

Abstract
We presented small groups of chimpanzees with two collective action situations, in which action was necessary for reward but there was a disincentive for individuals to act owing to the possibility of free-riding on the efforts of others. We found that in simpler scenarios (experiment 1) in which group size was small, there was a positive relationship between rank and action with more dominant individuals volunteering to act more often, particularly when the reward was less dispersed. Social tolerance also seemed to mediate action whereby higher tolerance levels within a group resulted in individuals of lower ranks sometimes acting and appropriating more of the reward. In more complex scenarios, when group size was larger and cooperation was necessary (experiment 2), overcoming the problem was more challenging. There was highly significant variability in the action rates of different individuals as well as between dyads, suggesting success was more greatly influenced by the individual personalities and personal relationships present in the group.
Keywords: cooperation, collective action, free-riding, chimpanzees, volunteer's dilemma, dominance
1. Introduction
Collective action problems (CAPs) arise in situations in which effortful action is required, at a cost to the actor(s), and such action results in the distribution or availability of a shared good of which non-actors may also benefit. In collective action scenarios, the optimal strategy for any individual is to free-ride—that is, let others assume the cost of action yet benefit from the rewards. This disincentive to action produces what is known as a CAP [1].
One of the best-known examples of a CAP in a non-human species involves individual variability in participation in group-territorial conflict among female lions, in which certain females were prone to lead the move towards an intruder, whereas others lagged behind, avoiding the risks of conflict yet benefiting in the renewed safety of their territory [2]. Other examples of collective action include aggressive extra-group encounters in monkeys and lemurs [1,3] and nested female guarding alliances in dolphins [4].
There are two outstanding questions important to determining how groups overcome CAPs. First, what mechanisms allow for impasse being averted? Nunn [1] suggests different classes of factors that may potentially promote action, including: (i) asymmetrical benefits and privileged groups, which would provide extra incentive for certain individuals to act (e.g. Mitani and Watts found a positive correlation between male chimpanzee participation in boundary patrols and mating success, suggesting that participants are individuals with the most to gain [5]); (ii) private incentives, additional goods only received by contributors and (iii) coercion. Second, in situations where a CAP is successfully overcome, what governs the strategies different individuals adopt, and do patterns in decision-making arise? For instance, it is possible that rank or tolerance between group members may influence an individual's motivation to act.
Among wild chimpanzees, one example of a potential CAP is group hunting of colobus monkeys. All chimpanzees in a group are motivated to obtain nutrient-rich scraps of meat from a monkey carcass. It is important to note the probability of a male obtaining a scrap does increase with the number of hunters [6], and therefore, there is an incentive for an individual to act; however, two possible types of deterrent are also still present. First, hunting always entails opportunity cost, risk of injury and energy cost. Second, in the largest groups, hunters and non-hunters may be equally likely to get some of the prize [6], suggesting that at least in large party contexts an individual still has an incentive to free-ride (although this appears to vary by site, see [6–9]). These factors remain reasons an individual might limit participation and suggest that hunts can represent CAPs, especially in large groups. How chimpanzees overcome the disincentives to action and initiate and complete successful hunts remains unclear [6,9].
Gilby et al. [6] argue that individual variation in hunting motivation is the most important factor predicting the likelihood of a hunt (i.e. solution of the CAP).They identified certain highly motivated males as ‘impact hunters,’ owing to their critical role in triggering hunts. It remains unclear which traits are most important for shaping an impact hunter. Age is unlikely to be a determining factor, as impact hunters seem to retain this role over several years [9]. Dominance could potentially have an effect; however, it is hard to disentangle dominance from qualities important to hunting ability such as strength and agility [9]. Skill is most likely essential, and personality appears central [9].
With few exceptions, experimental approaches exploring chimpanzee cooperative problem-solving have focused on dyadic interactions [10–13], which limits their application to more complex, and perhaps more naturalistic, interactions such as CAPs. Controlled experimental examination of carefully constructed CAP scenarios is necessary to enrich our understanding of the mechanisms underlying the behaviours documented in such contexts in the field.
Different types of CAP reflect particular payoff structures. Our experiments are modelled on the Volunteer's Dilemma, a scenario in which a shared good is produced only if at least one individual volunteers to pay a cost. Each individual wants the good to be produced but prefers someone else volunteers. This preference leads some to behave as free-riders, defined as individuals who receive rewards from the efforts of others without contributing to their production. However, if no one else volunteers, all individuals lose [14–16].
In a first experiment, we took the initial step in looking beyond dyadic interactions to investigate the factors that potentially govern individual strategy in a shared goods game. We designed a scenario as follows: groups of chimpanzees were given access to three indoor testing rooms in which any individual could choose to push a button resulting in juice being dispensed into drinking troughs on the opposite side of the testing area. We manipulated group size (such that subjects were either tested in dyads or triads) and dispersion (such that the juice flowed into either a single trough or three troughs side by side, with higher dispersion making the reward less excludable).
Our second experiment had three primary differences from the first: we made the action cooperative, the reward was more widely dispersed (but kept constant across conditions) and we increased the difference between group sizes. Groups of chimpanzees were tested in a set up that allowed any two individuals to choose to coordinate pulling two ends of a rope, which would result in peanuts being sprayed into an opposing room. Subjects were tested in either triads or sextets, which should intensify the CAP and make it more striking for the subjects.
Because of the rivalrous nature of the reward, and the distance between the action and reward locations, the principle cost for the actor is a reduction in the amount of potential reward. While hunting, wild chimpanzees face more severe costs such as opportunity costs, energy expenditure and risk of injury, and while it is not an option to mimic all of these experimentally, it is important to keep this difference in mind.
We hypothesized that changes to group size and reward dispersion would alter the payoff structure among subjects, resulting in individuals demonstrating different strategies and propensities for action dependent on condition. Specifically, that dominant individuals would be most likely to act across conditions, whereas subordinate individuals would act more when the reward was more greatly dispersed and group size was small.
2. Experiment 1
(a). Subjects
Subjects include 12 chimpanzees housed at the Wolfgang Köhler Primate Research Center at the Leipzig Zoo, Germany. Subjects were seven females (age range: 7–16 years old) and five males (age range: 5–33 years old). Three subjects lived in a social group comprised of six individuals; nine subjects lived in a social group comprised of 17 individuals. See the electronic supplementary material, S1 for more details concerning the subjects' sex, age, relations, grouping and living conditions. Subjects were separated into four groups of three individuals and tested in dyad and triad combinations within these groups. Groups were formed with the guidance of the keepers to ensure sufficient tolerance levels between individuals to prevent undue stress and aggression. A dominance hierarchy for each group was generated in consultation with the keepers.
(b). Apparatus
The apparatus consisted of four principal parts: an action box, a collecting box, reward troughs and the tubes that connected them (figure 1). The action box had an inner rectangular basin into which juice could be poured by the experimenter. The action box was connected to the collecting box (a smaller juice basin attached to an adjacent room) by a short clear tube. Continuous pushing of a button on the action box would cause the level of liquid in the collecting box to rise until it was high enough for juice to flow out of the three long tubes anchored to its top edge, allowing the liquid to become accessible to the subjects as it moved through the tubes and spread out in the reward troughs two rooms away. See the electronic supplementary material, S2 for a more detailed description.
Figure 1.
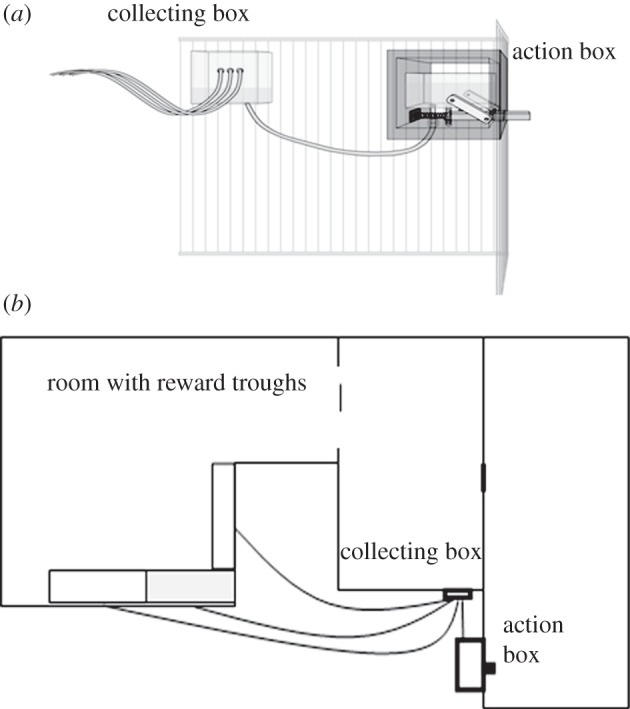
(a) Close up of the action box and collecting box; (b) bird's eye view of the layout used in experiment 1. The trough used in the 1trough condition is shaded grey.
(c). Procedure and design
(i). Individual introduction to the apparatus
Individuals passed through a series of pre-tests to ensure that they understood how the apparatus functioned and were motivated to push the button for the desired length of time when alone. The pre-tests and test were ordered as follows (see the electronic supplementary material, S3 for more details on each pre-test):
One trough, one room. Subjects were in one room and had access only to the action box and one drinking trough (directly underneath), and learned how to dispense the reward.
One trough, three rooms. Subjects had access to the action box and one trough placed two rooms away and learned to dispense juice and then leave the action box and move to the trough room to retrieve the reward.
Three troughs, three rooms. Subjects had access to the action box and all three troughs placed two rooms away and learned that when they dispensed juice it would spread out in all the troughs.
Half liquid. Same as previous pre-test except that the amount of juice in the action box was reduced by half such that subjects had to learn to push longer before juice was dispensed.
Test. Subjects were separated into four distinct groups of three individuals. We manipulated two variables (number of troughs and group size) resulting in four different conditions: dyad-1trough, dyad-3troughs, triad-1trough and triad-3troughs. Number of troughs was counterbalanced across groups such that two test groups started with three troughs and continued to one trough and two groups received one trough first. Group size followed an ABA design such that subjects underwent six dyad sessions followed by six triad sessions followed by six dyad sessions. Each dyad session consisted of three trials (one for each possible dyad combination, with each individual being tested twice). Each triad session consisted of two trials (again each individual was tested twice).
Subjects began each trial confined to the room with the reward troughs. A trial began when the door of this room was raised, allowing all individuals access to the adjacent room as well as to the room with the action box. Trials lasted 5 min. The experimenter and keeper were absent from the testing area throughout the duration of the trial.
(ii). Coding and analysis
All trials were recorded by three cameras. One camera focused on the action box and collecting box, while the two others were directed at the drinking trough(s). After each trial, the amount of juice that had been released was measured. The following variables were later coded from videotape: (i) latency to first push (latency); (ii) whether the subject pushed the button (push.yn); (iii) the duration of each pushing bout (push_total) (a bout begins when the subject pushes the button and ends when they release the button for more than 3 s); (iv) whether the subject drank from the trough (drink.yn); (v) the duration of each drinking bout (drink_total) (a drinking bout begins when the subject drinks or licks the trough and ends when they stop for 3 or more seconds).
To test whether pushing behaviour (push.yn, pushing duration) and drinking duration were influenced by the number of troughs, dominance rank or the size of the group, we used a generalized linear mixed model (GLMM) [17]. We included the following predictors as fixed effects: rank, group size (dyad versus triad), number of troughs (one versus three; and all interactions between them up to the three-way interaction), session number and trial number, and as random effects: trial, group_id, subject and triad_id. We did two additional analyses to test whether the amount released or the latency to first push were influenced by the number of troughs or the size of the group, into which we included the following predictors as fixed effects: group size (dyad versus triad), number of troughs (one versus three; and all interactions between them), session and trial and as random effects group_id and triad_id (see the electronic supplementary material, S4). Data: doi:10.5061/dryad.b5c1f.
(d). Results
Owing to the small sample size of four triad groups, all results must be interpreted with caution and we complement these statistical results with more qualitative descriptions of behaviour within groups and overall patterns (see the electronic supplementary material, S8).
In testing whether an individual pushed, we found a three-way interaction effect between rank, group size and number of troughs (GLMM, p = 0.0116, electronic supplementary material, S5). In general, there appears to be a positive relationship between rank and tendency to push, with higher-ranking individuals pushing more. This relationship breaks down in the triad-3troughs condition. Pushing time is more uniformly distributed in the triad condition. Number of troughs appears to have an effect only in the triad condition (figure 2).
Figure 2.

Median (bar) and quartile (line) pushing probability in 1trough and 3troughs conditions as a function of group and rank. The GLMM revealed a three-way interaction between group size, rank, and number of troughs (see text). Rank from highest (1) to lowest (3).
With respect to pushing duration, we found a two-way interaction effect between rank and number of troughs as well as between number of troughs and group size (GLMM, estimated pMCMC = 0.0010 and 0.0094, respectively; electronic supplementary material, S6). Among dyads, there is little difference in pushing duration regardless of number of troughs, while in triads the middle-ranking individual pushes more in the 3troughs than in the 1trough condition.
In testing for the factors affecting drinking duration, we found a two-way interaction effect between rank and number of troughs (GLMM, estimated pMCMC = 0.0001, electronic supplementary material, S7). In the 1-trough condition, drinking duration was positively associated with rank, with the most dominant individual drinking the longest on average. In the 3troughs condition, drinking durations were almost equal.
Finally, none of the factors had a significant influence on the amount of liquid released or on latency.
(e). Discussion
In this Volunteer's Dilemma context, reward dispersion did affect the payoff expectations of individuals of different rank and thus also their willingness to act. In all conditions except for triad-3troughs, dominants tended to face a relatively low cost for action, as they could always be assured a worthwhile portion of the reward while for subordinates the opposite was true. In the triad-3troughs condition, however, both dispersion and rivalry for the reward were maximized and thus dominants were no longer assured a large portion of the juice after acting (their expected payoff decreased), whereas subordinates would be more interested in making sure the reward is produced, as their chance to sneak into the trough area and drink increased. These changes in strategy are reflected in figure 2. While dispersion and rivalry affected all subjects, what strategies were viable in each particular group was likely due to interactions between dominance and group tolerance levels [10,18]. While each dyad and triad had the same dominance structure (dominant/subordinate in the dyad and dominant/middle/subordinate in the triad), the magnitude of the differences in rank varied between groups. The degree of tolerance between individuals also varied.
Overall, role patterns can be summarized as follows (see the electronic supplementary material, S8 for details about strategies evident in each group as well as the identity of group members): pushers tended to be individuals higher in rank, free-riders tended to be subordinate individuals in groups with high tolerance levels or subordinate individuals with particularly tolerant relationships with the dominant in their group. Dominants who pushed were always assured a share of the reward, which lessened the risk of action. It was also the case that individuals in groups with medium tolerance levels tended to use flexible strategies, switching roles based on reward dispersion. Finally, in dyads with low tolerance if the subordinate was especially fearful of the dominant, they generally ended up obtaining scant reward.
3. Experiment 2
In our second experiment, we sought to exacerbate the dilemma (through larger group size and more dispersed reward).
(a). Subjects
Subjects were 12 semi-free ranging residents of the Ngamba Island Chimpanzee Sanctuary in Lake Victoria, Uganda (www.ngambaisland.org). Subjects were eight males (age range: 8–13 years) and four females (age range: 8–13 years). See the electronic supplementary material, S9 for details concerning the subjects' sex, age, experimental history, grouping and living conditions. All subjects had demonstrated an understanding of a similar cooperative rope mechanism in previous studies [10–12,19]. With input from keepers, six different groups of six individuals were formed for the test phase. Each of these groups was further divided into four groups of three individuals to allow for two group-size conditions.
(b). Apparatus
The principal testing area consisted of two opposing rooms connected by an overhead raceway. A large funnel was attached to one room (figure 3). The funnel's circular opening was blocked by a sliding piece of Plexiglas with a hole at the far end. A rope was guided around two wooden dowels attached to the Plexiglas and the ends passed through the cage bars such that when the two ends were pulled in synchrony, the piece would slide, the holes align and the peanuts would be released. A long ramp was attached directly under where the holes of the funnel and Plexiglas piece aligned, allowing the peanuts that were released to roll down across the hallway to a battery-operated feed dispenser attached to the opposing cage. Once peanuts reached the feeder, E would start the motor at a distance by remote. The peanuts (up to 320, depending on how long the ropes were pulled) would then spray into the cage. See the electronic supplementary material, S10 for a more detailed description.
Figure 3.
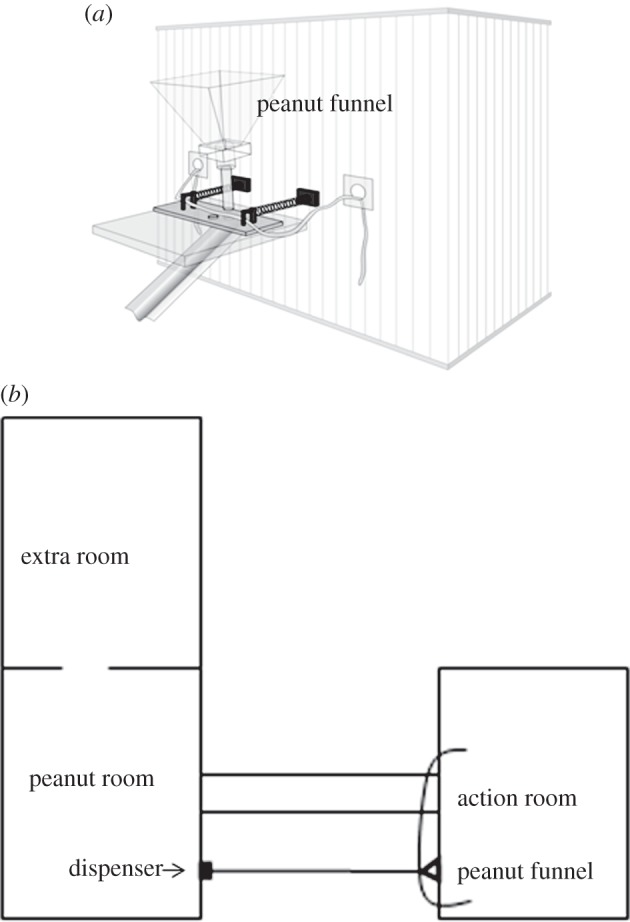
(a) Close up of the peanut dispenser; (b) bird's eye view of the layout used in experiment 2. Note that the peanuts spray into the room opposite where the rope ends are pulled forcing subjects to climb up to and cross an overhead raceway to transition from action to reward access.
(c). Procedure and design
(i). Individual introduction to the apparatus
Rope ends were inserted at a distance which allowed the subjects to pull alone. They learned to release peanuts and climb from the action room over to the peanut room to eat them. See the electronic supplementary material, S11 for more details.
(ii). Dyad pre-test
Cooperation trials. All possible dyad combinations within each sextet were tested to assess their willingness to work together to release peanuts. It has already been demonstrated that tolerance levels have a great influence on success in cooperative tasks [10], and it was important to decipher which pairs would work together in a simpler non-CAP context to potentially better understand their behaviour in the larger group dynamic. See the electronic supplementary material, S11 for more details.
Dominance trials. Cooperation trials were followed by two dominance trials in which an eighth of a watermelon was placed within reach in the corridor. Pairs were allowed to enter the baited room through the door furthest from the reward. The first individual to reach the watermelon and to eat all the most attractive bits (flesh and light green rind) was noted; this individual was considered dominant over their partner. Once all dyad combinations had been tested, the results were used to form a dominance hierarchy of all subjects.
As members of the first two groups of six individuals/sextets were non-overlapping, we ran the dyad combinations through the cooperation and dominance pre-tests and continued with the first two rounds of the test phase before individuals underwent the remainder of the dyad trials in the combinations that would appear in the remaining four sextets (and four rounds) of the test.
(iii). Test phase
To equalize subject experience as much as possible, triads were formed such that each individual appeared in two of the four triads within each group, and particular dyads appeared in no more than one triad within each group. Each individual appeared in three of the six groups. The majority of dyad combinations appeared in two groups (the rest appeared only in one).
Triad condition. Groups of three started in the room where peanuts could potentially be released. In addition, the room adjacent to the peanut room was open to reduce the potential for fighting over the reward. Trials started when the door to the overhead raceway was opened, allowing free access to the rope room. Trials lasted a maximum of 2 min.
Sextet condition. Same as for triad condition, except six individuals were tested together.
Testing order. Group size was counterbalanced such that three groups began with the triad condition, and three groups started with the sextet condition. Subjects experienced three sessions of three trials per condition. Trials were repeated up to three times if the rope was pulled out or on the few occasions an individual managed to pull both ends alone—in this case, the long rope used in the test was replaced on subsequent trials by a rope measuring 345 cm, the shortest length that still allowed for possible success. Each individual trial thus had three possible ‘takes’ (repeated trials). Original trial outcome could be success or 2 min—no repetitions needed and continue to trial 2, or rope out or pulled alone—up to a maximum of three further repetitions available. All six groups experienced two sessions of each condition before adding a third round of each condition.
(d). Coding and analysis
All trials were recorded by four cameras: one focused on one rope end and the funnel, one directed at the second rope end, one aimed at the feed dispenser and the area around it, and one capturing as much of the peanut room as possible. After each successful trial, the amount of peanuts released was measured. The variables coded were: (i) the result of the trial—success (when two individuals pulled the rope, releasing peanuts) versus no success (when 2 min elapsed without any action on the rope or the rope was pulled out); (ii) latency to pull (iii) who pulled; (iv) an approximation of how much each individual ate—could be none (score 0), scrounging from the floor (score 1) or sitting directly in front of the feed dispenser (score 2).
(i) Relationship between pulling and feeding. We analysed whether pulling the rope in a particular trial had an effect on the respective individuals' feeding success in that trial using a GLMM. We also used a GLMM to check whether cost (pulling the rope) and benefit (food eaten) might balance out over the sequence of trials within the same group (calculating the proportion of trials in which an individual pulled as well as the average food reward they got). For each analysis, we fitted a model into which we included whether or not the individual pulled in a given trial (or proportion pulling over the course of trials), the individual's rank, group size, the session and order of conditions as fixed effects and the ID of the group and the subject as random effects (see the electronic supplementary material, S12.1). Note that for the main analysis (all GLMMs), we counted any instance of success whether it occurred on an original trial or a repeated trial.
(ii) Factors influencing action (i.e. pulling) at an individual and dyadic level. To test what determined whether an individual pulled the rope (yes or no) in a successful trial, we used a GLMM. Into this, we included group size, session and order of conditions and all their interactions up to order three as fixed effects. To control for possible effects of rank, we also included this variable. The identity of the specific group, subject and individual trial were included as random effects. Another GLMM was used to test whether a given dyad pulled the rope in a successful trial. We included as fixed effects group size, the rank of the individual, the session and order of conditions. The identity of the specific group, of the dyad, of the lower- and higher-ranking individual and the individual trial were included as random effects (see the electronic supplementary material, S12.2).
(iii) Time to success. We analysed latencies (on a dyadic level) using a GLMM in which we included group size, the rank of the individual, the session and order of conditions as fixed effects, as well as group, dyad and the identities of the higher- and lower-ranked individual as random effects (see the electronic supplementary material, S12.3). Data: doi:10.5061/dryad.b5c1f.
(e). Results
(i). Dyad pre-test
In the dyad pre-test, 88 per cent of dyad combinations had some level of success (45% of pairs succeeded immediately on all four trials with no mistakes while the rest were successful on a repeated trial). In comparison, in the test phase, 41 per cent of dyad combinations pulled successfully (16% pulled in both the triad and sextet condition, 20% pulled only in the triad condition and 5% pulled only in the sextet condition). Of these successful dyads, 60 per cent had a perfect success record in the pre-test, 24 per cent succeeded on a repeated trial in the pre-test and 16 per cent had never succeeded in the pre-test.
(ii). Test phase
We found that subjects succeeded in 34.4 per cent of the triad and 42 per cent of the sextet initial trials. Subjects did not pull the rope during the entire 2 min trial duration in 28 per cent of the triad trials and 22 per cent of the sextet trials. The remaining trials (37.6% and 36% of the triad and sextet trials, respectively) were repeated owing to one individual pulling the rope out and/or succeeding alone. Of these repeated trials, 32 per cent in the triad condition and 56 per cent in the sextet condition resulted in success, and in 40 per cent of the triad trials and 22 per cent of the sextet trials, subjects let 2 min elapse without acting on the rope. In the remaining trials, subjects were unsuccessful owing to one individual pulling the rope out.
(i) Relationship between pulling and feeding. To investigate whether pulling resulted in a cost to the actor, we analysed whether pulling the rope in a particular trial influenced the respective individuals' feeding success. Pulling the rope clearly reduced the amount of food an individual received (GLMM, p < 0.0001). In addition, higher-ranking individuals received more food (p < 0.0001). The interaction between rank and pulling was not significant (p = 0.153; electronic supplementary material, S13.1). The second analysis looked at the costs (pulling the rope) and benefits (getting food) over the course of trials. There was a clearly significant interaction, over the course of trials, between pulling rate and rank (p = 0.005; electronic supplementary material, S13.2). While among individuals with low pulling rates, dominants ate more than subordinates, high- and low-ranking individuals who pulled the rope frequently had similar feeding success. Hence, high-ranking subjects suffered a more striking cost for pulling than lower-ranking individuals. There was a greater reduction in the amount eaten by a dominant puller versus a dominant free-rider than there was in the amount eaten by a subordinate puller versus a subordinate free-rider (figure 4).
Figure 4.
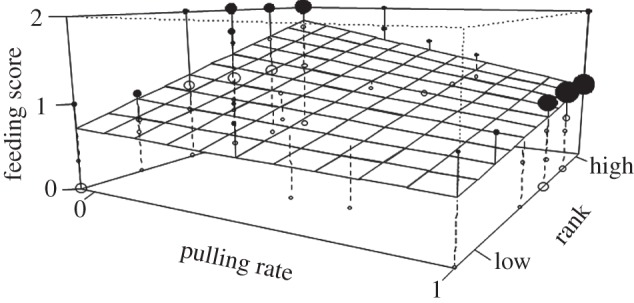
Influence of rank and pulling on feeding success. The surface represents results from the GLMM looking at pulling and feeding over the course of trials. The x-axis represents rank. Dots above the surface are in black, and dots below the surface are open. The area of the dots corresponds to the number of subjects in the respective combination of pulling rate and rank. Pulling rate is represented on the y-axis, with 0 indicating no pulling and 1 indicating pulling. Feeding score is represented on the z-axis: 2 = dispenser, 1 = ground, 0 = none (see methods for more details).
(ii) Factors influencing action. Whether an individual pulled or not was clearly affected by the factors investigated and the interactions between them. More specifically, the three-way interaction between group size, order and session was significant (p = 0.044; electronic supplementary material, S14), meaning that all three factors impacted the individuals' pulling probabilities. But note that rank did not have a significant impact, in contrast to experiment 1. When individuals received the triad condition first, the individual pulling rate started relatively high and decreased as the study progressed. When individuals received the sextet condition first, the individual pulling rate started low and increased in the triad condition (figure 5). Furthermore, pulling probabilities clearly differed between individuals (likelihood ratio test comparing full model with model not including the random effect of subject but everything else: χ2 = 88.43, d.f. = 1, p < 0.0001).
Figure 5.

Pulling probability in the triad and sextet conditions as a function of order (triad or sextet treatment first) and session. The GLMM revealed a three-way interaction between group size, order and session (see text). Rank from highest (1) to lowest (3).
We were also interested in investigating whether particular dyads were more prone to pulling. There was a clear effect of dyads (χ² = 30.55, d.f. = 1, p < 0.0001), such that certain combinations were more likely to pull than others (see the electronic supplementary material, S15 for non-significant results on the factors influencing dyadic pulling).
(iii). Time to success
Latencies were clearly influenced by the three factors investigated and their interactions. Specifically, the three-way interaction was significant (pMCMC = 0.0096, electronic supplementary material, S16). Regardless of order of conditions (triad or sextet first), latencies tended to increase in the first condition and decrease in the second condition. Generally, latencies were longer and changes were bigger when the sextet condition was received first.
(f). Discussion
The results demonstrate that the experimental design successfully engaged subjects in a Volunteer's Dilemma scenario, in which there was a distinct cost to acting. More dominant individuals were better able to feed than subordinate individuals on a given trial because if they did not pull, they could guard the position in front of the dispenser, and if they did pull, they were more likely than a subordinate to still successfully feed from the floor. Over the course of trials, dominants had to pay a higher cost for pulling than subordinates most likely because if they pulled they could not occupy the coveted dispenser position and could only feed from the floor.
Pulling rate was clearly affected by group size, order of conditions and session. It seems most likely that the subjects were not able to calculate the complexities of the dilemma offhand but adjusted their pulling rates as they experienced the dilemma reward structure over sessions. It is important to remember that while all group constellations were unique, individuals reappeared in more than one group and thus potentially already had experience in the two group sizes with other individuals.
One result of interest is that in the triad condition, pulling rates appear to be converging on some low but stable value in both the sextet first and triad first conditions. For those groups that experienced the sextet first, pulling rate increases across sessions, likely as a result of the greater potential for reward in the triad versus sextet condition. Conversely, in the groups that experience the triad first, pulling rate declines across triad sessions and remains low in sextet sessions. The first decline may be partly a function of their experiencing the reduced potential for reward relative to the dyad pre-test which came just before.
While it could be argued that pulling rates decrease simply as a result of a conditioning and extinction pattern prompted by receiving less reward, we believe this explanation is unlikely. Extinction from conditioning is typically very slow, and given the small number of trials and the incomplete cessation of reward, it is highly unlikely that extinction would occur in our experimental context. Furthermore, subjects showed flexibility in their decision-making, evident in their being motivated to pull when alone in the first pre-test, then acting or not acting in the dyad pre-test depending on their partner, and overall, displaying the tendency to pull less when in larger groups than in smaller ones regardless of order of condition. One may further note that some subjects who were unwilling to pull in the dyad pre-test did pull in the test phase when the amount of reward was reduced but group context had changed. Previous experiments have shown chimpanzees to be very sensitive to social intricacies in dyadic cooperative contexts [10–12], and it seems likely that their behaviour would be similarly influenced in our tests as well.
In the sextet condition, pulling rates either remain low (in the sextet first groups) or decline to low values (in triad first groups), which suggests resistance to solving the dilemma and potential for breakdown if further sessions were added. The highly significant variability in the pulling rates of different individuals suggests that certain subjects acted as ‘impact pullers,’ analogous to the impact hunters reported by Gilby and co-workers [6,9]. In the experiment, ‘impact pullers’ are highly motivated to pull the rope in any given trial, regardless of group size, order of condition or session number. Gilby & Connor [9] suggest that personality may be a significant factor determining motivation to initiate and persist in a hunt, and that those individuals most motivated to do so precipitate the collective hunt, most likely by reducing the cost of participation to subsequent hunters. It is possible that personality may account for some of the variance observed in pulling rates between individuals. However, it is still unclear whether these individuals influence success in the same way in an experimental setting as they do in the wild. While it has been suggested that an impact hunter reduces the costs for whoever follows by engendering chaos among the prey and increasing the likelihood that another hunter may succeed, it is as of yet unclear in exactly what ways an impact puller reduces the cost of pulling for another other than by assuring whoever follows that they have a willing partner and therefore, that the action would most likely be successful.
Furthermore, we detected a significant variation in pulling rates between dyads. This suggests that dyadic dynamics, perhaps revolving around tolerance between individuals, are also affecting variation in success rates.
4. General discussion
These experiments presented subjects with a Volunteer's Dilemma in which action was required to release a reward that could potentially be shared by all individuals present. As with any CAP, there were higher rewards associated with free-riding and disincentives to action. In experiment 1, the dilemma was overcome in all conditions, the typical pattern being that a particular higher-ranking individual would consistently choose to push in the majority of trials. The payoff structure appeared to be influenced by the dispersion of the reward, the strength of the hierarchy in each group as well as by the level of tolerance displayed between individuals. These results suggest that in this case, dominance was the primary force mediating the costs of action. This is an example of Nunn's [1] first class of influential factors: asymmetrical benefits and privileged groups. Because higher-ranked individuals have the security of knowing they will be rewarded for their effort, it is in their interest to produce the goods even when free-riding occurs, whereas subordinates typically pay a very high cost for pushing as they are subsequently unable to gain much access to the reward and thus they tend not to act. The only exception being when dispersion and rivalry are maximized, making it hard for dominants to take control of the reward and easier for subordinates to profit. Similar dominance effects in an experimental CAP have been observed in pigs [20].
In experiment 2, the dilemma was overcome in most triad trials but there appeared to be a decrease in action over sessions in the sextet condition. In our particular design, because the peanuts were sprayed into the room and thus highly dispersed, all individuals should have some motivation to act. The reward distribution was complicated by the fact that individuals learned to sit directly in front of the dispenser in order to catch the majority of peanuts. Dominants could use their rank to monopolize this favourable position if they remained in the peanut room and did not pull. In triad trials, lower-ranking individuals may be encouraged to pull because they would only have to compete with two other individuals; for the same reason, dominants may be more willing to sacrifice the dispenser strategy. In the sextet condition, the number of competitors also vying for a share of the peanuts more than doubles (going from two to five from the perspective of the individual actor), and thus subordinates have little incentive to pull while dominants would be reluctant to leave the coveted dispenser position. The cooperative aspect of the CAP in the second experiment also hindered success as not only one, but two individuals would have to decide the cost of action was worthwhile. Because of the changes in dispersion, group size and type of action (individual to cooperative), dominance (as an example of privileged groups) is unlikely to have been a strong mechanism leading to overcoming the CAP, particularly in the sextet condition.
Instead, in experiment 2, significant variation in individual motivation to act seemed to be the force propelling success. Impact pullers may have increased chances of peanuts being released in two ways. First, two impact pullers could have been tested in the same group and been willing to collaborate. This may explain in part the variation in pulling rates between dyads. Second, the movement of one impact puller towards a rope end may have acted as a catalyst for another individual deciding to act, in particular if the second individual could have access to the rope end under the raceway, which allowed the fastest return to the peanut room. At present, it is unclear what characteristics mark the personality of an impact puller.
The results of these experiments suggest that chimpanzees are able to overcome a Volunteer's Dilemma scenario, but that they may have a limited set of mechanisms available to them to do so. Asymmetrical benefits accruing from membership in a privileged group (higher-ranking individuals) may be one solution in simple small group scenarios. However, when dominants pay a higher cost for acting or lose opportunities to reap high rewards, this mechanism breaks down. Impact actors may play an important role in maintaining levels of success when general motivation levels are sufficiently high to encourage interest in collaboration, but in larger groups, when expected payoff drops, the influence of these impact individuals weakens. While a couple of subjects in experiment 2 began routinely claiming the dispenser position and proceeding to vocalize and bang on the bars and floor, and one individual was observed reaching towards another who was commonly an actor, these attention getters did not seem to have a direct effect on the success of the trial. No additional long-term mechanism for increasing the likelihood that someone volunteers to produce the shared good, such as increased coordination allowing for turn-taking, emerged. In this study, the principal cost of action was reducing the potential for reward. This is not exactly analogous to chimpanzee collective hunts where hunters increase rather than decrease their potential for possessing the reward or securing larger shares [7]. The costs of hunting are instead energetic effort and risk of injury. Therefore, in future, it would be interesting to examine CAP scenarios that reflect the payoff structure of hunting contexts more closely.
Acknowledgements
We are very grateful to the animal caretakers at the Wolfgang Köhler Research Center and to L. Ajarova, the trustees and all the staff of Ngamba Island Chimpanzee Sanctuary for their continuous help and support. Many special thanks to the animal caretakers of Ngamba Island. We also thank O. Boniface and R. Pieszek for building the apparatuses. We also appreciate permission from the Ugandan National Council for Science and Technology and the Uganda Wildlife Authority for allowing us to conduct our research in Uganda. We are extremely thankful to Roger Mundry for his invaluable help analysing the data.
References
- 1.Nunn C. L. 2000. Collective benefits, free-riders, and male extra-group conflict. In Primate males (ed. Kappeler P. M.), pp. 192–204 Cambridge, UK: Cambridge University Press [Google Scholar]
- 2.Heinsohn R., Packer C. 1995. Complex cooperative strategies in group territorial African lions. Science 269, 1260–1262 10.1126/science.7652573 (doi:10.1126/science.7652573) [DOI] [PubMed] [Google Scholar]
- 3.Nunn C. L., Deaner R. O. 2004. Patterns of participation and free riding in territorial conflicts among ringtailed lemurs (Lemur catta). Behav. Ecol. Sociobiol. 57, 50–61 10.1007/s00265-004-0830-5 (doi:10.1007/s00265-004-0830-5) [DOI] [Google Scholar]
- 4.Connor R. C. 2010. Cooperation beyond the dyad: on simple models and a complex society. Phil. Trans. R. Soc. B 365, 2687–2697 10.1098/rstb.2010.0150 (doi:10.1098/rstb.2010.0150) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Watts D. P., Mitani J. C. 2001. Boundary patrols and intergroup encounters in wild chimpanzees. Behaviour 138, 299–327 10.1163/15685390152032488 (doi:10.1163/15685390152032488) [DOI] [Google Scholar]
- 6.Gilby I. C., Eberly L. E., Wrangham R.W. 2008. Economic profitability of social predation among wild chimpanzees: individual variation promotes cooperation. Anim. Behav. 75, 351–360 10.1016/j.anbehav.2007.06.008 (doi:10.1016/j.anbehav.2007.06.008) [DOI] [Google Scholar]
- 7.Boesch C. 2002. Cooperative hunting roles among Taï chimpanzees. Hum. Nat. 13, 27–46 10.1007/s12110-002-1013-6 (doi:10.1007/s12110-002-1013-6) [DOI] [PubMed] [Google Scholar]
- 8.Watts D. P., Mitani J. C. 2002. Hunting behavior of chimpanzees at Ngogo, Kibale National Park, Uganda. Int. J. Primatol. 23, 1–28 10.1023/A:1013270606320 (doi:10.1023/A:1013270606320) [DOI] [Google Scholar]
- 9.Gilby I. C., Connor R. C. 2010. The role of intelligence in group hunting: are chimpanzees different than other social predators? In The mind of the chimpanzee: ecological and experimental perspectives (eds Lonsdorf E. V., Ross S. R., Matsuzawa T.), pp. 220–232 Chicago, IL: University of Chicago Press [Google Scholar]
- 10.Melis A. P., Hare B., Tomasello M. 2006. Engineering cooperation in chimpanzees: tolerance constraints on cooperation. Anim. Behav. 72, 275–286 10.1016/j.anbehav.2005.09.018 (doi:10.1016/j.anbehav.2005.09.018) [DOI] [Google Scholar]
- 11.Melis A. P., Hare B., Tomasello M. 2006. Chimpanzees recruit the best collaborators. Science 311, 1297–1300 10.1126/science.1123007 (doi:10.1126/science.1123007) [DOI] [PubMed] [Google Scholar]
- 12.Melis A. P., Hare B., Tomasello M. 2009. Chimpanzees coordinate in a negotiation game. Evol. Hum. Behav. 30, 381–392 10.1016/j.evolhumbehav.2009.05.003 (doi:10.1016/j.evolhumbehav.2009.05.003) [DOI] [Google Scholar]
- 13.Bullinger A. F., Wyman E., Melis A. P., Tomasello M. 2011. Coordination of chimpanzees (Pan troglodytes) in a stag hunt game. Int. J. Primatol. 32, 1296–1310 10.1007/s10764-011-9546-3 (doi:10.1007/s10764-011-9546-3) [DOI] [Google Scholar]
- 14.Diekmann A. 1985. Volunteer's Dilemma. J. Conflict Resolution 29, 605–610 10.1177/0022002785029004003 (doi:10.1177/0022002785029004003) [DOI] [Google Scholar]
- 15.Archetti M. 2009. The volunteer's dilemma and the optimal size of a social group. J. Theor. Biol. 261, 475–480 10.1016/j.jtbi.2009.08.018 (doi:10.1016/j.jtbi.2009.08.018) [DOI] [PubMed] [Google Scholar]
- 16.Archetti M. 2009. Cooperation as a volunteer's dilemma and the strategy of conflict in public goods games. J. Evol. Biol. 22, 2192–2200 10.1111/j.1420-9101.2009.01835.x (doi:10.1111/j.1420-9101.2009.01835.x) [DOI] [PubMed] [Google Scholar]
- 17.Baayen H. 2008. Analyzing linguistic data: a practical introduction to statistics using R. Cambridge, UK: Cambridge University Press [Google Scholar]
- 18.de Waal F. B. M., Davis J. M. 2003. Capuchin cognitive ecology: cooperation based on projected returns. Neuropsychologia 41, 221–228 10.1016/S0028-3932(02)00152-5 (doi:10.1016/S0028-3932(02)00152-5) [DOI] [PubMed] [Google Scholar]
- 19.Melis A. P., Hare B., Tomasello M. 2008. Do chimpanzees reciprocate received favours? Anim. Behav. 76, 951–962 10.1016/j.anbehav.2008.05.014 (doi:10.1016/j.anbehav.2008.05.014) [DOI] [Google Scholar]
- 20.Baldwin B. A., Meese G. B. 1979. Social behaviour in pigs studied by means of operant conditioning. Anim. Behav. 27, 947–957 10.1016/0003-3472(79)90033-2 (doi:10.1016/0003-3472(79)90033-2) [DOI] [Google Scholar]