–120. [PubMed] [Google Scholar]
- 185.Dong HL, Fukuda S, Murata E, et al. Orexins increase cortical acetylcholine release and electroencephalographic activation through orexin-1 receptor in the rat basal forebrain during isoflurane anesthesia. Anesthesiology. 2006;104(5):1023–1032. doi: 10.1097/00000542-200605000-00019. [DOI] [PubMed] [Google Scholar]
- 186.Hong ZY, Huang ZL, Qu WM, et al. Orexin A promotes histamine, but not norepinephrine or serotonin, release in frontal cortex of mice. Acta Pharmacol Sin. 2005;26(2):155–159. doi: 10.1111/j.1745-7254.2005.00523.x. [DOI] [PubMed] [Google Scholar]
- 187.Ishizuka T, Yamamoto Y, Yamatodani A. The effect of orexin-A and -B on the histamine release in the anterior hypothalamus in rats. Neurosci Lett. 2002;323(2):93–96. doi: 10.1016/s0304-3940(01)02552-6. [DOI] [PubMed] [Google Scholar]
- 188.Tao R, Ma Z, McKenna JT, et al. Differential effect of orexins (hypocretins) on serotonin release in the dorsal and median raphe nuclei of freely behaving rats. Neuroscience. 2006;141(3):1101–1105. doi: 10.1016/j.neuroscience.2006.05.027. [DOI] [PubMed] [Google Scholar]
- 189.Bernard R, Lydic R, Baghdoyan HA. Hypocretin (orexin) receptor subtypes differentially enhance acetylcholine release and activate g protein subtypes in rat pontine reticular formation. J Pharmacol Exp Ther. 2006;317(1):163–171. doi: 10.1124/jpet.105.097071. [DOI] [PubMed] [Google Scholar]
- 190.Brevig HN, Watson CJ, Lydic R, et al. Hypocretin and GABA interact in the pontine reticular formation to increase wakefulness. Sleep. 2010;33(10):1285–1293. doi: 10.1093/sleep/33.10.1285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Winrow CJ, Gotter AL, Cox CD, et al. Pharmacological characterization of MK-6096 - A dual orexin receptor antagonist for insomnia. Neuropharmacology. 2011 doi: 10.1016/j.neuropharm.2011.10.003. in press. [DOI] [PubMed] [Google Scholar]
- 192.Brisbare-Roch C, Dingemanse J, Koberstein R, et al. Promotion of sleep by targeting the orexin system in rats, dogs and humans. Nat Med. 2007;13(2):150–155. doi: 10.1038/nm1544. [DOI] [PubMed] [Google Scholar]
- 193.Taheri S, Lin L, Austin D, et al. Short sleep duration is associated with reduced leptin, elevated ghrelin, and increased body mass index. PLoS Med. 2004;1(3):e62. doi: 10.1371/journal.pmed.0010062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Spiegel K, Tasali E, Penev P, et al. Brief communication: Sleep curtailment in healthy young men is associated with decreased leptin levels, elevated ghrelin levels, and increased hunger and appetite. Ann Intern Med. 2004;141(11):846–850. doi: 10.7326/0003-4819-141-11-200412070-00008. [DOI] [PubMed] [Google Scholar]
- 195.Basoglu OK, Sarac F, Sarac S, et al. Metabolic syndrome, insulin resistance, fibrinogen, homocysteine, leptin, and C-reactive protein in obese patients with obstructive sleep apnea syndrome. Ann Thorac Med. 2011;6(3):120–125. doi: 10.4103/1817-1737.82440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196.Ursavas A, Ilcol YO, Nalci N, et al. Ghrelin, leptin, adiponectin, and resistin levels in sleep apnea syndrome: Role of obesity. Ann Thorac Med. 2010;5(3):161–165. doi: 10.4103/1817-1737.65050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197.Sanchez-de-la-Torre M, Barcelo A, Pierola J, et al. Plasma levels of neuropeptides and metabolic hormones, and sleepiness in obstructive sleep apnea. Respir Med. 2011;105(12):1954–1960. doi: 10.1016/j.rmed.2011.08.014. [DOI] [PubMed] [Google Scholar]
- 198.Wathen AB, West EM, Lydic R, et al. Olanzapine causes a leptin-dependent increase in acetylcholine release in mouse prefrontal cortex. Sleep. 2012;35(3):315–323. doi: 10.5665/sleep.1686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Wang W, Baghdoyan HA, Lydic R. Leptin replacement restores supraspinal cholinergic antinociception in leptin-deficient obese mice. J Pain. 2009;10(8):836–843. doi: 10.1016/j.jpain.2009.02.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.Laposky AD, Shelton J, Bass J, et al. Altered sleep regulation in leptin-deficient mice. Am J Physiol Regul Integr Comp Physiol. 2006;290(4):R894–903. doi: 10.1152/ajpregu.00304.2005. [DOI] [PubMed] [Google Scholar]
- 201.Douglas CL, Bowman GN, Baghdoyan HA, et al. C57BL/6J and B6. V-LEPOB mice differ in the cholinergic modulation of sleep and breathing. J Appl Physiol. 2005;98(3):918–929. doi: 10.1152/japplphysiol.00900.2004. [DOI] [PubMed] [Google Scholar]
- 202.Laposky AD, Bradley MA, Williams DL, et al. Sleep-wake regulation is altered in leptin-resistant (db/db) genetically obese and diabetic mice. Am J Physiol Regul Integr Comp Physiol. 2008;295(6):R2059–2066. doi: 10.1152/ajpregu.00026.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 203.Szentirmai E, Kapas L, Krueger JM. Ghrelin microinjection into forebrain sites induces wakefulness and feeding in rats. Am J Physiol Regul Integr Comp Physiol. 2007;292(1):R575–585. doi: 10.1152/ajpregu.00448.2006. [DOI] [PubMed] [Google Scholar]
- 204.Roehrs T, Hyde M, Blaisdell B, et al. Sleep loss and REM sleep loss are hyperalgesic. Sleep. 2006;29(2):145–151. doi: 10.1093/sleep/29.2.145. [DOI] [PubMed] [Google Scholar]
- 205.Chhangani BS, Roehrs TA, Harris EJ, et al. Pain sensitivity in sleepy pain-free normals. Sleep. 2009;32(8):1011–1017. [PMC free article] [PubMed] [Google Scholar]
- 206.Mystakidou K, Clark AJ, Fischer J, et al. Treatment of chronic pain by long-acting opioids and the effects on sleep. Pain Pract. 2011;11(3):282–289. doi: 10.1111/j.1533-2500.2010.00417.x. [DOI] [PubMed] [Google Scholar]
- 207.Lavigne G, Sessle BJ, Choinière M, et al., editors. Sleep and Pain. Seattle: International Association for the Study of Pain Press; 2007. [Google Scholar]
- 208.Shaw IR, Lavigne G, Mayer P, et al. Acute intravenous administration of morphine perturbs sleep architecture in healthy pain-free young adults: a preliminary study. Sleep. 2005;28(6):677–682. doi: 10.1093/sleep/28.6.677. [DOI] [PubMed] [Google Scholar]
- 209.Dimsdale JE, Norman D, DeJardin D, et al. The effect of opioids on sleep architecture. J Clin Sleep Med. 2007;3(1):33–36. [PubMed] [Google Scholar]
- 210.Bonafide CP, Aucutt-Walter N, Divittore N, et al. Remifentanil inhibits rapid eye movement sleep but not the nocturnal melatonin surge in humans. Anesthesiology. 2008;108(4):627–633. doi: 10.1097/ALN.0b013e3181684bc3. [DOI] [PubMed] [Google Scholar]
- 211.Sharkey KM, Kurth ME, Anderson BJ, et al. Assessing sleep in opioid dependence: a comparison of subjective ratings, sleep diaries, and home polysomnography in methadone maintenance patients. Drug Alcohol Depend. 2011;113(2–3):245–248. doi: 10.1016/j.drugalcdep.2010.08.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 212.Trksak GH, Jensen JE, Plante DT, et al. Effects of sleep deprivation on sleep homeostasis and restoration during methadone-maintenance: a [31]P MRS brain imaging study. Drug Alcohol Depend. 2010;106(2–3):79–91. doi: 10.1016/j.drugalcdep.2009.07.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 213.Xiao L, Tang YL, Smith AK, et al. Nocturnal sleep architecture disturbances in early methadone treatment patients. Psychiatry Res. 2010;179(1):91–95. doi: 10.1016/j.psychres.2009.02.003. [DOI] [PubMed] [Google Scholar]
- 214.Moore JT, Kelz MB. Opiates, sleep, and pain: the adenosinergic link. Anesthesiology. 2009;111(6):1175–1176. doi: 10.1097/ALN.0b013e3181bdfa2e. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 215.Keifer JC, Baghdoyan HA, Lydic R. Sleep disruption and increased apneas after pontine microinjection of morphine. Anesthesiology. 1992;77(5):973–982. doi: 10.1097/00000542-199211000-00021. [DOI] [PubMed] [Google Scholar]
- 216.Watson CJ, Lydic R, Baghdoyan HA. Sleep and GABA levels in the oral part of rat pontine reticular formation are decreased by local and systemic administration of morphine. Neuroscience. 2007;144(1):375–386. doi: 10.1016/j.neuroscience.2006.09.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 217.Wade N. The New York Times. Jun 12, 2010. A decade later, genetic map yields few new cures. [Google Scholar]
- 218.Pollack A. The New York TImes. Jun 14, 2010. Awaiting the genome payoff. [Google Scholar]
- 219.Lin JS, Sakai K, Vanni-Mercier G, et al. A critical role of the posterior hypothalamus in the mechanisms of wakefulness determined by microinjection of muscimol in freely moving cats. Brain Res. 1989;479(2):225–240. doi: 10.1016/0006-8993(89)91623-5. [DOI] [PubMed] [Google Scholar]
- 220.Kryger MH, Roth T, Dement WC, editors. Principles and Practice of Sleep Medicine. 5. Philadelphia: W.B. Saunders; 2010. [Google Scholar]
- 221.Klipp E, Herwig R, Kowald A, et al. Systems Biology in Practice: Concepts, Implementation, and Application. Weinham: Wiley-VCH; 2005. [Google Scholar]
- 222.Paxinos G, Watson C. The Rat Brain in Stereotaxic Coordinates. 6. New York: Academic Press; 2007. [Google Scholar]