

Abstract
The four major oligomeric reaction products from saponified modified hairy regions (MHR-S) from apple, produced by recombinant rhamnogalacturonan (RG) α-l-rhamnopyranosyl-(1,4)-α-d-galactopyranosyluronide lyase (rRG-lyase) from Aspergillus aculeatus, were isolated and characterized by 1H-nuclear magnetic resonance spectroscopy. They contain an alternating RG backbone with a degree of polymerization of 4, 6, 8, and 10 and with an α-Δ-(4,5)-unsaturated d-galactopyranosyluronic acid at the nonreducing end and an l-rhamnopyranose at the reducing end. l-Rhamnopyranose units are substituted at C-4 with β-galactose. The maximum reaction rate of rRG-lyase toward MHR-S at pH 6.0 and 31°C was 28 units mg−1. rRG-lyase and RG-hydrolase cleave the same alternating RG I subunit in MHR. Both of these enzymes fragment MHR by a multiple attack mechanism. The catalytic efficiency of rRG-lyase for MHR increases with decreasing degree of acetylation. Removal of arabinose side chains improves the action of rRG-lyase toward MHR-S. In contrast, removal of galactose side chains decreased the catalytic efficiency of rRG-lyase. Native RG-lyase was purified from A. aculeatus, characterized, and found to be similar to the rRG-lyase expressed in Aspergillus oryzae.
Highly branched RG structures are found associated with pectin in the cell walls of many different plants (O'Neill et al., 1990; Schols and Voragen, 1994; Yamada, 1994). Polygalacturonases, pectin lyases, and pectate lyases fragment the homogalacturonan regions of pectin but do not degrade RGs. However, these enzymes do release high-Mr, branched RGs from cell walls (De Vries et al., 1982; O'Neill et al., 1990; Schols et al., 1995b). The availability of RG-degrading enzymes is of great value to the structural elucidation of RG structures in the plant cell wall. Furthermore, the usefulness of these enzymes has been indicated in the processing of fruit, where it is important that the commercial pectolytic enzyme preparations solubilize and hydrolyze the branched RG structures, which otherwise remain as colloidally dissolved polymers in the juice and lead to problems during filtration and clarification (Voragen et al., 1992; Will and Dietrich, 1994).
RGase, the first enzyme with activity toward RG, was found by Schols et al. (1990a). Subsequently, an RG-acetylesterase (Searle-Van Leeuwen et al., 1992), an RG-rhamnohydrolase (Mutter et al., 1994), and an RG-galacturonohydrolase were identified (Mutter et al., 1996b). These enzymes were all purified from the commercial enzyme preparation, Pectinex Ultra-SP, produced by Aspergillus aculeatus. A second RGase (RGase B) was discovered in the authors' laboratory and later cloned and expressed in Aspergillus oryzae (Kofod et al., 1994). This recombinant RGase B is a specific RG-lyase (Azadi et al., 1995; Mutter et al., 1996a). In this report we describe the characterization of all four major oligomers formed by rRG-lyase from MHR (Schols et al., 1990b) by 1H-NMR spectroscopy. The recombinant enzyme is further characterized with respect to the influence of various MHR substituents on kinetic parameters. Native RG-lyase from the commercial A. aculeatus preparation was purified and compared with the recombinant enzyme.
MATERIALS AND METHODS
Substrates
MHR were isolated from apple liquefaction juice produced using Rapidase C600 (Gist-brocades, Delft, The Netherlands), according to the work described by Schols et al. (1990b). MHR were then saponified to yield MHR-S (Schols et al., 1990b), since RG-lyase and RG-hydrolase were hindered by acetyl groups. The sugar composition of MHR-S was reported previously (Mutter et al., 1994).
Isolation and characterization of RGmed and RGpoly degradation products of MHR-S produced by RG-hydrolase (previously RGase or RGase A) were described by Mutter et al. (1994).
Substrates used to determine side activities were polygalacturonic acid (Fluka), highly methoxylated pectin (DM92.3) prepared in our laboratory according to the procedure of Van Deventer-Schriemer and Pilnik (1976), larchwood arabino-β-(1,3)/(1,6)-galactan (“stractan”, Meyhall Chemical AG, Kreuzlingen, Switzerland), xylan from oat spelts (Koch and Light Ltd., Haverhill, England), carboxymethylcellulose (Akucell AF-0305, Akzo, Arnhem, The Netherlands), soluble starch (Merck AG, Darmstadt, Germany), a linear arabinan kindly provided by British Sugar (Peterborough, UK), potato arabino-β-(1,4)-galactan (isolated from potato fiber according to the method of Labavitch et al. [1976]), and a hexamer of galacturonic acid as described previously by Voragen (1972).
The following pnp-glycosides used for screening of glycosidase activities were obtained from Koch and Light Ltd. and from Sigma: pnp-α-l-Araf, pnp-α-l-Arap, pnp-α-l-Galp, pnp-β-d-Galp, pnp-α-d-Xylp, pnp-β-d-Xylp, and pnp-α-l-Rhap.
Isolation of rRG-Lyase Oligomers from MHR-S
MHR-S were degraded (2.5 g, 4.8% [w/v], 16.5 h, 40°C, in 5 mm NaOAc buffer, pH 6.0) by rRG-lyase (0.23 mg). The degradation products were separated on a column of Sephadex G-50, as described by the method of Schols et al. (1990b), using demineralized water for elution. Fractions were assayed by automated colorimetric methods for uronic acids and total neutral sugars as described by Mutter et al. (1994). Fractions were analyzed by HPAEC and those containing the rRG-lyase MHR oligomers (fractionated as 5, 6, and 7, see Fig. 1a) were further purified by preparative HPAEC, essentially as described by Schols et al. (1994), using a Dionex (Sunnyvale, CA) PA-100 (22- × 250-mm) column at 25 mL min−1 with the following gradient of NaOAc in 100 mm NaOH: 0 to 50 min, 200 to 300 mm; 50 to 55 min, 300 to 1000 mm; 55 to 70 min, 200 mm. Fractions were neutralized using acetic acid, pooled, dialyzed, and lyophilized.
Figure 1.

a, Size-exclusion chromatography of the rRG-lyase digest of MHR-S on a Sephadex G-50 column using demineralized water for elution. Fractions were pooled as indicated (1–7; ▴, neutral sugars; ▪, uronic acid); b, HPAEC elution patterns of fractions 5 to 7. PAD, Pulsed amperometric detection.
The neutral sugar composition of the Sephadex G-50 fractions was determined by hydrolyzing lyophilized material in 1 m sulfuric acid (3 h at 100°C) and subsequently by converting the released neutral sugars to their alditol acetates (Englyst and Cummings, 1984). These were analyzed on a 15-m × 0.53-mm i.d. wide-bore capillary DB 225 column in a Carlo Erba 4200 (Milan, Italy) gas chromatograph. The temperature program was set at 180°C for 1 min, from 180 to 220 at 2.5°C/min and 220°C for 3 min. The system was equipped with a flame ionization detector set at 275°C. Inositol was used as the internal standard. Uronic acid was determined using an automated colorimetric method based on the method described by Ahmed and Labavitch (1977).
Preparative Deacetylation of MHR
Eight batches of MHR (approximately 60 mg each) were treated for 1 h at 40°C as 0.27% (w/v) solutions in 50 mm NaOAc buffer (pH 5.0) with different amounts (between 1 and 40 μg, and one batch with an excess amount of 4 mg) of an rRG-acetylesterase from Aspergillus aculeatus (Kauppinen et al., 1995). After inactivation for 20 min at 100°C, the batch treated with 4 mg of enzyme was centrifuged to remove denaturated protein, and all batches were dialyzed (molecular mass cut-off, approximately 20 kD) one night against running tap water and 7 d against distilled water (at 4°C) and lyophilized. The DA and DM were determined by HPLC according to the method of Voragen et al. (1986).
Preparative Removal of Ara and Gal from MHR-S by Enzymes
MHR-S (in batches of approximately 50 mg) were treated with various enzymes and enzyme combinations in excess amounts (2–5 mg). Recombinant α-arabinofuranosidase, recombinant arabinanase, and recombinant β-(1,4)-galactanase from A. aculeatus (Christgau et al., 1995) were experimental batches kindly provided by Novo Nordisk A/S (Copenhagen, Denmark). A β-galactosidase from Aspergillus niger was purified following the procedure of Van de Vis (1994). To remove Ara, MHR-S were treated with the arabinofuranosidase (MHR-S deAra-1), with the endo-arabinanase (MHR-S deAra-2), and with the combination of these two enzymes (MHR-S deAra-3). To remove Gal, MHR-S was treated with the β-galactosidase (MHR-S deGal-1), with the endo-β-(1,4)-galactanase (MHR-S deGal-2), and with the combination of these two enzymes (MHR-S deGal-3). The removal of all Ara- and Gal-containing side chains was attempted with a combination of all formerly mentioned enzymes (MHR-S deAra-deGal). Reaction mixtures were incubated at 40°C for 24 h in 0.18 to 0.23% (w/v) solutions in 50 mm NaOAc (pH 5.0). After inactivation for 20 min at 100°C, the incubation mixtures were centrifuged to remove precipitated material. After dialysis the samples were lyophilized. Sugar composition was determined after methanolysis and subsequent hydrolysis with trifluoroacetic acid as described by De Ruiter et al. (1992).
Enzyme Purification
Native RG-lyase was partially purified from the commercial preparation Pectinex Ultra SP-L produced by A. aculeatus, kindly provided by Novo Nordisk Ferment Ltd. (Dittingen, Switzerland). Purification steps involved Bio-Gel P10, DEAE Bio-Gel A (Bio-Rad), and Mono-S HR 5/5 (Pharmacia LKB), and were performed as described by Mutter et al. (1994). Concentration between DEAE and Mono-S chromatography steps was carried out by ultrafiltration using a YM 10K membrane from Amicon (Danvers, MA). Fractions were screened for RG-lyase activity by incubation with MHR-S, followed by HPSEC and HPAEC (gradient A, see “Analytical Methods”).
rRG-lyase from A. aculeatus was purified essentially as described by Kofod et al. (1994), starting from lyophilized crude culture supernatant of an A. oryzae transformant (A 1560) producing rRG-lyase, kindly provided by Novo Nordisk A/S. Native RG-lyase was characterized and found to be essentially the same enzyme as rRG-lyase. Since rRG-lyase was available in large amounts, it was used in all further experiments.
RG-hydrolase was purified using the method of Schols et al. (1990a).
Enzyme Incubations and Enzyme Assays
Determination of Side Activities of Native RG-lyase from A. aculeatus
Native RG-lyase (0.5 μg mg−1 substrate) was screened for contaminating glycanase activities by incubation for 24 h at 40°C with 0.24% (w/v) solutions of selected substrates in 50 mm NaOAc buffer (pH 5.0). The digests from the glycanase assays were analyzed by HPSEC and HPAEC (gradient A). For contaminating glycosidase activities, screening was performed by incubating RG-lyase (12 μg mg−1 substrate) for 1 h at 30°C with 0.02% (w/v) solutions of pnp-glycosides in 50 mm NaOAc buffer (pH 5.0). The release of pnp from pnp-glycosides was measured spectrophotometrically at 405 nm and activity was calculated using the molar extinction coefficient of 13,700 m−1 cm−1.
Influence of pH and Temperature on Native RG-lyase from A. aculeatus
The optimum pH for RG-lyase was determined by incubating RG-lyase (1.25 μg mg−1 substrate) for 3 h at 40°C in 0.24% (w/v) substrate solutions in McIlvaine buffers (mixtures of 0.1 m citric acid and 0.2 m sodium hydrogen phosphate) in the pH range 2.0 to 8.0. The same buffers were used for preincubation of RG-lyase (4.5 h at 40°C) to measure the stability of RG-lyase at different pH values. After preincubation, 1.5 m NaOAc buffer (pH 5.0) was added to adjust the pH, and substrate solution was added to start incubation for 3 h at 40°C. The optimum temperature was determined by incubating RG-lyase (1.25 μg mg−1 substrate) for 3 h in 0.24% (w/v) substrate solutions in 50 mm NaOAc buffer (pH 5.0) at different temperatures in the range 3 to 60°C. The stability of RG-lyase at different temperatures was measured by preincubation for 4.5 h in 50 mm NaOAc buffer (pH 5.0). After the temperature was adjusted, substrate solution was added to start incubation for 3 h at 40°C. Incubation mixtures were inactivated by heating for 10 min at 100°C. Temperature and pH optima and stability were determined from the total amount of oligomeric fragments in digests based on peak area (HPAEC, gradient A). The pH optimum was also determined from the increase in A235.
Determination of Kinetic Parameters of rRG-Lyase toward MHR-Derived Samples
General reaction conditions were 50 mm NaOAc buffer (pH 6.0), 31°C, 0.42 μg rRG-lyase mL−1. Substrate concentrations in MHR samples with different DAs (Fig. 3; Table III) ranged between 0.03 and 0.6% (w/v). Substrate concentrations in various enzyme-treated MHR-S samples (Table IV) ranged between 0.01 and 0.4% (w/v). RG-lyase activity was calculated from the increase in A235 as measured in duplicate every 30 or 60 s using a spectrophotometer (model DU-62, Beckman) equipped with a Soft-Pac kinetics module (Beckman). The number of linkages cleaved was expressed in activity units (1 unit of enzyme producing 1 μmol unsaturated products min−1) using a molar extinction coefficient of 4800 m−1 cm−1 (MacMillan et al., 1964). Data analysis for calculation of kinetic parameters, using nonlinear regression, was performed by the program Enzfitter (Biosoft, Cambridge, UK)
Figure 3.

Influence of the DA of MHR on the Vmax (•) and Km (▪) of rRG-lyase. U, Unit.
Table III.
Specificity constant of rRG-lyase toward MHR samples in various stages of deacetylation (Km and Vmax, see Fig. 3)
| Substrate | (Vmax/[E])/Km |
|---|---|
| units mL mg−2 | |
| MHR DA 57 | 3.7 |
| MHR DA 48 | 8.8 |
| MHR DA 43 | 12 |
| MHR DA 38 | 26 |
| MHR DA 33 | 36 |
| MHR DA 30 | 38 |
| MHR DA 27 | 46 |
| MHR DA 24 | 49 |
| MHR DA 16 | 70 |
| MHR-S DA 1a | 89 |
This is saponified MHR; therefore, both acetyl and methoxyl esters have been removed.
Table IV.
Sugar composition of the enzyme-treated MHR-S fractions
| Sugar | MHR-S | deAra-1 | deAra-2 | deAra-3 | deGal-1 | deGal-2 | deGal-3 | deAra-deGal |
|---|---|---|---|---|---|---|---|---|
| mol % | ||||||||
| Rha | 14 | 18 | 16 | 19 | 12 | 15 | 15 | 14 |
| Ara | 19 | 9 | 9 | 5 | 16 | 17 | 17 | 7 |
| Gal | 18 | 20 | 19 | 19 | 13 | 16 | 14 | 13 |
| Glc | 5 | 2 | 4 | 4 | 4 | 5 | 3 | 3 |
| Xyl | 14 | 19 | 17 | 21 | 13 | 18 | 19 | 27 |
| GalA | 30 | 32 | 35 | 32 | 42 | 29 | 32 | 36 |
Activity of rRG-Lyase and RG-Hydrolase toward Other MHR Subunits
rRG-lyase (0.20 μg mL−1 reaction mixture) and RG-hydrolase (0.086 μg mL−1 reaction mixture) were both incubated with the following substrates: combined Sephadex G-50 fractions 1 and 2 (see Table I), combined Sephadex G-50 fractions 3 and 4, RGpoly, and RGmed. The latter two substrates were obtained from earlier work (Mutter et al., 1994). The enzymes were incubated with 0.1% (w/v) substrate solutions in 50 mm NaOAc (pH 5.0) for 20 h at 40°C. Mixtures were inactivated by heating for 5 min at 100°C and analyzed using HPSEC and HPAEC (gradient B).
Table I.
Sugar composition of Sephadex G-50 fractions of the rRG-lyase degradation products from MHR-S
| Sugar | Fraction
|
MHR-S | ||||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | ||
| mol % | ||||||||
| Rha | 4 | 6 | 9 | 12 | 18 | 22 | 27 | 16 |
| Fuc | 1 | 4 | 1 | 0 | 1 | 0 | 1 | 0 |
| Ara | 18 | 26 | 43 | 33 | 18 | 11 | 4 | 20 |
| Xyl | 27 | 21 | 11 | 4 | 2 | 2 | 4 | 11 |
| Man | 2 | 1 | 0 | 1 | 1 | 1 | 1 | 0 |
| Gal | 14 | 13 | 14 | 21 | 30 | 33 | 35 | 18 |
| Glc | 2 | 2 | 1 | 0 | 0 | 0 | 0 | 2 |
| GalA | 32 | 27 | 21 | 29 | 30 | 31 | 28 | 33 |
Determination of the Degree of Multiple Attack of rRG-Lyase
MHR-S solutions of 0.23% (w/v) in 50 mm NaOAc buffer (pH 6.0) and 0.01% (w/v) NaN3 were incubated at 40°C for 1 h using varying amounts of rRG-lyase per milligram of MHR-S. After inactivation for 10 min at 100°C, samples were analyzed using HPSEC. For RG-hydrolase the experiment was carried out similarly, with the exception that the pH of the 50 mm NaOAc buffer was 4.0 instead of 6.0.
Gel Electrophoresis and Dot Blotting
SDS-PAGE was carried out as described by Mutter et al. (1994). The pIs were determined using zymography (M. Mutter, G. Beldman, V.L.C. Klostermann, Y. Schnell, K. Dörreich, H. Berends, H.A. Schols, and A.G.J. Voragen, unpublished results).
RG-hydrolase and native RG-lyase were applied onto nitrocellulose membranes (Bio-Rad), which were previously washed in TBS (20 mm Tris buffer, pH 7.5, containing 500 mm NaCl). Unreacted binding sites were blocked by incubation for 45 min at room temperature in TBS containing 3% (w/v) gelatin. For immunoblotting, the membrane was incubated for 1.5 h at room temperature with a polyclonal rabbit antiserum raised against purified RG-hydrolase as described by Harlow and Lane (1988), diluted 1:2500 in TBS containing 0.05% (w/v) Tween 20 and 1% (w/v) gelatin. Bound rabbit antibodies were visualized by incubation of 5-bromo-4-chloro-3-indolylphosphate and nitroblue tetrazolium with an alkaline phosphatase-labeled goat anti-rabbit antibody (Sigma).
Determination of Molecular Mass of Native RG-Lyase Using Size-Exclusion Chromatography
A Superose 12 HR 10/30 column (Pharmacia) was calibrated with endopolygalacturonase (43 kD), RG-rhamnohydrolase (84 kD), RG-hydrolase (53 kD), and several partially purified proteins with molecular masses of 78, 76, 52, 45, and 32 kD, as characterized by SDS-PAGE. A buffer of 150 mm NaOAc (pH 6.0) was used for elution. Retention of RG-hydrolase (17.1 μg) and native RG-lyase (2.4 μg) on this column was monitored by collecting fractions and determining their activity toward MHR-S (analysis by HPSEC and HPAEC [gradient A] as described below).
Analytical Methods
HPSEC
The molecular mass distribution of substrates before and after enzyme treatment was determined by HPSEC on three Bio-Gel TSK columns in series (40XL, 30XL, and 20XL) as described by Schols et al. (1990a). Pectin standards of 100, 82, 77.6, 63.9, 51.4, 42.9, 34.6, and 10 kD (Van Deventer-Schriemer and Pilnik, 1987) and GalA and diGalA were used for calibration of the system. GPC/PC software from Spectra Physics (San Jose, CA) was used for determination of the number-average molecular mass.
HPAEC
HPAEC was performed using a Dionex Bio-LC system equipped with a Dionex CarboPac PA-100 (4 × 250 mm) column and a Dionex PED detector in the pulsed amperometric detection mode. Gradients of NaOAc in 100 mm NaOH (1 mL min−1) were used as follows: gradient A, 0 to 5 min, 0 mm; 5 to 35 min, 0 to 430 mm; 35 to 40 min, 430 to 1000 mm; 40 to 45 min, 1000 mm; 45 to 60 min, 0 mm; gradient B, 0 to 45 min, 100 to 380 mm; 45 to 55 min, 380 to 500 mm; 55 to 60 min, 500 to 1000 mm; 60 to 80 min, 100 mm.
1H-NMR Spectroscopy
1H-NMR spectra of the oligosaccharides in deuterated water were obtained at 400 MHz using a Jeol GX400 spectrometer. The sample temperature was 27°C (or 50°C to reveal signals at δ 4.75) and chemical shifts were determined using acetone (δ 2.217 with respect to tetramethylsilane) as an internal reference. Phase-sensitive two-dimensional NMR experiments (correlation spectroscopy, ROESY, and HOHAHA) were carried out as described previously (Colquhoun et al., 1990). For the two-dimensional experiments the spectral width was 2500 Hz in both dimensions, matrix size 2048(t2) × 256(t1), spin-locking times were 200 ms (ROESY) and 110 ms (HOHAHA), and spin-lock power 6.25 kHz (both experiments). Data processing was carried out using FELIX software (Molecular Simulations, Burlington, MA). Precise values of integrals were determined using the curve-fitting option of FELIX.
RESULTS AND DISCUSSION
Isolation of rRG-Lyase Oligomers Produced from MHR-S
Structural characterization of the oligomers that rRG-lyase release from MHR-S will lead to a better understanding of the mode of action of the enzyme. Therefore, MHR-S was incubated on a preparative (gram) scale with rRG-lyase. The digest was separated into seven fractions using a Sephadex G-50 column (Fig. 1a). The sugar composition of the seven fractions is shown in Table I. Fractions 1 to 4 contained larger fragments, whereas fractions 5 to 7 contained oligomeric reaction products (I–IV in Fig. 1b) that could be detected using HPAEC, and contained Gal, GalA, and Rha as predominant sugars. Thus, the oligomers generated by rRG-lyase treatment of MHR-S and the oligomers produced by RG-hydrolase treatment of MHR-S are composed of the same sugar residues. Fractions 5 to 7 were further fractionated using preparative HPAEC (Fig. 1b). Seven HPAEC fractions (7.1–7.7) were obtained from fraction 7, eight HPAEC fractions were obtained from fraction 6 (6.1–6.8), and 18 HPAEC fractions (5.1–5.18) were obtained from fraction 5. Finally, fraction 7.2 (containing oligomer I), fraction 6.5 (containing oligomer II), and fraction 5.12 (containing the two largest oligomers that could not be separated by HPAEC, III and IV) were selected because sufficient amounts of material were present for 1H-NMR spectroscopic analysis.
The trend in sugar composition of the Sephadex G-50 fractions (Table I) over the elution profile of the rRG-lyase digest of MHR-S resembles that of the Sephacryl S-200 fractions of the RG-hydrolase digest of MHR-S population A (the highest molecular mass population; Schols et al., 1995a). From the work of these authors on the characterization of the degradation products of apple MHR population A, a detailed model for the chemical structure of this population emerged. Three subunits were distinguished: subunit I, xylogalacturonan type of polymers (recognizable in G-50 fractions 1 and 2); subunit II, RG backbone stubs with an Rha:GalA ratio of less than 1 and arabinan side chains (recognizable in G-50 fractions 3 and 4); and subunit III, strictly alternating RG fragments with Gal side chains (recognizable in G-50 fractions 5–7). Subunit III and RG I as described by Albersheim et al. (1996) are essentially the same type of polysaccharide. The comparable S-200 fractions described by Schols et al. (1995a) were richer in Ara, but their starting material already contained more Ara. The results suggest that rRG-lyase attacks the same type of subunit in MHR as does RG-hydrolase, namely, the strictly alternating RG I regions with single-unit Gal side chains attached to the C-4 of Rha. This was confirmed by the fact that rRG-lyase was not active toward the higher molecular mass MHR-S degradation products (RGpoly and RGmed) liberated by RG-hydrolase after producing RG oligomers (not shown). Similarly, RG-hydrolase was not active toward the higher molecular mass MHR-S degradation products (combined Sephadex G-50 fractions 1 and 2 and 3 and 4) liberated by rRG-lyase after producing oligomers (not shown).
Characterization of rRG-Lyase Oligomers from MHR-S Using NMR Spectroscopy
Fraction 7.2
Figure 2 shows the 1H-NMR spectra of fractions 7.2, 6.5, and 5.12. Figure 2a is the region to low field of the residual water signal and Figure 2b shows all the peaks to high field of this position, except for a group of doublets at approximately 1.3 μg g−1. The spectrum of fraction 7.2 was discussed previously (Mutter et al., 1996a). Assignments made with the help of two-dimensional NMR spectra allowed the primary structure of the major oligosaccharide in fraction 7.2 to be determined as oligomer I. The feature that most clearly distinguished the degradation products of rRG-lyase from those of RG-hydrolase was the doublet a (δ 5.81) in Figure 2a, which was assigned to H-4 of a α-Δ-(4,5)-us-GalA residue at the nonreducing terminus of the oligosaccharide. Other assignments of well-resolved signals are indicated in the figure captions. They established that a Rha residue (H-1 signals c and g) was at the reducing end while the relative intensities of the remaining anomeric signals (d, f for α-GalA; b for α-Rha) showed that in addition to the terminal residues the oligosaccharide in fraction 7.2 had one internal Rha-GalA disaccharide unit. The linkage positions were confirmed by a ROESY experiment. H-1 signals of β-Gal residues were also identified (doublets j at δ 4.63 in Fig. 2b). These residues were linked to C-4 of the Rha units, as found for the oligosaccharides resulting from RG-hydrolase treatment of MHR-S (Colquhoun et al., 1990). Fraction 7.2 consisted almost entirely of the hexasaccharide I. The integration of the β-Gal H-1 signal was difficult (because of proximity to the irradiated water signal and overlap with GalA H-5). The presence of unsubstituted Rha units would be indicated by Rha H-4 signals below a δ of approximately 3.5, or H-6 signals below a δ of approximately 1.28 (Colquhoun et al., 1990). Since neither of these signals was observed, we conclude that all the Rha residues are substituted at C-4. The two weak triplets at δ 3.3 to 3.4 have a different origin, as discussed for fraction 6.5.
Figure 2.

The 400-MHz 1H-NMR spectrum of fractions 5.12, 6.5, and 7.2 (27°C). RE, Reducing end; NR, nonreducing end. a, Low-field region. Peak a, H-4 us-α-GalA (NR); peak b, H-1 α-Rha (linked to NR); peak c, H-1 α-Rha (RE); peak d, H-1 α-GalA (linked to β-RE); peak e, H-1 us-α-GalA (NR); peak f, H-1 α-GalA (linked to α-RE); peak g, H-1 β-Rha (RE); peak h, H-1 α-Rha (unit C, Table II); and peak i, H-1 α-GalA (unit D, Table II). b, High-field region. Peak j, H-β-Gal and H-5 α-GalA; peak k, H-4 α-GalA; peak l, H-3 us-α-GalA (NR); and peak m, H-2 α-Rha (linked to NR). An impurity (lactate?) is present in all of the spectra with signals at δ of 4.11 (quartet) and 1.33 (doublet).
Fractions 5.12 and 6.5
The anomeric signals discussed above were also clearly identifiable in spectra of fractions 5.12 and 6.5. A detailed two-dimensional NMR study (correlation spectroscopy, ROESY, and HOHAHA) was made of fraction 6.5 to determine the 1H chemical shifts of the major oligosaccharide, and these are presented in Table II. The basic features were very similar to those reported for fraction 7.2 (see above) but with the addition of a further internal Rha-GalA unit. It is seen from Table II that the Rha H-1 and H-2 chemical shifts are sensitive to whether the neighboring residue is GalA or us-GalA. Otherwise, the chemical shifts of protons in comparable residues were nearly the same. In particular, the Rha H-4 chemical shifts (δ 3.6–3.7) showed again that all the Rha residues carried a β-Gal substituent. The two new H-1 resonances are labeled h (Rha) and i (GalA) for fraction 6.5 in Figure 2a.
Table II.
1H Chemical shifts of oligosaccharide II (fraction 6.5)
| Unit | Codea | Chemical Shift (δ)
|
|||||
|---|---|---|---|---|---|---|---|
| H-1 | H-2 | H-3 | H-4 | H-5 | H-6 | ||
| Rha | Aα | 5.22 | 3.97 | 4.09 | 3.71 | 3.95 | 1.34 |
| Aβ | 4.95 | 4.06 | 3.86b | 3.60b | 3.52b | 1.35b | |
| α-GalA | B | 5.08, 5.16c | 3.92, 3.97c | 4.13, 4.15c | 4.42 | 4.64 | —d |
| α-Rha | C | 5.27 | 4.13 | 4.09 | 3.67 | 3.85 | 1.30 |
| α-GalA | D | 5.04 | 3.93 | 4.12 | 4.42 | 4.64 | — |
| α-Rha | E | 5.32 | 4.31 | 4.09 | 3.62 | 3.85 | 1.29 |
| us-α-GalA | F | 5.14 | 3.80 | 4.34 | 5.81 | — | — |
| β-Gale | 4.63 | 3.51 | 3.66 | 3.90 | n.d.f | n.d. | |
| ?β-Xylg | 4.48 | 3.29 | 3.55 | 3.78 | 3.37, 4.10 | — | |
Backbone sugar units are coded A to F from reducing end to nonreducing end.
Tentative assignment (weak signals).
Two values are for unit B linked to Aα and Aβ, respectively.
−, Not present.
Three residues, linked to A, C, and E. Not distinguished, δ values differ by <0.01 ppm.
n.d., Not determined.
Detected in fraction 6 but not part of oligosaccharide II (see text).
Two signals appeared in the same positions, but with greater relative intensity, in the spectrum of fraction 5.12. Signal h had integrated intensity 1.1 (for fraction 6.5) and 2.5 (for fraction 5.12) with respect to signal b (known to represent a single proton). This suggests that fraction 6.5 contains a mixture of II (major) and III (minor) with fraction 5.12 containing a mixture of III and IV in roughly equal proportions. Thus, it appears that the four major peaks found in the HPAEC analysis (Fig. 1b) can be identified with the oligosaccharides I–IV:
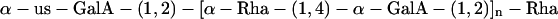 |
 |
 |
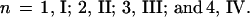 |
Four major oligomers are identified, in contrast with Kofod et al. (1994), who reported only two major oligosaccharides released by rRG-lyase from MHR-S. However, the MS data of Azadi et al. (1995) showed that rRG-lyase released partially galactosylated oligomers with an RG backbone of DP 4, 6, and 8 from sycamore RG I.
The overall sugar analysis (Table II) indicates some deviation from the expected 1:1:1 ratios for Rha:Gal:GalA, as well as the presence of additional sugars, especially Ara. The presence of a very large number of minor components can be seen from the HPAEC elution patterns (Fig. 1b) and is most marked for fraction 6 (between 2 and 20 min). Although the great majority of the NMR signals observed could be attributed to I to IV, there were additional signals in the spectrum of fraction 6.5 that could not be assigned to these RG oligosaccharides. None of these additional signals could definitely be assigned to Ara residues, but one complete network of signals was identified as originating from a β-Xyl residue (Table III). These signals were most prominent in fraction 6.5, weak in fraction 7.2, and absent from fraction 5.12. Well-resolved signals belonging to the network are the H-1 doublet at δ 4.48 and two triplets at δ 3.3 to 3.4 (Fig. 2b, fraction 6.5). The chemical shifts and the coupling pattern indicated that the signals might arise from a β-Xyl residue. A further unassigned signal at δ 4.75 (singlet or unresolved doublet) was present, again strong in fraction 6.5 and weak in the other two samples. Schols et al. (1995a) reported that this chemical shift arose from H-1 of GalA, branched at C-3 with β-Xyl, as in xylogalacturonan subunits of the pectic hairy regions. However, A.J. Mort (personal communication) found a signal at approximately 4.68 ppm, which is from H-4 of GalA with a Xyl linked to its 3-position in a xylogalacturonan oligomer. It suggests that some oligosaccharides of the xylogalacturonan type may be present in fraction 6.5, but there is no evidence that they are linked to the well-characterized RG oligosaccharides.
Influence of MHR Acetyl Groups on the Activity of rRG-Lyase
The effects of the acetyl substituents, reported to occur on the RG part of pectin (Komalavilas and Mort, 1989; Schols et al., 1990b; Ishii, 1995), on the Vmax and the Km of rRG-lyase for MHR degradation was investigated. The DA of MHR is 57, whereas treatment of MHR with NaOH resulted in an almost complete removal of acetyl groups (DA of MHR-S is 1). The RG-acetylesterase was able to remove 72% of the acetyl groups originally present in MHR. This is in agreement with the findings of Searle-Van Leeuwen et al. (1992). According to the work of Kauppinen et al. (1995), only 60% of the available acetyl groups could be removed, which they attributed to the use of a different MHR batch. The DM of MHR treated with excess RG-acetylesterase and of parental MHR were similar (DM 23), confirming that no methoxyl groups were removed by the RG-acetylesterase. Km and Vmax values of rRG-lyase were determined using MHR, MHR-S, and MHR batches with different DA values. The results are shown in Figure 3.
It is clear that the affinity of rRG-lyase decreases drastically when the DA increases: the Km is more than 10 times larger for MHR than for MHR-S. It must be noted that MHR-S (data point at DA 1 in Fig. 3) is the only sample that has all methoxyl groups removed as a result of the saponification. However, the minor difference in Vmax of rRG-lyase toward MHR-S and toward MHR DA 16 suggests that the methoxyl groups do not have a large influence on rRG-lyase action, similar to RG-hydrolase (Schols et al., 1990a). It can be concluded from Figure 3 that the affinity for highly acetylated MHR will be extremely low. The effect of the DA on the Vmax of rRG-lyase is only moderate, but the 40% lower Vmax of rRG-lyase toward MHR compared with MHR DA 16 shows that it becomes catalytically more difficult to cleave the RG backbone when more acetyl groups are present. The overall effect of the kinetic parameters can be expressed as the “specificity constant”: kcat/Km, where kcat is the catalytic constant, which equals (Vmax/[E])/Km (Table III; Fersht, 1985). The calculated constants indicate that rRG-lyase is 25 times more specific for MHR-S than for MHR and that rRG-lyase is 20 times more specific for MHR DA 16 than for MHR DA 57. For comparison, the majority of alginate lyases are also known to be hindered by the presence of O-acetyl groups on the C-2 and C-3 positions of d-mannuronosyl residues (Sutherland, 1995).
Influence of MHR Side Chains on the Activity of rRG-Lyase
The influence of the Ara and Gal containing side chains (together making up 38 mol %) of MHR-S on rRG-lyase activity was investigated. MHR-S were treated with different glycosylhydrolases and combinations. No enzymes available were able to remove the β-(1,3)-linked Xyl from the xylogalacturonan part of MHR (Schols et al., 1995a). However, from the data collected so far, rRG-lyase did not act toward the xylogalacturonan part of the hairy regions. After the enzyme treatment, the molecular mass distribution of the MHR-S samples (as determined using HPSEC, not shown) had not changed, which indicated that the backbone of the samples was not degraded.
In Table IV the sugar composition of the enzyme-treated substrates is presented. Calculations were performed to check whether sugars other than Ara (e.g. in deAra-1) were removed. Therefore, for each batch the increase in mole percent of other sugars was calculated assuming that only the Ara, or Gal, or Ara plus Gal (in deAra-deGal) content decreased. From the difference between these theoretical data and the sugar composition data actually found, it could be concluded that from most enzyme-treated MHR-S batches some Glc was also liberated, in the deGal samples also some Ara, and some Rha when the β-galactosidase was used (results not shown), presumably by contaminating enzyme activities. It can be seen in Table IV that not all Ara could be removed, not even by the combination of the arabinofuranosidase and the arabinanase (maximum 74% of Ara). Attempts to remove Gal were even less successful (maximum 28% of Gal). Apparently, other types of these enzymes with other specificities are required. The fact that MHR-S can be completely degraded by the A. aculeatus preparation Pectinex Ultra SP after prolonged incubation suggests that all required enzyme activities for side chain and backbone degradation are present in the preparation, but probably as very minor constituents, like the RG-rhamnohydrolase (Mutter et al., 1994) or the rRG-lyase.
The Vmax and the Km of rRG-lyase toward these substrates were determined. Since the RG region of MHR is the true substrate for rRG-lyase, and because the enzyme treatments caused the various MHR-S batches to have varying amounts of RG, the Km values initially obtained were adjusted to represent the same RG content (based on the Rha content, Table IV) as MHR-S. These corrected values are presented in Table V. The Km values for the substrates in which treatment involved endo-arabinanase (MHR-S deAra-2 and -3) or endo-β-(1,4)-galactanase (MHR-S deGal-2) were lower than the value for MHR-S. Treatment with arabinofuranosidase only (MHR-S deAra-1) with the β-galactosidase only (MHR-S deGal-1), with the combination of β-galactosidase and β-(1,4)-galactanase (MHR-S deGal-2), and with the combination of all four enzymes (MHR-S deAra-deGal) did not significantly influence the Km. It is inconsistent that the Km for the latter two samples had not decreased, since for the modification of these samples the same enzymes were used as for MHR-S deAra2, deAra3, and deGal2.
Table V.
Vmax and Km of rRG-lyase toward enzyme-treated MHR-S samples
| Substrate | Enzymes Used in Treatment | Vmax/[E] | Km | (Vmax/[E])/Km |
|---|---|---|---|---|
| units mg−1 | mg mL−1 | units mL mg−2 | ||
| MHR-S | None | 28 (±1.3) | 0.33 (±0.09) | 85 |
| MHR-S deAra-1 | Arafur.a | 26 (±0.8) | 0.32 (±0.03) | 81 |
| MHR-S deAra-2 | Endo-ara.b | 25 (±0.8) | 0.20 (±0.03) | 125 |
| MHR-S deAra-3 | Arafur. + endo-ara. | 24 (±0.1) | 0.20 (±0.004) | 120 |
| MHR-S deGal-1 | β-Gal.c | 20 (±0.5) | 0.32 (±0.02) | 63 |
| MHR-S deGal-2 | Endo-gal.d | 16 (±0.4) | 0.25 (±0.02) | 64 |
| MHR-S deGal-3 | β-Gal. + endo-gal. | 18 (±0.9) | 0.35 (±0.05) | 51 |
| MHR-S deAra-deGal | Arafur. + endo-ara. + β-gal. + endo-gal | 17 (±1.6) | 0.39 (±0.11) | 44 |
The initially obtained Km values for the enzyme-treated MHR-S batches were adjusted to represent the same RG content (based on the Rha content) as the original MHR-S.
Arafur., Arabinofuranosidase.
Endo-ara., Endo-arabinanase.
β-Gal., β-Galactosidase.
Endo-gal., Endo-galactonase.
The improved action of rRG-lyase after removal of Ara side chains from MHR-S by endo-arabinanase suggests that the Ara side chains sterically hinder the rRG-lyase. Although the arabinofuranosidase is able to remove the same amount of Ara from MHR-S, the Km is not affected. This could be explained by the fact that the arabinofuranosidase, able to cleave off terminal α-(1,2)-, α-(1,3)-, and α-(1,5)-linked Ara units (Beldman et al., 1993), cannot pass a Gal unit if present, e.g. in the subbranches of the longer Ara side chains, thus leaving large stretches of these longer side chains intact that might sterically hinder the rRG-lyase. The endo-arabinanase, on the other hand, is able to attack the longer side chains at locations closer to the RG backbone; therefore, only small or no side chains will be left. Improved action of rRG-lyase toward RG I regions when Ara side chains were removed was also reported by Azadi et al. (1995). They found that neither rRG-hydrolase or rRG-lyase was able to fragment sycamore RG I unless most Ara units had been removed by trifluoroacetic acid hydrolysis.
The Vmax of rRG-lyase toward the treated MHR-S substrates was slightly lower when Ara was removed, but the Gal removal, although marginal, decreased the Vmax markedly. Apparently, Gal had been removed near regions where cleavage could occur. The results suggest that the single-unit Gal side chains attached to C-4 of Rha might play an important role in the cleavability of the Rha-GalA linkage by rRG-lyase. The effectivity of cleavage by rRG-lyase of the various enzyme-treated MHR-S batches was expressed in the specificity constant (Vmax/[E])/Km (Fersht, 1985). Clearly, the substrates treated with endo-arabinanase were cleaved with the highest catalytic efficiency.
Degree of Multiple Attack of rRG-Lyase
When a time-course experiment of degradation of MHR-S by rRG-lyase was done, the rapid shift in molecular mass of MHR-S upon HPSEC was accompanied by a rapid increase in oligomers formed as detected using HPAEC (not shown). The same set of four oligomers was formed from the start, in a ratio that did not significantly change during the progress of the degradation. Therefore, it was hypothesized that rRG-lyase fragmented MHR-S with a certain degree of multiple attack. In a multiple attack mechanism, once the enzyme forms an enzyme-polymer complex, the enzyme may catalyze the hydrolysis of several bonds before it dissociates and forms a new active complex with another polymer chain. The multiple attack mechanism can be seen as a general concept including single chain (“zipper” fashion) and multichain (random attack) mechanisms as extreme special cases (Robyt and French, 1967).
The degree of multiple attack may be defined as the average number of catalytic events, following the first, during the lifetime of an individual enzyme-substrate complex (Robyt and French, 1967). For calculation of the degree of multiple attack the ratio (r) has to be calculated. This is the ratio between the total amount of linkages that are split, i.e. the sum of both newly produced polymer and oligomer fragments, and the number of effective encounters, i.e. newly produced polymer fragments. This ratio gives the number of bonds that is broken per effective encounter. Since the first bond broken releases a polymer fragment, the average number of subsequent broken bonds (r − 1) is numerically equal to the degree of multiple attack (Robyt and French, 1967).
As a measure for the total amount of linkages split by α-amylase acting toward amylose, Robyt and French (1967) determined the increase in reducing value of the total amylose digest during degradation. To obtain the number of effective encounters, they determined the increase in reducing value of the 67% ethanol precipitate of the digest, which contains polymers. In this study, instead of ethanol precipitation to separate polymers from oligomers, HPSEC was performed on MHR-S digests, and a separation between oligomers and polymers was made at the retention time of 29 min, corresponding to a molecular mass of approximately 3000 D (approximately DP 20). Under the conditions used by Robyt and French (1967), the smallest polymer that could be precipitated by 67% ethanol had an average DP of 20 as well. Subsequently, the number-average molecular mass was calculated, using GPC/PC software, for the total digest and the thus-defined polymer fraction. From the number-average molecular mass and the carbohydrate content of the total digest, the total number of molecules present in a sample was calculated. The area percentage from the total HPSEC chromatogram made up by the polymers was used to determine the carbohydrate content of polymers in a sample. From this carbohydrate content and the number-average molecular mass, the number of polymers in a sample was calculated. Finally, the parameter r was calculated from the increase in the total number of molecules in the digest, divided by the increase in the number of polymers in the samples during degradation.
While Robyt and French (1967) used a fixed concentration of enzyme per milligram of substrate and took samples at increasing incubation times, in this study samples of progressing degradation were obtained by using increasing amounts of enzyme per milligram of substrate with a fixed incubation time. In Figure 4a the number of total molecules and the number of polymers present in the samples are shown. Up to a concentration of 60 ng rRG-lyase mg−1 substrate, the number of polymers still increased. Above this concentration, the number of polymers decreased, which means that degradation has advanced so far that all newly formed fragments fall into the category of oligomers. Obviously, this results in a rapidly increasing r − 1 value, as shown in Figure 4b. Therefore, true r − 1 values can be obtained only from samples in which the number of polymers is still increasing. The average degree of multiple attack, r − 1, calculated from the data points up to 60 ng rRG-lyase mg−1 substrate was 2.5. For RG-hydrolase a similar experiment was carried out and the degree of multiple attack was found to be almost twice as high: 4.0 (not shown). For different α-amylases values of multiple attack between 1.9 and 6.0 were measured (Robyt, 1984). Since the method of Robyt and French has not been used for other polymers, no comparison other than with starch-degrading enzymes could be made.
Figure 4.

Various parameters for determination of the degree of multiple attack (r − 1) of rRG-lyase toward MHR-S. a, The number of total molecules (○) and polymers (□) present; b, the (r − 1) values calculated from r; see text for explanation.
A major difference with the experiments of Robyt and French is that, instead of long starch homopolymers (average DP of 1,000), the MHR as used here is a heterogeneous substrate, with a very broad distribution in molecular mass (major populations ranging between 80,000 and 7,000 D, i.e. approximately between DP 500 and 40). The strictly alternating RG sequences with single-unit Gal substituents in MHR might not be very long, in which case the degree of multiple attack as determined is underestimated. However, the values obtained for rRG-lyase and RG-hydrolase do give us valuable information about the length of these RG sequences in MHR. They show that the average length of alternating RG sequences in MHR would have to allow for the release of 4 (RG-hydrolase) and 2.5 (rRG-lyase) consecutive oligomers, respectively. It is known which oligomers are released from MHR-S by RG-hydrolase (major products with RG backbones of DP 4 and 6, Mutter et al., 1994), and therefore the RG sequences in MHR would have to be on average 20 sugar residues long to release two oligomers of DP 4 and two of DP 6. For rRG-lyase, the average sequence length would have to be 20 sugar residues to release three oligomers of DP 4, 6, and 8. The length of the RG I regions in MHR must therefore be rather longer than the average of 13 units, as suggested by Schols and Voragen (1996) in their model.
Comparison of Native and rRG-Lyase from A. aculeatus
The crude enzyme mixture Pectinex Ultra SP, produced by A. aculeatus, was first desalted on a Bio-Gel P10 column. Desalted protein was applied to a DEAE Bio-Gel A anion-exchanger at pH 5.0. RG-lyase was present in the unbound protein. On subsequent chromatography of the unbound protein using a Mono-S HR 5/5 cation-exchanger at pH 4.25, RG-lyase eluted in a distinct peak at 40 mm NaCl.
The characteristics of native RG-lyase are summarized in Table VI. SDS-PAGE of RG-lyase revealed a major protein band at 76 kD and a minor one at 57 kD. Upon chromatography of RG-lyase using a calibrated Superose 12 column, RG-lyase eluted in the tail (57 kD) of the major protein peak. The nature of the major protein is as yet unknown, since no activity toward various glycans and pnp-glycosides could be detected. The pI of RG-lyase, determined using zymography (M. Mutter, G. Beldman, V.L.C. Klostermann, Y. Schnell, K. Dörreich, H. Berends, H.A. Schols, and A.G.J. Voragen, unpublished results), was 5.1 to 5.3. RG-lyase was found to be most active between 50 and 60°C and was stable for 4.5 h up to 40°C. A pH optimum of 6.0 was found, and the enzyme was stable at pH 6.0 and higher (measured up to pH 8.0). The pH optimum determined from the increase in A235 was the same as that derived from the increase of HPAEC peak area. Dot blotting was performed to investigate whether native RG-lyase showed immunological cross-reactivity to a polyclonal rabbit antiserum raised against purified native RG-hydrolase. The antiserum reacted only in negligible amounts with RG-lyase but strongly with RG-hydrolase, suggesting that the two polypeptides are structurally different, representing two different enzymes.
Table VI.
Characteristics of native RG-lyase from A. aculeatus, compared with rRG-lyase from A. aculeatus
| Characteristic | Native RG-Lyase | rRG-Lyasea |
|---|---|---|
| Molecular mass (kD) | 57 | 55 |
| pI | 5.1–5.3 | 5.2 |
| pH optimum | 6 | 6 |
| pH at which stable for 4.5 h | >6 | >6 |
| Temperature optimum | 50–60°C | 50°C |
| Temperature stability | Stable 4.5 h up to 40°C (100% activity) | Activity after 2 h at 60°C is 0% |
| Recognition by antiserum raised against RG-hydrolase | Negligible (dot blot) | No (western blot) |
Data from Kofod et al. (1994).
rRG-lyase from A. aculeatus was purified from the culture supernatant of an A. oryzae transformant. The reaction products that are formed from MHR-S upon incubation with native RG-lyase from A. aculeatus or rRG-lyase showed similar HPSEC and HPAEC elution patterns (not shown). The data on temperature and pH optimum and stability of rRG-lyase, as determined by Kofod et al. (1994), are shown in Table VI. Small differences in characteristics can be explained by interlaboratory assay variation, e.g. in the different assays and assay conditions used. It can be concluded that rRG-lyase and native RG-lyase from A. aculeatus are essentially the same enzyme.
The importance of RG-degrading enzymes such as RG-lyase will most certainly be established in the future, e.g. in the field of the oligosaccharins. RG I generated from cultured Acer spp. cell walls by pectinase digestion has been demonstrated to have wound-signal activity (Ryan et al., 1981). From mucilage of germinated cress seeds α-Δ-(4,5)-us-GalA-(1,2)-Rha disaccharides were isolated (as the sodium salt and named lepidimoide), which appeared to promote Amaranthus caudatus hypocotyl elongation (Hasegawa et al., 1992). Interestingly, in Pectinex Ultra SP, from which RG-lyase was purified, enzyme activities have been discovered by the authors that are able to degrade all four RG-lyase MHR-S oligomers completely into Gal and one unknown product, which, regarding the elution behavior is tentatively identified as the α-Δ-(4,5)-us-GalA-(1,2)-Rha dimer. This implies that a β-galactosidase and a new lyase have been active and that more new enzyme activities can still be purified from the A. aculeatus preparation. From these observations and the literature reports of this subject, it is anticipated that RG-lyase will be very useful in the investigation of the biological activity of Δ-(4,5)-unsaturated-RG oligosaccharides.
CONCLUSIONS
rRG-lyase releases α-Δ-(4,5)-unsaturated RG oligosaccharides I to IV from MHR-S, which are completely galactosylated and have backbone DPs of 4, 6, 8, and 10, confirming the results of Azadi et al. (1995), who found RG backbones of DP 4, 6, and 8. However, Azadi et al. (1995) found only partially galactosylated oligomers, which is probably due to the different source (sycamore RG I). The oligomers released by rRG-lyase are larger than the major products released from MHR-S by RG-hydrolase (backbone DPs of 4 and 6 [Colquhoun et al., 1990; Mutter et al., 1994]). Apparently rRG-lyase has more subsites for sugar binding in its active site than RG-hydrolase. This is confirmed by the observation that RG-hydrolase is able to cleave smaller linear RG oligomers than rRG-lyase (Mutter et al., 1996b). The degree of multiple attack toward MHR-S of RG-lyase (2.5) and RG-hydrolase (4) reveals that in MHR the alternating RG I sequences have to be at least 20 units long, in contrast to the model of Schols and Voragen (1996), in which the RG I regions are suggested to be, on average, 13 units long. The catalytic efficiency of rRG-lyase for MHR was shown to increase by removal of acetyl groups and Ara side chains. In contrast, when Gal side chains were degraded, the catalytic efficiency of rRG-lyase decreased. Native RG-lyase was purified from A. aculeatus, characterized, and found to be similar to the rRG-lyase expressed in A. oryzae (Kofod et al., 1994; Mutter et al., 1996a).
ACKNOWLEDGMENTS
Thanks are due to Jan van Iersel for his contribution to the isolation of RG-lyase MHR oligomers; to Marjo Searle-Van Leeuwen for purification of the β-galactosidase from A. niger; to Ben van den Broek for his work on the immunoblotting; to Marcel Mischler, Yvette Schnell, and Dr. Kurt Dörreich from Novo Nordisk Ferment Ltd (Dittingen, Switzerland) for purification of RG-hydrolase; and to Dr. Lene Venke Kofod and Dr. Hans-Peter Heldt-Hansen from Novo Nordisk (Denmark) for supplying crude rRG-lyase and the antiserum raised against RG-hydrolase.
Abbreviations:
- DA
degree of acetylation: no. of mol of acetyl groups per 100 mol of GalA residues
- DM
degree of methoxylation: no. of mol of methoxyl groups per 100 mol of GalA residues
- DP
degree of polymerization
- [E]
enzyme concentration
- GalA
d-galactopyranosyluronic acid
- HOHAHA
homonuclear Hartmann-Hahn spectroscopy
- HPAEC
high-performance anion-exchange chromatography
- HPSEC
high-performance size-exclusion chromatography
- MHR
modified hairy regions of pectin
- MHR-S
saponified MHR
- pnp
p-nitrophenyl
- RG
rhamnogalacturonan
- RGase
rhamnogalacturonase
- RG-hydrolase
RG α-d-galactopyranosyluronide-(1,2)-α-l-rhamnopyranosyl hydrolase
- RG-lyase
RG α-l-rhamnopyranosyl-(1,4)-α-d-galactopyranosyluronide lyase
- RGmed
intermediate-sized fragments produced from MHR-S by RGase
- RGpoly
high-molecular-mass fragments produced from MHR-S by RGase
- RG-rhamnohydrolase
RG α-l-rhamnopyranosylhydrolase
- Rha
l-rhamnopyranose
- ROESY
rotating frame Overhauser effect spectroscopy
- rRG-lyase
recombinant RG-lyase
- us-GalA
α-Δ-(4,5)-unsaturated GalA
- Vmax
maximum reaction rate
Footnotes
This work was supported by Novo Nordisk A/S (Copenhagen, Denmark).
LITERATURE CITED
- Ahmed A el R, Labavitch JM (1977) A simplified method for accurate determination of cell wall uronide content. J Food Biochem 1: 361–365
- Albersheim P, Darvill AG, O'Neill MA, Schols HA, Voragen AGJ. An hypothesis: the same six polysaccharides are components of the primary cell walls of all higher plants. In: Visser J, Voragen AGJ, editors. Progress in Biotechnology, Vol 14: Pectins and Pectinases. Amsterdam, The Netherlands: Elsevier; 1996. pp. 47–55. [Google Scholar]
- Azadi P, O'Neill MA, Bergmann C, Darvill G, Albersheim P. The backbone of the pectic polysaccharide rhamnogalacturonan I is cleaved by an endohydrolase and an endolyase. Glycobiology. 1995;5:783–789. doi: 10.1093/glycob/5.8.783. [DOI] [PubMed] [Google Scholar]
- Beldman G, Searle-Van Leeuwen MJF, De Ruiter GA, Siliha HA, Voragen AGJ. Degradation of arabinans by arabinanases from Aspergillus aculeatus and Aspergillus niger. Carbohydr Polym. 1993;20:159–168. [Google Scholar]
- Christgau S, Sandal T, Kofod LV, Dalbøge H. Expression cloning, purification and characterization of a β-1,4-galactanase from Aspergillus aculeatus. Curr Genet. 1995;27:135–141. doi: 10.1007/BF00313427. [DOI] [PubMed] [Google Scholar]
- Colquhoun IJ, De Ruiter GA, Schols HA, Voragen AGJ. Identification by N.M.R. spectroscopy of oligosaccharides obtained by treatment of the hairy regions of apple pectin with RGase. Carbohydr Res. 1990;206:131–144. doi: 10.1016/0008-6215(90)84012-j. [DOI] [PubMed] [Google Scholar]
- De Ruiter GA, Schols HA, Voragen AGJ, Rombouts FM. Carbohydrate analysis of water-soluble uronic acid containing polysaccharides with high performance anion-exchange chromatography using methanolysis combined with TFA hydrolysis is superior to four other methods. Anal Biochem. 1992;207:176–185. doi: 10.1016/0003-2697(92)90520-h. [DOI] [PubMed] [Google Scholar]
- De Vries JA, Rombouts FM, Voragen AGJ, Pilnik W. Enzymic degradation of apple pectins. Carbohydr Polym. 1982;2:25–33. [Google Scholar]
- Englyst HN, Cummings JH. Simplified method for the measurement of total non-starch polysaccharides by gas-liquid chromatography of constituent sugars as alditol acetates. Analyst. 1984;109:937–942. doi: 10.1039/an9820700307. [DOI] [PubMed] [Google Scholar]
- Fersht A (1985) Enzyme Structure and Mechanism. WH Freeman and Co., New York
- Harlow E, Lane D. Antibodies: A Laboratory Manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press; 1988. [Google Scholar]
- Hasegawa K, Mizutani J, Kosemura S, Yamamura S. Isolation and identification of lepidomoide, a new allelopathic substance from mucilage of germinated cress seeds. Plant Physiol. 1992;100:1059–1061. doi: 10.1104/pp.100.2.1059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ishii T. Isolation and characterization of acetylated rhamnogalacturonan oligomers liberated from bamboo shoot cell-walls by Driselase. Mokuzai Gakkaishi. 1995;41:561–572. [Google Scholar]
- Kauppinen S, Christgau S, Kofod LV, Halkier T, Dörreich K, Dalbøge H. Molecular cloning and characterization of a rhamnogalacturonan acetylesterase from Aspergillus aculeatus. J Biol Chem. 1995;270:27172–27178. doi: 10.1074/jbc.270.45.27172. [DOI] [PubMed] [Google Scholar]
- Kofod LV, Kauppinen S, Christgau S, Andersen LN, Heldt-Hansen HP, Dörreich K, Dalbρge H. Cloning and characterization of two structurally and functionally divergent rhamnogalacturonases from Aspergillus aculeatus. J Biol Chem. 1994;269:29182–29189. [PubMed] [Google Scholar]
- Komalavilas P, Mort AJ. The acetylation at O-3 of galacturonic acid in the rhamnose-rich portion of pectins. Carbohydr Res. 1989;189:261–272. [Google Scholar]
- Labavitch JM, Freeman LE, Albersheim P. Structure of plant cell walls. Purification and characterization of a β-(1,4)-galactanase which degrades a structural component of the primary cell wall of dicots. J Biol Chem. 1976;251:5904–5910. [PubMed] [Google Scholar]
- MacMillan JD, Phaff HJ, Vaughn RH. The pattern of action of an exopolygalacturonic acid-trans-eliminase from Clostridium multifermentans. Biochemistry. 1964;3:572–577. doi: 10.1021/bi00892a017. [DOI] [PubMed] [Google Scholar]
- Mutter M, Beldman G, Schols HA, Voragen AGJ. Rhamnogalacturonan α-l-rhamnopyranohydrolase. A novel enzyme specific for rhamnogalacturonan regions of pectin. Plant Physiol. 1994;106:241–250. doi: 10.1104/pp.106.1.241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mutter M, Colquhoun IJ, Schols HA, Beldman G, Voragen AGJ. Rhamnogalacturonase B from Aspergillus aculeatus is a rhamnogalacturonan α-l-rhamnopyranosyl-(1,4)-α-d-galactopyranosyluronide lyase. Plant Physiol. 1996a;110:73–77. doi: 10.1104/pp.110.1.73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mutter M, Renard CMGC, Beldman G, Schols HA, Voragen AGJ. Characterization of RG degradation products of new RGases using RG-rhamnohydrolase and RG-galacturonohydrolase. In: Visser J, Voragen AGJ, editors. Progress in Biotechnology, Vol 14: Pectins and Pectinases. Amsterdam, The Netherlands: Elsevier; 1996b. pp. 263–274. [Google Scholar]
- O'Neill M, Albersheim P, Darvill A. The pectic polysaccharides of primary cell walls. Methods Plant Biochem. 1990;2:415–441. [Google Scholar]
- Robyt JF. Enzymes in the hydrolysis and synthesis of starch. In: Whistler RL, Bemiller JN, Paschall EF, editors. Starch: Chemistry and Technology. London: Academic Press; 1984. pp. 87–123. [Google Scholar]
- Robyt JF, French D. Multiple attack hypothesis of α-amylase action: action of porcine pancreatic, human salivary, and Aspergillus oryzae α-amylases. Arch Biochem Biophys. 1967;122:8–16. doi: 10.1016/0003-9861(67)90118-x. [DOI] [PubMed] [Google Scholar]
- Ryan CA, Bishop P, Pearce G, Darvill AG, McNeil M, Albersheim P. A sycamore cell wall polysaccharide and a chemically related tomato leaf polysaccharide possess similar proteinase inhibitor-inducing activities. Plant Physiol. 1981;68:616–618. doi: 10.1104/pp.68.3.616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schols HA, Bakx EJ, Schipper D, Voragen AGJ. A xylogalacturonan subunit present in the modified hairy regions of apple pectin. Carbohydr Res. 1995a;279:265–279. [Google Scholar]
- Schols HA, Geraeds CCJM, Searle-Van Leeuwen MJF, Kormelink FJM, Voragen AGJ. Rhamnogalacturonase: a novel enzyme that degrades the hairy regions of pectins. Carbohydr Res. 1990a;206:105–115. [Google Scholar]
- Schols HA, Posthumus MA, Voragen AGJ. Structural features of hairy regions of pectins isolated from apple juice produced by the liquefaction process. Carbohydr Res. 1990b;206:117–129. [Google Scholar]
- Schols HA, Vierhuis E, Bakx EJ, Voragen AGJ. Different populations of pectic hairy regions occur in apple cell walls. Carbohydr Res. 1995b;275:343–360. doi: 10.1016/0008-6215(95)00155-m. [DOI] [PubMed] [Google Scholar]
- Schols HA, Voragen AGJ. Occurrence of pectic hairy regions in various plant cell wall materials and their degradability by RGase. Carbohydr Res. 1994;256:93–95. [Google Scholar]
- Schols HA, Voragen AGJ. Complex pectins: structure elucidation using enzymes. In: Visser J, Voragen AGJ, editors. Progress in Biotechnology, Vol 14: Pectins and Pectinases. Amsterdam, The Netherlands: Elsevier Science; 1996. pp. 3–19. [Google Scholar]
- Schols HA, Voragen AGJ, Colquhoun IJ. Isolation and characterization of rhamnogalacturonan-oligomers, liberated during degradation of pectic hairy regions by RGase. Carbohydr Res. 1994;256:97–111. doi: 10.1016/0008-6215(94)84230-2. [DOI] [PubMed] [Google Scholar]
- Searle-Van Leeuwen MJF, Van den Broek LAM, Schols HA, Beldman G, Voragen AGJ. Rhamnogalacturonan acetylesterase: a novel enzyme from Aspergillus aculeatus, specific for the deacetylation of hairy (ramified) regions of pectins. Appl Microbiol Biotechnol. 1992;38:347–349. [Google Scholar]
- Sutherland IW. Polysaccharide lyases. FEMS Microbiol Rev. 1995;16:323–347. doi: 10.1111/j.1574-6976.1995.tb00179.x. [DOI] [PubMed] [Google Scholar]
- Van Deventer-Schriemer WH, Pilnik W. fractionation of pectins in relation to their degree of esterification. Lebensm Wiss Technol. 1976;9:42–44. [Google Scholar]
- Van Deventer-Schriemer WH, Pilnik W. Studies on pectin degradation. Acta Alimentaria. 1987;16:143–153. [Google Scholar]
- Van de Vis JW (1994) Characterization and Mode of Action of Enzymes Degrading Galactan Structures of Arabinogalactans. Koninklijke Bibliotheek, The Hague, The Netherlands, pp 89–108
- Voragen AGJ. Characterization of pectin lyases on pectins and methyl oligogalacturonates. Agric Res Rep. 1972;780:1–121. [Google Scholar]
- Voragen AGJ, Schols HA, Beldman G. Tailor-made enzymes in fruit juice processing. Fruit Processing. 1992;2:98–102. [Google Scholar]
- Voragen AGJ, Schols HA, Pilnik W. Determination of the degree of methylation and acetylation of pectins by h.p.l.c. Food Hydrocolloids. 1986;1:65–70. [Google Scholar]
- Will F, Dietrich H. Characterization of residual pectins from raspberry juices. Carbohydr Polym. 1994;25:155–160. [Google Scholar]
- Yamada H. Pectic polysaccharides from Chinese herbs: structure and biological activity. Carbohydr Polym. 1994;25:269–275. [Google Scholar]